Back to Journals » International Journal of Nanomedicine » Volume 21
Biomimetic Nanoparticles for Bone Regeneration: Construction Strategies and Therapeutic Mechanisms
Authors Wang Z, Shen N, Yang T, Dong S, Xu K, Zhao K, Zhang Y, Jiang Y, Li T ![]()
Received 14 December 2025
Accepted for publication 19 March 2026
Published 26 March 2026 Volume 2026:21 589003
DOI https://doi.org/10.2147/IJN.S589003
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Professor Lijie Grace Zhang
Zehua Wang,1,2,* Ning Shen,3,* Tao Yang,2,* Shuhang Dong,1,2,* Kaige Xu,1 Kunyi Zhao,1 Yingze Zhang,1,4 Yaping Jiang,5 Tao Li1
1Department of Joint Surgery, The Affiliated Hospital of Qingdao University, Qingdao, 266003, People’s Republic of China; 2Department of Clinical Medicine, Qingdao Medical College of Qingdao University, Qingdao, 266003, People’s Republic of China; 3Institute of Cancer Stem Cell, Dalian Medical University, Dalian, 116044, People’s Republic of China; 4Department of Orthopedics, The Third Hospital of Hebei Medical University, Shijiazhuang, 050000, People’s Republic of China; 5Department of Oral Implantology, The Affiliated Hospital of Qingdao University, Qingdao, 266003, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yaping Jiang, Department of Oral Implantology, The Affiliated Hospital of Qingdao University, Qingdao, 266003, People’s Republic of China, Email [email protected] Tao Li, Department of Joint Surgery, The Affiliated Hospital of Qingdao University, No. 59, Haier Road, Qingdao, 266000, People’s Republic of China, Email [email protected]
Abstract: Bone tissue is the hardest and most dynamic connective tissue in the human body, and its integrity is essential for maintaining both mechanical support and physiological functions. However, with the aging population, the incidence of fractures, osteoporosis, and bone defects has risen significantly, severely impairing patients’ quality of life and creating a substantial social burden. Although autologous or allogeneic bone grafts and metallic or non-metallic implants can partially restore bone defects, their long-term efficacy is constrained by donor shortages, immune rejection, and limited regenerative capacity. Consequently, the development of efficient, precise, and biomimetic bone regeneration strategies is of great importance. In recent years, biomimetic nanoparticles have shown unique advantages in mimicking the bone microenvironment, delivering bioactive factors, and modulating cellular behavior due to their tunable structures and functions. This review summarizes the structural composition of bone tissue and its repair processes, highlights biomimetic nanoparticle construction strategies based on cell membranes, exosomes, proteins, and peptides, and discusses their roles in osteogenesis, mineralization, immune regulation, and neurovascular development. Finally, it explores their clinical translation prospects and associated challenges.
Keywords: biomimetic nanoparticles, bone regeneration, exosomes, targeting, tissue engineering
Introduction
Bone is one of the hardest tissues in the human body and constitutes the primary structural component of the skeletal system. It is characterized by a dense and rigid architecture as well as the biological capacity for continuous remodeling, thereby providing essential support and maintaining skeletal function.1 Bone tissue comprises cellular components—such as osteoblasts, osteoclasts, and osteocytes—alongside matrix elements including collagen and hydroxyapatite. Together, these components coordinate bone formation, resorption, and mechanical maintenance.2,3 Structurally, bone exists in two forms: cortical bone and cancellous bone. Cortical bone is compact and rigid, responsible for bearing the majority of mechanical loads, whereas cancellous bone is spongy, enriched with bone marrow and blood vessels, and facilitates material exchange and metabolic regulation.4,5 In addition to structural support and enabling movement, bone performs essential physiological functions, including protection of visceral organs, regulation of calcium–phosphorus metabolism, and hematopoiesis.6–8 With the global trend of population aging, the incidence of musculoskeletal disorders—including fractures, osteoporosis, infections, tumors, and degenerative diseases—has markedly increased.9 Current estimates indicate that approximately 1.71 billion people worldwide are affected by musculoskeletal diseases.10,11 These conditions compromise bone integrity by enhancing resorption, inhibiting osteogenesis, inducing local inflammation, or disrupting bone metabolism. As a result, they often cause bone injury, defects, or nonunion, severely impairing both the mechanical function and regenerative capacity of bone tissue.12 This pathological process not only diminishes patients’ quality of life but also imposes significant socioeconomic and healthcare burdens. Although several clinical strategies are currently employed to treat bone defects—such as autologous and allogeneic bone grafting, metal implants, and bone substitute materials13—these approaches remain limited by restricted donor availability, risks of donor-site morbidity, low rates of graft resorption and integration, immune rejection, and suboptimal restoration of bone structure and function (Table 1).14,15 Therefore, developing cost-effective and efficient bone regeneration strategies could yield substantial global health and economic benefits.
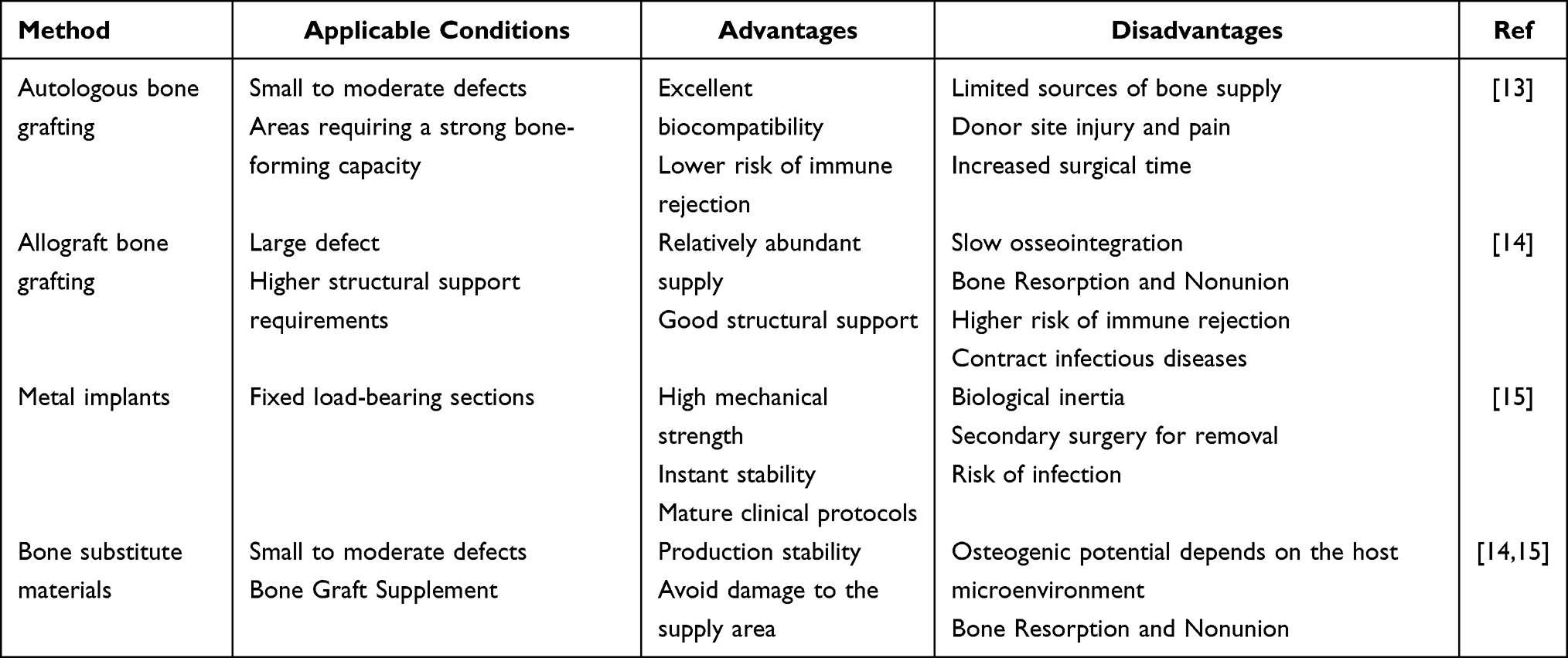 |
Table 1 Comparative Analysis of Clinical Bone Defect Repair Strategies |
In recent years, advances in regenerative medicine and tissue engineering have driven bone repair strategies toward greater precision and biomimicry. The development of biomimetic materials with structural and functional similarities to native tissue has shown considerable potential for reconstructing the microstructure, mechanical properties, and biological activity of bone, thereby offering new insights and technical support for repairing complex bone defects.16 Nanoparticles (NPs), defined as solid particles with dimensions of 1–100 nm, have attracted significant attention in the biomedical field due to their high surface area, excellent biocompatibility, and ease of surface modification.17 The integration of nanotechnology with regenerative medicine has further accelerated their application in drug delivery systems, bioactive carriers, and smart responsive platforms.18 In bone repair, nanoparticles not only enable controlled delivery of growth factors or drugs by mimicking the nanoscale architecture of bone but also modulate cellular behavior and the microenvironment, thereby promoting osteogenic differentiation and tissue regeneration.19,20 Through strategies such as surface modification and multifunctional coupling, nanoparticles can achieve targeted delivery, controlled release, and tissue-specific repair, establishing themselves as essential materials for bone regeneration research.21 However, conventional nanoparticles face significant obstacles in complex physiological environments, including immune clearance, nonspecific distribution, and limited ability to regulate biological signaling.22–24 To address these limitations, biomimetic design principles have been increasingly applied in nanomaterial development. By mimicking the structural and functional characteristics of natural cells, tissues, or biomolecules, biomimetic nanoparticles achieve enhanced biocompatibility, targeting, and functional integration.25,26 Representative strategies include cell membrane coating,27–29 exosome-inspired nanostructures,30,31 and protein- or peptide-based templating.32–34 These approaches significantly improve nanoparticle stability and recognition in vivo, while endowing them with multifunctional biological activities such as osteoinduction, immunomodulation, and neurovascular regeneration. Collectively, biomimetic nanoparticles represent a promising next-generation platform for more efficient and precise bone repair and regeneration.
This paper first reviews the structural composition of bone tissue and its microenvironment, along with the healing processes following bone injury. It then systematically summarizes the principal methods and design strategies for constructing biomimetic nanoparticles. Next, it highlights the mechanisms by which biomimetic nanoparticles contribute to bone regeneration. Finally, it discusses current development trends and future application prospects in bone repair, with the aim of providing valuable references and insights for the design of biomimetic nanoparticles and related biomedical research (Figure 1).
 |
Figure 1 Strategies for Constructing Biomimetic Nanoparticles Based on Different Biological Templates and Their Role in Bone Repair and Regeneration. |
Biological Basis of Bone Tissue Regeneration and Repair
Structure and Composition of Bone
Bone is a highly specialized dynamic connective tissue composed primarily of a small population of bone cells embedded within a large bone matrix. Bone tissue is maintained by coordinated interactions among osteoblasts, osteoclasts, and osteocytes, which collectively regulate matrix deposition, resorption, and homeostatic remodeling.35 The bone matrix consists of approximately 30% organic and 70% inorganic mineral components. The organic fraction is primarily composed of type I collagen, which accounts for over 90% of the organic matrix and lends bone its toughness and tensile strength. Non-collagenous proteins, such as osteopontin, osteocalcin, and proteoglycans, contribute to mineralization and mediate cell adhesion.36 The inorganic fraction is primarily composed of hydroxyapatite crystals, which are regularly deposited within collagen fibrils, conferring high hardness and compressive strength to bone.37 At the nanoscale, the organic and inorganic phases form a composite architecture that combines rigidity with elasticity and toughness, thereby enabling bones to withstand substantial external mechanical loads.38
Based on structural characteristics and density, bone tissue is classified into two types: compact bone and cancellous bone (Figure 2).39 Compact bone, or cortical bone, is dense and rigid. Its matrix contains abundant organic substances and inorganic salts,40 while bone cells are housed within small cavities called lacunae.41 Approximately 80% of the human skeleton is composed of cortical bone, primarily located in the shafts of long bones and in the outer layers of other bones. It serves as the main mechanical support of the body, capable of withstanding substantial tensile, compressive, and bending stresses.42 Mature compact bone exhibits a lamellar arrangement, penetrated by interlacing vascular and neural networks that form the Haversian system. This system, organized around osteons, provides essential nutritional and metabolic support to osteocytes.43 By contrast, immature cortical bone displays a woven architecture without distinct osteons, characterized by randomly oriented collagen fibers.44 During bone remodeling, involving resorption and new bone deposition, this woven bone is progressively replaced by mature lamellar bone with well-defined osteons.43 Cancellous bone, or spongy bone, is lightweight and porous, with a honeycomb-like structure composed of interconnected trabeculae. The intertrabecular spaces are filled with bone marrow and vascular tissue.45 Accounting for roughly 20% of skeletal mass, cancellous bone is predominantly found in the epiphyses of long bones, ribs, scapulae, and flat bones such as the cranium. Its high surface area allows effective dispersion and absorption of sudden mechanical impacts and supports elevated metabolic activity during bone remodeling.46 Moreover, the porous structure facilitates vascularization and hematopoiesis within the bone marrow, serving as a critical foundation for homeostasis and the continuous renewal of bone tissue.47
 |
Figure 2 The Composition and Structure of Compact Bone and Spongy Bone. |
The bone unit is the fundamental structural and functional element of mature cortical bone, representing the highly ordered microscopic architecture of bone tissue. Each osteon consists of concentric lamellae encircling a central Haversian canal.48,49 These canals run longitudinally along the bone axis and contain blood vessels, lymphatic vessels, and nerve fibers that provide essential nutritional and metabolic support to compact bone.6 Between the lamellae lie lacunae that house osteocytes, which are interconnected by canaliculi to form a three-dimensional communication network that facilitates intercellular signaling and material exchange.50 Different osteons are linked by transverse or oblique Volkmann’s canals, establishing an interwoven vascular network that connects with the blood supply of the periosteum and bone marrow cavity.51 This intricate tubular–lamellar architecture ensures sufficient vascularization within dense cortical bone and provides the structural foundation for mechanical adaptation and microdamage repair. The ordered arrangement of osteons confers cortical bone with exceptional resistance to bending, torsion, and compression, forming the microscopic basis for balancing high strength with high toughness in bone tissue.52
The Components of the Bone Microenvironment
The Extracellular Matrix in the Bone Microenvironment
The extracellular matrix (ECM) of bone tissue is a critical component of the bone microenvironment. It not only provides a structural scaffold for cell adhesion, growth, and migration but also regulates bone development, maintenance, and regeneration by mediating mechanical signal transduction, storing bioactive factors, and influencing cell fate.53 The bone ECM demonstrates high specificity and dynamic plasticity, consisting mainly of an organic matrix and inorganic minerals, whose synergistic interactions endow bone tissue with superior mechanical properties and biological activity.54 The organic fraction represents approximately 30% of the ECM’s dry weight, with type I collagen as the predominant constituent, comprising more than 90% of the organic portion. Type I collagen is the key molecule that imparts toughness and tensile strength to bone.55 Its ordered fiber arrangement not only provides a template for mineral deposition but also regulates osteoblast adhesion and differentiation. In addition, non-collagenous proteins (NCPs), including osteopontin, osteocalcin, osteoglycin, and proteoglycans, play essential roles in bone formation and remodeling.56,57 These molecules participate in matrix mineralization, mediate extracellular signaling, and regulate cell–matrix interactions. The inorganic fraction is composed primarily of hydroxyapatite crystals, which are deposited along the axial orientation of collagen fibers, forming nanoscale bone plates that confer compressive strength and structural stability to bone tissue.58 The nanostructural characteristics of the ECM—including collagen fiber diameter, spatial arrangement, and crystal size—directly affect the mechanosensing and responsiveness of osteogenic cells, thereby regulating their proliferation, differentiation, and migration.59 Moreover, the ECM undergoes continuous remodeling during osteogenesis and resorption, reflecting its dynamic plasticity.60 Matrix metalloproteinases (MMPs) and tissue inhibitors of metalloproteinases (TIMPs) synergistically coordinate ECM degradation and reconstruction to maintain the structural and functional homeostasis of bone.61 Furthermore, the ECM functions as a reservoir and controlled-release system for diverse growth factors, including bone morphogenetic proteins (BMPs), fibroblast growth factors (FGFs), and vascular endothelial growth factor (VEGF).62 These factors interact with the ECM to regulate osteogenesis, angiogenesis, and the immune microenvironment, thereby promoting bone regeneration. Consequently, mimicking the composition and nanostructure of the ECM has emerged as a central strategy for the design of biomimetic nanomaterials aimed at enhancing biological functionality and improving integration in bone defect repair.
Because the ECM provides structural support, mediates biochemical signaling, and regulates the local microenvironment, current ECM-inspired biomimetic nanoparticle design strategies can be broadly categorized into two complementary approaches. The first, interfacial biomimicry, focuses on reconstructing a cell-instructive interface by incorporating ECM-derived components (eg., collagen, gelatin, and hyaluronic acid) or adhesive motifs, together with precise control over ligand density and surface physicochemical properties to promote cell adhesion, spreading, and osteogenic differentiation. The second, functional biomimicry, seeks to recapitulate key matrix functions by integrating hydroxyapatite or bone-associated ions to mimic the mineral phase and by incorporating heparinized or sulfated shells, or multivalent binding sites, to enable the spatiotemporal presentation of bioactive factors such as BMPs and VEGF. Through these combined design elements, functional biomimicry provides osteogenic and pro-angiogenic cues that more closely align with the physiological sequence of bone regeneration.
Bone Immune Microenvironment
In recent years, advances in bone biology have highlighted that bone tissue functions not only as a structural support for mechanical forces but also as a highly dynamic site of immune regulation.63 The emergence of the field of “osteoimmunology” has revealed the intimate spatial and functional interactions between the skeletal and immune systems, thereby defining the bone immune microenvironment (Figure 3A).64,65 This microenvironment comprises diverse bone cells, including osteoblasts and osteoclasts, immune cells such as macrophages, T cells, B cells, and dendritic cells, as well as their secreted cytokines, chemokines, and bone-regulatory factors. Together, these components coordinate to maintain bone homeostasis and facilitate regenerative repair following injury.66
 |
Figure 3 Composition of the Bone Microenvironment. (A) Bone microenvironment as a loosely compartmentalized lymphoid organ: primarily comprising T* cells (memory T cells and circulating T cells), B* cells (B cells, memory B cells, and circulating mature B cells), stromal cells, M* cells (monocytes and their derivatives), and newly formed uncalcified osteoid matrix.65 (B) 3D imaging of blood vessels and perivascular cells under homeostasis and radiation stress.67 (C) Neural tissue within the femur.68 (i) A simplified schematic representation of the neuronal distribution in the mouse femur. (ii) A mosaic stitched, 3D reconstructed image of the bone marrow after surface rendering (scale bars, 1000 μm). (iii) Insets show 300 μm maximum intensity z-projections of PGP9.5-labeled nerve fibers are indicated in (ii). Arrow indicates the entry site of a nerve bundle into the marrow cavity (scale bars, 1000 μm). (iv) Insets show 200 μm z-projections of nerve fibers in the trabecular bone indicated in (ii). Nerve bundles enter through the distal epiphysis (arrows), where they branch out and terminate in the marrow cavity (arrowheads). Dotted lines indicate the growth plate (scale bars, 500 μm). (v) 300 (left) and 200 μm (right) z-projections of nerve fibers imaged at higher power, showing PGP9.5-labeled axons wrapping around or running parallel to blood vessels (open arrows) and branching out and terminating near the endosteum (arrows) or in the marrow cavity (arrowheads) (scale bars, 100 μm). (vi) A 100 μm z-projection showing PGP9.5-labeled axons following vessels in the marrow cavity (open arrows) and running through canals in the cortical bone (arrows) (scale bar, 100 μm). (vii, left) A 3D reconstructed image of PGP9.5-labeled nerve fibers taken in the lower diaphysis (scale bar, 200 μm). (Right) Insets show a 3D reconstruction and tracing of a cluster of complex nerve endings indicated in the white box viewed from the front (XY-plane; left) and side (YZ-plane; right). Asterisks indicate parent axons (scale bars, 50 μm). Dotted lines indicate cortical bone. (D) Neural tissue within the mandible.68 (i) A simplified schematic representation of the neuronal distribution in the mouse mandible. (ii) An 800 μm maximum intensity z-projection showing the distribution of inferior alveolar nerve branches through the mandible (scale bar, 500 μm). (iii) A 3D-reconstructed image of the dental pulp after surface rendering (scale bar, 500 μm). (iv) Schematic representation indicating the location from which (v–x) were sampled and viewed. (v) A 400 μm z-projection showing a bundle of nerve fibers that entered the molar through the apical foramen (arrows) and an accessory canal (arrowhead) (scale bar, 100 μm). (vi) A 150 μm z-projection showing nerve fibers running through the mandibular alveolar bone, including the marrow, and nerve terminal endings in the periodontal ligament (arrowheads). Arrows indicate PGP9.5-labeled cells in the alveolar bone (scale bar, 100 μm). (vii) An 80 μm z-projection showing nerve fibers running through the mandibular alveolar bone, including the marrow, and complex Ruffini-like endings in the periodontal ligament (open arrow) (scale bar, 100 μm). (viii) A 100 μm z-projection showing innervation of the gingiva around the tooth (scale bar, 200 μm). (ix) 100 μm z-projections showing a PGP9.5-labeled nerve bundle entering the apical foramen (arrow) or branching and terminating around the periodontal ligament in a “basket”-like manner around the molar root. The periodontal nerve fibers terminated as free endings (arrowheads) or Ruffini-like endings (open arrows) (scale bars, 100 μm). (x) A 100 μm z-projection showing complex Ruffini-like endings (open arrows) in the incisor periodontal ligament (scale bar, 500 μm). (E) Light sheet imaging of intact skeletal elements identifying lymphatic vessels within the knee joint69 (scale bar, 50 μm). Abbreviations: GP, growth plate; MP, metaphyseal; CB, condensed bone; DP, diaphyseal; Emcn, endomucin; PDGFRβ, platelet-derived growth factor beta (scale bars, 100 μm); CB, cortical bone; MC, marrow cavity; AB, alveolar bone; D, dentin; DP, dental pulp; G, gingiva; IAN, inferior alveolar nerve; M, muscle; PDL, periodontal ligament. |
Under physiological conditions, the bone marrow serves as the primary site for hematopoietic and immune cell production, providing a rich cellular reservoir for intramedullary immune activity.70 Resident immune cells, including macrophages, dendritic cells, and natural killer cells, not only mediate immune surveillance of microenvironmental changes (33872520) but also engage in cross-regulation with bone cells such as osteoblasts and osteoclasts through direct contact or paracrine signaling. Among these interactions, the RANK–RANKL–OPG axis represents a central pathway in osteoimmune regulation. RANKL, secreted by osteoblasts and activated T and B lymphocytes, induces osteoclast differentiation and maturation.71 Osteoprotegerin (OPG), acting as a soluble decoy receptor, competitively binds RANKL and prevents its interaction with RANK, thereby inhibiting osteoclast formation and maintaining the balance between bone resorption and formation.72 The subtypes and polarization states of immune cells also play essential roles in shaping the bone microenvironment. Macrophages dynamically transition between M1 (pro-inflammatory) and M2 (anti-inflammatory) phenotypes depending on local stimuli. This polarization modulates inflammatory responses73,74 and indirectly regulates osteoblast and osteoclast activity via cytokine secretion, thereby influencing bone remodeling.25 T cells display similarly diverse functions: subsets such as Th1, Th2, Th17, and Treg exhibit complex and distinct roles in bone metabolism. By secreting specific cytokines, they regulate the balance of bone resorption and formation and contribute to bone regeneration by maintaining immune homeostasis.75,76 Through these synergistic interactions and cross-regulatory mechanisms, immune cells establish a multidimensional and finely tuned network within the bone immune microenvironment. Collectively, this microenvironment is essential for preserving bone tissue function. Its highly integrated immunoregulatory mechanisms not only maintain metabolic equilibrium under homeostatic conditions but also enable rapid adaptive responses to internal and external perturbations, reflecting the multifunctional regulatory system unique to bone tissue.
Because bone regeneration follows a characteristic temporal sequence of inflammation, repair, and remodeling, immune-inspired biomimetic nanoparticle design generally focuses on reshaping the regenerative microenvironment through controlled modulation of inflammatory intensity and immune cell function. One strategy is to target, or preferentially interact with, key immune cells such as macrophages in order to promote pro-repair phenotypes (eg., M2-associated polarization) while attenuating excessive inflammation, thereby creating an immune milieu more conducive to osteogenesis and angiogenesis. A complementary strategy is to incorporate stimulus-responsive release mechanisms that are tailored to defect-site microenvironmental cues, enabling the spatiotemporal separation and staged delivery of immunoregulatory signals together with osteogenic or pro-angiogenic factors.
Vascularization in the Bone Microenvironment
As a highly vascularized and metabolically active tissue, the bone microenvironment regulates the dynamic equilibrium between osteoblasts and osteoclasts through immune cell activity, while the vascular system plays a pivotal role in bone development, remodeling, and regeneration (Figure 3B).67,77 The intraosseous vascular network supplies oxygen and nutrients to bone tissue and efficiently removes metabolic waste, thereby preserving local homeostasis and supporting normal skeletal function.78
Blood vessels in bone tissue are primarily located between trabeculae in cancellous bone and within the Haversian and Volkmann’s canals of cortical bone, forming a complex microcirculatory system with diverse functions.6 Recent studies have shown that intraosseous vessels display marked structural and functional heterogeneity, particularly H-type vessels, which are strongly associated with high osteogenic activity.79 These vessels regulate the proliferation, differentiation, and localization of bone progenitor cells through the secretion of angiogenic signaling molecules such as VEGF, Platelet-Derived Growth Factor (PDGF), and Notch ligands.80 In parallel, vascular endothelial cells establish synergistic regulatory networks with osteoclasts and immune cells, contributing to bone remodeling and immune homeostasis.81 The intricate interplay between the vascular system and bone cells forms the so-called “vascular–bone axis.” This functional unit plays a pivotal role in maintaining bone homeostasis and provides a theoretical basis and therapeutic targets for understanding bone repair and regeneration.82 Informed by the biological coupling between angiogenesis and osteogenesis, recent biomaterials and nanodelivery strategies have increasingly focused on improving bone defect repair by promoting vascularization, enhancing local perfusion, and coordinating pro-angiogenic and osteogenic cues in a spatiotemporally controlled manner.
Nerves in the Bone Microenvironment
Bone tissue functions not only as a critical organ for mechanical support and mineral storage but is also extensively innervated by a complex neural network.14 Anatomically, similar to the intricate vascular system within bone, neural fibers often accompany blood vessels and are widely distributed throughout cortical bone, cancellous bone, bone marrow, and the periosteum (Figure 3C and D).68,83,84 Through extensive branching and interconnections, these nerve fibers facilitate signal transmission and nutritional regulation across skeletal structures. Moreover, they act synergistically with bone marrow mesenchymal stem cells (BMSCs), osteoblasts, osteoclasts, and vascular endothelial cells to maintain metabolic homeostasis and dynamic equilibrium within bone tissue.85
Intraosseous nerves consist primarily of sensory and sympathetic fibers, which together form a complex neural regulatory system involved in bone formation, continuous tissue renewal, and the modulation of repair mechanisms.86 Sensory nerve endings release neuropeptides such as calcitonin gene-related peptide (CGRP) and substance P (SP), directly influencing the activity of osteoblasts and osteoclasts to promote bone formation or inhibit bone resorption.87 By contrast, sympathetic nerves indirectly suppress osteogenesis and enhance bone resorption through norepinephrine secretion, which acts on β-adrenergic receptors, thereby contributing to the inhibition of bone formation within metabolic regulation.88 The neural network within bone tissue not only maintains metabolic balance under physiological conditions but also plays a pivotal role in bone repair. Axonal regeneration, neurotrophic factor release, and coordinated vascular reconstruction are considered essential biological events in bone regeneration.88,89 Therefore, advancing our understanding of neural regulatory mechanisms in bone tissue is critical for elucidating the pathophysiology of bone regeneration disorders and for developing neuro-mediated therapeutic strategies.
Building on the roles of neural signaling in bone regeneration—including cell recruitment, inflammatory regulation, and coordinated neurovascular reconstruction—recent studies have increasingly incorporated neuroregulatory principles into the design of biomimetic nanodelivery systems. One approach uses nanoplatforms to achieve the localized enrichment and controlled release of neuropeptides, such as CGRP and SP, as well as neurotrophic factors, including NGF and BDNF, thereby providing phase-appropriate neural cues during regeneration. Another strategy exploits the influence of sympathetic signaling on bone metabolism by modulating the neural–immune–bone signaling axis, thereby indirectly optimizing the regenerative microenvironment and promoting bone repair.
Lymphatic Vessels in the Bone Microenvironment
Within the bone microenvironment, the lymphatic system is a critical regulator of immunity and fluid homeostasis. Although its spatial distribution is relatively restricted compared with the vascular system, its biological functions are indispensable. Recent studies have identified abundant lymphatic structures within bone tissue, particularly in the periosteum, cancellous bone, and perimedullary regions (Figure 3E).69 These structures maintain interstitial fluid balance, regulate local immune responses, and facilitate metabolic waste clearance.90–92 In addition, the lymphatic network contributes to antigen presentation, inflammatory signaling, and immune cell trafficking, thereby indirectly modulating the dynamic balance between osteogenesis and osteoclast activity.88 Under pathological conditions such as bone injury, inflammatory disease, and tumor-related bone disorders, functional alterations in lymphatic vessels within bone or adjacent tissues can markedly reshape the local immune environment, compromising the stability of the bone regeneration microenvironment.93 Therefore, elucidating the spatial distribution, structural features, and regulatory mechanisms of lymphatic vessels within the bone microenvironment is essential for advancing a systematic understanding of immune–bone interactions in both homeostasis and disease.
Given the roles of lymphatic vessels in tissue fluid drainage and immune cell trafficking, recent studies have begun incorporating lymphatic-associated processes into the design of nanodelivery systems for bone regeneration. Such strategies typically employ inflammation-responsive nanoplatforms to achieve controlled release of immunomodulatory factors, while optimizing particle size and surface physicochemical properties to enhance accumulation at the defect site. Together, these design features.
The Healing Process of Bone Tissue Injury
The healing of bone tissue is a highly dynamic and tightly regulated biological process that integrates multiple cellular and molecular signaling pathways, with the primary goal of restoring structural integrity and mechanical function.94 Bone healing is generally classified into two types: primary (direct) healing and secondary (indirect) healing.95
When displacement or compression at the site of bone injury is mild and occurs within a stable mechanical environment, bone repair proceeds through primary healing.96 This process predominantly relies on intramembranous ossification, closely resembling normal bone remodeling, and is most common in flat bones such as the cranium, mandible, and clavicle.97 Bone marrow mesenchymal stem cells, osteoblasts, and osteoclasts collectively contribute to repair. In the early phase, osteoclasts migrate to the injury site to remove necrotic and damaged matrix,98 while BMSCs differentiate into osteoblasts under the regulation of signaling pathways such as BMPs and Wnt/β-catenin. These osteoblasts secrete type I collagen and other extracellular matrix components, promoting hydroxyapatite deposition and gradually forming dense, lamellar bone.99 Ultimately, the newly formed lamellae organize concentrically around the central canal, reconstructing the Haversian system of mature cortical bone and restoring both structural integrity and mechanical function.100 However, in most clinical cases, inadequate stability or rigidity of external fixation devices results in displacement at the injury site. Under these conditions, bone healing typically follows the classical multi-stage secondary healing process.101 Based on dynamic histological changes during regeneration, this process can be divided into four sequential stages: hematoma and inflammatory response, soft callus formation, hard callus formation, and bone remodeling (Figure 4).102
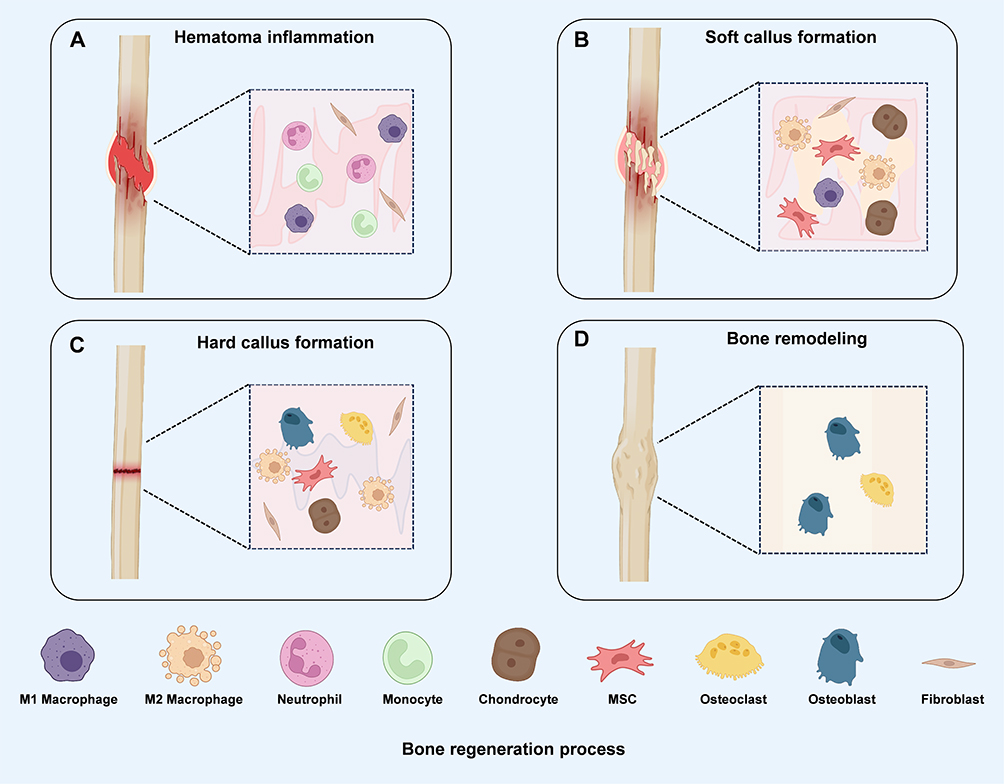 |
Figure 4 Schematic Diagram of Bone Tissue Injury Healing Process. (A) Hematoma and Inflammatory Response Phase: Following fracture, a hematoma forms at the injury site and is accompanied by infiltration of inflammatory cells, primarily neutrophils, monocytes, and M1 macrophages, along with fibroblasts, thereby initiating the early inflammatory response. (B) Formation of Soft Healing Tissue Phase: As the inflammatory response gradually subsides, macrophages begin to transition toward a reparative phenotype, while mesenchymal stem cells, chondrocytes, fibroblasts, and macrophages collectively contribute to soft callus formation. (C) Formation of Hard Healing Tissue Phase: During endochondral ossification, osteoblasts and osteoclasts, together with MSCs, chondrocytes, fibroblasts, and reparative macrophages, promote mineralized callus formation and bone consolidation. (D) Bone remodeling phase: Newly formed bone is further remodeled through the coordinated activities of osteoblasts and osteoclasts, gradually restoring the structural integrity and function of the injured bone. |
The hematoma and inflammatory phase represent the initial and indispensable stage of bone healing.103 Following bone injury, blood vessels in the surrounding soft tissues rupture, leading to blood extravasation and the formation of a hematoma enriched with platelets and fibrin.104 This hematoma not only seals the injury site but also acts as a reservoir for bioactive factors, releasing signaling molecules such as PDGF, transforming growth factor-β (TGF-β), and VEGF to promote angiogenesis and recruit mesenchymal stem cells.105 Concurrently, neutrophils, monocytes, and macrophages migrate to the site, where they remove necrotic tissue and cellular debris by secreting inflammatory mediators including IL-1β, TNF-α, and IL-6, while simultaneously activating osteogenesis-related signaling pathways.95 Subsequently, the process transitions into the soft callus formation phase. Under the influence of multiple growth factors in the bone microenvironment, subperiosteal osteogenic progenitors, BMSCs, and perivascular cells differentiate into chondrocytes and osteoblasts.106 The locally generated hyaline cartilage matrix, rich in type II collagen and proteoglycans, provides preliminary mechanical stability and partially supports external loading.107 At the same time, some BMSCs differentiate directly into osteoblasts, depositing non-mineralized collagen matrix that serves as a scaffold for subsequent mineralization.108 Meanwhile, VEGF secreted by the cartilage matrix stimulates vascular invasion into the injury site, enhancing local oxygenation and establishing conditions necessary for hard callus formation.109
During the hard callus formation phase, angiogenesis further intensifies, resulting in a marked increase in local oxygen partial pressure. This enhanced oxygenation suppresses the continued differentiation of chondrocytes while simultaneously promoting osteoblast activation and bone matrix deposition.110 As osteogenesis progresses, hyaline cartilage is gradually replaced by calcified cartilage and ultimately by lamellar bone secreted by osteoblasts.111 In parallel, intramembranous ossification occurs in the subperiosteal region, directly generating bone plate structures. Hydroxyapatite crystals deposit within collagen fiber interspaces, conferring high compressive strength and structural stability to the newly formed bone tissue, thereby markedly improving its mechanical properties.112
In the bone remodeling phase, osteoclasts initially resorb woven bone, after which osteoblasts deposit structurally dense lamellar bone to reconstruct the Haversian system characteristic of mature cortical bone.113 This process depends on the functional coordination between osteoclasts and osteoblasts, primarily regulated by the RANKL–RANK signaling pathway, to maintain dynamic equilibrium between bone resorption and formation.114 As trabeculae and cortical bone gradually align with mechanical loading, the tissue’s resistance to bending, torsion, and compression markedly increases. Ultimately, the anatomical architecture and function of the medullary cavity and vascular network are restored to steady-state levels, thereby fully re-establishing the mechanical support and metabolic functions of bone tissue.115
The endogenous bone-healing process depends critically on the proper initiation and timely resolution of inflammation, effective vascular regeneration with restoration of tissue oxygenation, and precise coordination of osteoblast–osteoclast coupling. In clinically common pathological conditions—including aging and osteoporosis, diabetes, infection, post-tumor resection defects, and nonunion—the defect microenvironment is often characterized by persistent or dysregulated inflammation, elevated oxidative stress, impaired angiogenesis, and compromised reparative cell function. As a result, healing may remain arrested in the inflammatory or cartilaginous stages, ultimately leading to delayed union or nonunion. In this context, biomimetic nanoparticles provide several rational strategies to overcome pathological barriers to bone repair. First, immune-biomimetic and immunoregulatory designs can facilitate the transition from inflammation to repair by modulating macrophage phenotypes and limiting excessive release of inflammatory mediators. Second, strategies aimed at restoring vascularization and perfusion can enhance angiogenic–osteogenic coupling and improve local oxygenation through the delivery of pro-angiogenic factors or the incorporation of microenvironment-responsive release mechanisms triggered by hypoxia, reactive oxygen species (ROS), or pH changes. Third, ECM and mineral phase–mimetic approaches can partially compensate for the compromised osteogenic niche under pathological conditions. In more complex settings, such as infectious or tumor-associated bone disease, nanoplatforms may further integrate antimicrobial and regenerative functions to simultaneously address pathological control and tissue reconstruction.
Strategies for Constructing Biomimetic Nanoparticles Based on Different Biological Templates
In bone tissue engineering and regenerative medicine, the development of biomimetic nanoparticles has emerged as a pivotal strategy to improve bone repair efficiency and regeneration outcomes.116 These nanoscale platforms achieve precise biomimicry of natural biological components through modifications in chemical composition, spatial configuration, and surface functionalization. Such designs enable high compatibility with host tissues in vivo while effectively replicating—or even amplifying—critical signaling pathways and biological functions involved in bone repair.25 This approach not only enhances immune regulation within the bone microenvironment but also promotes osteoblast differentiation, accelerates mineralization, and enables targeted drug delivery to sites of injury. In this review, we summarize construction strategies for biomimetic nanoparticles based on diverse biological templates, including cell membrane-based, exosome-based, protein template-based, and peptide template-based systems (Table 2). Table 3 outlines the advantages and limitations of these biomimetic nanoparticle categories, providing a theoretical framework for their rational selection and optimization in bone regeneration applications.
 |
Table 2 Comparative Analysis of Bionic Nanodelivery Strategies: Composition, Preparation, Targeting Mechanisms, and Therapeutic Functions |
 |
Table 3 Advantages and Disadvantages of Strategies for Constructing Biomimetic Nanoparticles Based on Different Biological Templates |
Cell Membrane-Based Nanoparticles
As natural biological interfaces, cell membranes exhibit exceptional biocompatibility. Their phospholipid bilayers encapsulate diverse functional proteins that are essential for signal transduction, cell recognition, and immune regulation.126 Membranes derived from different cell sources preserve the key biological traits of their parent cells, including “self” markers, targeting capabilities, and specific interactions with the immune system.27,127 Since the 1980s, cell membranes have been extensively investigated as natural carriers for drug delivery, enabling targeted transport and controlled release within the body.33 More recently, techniques such as hypotonic treatment and ultrasonic extrusion have been employed to coat various nanoparticle surfaces with cell membranes. The resulting nanoplatforms integrate the biological attributes of cell membranes with the physicochemical properties of nanoparticles, establishing themselves as a cutting-edge focus in active targeted drug delivery research.27 To date, this biomimetic strategy has been applied to multiple cell types, including erythrocytes,28 platelets,122 cancer cells,117 and macrophages.117
Red blood cells (RBCs) are the most abundant and longest-circulating cells in the human bloodstream, with a lifespan of approximately 100–120 days.128 Accordingly, they were among the earliest cell types investigated and applied in cell therapy. The erythrocyte membrane preserves multiple functional proteins, including CD47, C8bp, and HRP, which interact with signal-regulatory protein alpha (SIRPα) to transmit “self” recognition signals to the immune system, thereby suppressing phagocytic clearance by immune cells.129 This immune evasion mechanism prolongs the in vivo half-life of RBCs and extends the circulation time of RBC membrane-coated nanoparticles. Moreover, the absence of a nucleus and most organelles results in minimal cytoplasmic contents, reducing interference during membrane separation and extraction. This facilitates the acquisition of structurally intact and functionally preserved membranes for nanoparticle coating.130,131 Ji et al developed icariin (ICA)-loaded nanoparticles (iRINPs) coated with erythrocyte membranes functionalized with the tumor-penetrating peptide iRGD, creating a biomimetic camouflage layer to overcome ICA’s limitations, including poor solubility, high hydrophobicity, and limited tumor penetration. The iRINPs significantly enhanced ICA solubility, biocompatibility, and stability while markedly reducing macrophage uptake. In a lung cancer model, they inhibited A549 cell proliferation, migration, and invasion, demonstrating superior therapeutic efficacy compared with ICA alone.28 Overall, RBC membranes offer excellent biocompatibility, low immunogenicity, prolonged circulation, and facile membrane separation, making them a promising platform for biomimetic drug delivery and precision medicine. Nevertheless, additional functionalization strategies are required to overcome inherent limitations, particularly their lack of intrinsic tumor-targeting ability.
Platelets are naturally occurring, anucleate, disc-shaped cells in the bloodstream with a diameter of approximately 2–3 μm. They are released from megakaryocytes in the bone marrow during maturation.132 Beyond their central role in hemostasis and coagulation, platelets contribute to a range of physiological and pathological processes, including angiogenesis, immune regulation, and inflammation.133,134 Platelet membranes are enriched with adhesion molecules and receptors that promote preferential accumulation at ischemic or injured sites in the circulation.135 Accordingly, platelet membrane–derived biomimetic nanoparticle delivery systems have attracted growing interest. Nevertheless, difficulties in platelet isolation and ex vivo processing remain major barriers to clinical translation. Hu et al developed platelet membrane–cloaked nanoparticles (PNPs) by coating poly(lactic-co-glycolic acid) (PLGA) nanoparticles with human platelet membranes. These PNPs were separately loaded with docetaxel and vancomycin, yielding the PNP-DtX1 and PNP-Vanc delivery systems. Their therapeutic efficacy was subsequently evaluated in rat models of coronary artery injury and mouse models of systemic bacterial infection. Results demonstrated that PNP-DtX1 effectively suppressed neointimal hyperplasia induced by balloon injury, while PNP-Vanc significantly reduced bacterial loads in multiple organs of infected mice (Figure 5A and B).122
 |
Figure 5 Strategies for Constructing Cell Membrane-Based Bionic Nanoparticles (A) Enveloping poly(lactic-co-glycolic acid) (PLGA) nanoparticles with human platelet membranes endows them with platelet-like properties, conferring immunocompatibility, endothelial cell binding, and pathogen adhesion capabilities, thereby significantly enhancing in vivo stability and targeting efficiency.122 (B) Schematic of membrane-coated photosensitizer nanocarrier (MON) preparation and photosensitizer nanocarrier (ON) depolymerization processes. (C) Schematic of platelet membrane-cloaked nanoparticle (PNP) preparation. (D) Schematic of MON in vivo circulation, tumor targeting, and immune response induction during photodynamic therapy.136 |
Tumor cells exhibit a range of distinctive biological characteristics, including unlimited proliferative capacity, immune evasion, and homing and adhesion properties mediated by specific membrane proteins.137,138 Exploiting these features, nanoparticles cloaked with tumor cell membranes can achieve active targeting and selective adhesion to tumor tissues, thereby improving drug delivery efficiency and antitumor efficacy.139 For example, Meng et al engineered a peptide delivery system (SPIO NP@M-P) by encapsulating superparamagnetic iron oxide nanoparticles within lung cancer H460 cell membranes and conjugating them with a PD-L1 inhibitor peptide (TPP-1) and an MMP2 substrate peptide. This multifunctional system integrated homologous targeting, enzyme-triggered release, and magnetic resonance imaging capabilities. It significantly prolonged peptide half-life in vivo, enhanced T-cell activation, and inhibited tumor growth, demonstrating potential as an integrated platform for tumor diagnosis and therapy.117 Similarly, Wang et al first prepared ovalbumin (OVA) nanoparticles loaded with the photosensitizer Ce6 (ON) and subsequently cloaked them with B16-OVA cancer cell membranes, yielding membrane-coated nanoparticles (MON) with homotypic targeting capability. Under laser irradiation, MONs enhanced OVA antigen cross-presentation through ROS generation, effectively inducing immune cascade reactions that achieved complete tumor clearance and durable immune memory in tumor-bearing mice. Compared with conventional photodynamic therapy (PDT), this strategy substantially expanded PDT’s immunological potential, offering a novel and highly efficient approach to photodynamic immunotherapy (Figure 5C and D).136
Macrophage-coated nanoparticles exhibit remarkable immune evasion capacity due to the protective function of membrane-derived proteins, thereby extending their half-life in systemic circulation.140 In addition, the abundance of inflammation-related receptors on macrophage membranes mediates their inherent chemotaxis toward inflammatory microenvironments, enabling selective nanoparticle accumulation at lesion sites and conferring tissue-targeting properties. This significantly enhances the precision of drug delivery and therapeutic efficacy.141,142 Lu et al developed a biomimetic anti-inflammatory nanomedicine system (MM-CEP/NLCs) for the treatment of acute lung injury by encapsulating cefpodoxime (CEP) in nanostructured lipid carriers (NLCs) coated with macrophage membranes (MMs). This platform integrates the superior physicochemical properties of nanostructured lipid carriers with the inflammatory homing and immune regulatory functions of macrophage membranes. It achieves targeted accumulation and sustained drug release at pulmonary inflammatory sites while markedly reducing lung injury parameters, including pulmonary edema, bronchoalveolar lavage cell counts, inflammatory cell infiltration, and cytokine levels, thereby demonstrating promising potential in the treatment of inflammatory diseases.29
Within cell membrane–based biomimetic nanostrategies, recent studies have extended beyond conventional membrane templates—such as erythrocyte, platelet, and immune cell membranes—to incorporate membranes derived from cells directly involved in bone regeneration and remodeling. This shift enables more precise engagement of the key regulatory processes that govern the bone-defect microenvironment. In particular, stem cell membranes have attracted interest because of their lesion-homing capacity, low immunogenicity, and potential to modulate the local microenvironment. Accordingly, they have been used to enhance nanocarrier accumulation and retention at defect sites while providing a biomimetic interface for the integration of osteogenic, angiogenic, and immunomodulatory functions.143,144 Dong et al incorporated BMSC membrane–coated, ultrasound-responsive barium titanate nanoparticles into carboxymethyl cellulose-based hydrogels to create a dual-functional platform for diabetic bone regeneration. This system improved repair by simultaneously reshaping the immune microenvironment and enhancing osteogenic stimulation: CMS promoted macrophage polarization from the pro-inflammatory M1 phenotype toward the pro-reparative M2 phenotype via the PI3K–Akt–mTORC1 axis, whereas the released nanoparticles generated moderate levels of ROS under ultrasound stimulation, thereby activating Wnt/β-catenin signaling and enhancing BMSC proliferation and osteogenic differentiation.143
Osteoblast-derived membranes represent another functionally relevant template, as their surface adhesion molecules and bone-associated receptors can improve bone-site targeting and support a more favorable osteogenic microenvironment, thereby promoting osteogenic differentiation and mineralization.145 In this context, Li et al developed a bone-targeted delivery platform (OM/Cur@NPs) by coating curcumin-loaded PLGA nanoparticles with osteoblast-like membranes enriched in CXCR4 and tumor necrosis factor-α receptors. This design improved the in vivo stability of curcumin and increased accumulation at bone sites. In ovariectomized mice, the platform achieved greater bone enrichment and attenuated the inflammatory bone microenvironment through TNF-α neutralization, thereby exerting dual effects of promoting osteogenesis and suppressing bone resorption.146
Because cell membranes are directly derived from the host organism, most cell membrane–coated nanoparticles display excellent biocompatibility, biodegradability, and low immunogenicity. Nevertheless, their potential risks cannot be ignored. Certain cellular components and secreted molecules may exert toxic effects. For example, hemoglobin in blood has been reported to induce neuronal death,147 while bioactive factors released by platelets may exhibit neurotoxicity.148 Therefore, comprehensive safety assessments are essential before clinical application of biomimetic cell membrane–coated nanoparticles. In addition, the structural complexity of these systems often limits their drug-loading capacity. Despite these limitations, cell membrane–coated nanoparticles hold considerable promise for active tumor targeting due to their intrinsic membrane proteins and biorecognition properties. However, their mechanisms of action and safety profiles require further systematic investigation.
Exosome-Based Nanoparticles
Exosomes (EVs) are nanoscale lipid bilayer vesicles secreted by diverse cell types to mediate intercellular communication. With diameters of approximately 30–150 nm, they are widely distributed in bodily fluids such as blood, urine, and saliva. EVs originate from intracellular multivesicular bodies (MVBs) and are released into the extracellular microenvironment through exocytosis.149,150 They play pivotal roles in numerous physiological and pathological processes by transporting nucleic acids, proteins, and small molecules from donor cells to recipient cells, thereby regulating cellular biology and inducing functional or phenotypic changes.151–153 Owing to their broad distribution, nanoscale size, and natural tropism for specific tissues and cells, EVs are increasingly recognized as promising candidates for drug delivery systems and diagnostic biomarkers.154
Multiple studies have confirmed that exosomes, as key mediators of intercellular communication, possess selective recognition capabilities for specific target cells and display natural tropism toward particular tissues.155 This unique ability to interact with target cells enables exosomes to play pivotal roles not only in maintaining physiological homeostasis and mediating pathological processes but also as ideal natural carriers for therapeutic agents, thereby advancing precision medicine.30 Currently, drug loading into EVs is achieved through two primary strategies: indirect and direct methods (Figure 6A).116,123 Indirect loading exploits the endogenous biogenesis of EVs, during which therapeutic molecules are incorporated by donor cells and subsequently packaged into vesicles. This approach mainly includes two strategies: (i) co-incubation, in which donor cells are exposed to therapeutic molecules that are internalized and secreted into EVs through intrinsic uptake and secretion pathways;118 and (ii) genetic engineering, in which specific genes or proteins are introduced into donor cells via transfection or gene editing to direct selective packaging of RNA, proteins, or functional molecules into EVs.119 For example, Wang et al co-incubated paclitaxel (PTX) and doxorubicin (DOX) with breast and ovarian cancer cells, yielding exosomes enriched with these drugs.156 Similarly, Liu et al overexpressed miR-20a in BMSCs, generating EVs that promoted BMSC migration, osteogenesis, and enhanced osseointegration of porous titanium alloys in osteoporotic rats.31 Direct loading, in contrast, bypasses donor cell metabolism by modifying EV membrane permeability to allow drug entry. This method increases loading efficiency and is particularly suited for hydrophilic or macromolecular drugs.157 Common strategies include passive co-incubation and active electroporation. In passive co-incubation, isolated EVs are directly incubated with therapeutic molecules, enabling drug entry via diffusion or affinity.158 For instance, Wei et al loaded DOX into BMSC-derived EVs by co-incubation, followed by desalting and dialysis, producing Exo–Dox vesicles that enhanced uptake, inhibited osteosarcoma MG63 proliferation, and exhibited reduced cardiotoxicity in H9C2 cells.159 Electroporation, by contrast, generates transient pores in EV membranes under high voltage, facilitating efficient encapsulation of hydrophilic or large molecules such as siRNA.157,160 Faruqu et al successfully employed electroporation to load siRNA into exosomes derived from human embryonic kidney cells, achieving efficient delivery into cancer cells and establishing a standardized protocol for siRNA-based EV therapeutics (Figure 6B).161
 |
Figure 6 Different Methods for Exosome Isolation and Drug Encapsulation. (A) Commonly Used Exosome Isolation Method.157 (B) Commonly Used Exosome Drug Encapsulation Methods.157 (C) Schematic diagram of exosome-mediated delivery of different functionalized payloads. Following separation and purification, exosomes can be engineered to acquire specific targeting capabilities through surface modification or by covalent incorporation of functional motifs. Therapeutic payloads may be encapsulated within the exosomal lumen or conjugated to its surface. Upon uptake by target cells, the therapeutic motifs exert their intended effects. |
Although exosome-based therapeutic strategies have not yet received clinical approval, early clinical trials have demonstrated their favorable safety profile. Once regarded merely as cellular metabolic waste, exosomes are now increasingly recognized as promising delivery platforms for diverse therapeutic agents, with their roles in intercellular communication being progressively clarified. Compared with cell membrane-coated nanoparticles, exosome-based delivery systems offer several advantages, including high endogenous loading capacity, low immunogenicity, ease of modification, and enhanced cellular internalization efficiency. Nonetheless, clinical translation remains hindered by significant challenges, such as the development of efficient separation and purification methods, scalable production processes, and appropriate storage conditions (Figure 6C). Moreover, a deeper understanding of the uptake and transport mechanisms of EVs and their subpopulations in recipient cells is essential to advance their broad application in targeted therapies.
Protein Template-Based Nanoparticles
Protein-templated nanoparticles have emerged as an important research direction in nanomedicine. This strategy exploits the three-dimensional structure, functional group distribution, and self-assembly capacity of proteins as natural templates to guide nanoparticle nucleation and ordered growth.32 By mimicking biomineralization and biomolecular assembly, such approaches enable the synthesis of nanoparticles that closely resemble natural structures in size, morphology, and functionality. Based on design strategies, these nanoparticles can be broadly classified into three categories: protein–drug conjugates, engineered therapeutic proteins, and combinatorial complex platforms utilizing protein motifs.124,125 Protein-templated nanoparticles offer the advantage of personalized functional design through surface modification, while also improving drug targeting and minimizing off-target effects. As a result, they show great promise in cancer therapy, bone regeneration, and precision treatment of chronic diseases.120 Albumin, a highly soluble and stable natural protein, possesses low toxicity, excellent biocompatibility, and a long in vivo half-life.120 Its molecular surface contains multiple functional groups amenable to chemical modification, making it an ideal carrier for drug delivery systems. Albumin-based nanocarriers effectively improve drug stability, solubility, and tissue targeting.162 Zhu et al developed a novel biomimetic bone repair material, TD-BNP@DBBM, by loading thiazolidine-2,5-dione (TD) onto bovine serum albumin nanoparticles (BNPs) and combining them with deproteinized bovine bone mineral (DBBM). Experimental findings demonstrated that TD-BNP@DBBM significantly enhanced the osteogenic differentiation of MC3T3-E1 cells and accelerated bone regeneration in a rat cranial defect model, a process potentially associated with activation of the Wnt/β-catenin signaling pathway (Figure 7).163 Similarly, MinJoo Kim et al employed bovine serum albumin (BSA) to modify nanoparticle surfaces for biomimetic functionalization. Compared with scaffolds containing unmodified nanoparticles, BSA-coated scaffolds exhibited superior protein adsorption capacity and controllable degradation behavior, while significantly improving MC3T3-E1 cell adhesion, proliferation, and alkaline phosphatase (ALP) activity.164
 |
Figure 7 Schematic diagram of TD-BNP@DBBM preparation and its application in treating critical-sized cranial defects. (A) TD-BNP@DBBM preparation process.163 (B) Mechanism of action by which TD-BNP@DBBM promotes bone regeneration.163 (C) Schematic diagram of TD-BNP@DBBM application in repairing large-area bone defects in rats.163 (D) Schematic diagram of the process for constructing engineered EVs for bone targeting.165 |
Transferrin (Tf) is a natural protein widely employed in drug delivery systems. Its primary physiological role is to mediate iron transport across cell membranes by binding to transferrin receptors (TfR) on the cell surface, thereby maintaining cellular iron homeostasis and normal physiological function.166 The high expression of transferrin receptors on many tumor cells allows this receptor-mediated endocytosis pathway to serve not only in regulating iron metabolism but also as an efficient targeting mechanism for drug and gene delivery.167 As a result, transferrin and transferrin-derived nanoparticles show considerable promise in antitumor therapy and in facilitating drug transport across the blood–brain barrier. For example, Peng et al utilized an adenoviral vector to upregulate transferrin receptor expression in prostate cancer cells, which significantly enhanced the selective uptake and intracellular accumulation of transferrin–doxorubicin conjugates (Tf-DOX). This approach highlights transferrin’s pivotal role in precision delivery and antitumor therapy, offering novel insights for targeted cancer treatment.168
Protein-templated nanoparticles generally consist of an outer surface and an inner core, a structural feature that allows drug loading through two mechanisms: adsorption of drug molecules onto the outer surface or encapsulation within the inner core for storage and delivery. Modification of the protein template can further improve loading efficiency—for example, rendering albumin hydrophobic to accommodate hydrophobic drugs169 or cationizing proteins to bind nucleic acids.170 Once internalized by cells, these nanoparticles typically release their payload in a controlled manner through gradual protease-mediated degradation. Although proteins are often derived from endogenous molecules and therefore exhibit relatively low in vivo toxicity, preparation methods frequently require the use of harmful crosslinking agents, such as glutaraldehyde, to enhance stability. Consequently, the development of safer and more efficient preparation strategies remains a major challenge in this field.
Peptide Template-Based Nanoparticles
Targeting peptides are short peptide molecules that specifically recognize surface markers on cells with high affinity and selectivity.171 Compared with other targeting ligands, they provide multiple advantages, including higher targeting efficiency, straightforward synthesis, excellent biocompatibility, and low immunogenicity.172 In nanomedicine delivery systems, nanoparticle modification with targeting peptides markedly improves their accumulation in target tissues or cells, thereby enhancing drug delivery specificity and therapeutic efficacy.121 Consequently, a wide range of targeting peptides has been designed and applied in targeted therapies for diverse diseases, offering promising strategies for precision medicine.
DSS6 is an acidic, bone-targeting oligopeptide widely used to confer bone-selective accumulation on delivery systems. Its multiple aspartic acid residues exhibit strong affinity for hydroxyapatite within the bone mineral matrix, thereby promoting carrier localization and retention at bone surfaces.173,174 Accordingly, DSS6 is frequently grafted onto nanoparticles, liposomes, and extracellular vesicles as a bone-targeting module to enhance local delivery efficiency, tissue selectivity, and bioavailability in bone defect repair applications.175 Using this strategy, Zheng et al developed an engineered delivery platform for aging-associated bone repair based on extracellular vesicles derived from juvenile mouse serum. Surface modification with DSS6 enhanced EV accumulation and therapeutic efficacy at bone repair sites. Compared with unmodified EVs, DSS6-engineered EVs more effectively promoted fracture healing in aged mice. Mechanistically, these engineered EVs appeared to improve mitochondrial function in senescent cells and restore the stemness of BMSCs by activating the Tomm7-mediated Pink1/Parkin mitophagy pathway, thereby remodeling the senescent bone microenvironment.165
Peptides enriched in aspartic acid (Asp) display strong affinity for bone tissue due to electrostatic interactions between their carboxyl groups and calcium ions in hydroxyapatite (HAp), the primary mineral component of bone.176 Studies have shown that bone-targeting efficacy is closely correlated with the number of exposed Asp residues in the peptide chain, with higher residue counts conferring greater binding affinity.177 Among commonly used bone-targeting peptides, the d-aspartic acid octapeptide (Asp8), composed of eight Asp residues, exhibits particularly high affinity for bone tissue and is therefore widely employed in bone-targeted drug delivery and bone regeneration research.178 For instance, Wang et al developed Asp8-modified dendritic platinum–copper alloy nanoparticles (Asp-DPCN) and systematically evaluated their bone-targeting and antitumor properties. Compared with unmodified DPCN, Asp-DPCN showed stronger binding to hydroxyapatite and bone fragments in vitro and exhibited enhanced accumulation within the bone tumor microenvironment in vivo. Exploiting this property, Asp-DPCN significantly increased local tumor temperatures during photothermal therapy, effectively suppressing tumor growth while reducing osteoclast-mediated bone destruction.179 Similarly, the DSS6 peptide, composed of six Asp residues rich in carboxyl side chains, achieves high-affinity binding to bone tissue through strong coordination with calcium ions in hydroxyapatite crystals.180,181 This binding not only promotes preferential accumulation of drugs or nanocarriers in bone tissue but also elevates local drug concentrations, thereby enhancing therapeutic efficacy while minimizing systemic side effects. Zheng et al designed a bone repair platform by extracting engineered exosomes from juvenile mouse serum and modifying their surfaces with DSS6 peptides. Compared to unmodified EVs, DSS6-engineered EVs showed improved enrichment at bone repair sites, promoted osteogenesis and angiogenesis, and restored the pluripotency of BMSCs through activation of the Tomm7-mediated Pink1/Parkin mitophagy pathway, ultimately reversing the impaired microenvironment associated with aging bone.165
Peptide-templated nanoparticles exhibit strong binding affinity for specific targets, along with excellent biocompatibility and low immunogenicity. Current studies indicate that these nanoparticles generally do not trigger elevated inflammatory cytokine release or induce acute toxicity in major organs, making them promising candidates for systemic administration.33 Nonetheless, their clinical translation faces significant challenges due to the histological heterogeneity and genomic complexity of many diseases, which demand greater delivery precision and individualized therapeutic strategies. Therefore, the development of more specific, high-affinity targeting peptides is urgently needed to improve the accuracy of delivery systems and broaden their therapeutic applications.
Mechanism of Action of Biomimetic Nanoparticles in Bone Regeneration
Bone is a mineralized connective tissue composed of osteoblasts, osteoclasts, and osteocytes. Its extracellular matrix contains both inorganic minerals and organic components, conferring high mechanical strength and toughness.182 Although bone possesses an intrinsic self-repair capacity through continuous remodeling, this ability is limited. When injury surpasses the regenerative threshold, structural and functional restoration becomes challenging.183 Without timely and effective treatment, bone injuries can reduce mechanical strength and progress to nonunion or defects, severely compromising skeletal support and motor function.184 Consequently, developing new strategies to enhance bone repair and regeneration is of great clinical significance. Traditionally, treatment has relied on autologous bone grafting, allogeneic transplantation, and artificial metallic or non-metallic implants.13 While these methods can partially restore structural integrity and function, long-term clinical outcomes remain suboptimal due to persistent challenges, including limited donor availability, immune rejection, risk of disease transmission, poor corrosion resistance, and susceptibility to infection.14,15
Biomimetic nanoparticles have attracted growing attention in bone regeneration due to their ability to replicate the structural and compositional features of the ECM in bone tissue. By reconstructing a microenvironment analogous to the natural ECM, they facilitate osteoblast adhesion, differentiation, and mineralization, thereby accelerating new bone formation.20,185 Moreover, their surfaces can be readily functionalized to incorporate drugs, growth factors, or bone-targeting peptides, enabling precise delivery and controlled release.32,33 As a result, biomimetic nanoparticles offer distinct advantages in improving bone repair efficiency, reducing adverse reactions, and advancing clinical translation, providing innovative strategies for the treatment of bone injuries and defects. This section highlights the mechanisms through which biomimetic nanoparticles contribute to bone regeneration, including the promotion of osteogenesis and mineralization, modulation of the bone immune microenvironment, and stimulation of angiogenesis.
Promotion of Osteogenesis and Mineralization
Osteogenesis and mineralization are central biological processes in bone tissue regeneration.186 This process relies on the directed differentiation of BMSCs into osteoblasts, which subsequently secrete and deposit a collagen-rich organic matrix. Hydroxyapatite crystals then nucleate on this scaffold, leading to progressive mineralization and hardening of bone tissue.187 Multiple signaling pathways regulate these events synergistically. For example, the Wnt/β-catenin pathway enhances osteoblast differentiation and proliferation,188 while the BMP/Smad pathway is pivotal in inducing osteogenic differentiation and mineralization.189 Additional pathways, such as MAPK and PI3K/AKT, further regulate cytoskeletal reorganization, proliferation, and differentiation.190 The ECM of bone not only provides structural support but also creates a dynamic regulatory microenvironment through surface-bound growth factors and biomechanical cues, thereby exerting critical control over osteogenesis and mineralization.191,192 Zhang et al developed a novel biomimetic nanoparticle, ALN@BMSCM@PLGA-TK-PEG-SS31, designed for immune evasion and prolonged circulation via BMSC membrane coating, while incorporating alendronate (ALN) for bone targeting. Under oxidative stress, the Reactive oxygen species (ROS)-sensitive PLGA-TK-PEG structure ruptures to release the mitochondrial-targeting peptide SS31, thereby alleviating oxidative stress. In vitro, this nanoplatform suppressed RANKL-induced osteoclastogenesis under hydrogen peroxide exposure and significantly promoted BMSC osteogenic differentiation and bone formation. In vivo, in an ovariectomized mouse model, ALN@BMSCM@PLGA-TK-PEG-SS31 not only mitigated oxidative stress but also increased bone mass without significant systemic toxicity (Figure 8).193 The BMP/Smad signaling pathway remains one of the most critical molecular regulators of bone formation. BMPs first bind to cell surface receptors, which sequentially activate Smad proteins. Phosphorylated Smads form complexes that translocate into the nucleus, where they regulate transcription of osteogenesis-related genes, thereby promoting BMSC differentiation into osteoblasts and accelerating mineralized matrix deposition.194 Chen et al demonstrated that miR-486-5p from rheumatoid arthritis (RA) fibroblast-like synovial cell exosomes promotes osteoblast proliferation and differentiation by activating the BMP/Smad pathway and inhibiting Tob1.195 Furthermore, miRNAs secreted by osteogenically differentiated BMSCs, including let-7a-5p, let-7c-5p, miR-328a-5p, and miR-31a-5p, modulate the competitive balance between Bmpr2/Acvr2b and Bmpr-induced Smad1/5/9 phosphorylation, thereby fine-tuning downstream Smad signaling to promote bone regeneration.196
 |
Figure 8 Schematic Diagram of the Synthesis Process of ALN@BMSCM@PLGA-TK-PEG-SS31 and Its Functional Effects Following Systemic Injection.193 |
Modulating the Immune Microenvironment
Bone regeneration and repair is a complex biological process in which dynamic changes within the immune microenvironment exert critical regulatory functions.197 During the early stages of repair, immune cells such as macrophages, dendritic cells, and T cells modulate the intensity and duration of inflammation by secreting cytokines and chemokines, thereby directly influencing the balance between osteogenesis and osteoclastogenesis.198,199 Aberrant immune responses may induce excessive inflammation or bone resorption, whereas appropriate immune modulation establishes a microenvironment favorable for osteogenesis.200 In recent years, biomimetic nanoparticles, owing to their structural and functional versatility, have demonstrated the ability to resolve inflammation74 and induce macrophage polarization toward the M2 phenotype201,202 by modulating immune cell activation states and cytokine secretion patterns. These immunomodulatory effects further support osteogenic differentiation and tissue regeneration.25 Yin et al designed a biomimetic anti-inflammatory nanocapsule (BANC), in which lipopolysaccharide-treated macrophage membranes encapsulated gold nanocages (AuNC). The membrane surface receptors neutralized excessive pro-inflammatory cytokines, suppressing pathological inflammation. Simultaneously, Resolvin D1 loaded in the nanocages was released on demand under near-infrared irradiation, driving macrophage polarization toward the M2 phenotype and thereby enhancing anti-inflammatory activity and tissue repair. In vivo femoral defect studies showed that BANC combined with boron-containing mesoporous bioactive glass scaffolds not only attenuated inflammation but also promoted sequential M2 polarization and bone regeneration.74 Wu et al proposed a strategy targeting Icam1⁺ macrophages to address non-healing infectious bone defects. They synthesized magnetic and ultrasonically responsive iron-doped barium titanate nanoparticles (BFTO), coated with γ3-peptide-modified engineered mesenchymal stem cell membranes (EMM) and simultaneously loaded with curcumin, forming BFTO-Cur@EMM nanoparticles. These nanoparticles disrupted bacterial biofilms under alternating magnetic fields and activated oxidative phosphorylation and osteoimmune responses in Icam1⁺ macrophages via low-intensity pulsed ultrasound. In vitro and in vivo experiments confirmed that BFTO-Cur@EMM nanoparticles promoted macrophage polarization toward a reparative phenotype by activating the JAK2–STAT3 pathway and inhibiting the MAPK–JNK pathway, thereby enhancing the secretion of pro-angiogenic and osteogenic cytokines and facilitating bone defect repair (Figure 9).203
 |
Figure 9 Bifunctional iron-doped strontium barium titanate nanoparticles (BFTO–Cur@EMM) were implanted into an infectious bone defect model. Through sequential magnetic guidance for anti-infection and ultrasound-targeted activation of Icam1+ macrophages, they induced bone regeneration after controlling the infection.203 |
Biomaterials implanted in the body often elicit immune responses primarily mediated by macrophage polarization.204 Persistent activation of the M1 phenotype results in chronic inflammation and impaired osseointegration,205 whereas enhancement of the M2 phenotype fosters an immune microenvironment favorable for bone regeneration.206 Thus, precise regulation of M1/M2 macrophage transition is essential for improving bone regeneration and osseointegration. Lei et al developed a tannic acid-modified sulfonated polyether ether ketone (SPEEK) implant functionalized with bone marrow mesenchymal stem cell-derived exosomes. This system enabled sustained Exo release, significantly improving material biocompatibility after cellular uptake. It further promoted M2 macrophage polarization and BMSC osteogenic differentiation by modulating the NF-κB signaling pathway. In vivo studies confirmed its capacity to achieve effective immune modulation, robust new bone formation, and enhanced osseointegration in rat models, highlighting the therapeutic potential of BMSC-derived Exo-functionalized implants for bone immune regulation and osteogenesis.201 Similarly, Schwann cell-derived exosomes were shown to promote M2 macrophage polarization, enhance endothelial cell tubulogenesis, and facilitate osteogenic differentiation of BMSCs.202
Promoting Neurogenesis and Angiogenesis
Bone tissue repair and regeneration depend not only on osteogenesis but also on the coordinated interactions between the vascular and nervous systems.207 Blood vessels supply oxygen and nutrients to bone tissue while removing metabolic waste, thereby maintaining homeostasis and supporting bone healing.208,209 Meanwhile, nerve fibers regulate immune responses, angiogenesis, and osteoblast activity through the secretion of neuropeptides and growth factors.14 Increasing evidence indicates that blood vessels and nerves are spatially co-localized and functionally interdependent, forming a “vascular–nerve unit” critical for bone regeneration and repair.6,210 Consequently, the development of biomimetic nanomaterials that promote both angiogenesis and neuroregeneration has emerged as an important strategy in bone tissue engineering. Jiang et al designed a novel miRNA-based delivery system by loading miR-21 onto metal–organic frameworks (MOFs) and encapsulating them within membranes derived from CXCR4-overexpressing bone marrow mesenchymal stem cells. Leveraging the CXCR4–SDF1 axis, this nanoplatform achieved targeted delivery to ischemic bone regions. Upon release, miR-21 silenced PTEN and activated the PI3K–AKT pathway, thereby promoting angiogenesis and vascular repair, improving blood supply, and ultimately enhancing osteogenesis. This strategy demonstrated significant therapeutic potential for glucocorticoid-induced osteonecrosis of the femoral head (GIONFH) through neovascularization.211 Yang et al revealed a pathological vicious cycle in steroid-induced osteonecrosis of the femoral head (SANFH), in which exosomes from damaged endothelial cells induced adipogenic differentiation of BMSCs. In turn, adipogenically differentiated BMSCs secreted pro-inflammatory factors, exacerbating endothelial injury, impairing angiogenesis, and hindering bone regeneration. To address this, they developed M-SiO2@Icariin@EBHM, a biomimetic nanomaterial consisting of icariin-loaded mesoporous silica coated with a hybrid membrane from endothelial cells and BMSCs. Systemic administration enabled targeted accumulation at lesion sites, where it scavenged glucocorticoid-induced reactive oxygen species, repaired vascular endothelium, and promoted osteogenic differentiation of BMSCs. This dual mechanism effectively disrupted the cycle of endothelial injury, inflammation amplification, and impaired bone repair, significantly enhancing angiogenesis and bone regeneration, thereby delaying SANFH progression (Figure 10).212
 |
Figure 10 Pathological Mechanisms and Therapeutic Strategies for Steroid-Associated Necrosis of the Femoral Head (SANFH): Hormones and other factors induce vascular endothelial cell injury, leading to increased reactive oxygen species (ROS) production and upregulation of mucosal addressin cell adhesion molecule-1 (MAdCAM-1). This, in turn, promotes the secretion of abnormal exosomes by damaged vascular endothelial cells (DVECs), driving the formation of a vicious cycle of vascular injury and lipid metabolism disorders. Targeting this mechanism, mesoporous silica (M-SiO2) loaded with icariin combined with engineered bone matrix (EBHM) forms M -SiO2@Icariin@EBHM, constructed from mesoporous silica (M-SiO2) loaded with icariin and engineered bone matrix (EBHM), exerts therapeutic effects by scavenging ROS, targeting DVECs and BMSCs, delaying SANFH progression, promoting osteogenic-related positive feedback loops, inhibiting metabolic dysfunction cycles, and regulating BMSC differentiation.212 |
Growing evidence indicates that exosomes are promising candidates for promoting both vascular and neural regeneration. The miRNAs, proteins, and lipids they transport regulate the function of vascular endothelial and neural cells, thereby exerting dual effects during bone repair.213,214 To address the challenge of inadequate vascularization in large bone defect repair, Zha et al developed an acellular tissue-engineering system based on engineered exosomes. Specifically, exosomes derived from ATDC5 cells were functionalized with the anchoring peptide CP05, immobilized on 3D-printed porous bone scaffolds, and loaded with the VEGF gene. In vitro studies showed that these functionalized exosomes not only acted as osteogenic matrices to promote mesenchymal stem cell differentiation but also significantly enhanced angiogenesis through controlled VEGF release. In vivo experiments further confirmed their efficacy in inducing large-volume vascularized bone regeneration, underscoring the potential of functionalized exosomes in acellular tissue engineering.215 In addition, exosomes can simultaneously promote vascular and neural regeneration. Su et al constructed a biomimetic periosteum PCL@PEI@exosome@aptamer (PPEA) by conjugating phosphatidylserine (PS)-targeting aptamers with Schwann cell–derived exosomes and immobilizing them on electrospun fiber surfaces, thereby enabling precise repair of periosteal nerve injuries. Both in vitro and in vivo experiments demonstrated that Schwann cell–derived exosomes promoted neurogenesis and angiogenesis through activation of the JNK/MAPK signaling pathway, while also exhibiting osteogenic potential by upregulating neuropeptide expression.216
Challenges and Future Prospects
Biological Safety and Toxicity
Despite their significant advantages in bone regeneration, the clinical translation of biomimetic nanoparticles still faces substantial challenges. First, rigorous and comprehensive biosafety evaluations are essential. Previous experience with nanomedicine development underscores that biosafety risks are often the primary reason for the termination of research or clinical projects. Although biomimetic nanoparticles have shown favorable biocompatibility, biodegradability, and low immunogenicity in vitro and in small animal studies, uncertainties remain regarding their long-term metabolism, the toxicity of degradation products, and potential immune responses in vivo. Therefore, systematic biosafety assessments of all nanoparticle components are indispensable. Personalized biomimetic nanoparticles—such as those derived from patient-specific cell membranes or exosomes from mesenchymal stem cells or immune cells—may offer a promising precision medicine strategy, reducing immunogenicity risks while enhancing therapeutic outcomes.
Challenges in Large-Scale Production
Scalability and reproducibility remain major barriers to clinical application due to limitations in current production processes. Present fabrication methods are largely restricted to the laboratory stage and are hampered by low yields, batch-to-batch variability, and complex preparation steps. For example, cell membrane-coated nanoparticles often suffer from high production costs and limited drug-loading capacity, while exosome-based systems face issues of heterogeneity and labor-intensive extraction. Emerging technologies, however, provide potential solutions. High-throughput microfluidic platforms enable precise microscale manipulation of liquid flow, allowing particle sorting or enrichment through acoustic forces, electric fields, or size-based separation. These platforms can shorten processing times, reduce sample consumption, and improve purity and reproducibility. Tangential flow filtration (TFF) has also been applied to exosome isolation, offering 3–5 times higher yields than ultracentrifugation while preserving structural integrity. Importantly, integrating microfluidics with TFF holds promise for achieving scalable, cost-effective production with robust quality control, thereby enhancing the clinical translation potential of biomimetic nanoparticles.
Clinical Translational and Regulatory Standards Challenges
At present, most research on biomimetic nanoparticles remains at the preclinical stage, and verification of their long-term safety and efficacy in humans requires costly large-scale clinical trials. Regulatory gaps further hinder this process. The absence of unified international standards not only delays approval procedures but also exacerbates difficulties arising from material heterogeneity and inconsistent characterization methods. Rigorous evaluation of key physicochemical parameters—such as particle size, dispersibility, and metabolic stability—is essential, yet presents far greater challenges than conventional drug assessment. Moreover, limited experience in industrial-scale production significantly restricts controllable mass manufacturing. Addressing these barriers demands close interdisciplinary collaboration to advance regulatory standards, optimize analytical and characterization techniques, and streamline manufacturing and quality control, thereby accelerating the clinical translation of biomimetic nanoparticles.
Despite these challenges, biomimetic nanoparticles hold tremendous potential in bone regeneration. Ongoing advancements in materials science, nanotechnology, and fabrication strategies are expected to enable the design of personalized nanoparticles with controllable structures, precise functionalization, and enhanced biosafety. Such progress will provide critical assurance for their successful clinical translation.
Conclusion
In recent years, biomimetic nanoparticles have emerged as an important area of research in bone regeneration because of their tunable structural and functional properties. This review summarizes the structural organization and functional characteristics of bone tissue and its microenvironment, and outlines the key stages of bone repair following injury. It further discusses major strategies for constructing biomimetic nanoparticles using diverse biological templates, including cell membranes, exosomes, proteins, and peptides. These design approaches may be used individually or in combination to integrate complementary functions and generate multifunctional platforms that better interface with the host bone microenvironment. Collectively, biomimetic nanoparticles show broad potential to enhance osteogenesis and mineralization, modulate the immune microenvironment, and support vascular and neural regeneration, thereby providing a materials-based framework for the development of bone repair strategies. Future progress in this field will depend on establishing verifiable links between key material parameters and biological outcomes, improving scalable manufacturing, batch-to-batch consistency, and standardized characterization, and building a more rigorous evidence base for long-term biosafety and clinical translation. Such advances will be essential for the development of more translatable and personalized therapies for bone repair.
Abbreviations
NPs, Nanoparticles; ECM, Extracellular matrix; NCPs, Non-collagenous proteins; MMPs, Matrix metalloproteinases; TIMPs, Tissue inhibitors of metalloproteinases; BMPs, Bone morphogenetic proteins; FGFs, Fibroblast growth factors; VEGF, Vascular endothelial growth factor; OPG, Osteoprotegerin; PDGF, Platelet-Derived Growth Factor; BMSCs, Bone marrow mesenchymal stem cells; CGRP, Calcitonin gene-related peptide; SP, Substance P; TGF-β, Transforming growth factor-β; RBCs, Red blood cells; SIRPα, Signal-regulatory protein alpha; ICA, Icariin; PLGA, Poly(lactic-co-glycolic acid); PNPs, Platelet membrane–cloaked nanoparticles; OVA, Ovalbumin; PDT, Photodynamic therapy; CEP, Cefpodoxime; MMs, Macrophage membranes; NLCs, Nanostructured lipid carriers; EVs, Exosomes; MVBs, Multivesicular bodies; PTX, Paclitaxel; DOX, Doxorubicin; TD, Thiazolidine-2,5-dione; BNPs, Bovine serum albumin nanoparticles; DBBM, Deproteinized bovine bone mineral; BSA, Bovine serum albumin; ALP, Alkaline phosphatase; Tf, Transferrin; TfR, Transferrin receptors; Tf-DOX, Transferrin–doxorubicin conjugates; E7, EPLQLKMC; OPN, Osteopontin; PDA, PLGA/dopamine; KGN, Kartogenin; Asp, Aspartic acid; Hap, Hydroxyapatite; Asp8, d-aspartic acid octapeptide; Asp-DPCN, Asp8-modified dendritic platinum–copper alloy nanoparticles; ALN, Alendronate; ROS, Reactive oxygen species; RA, Rheumatoid arthritis; BANC, Biomimetic anti-inflammatory nanocapsule; EMM, Mesenchymal stem cell membranes; SPEEK, Sulfonated polyether ether ketone; MOFs, Metal–organic frameworks; GIONFH, Glucocorticoid-induced osteonecrosis of the femoral head; SANFH, Steroid-induced osteonecrosis of the femoral head; PPEA, PCL@PEI@exosome@aptamer; PS, Phosphatidylserine; TFF, Tangential flow filtration.
Data Sharing Statement
No data was used for the research described in the article.
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China; Grant number: 82272489, 82203588; TaiShan Scholars Project Special Fund; Grant number: NO.tsqn202306396; Qingdao Science and Technology Benefiting the People Demonstration Special Project; Grant number: 24-1-8-smjk-3-nsh. The funding body played no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Sivakumar PM, Yetisgin AA, Sahin SB, Demir E, Cetinel S. Bone tissue engineering: anionic polysaccharides as promising scaffolds. Carbohydr Polym. 2022;283:119142. doi:10.1016/j.carbpol.2022.119142
2. Permyakova ES, Kiryukhantsev-Korneev PV, Gudz KY, et al. Comparison of different approaches to surface functionalization of biodegradable polycaprolactone scaffolds. Nanomaterials. 2019;9(12):1769. doi:10.3390/nano9121769
3. Zhou C, Hu X, Jing Y, et al. Bidirectional crosstalk between the bone extracellular matrix and lysosomes in bone remodeling and osteoporosis. Front Endocrinol. 2025;16:1698404. doi:10.3389/fendo.2025.1698404
4. Wang L, You X, Zhang L, Zhang C, Zou W. Mechanical regulation of bone remodeling. Bone Res. 2022;10(1):16. doi:10.1038/s41413-022-00190-4
5. Mohanakrishnan V, Sivaraj KK, Jeong HW, et al. Specialized post-arterial capillaries facilitate adult bone remodelling. Nat Cell Biol. 2024;26(12):2020–32. doi:10.1038/s41556-024-01545-1
6. Zhang M, Lin R, Wang X, et al. 3D printing of Haversian bone-mimicking scaffolds for multicellular delivery in bone regeneration. Sci Adv. 2020;6(12):eaaz6725. doi:10.1126/sciadv.aaz6725
7. Jiao X, Sun X, Li W, et al. 3D-printed β-tricalcium phosphate scaffolds promote osteogenic differentiation of bone marrow-deprived mesenchymal stem cells in an N6-methyladenosine-dependent manner. Int J Bioprint. 2022;8(2):544. doi:10.18063/ijb.v8i2.544
8. Tang Q, Li X, Lai C, et al. Fabrication of a hydroxyapatite-PDMS microfluidic chip for bone-related cell culture and drug screening. Bioact Mater. 2021;6(1):169–178. doi:10.1016/j.bioactmat.2020.07.016
9. Zheng W, Meng Z, Zhu Z, et al. Metal-organic framework-based nanomaterials for regulation of the osteogenic microenvironment. Small. 2024;20(29):e2310622. doi:10.1002/smll.202310622
10. Wang Q, Mu M, Yu W, et al. Engineering polyphenol-based osteogenic system for bone and cartilage repair: transplantation, tissue engineering, and organoid. J Adv Res. 2025;(S2090–1232(25)00476–X). doi:10.1016/j.jare.2025.06.067
11. Li H, Fan R, Zou B, Yan J, Shi Q, Guo G. Roles of MXenes in biomedical applications: recent developments and prospects. J Nanobiotechnology. 2023;21(1):73. doi:10.1186/s12951-023-01809-2
12. Xu G, Chen J, Xiao Z, Xu J, Zhang L, Tang P. Bioaugmented osteosynthesis: precise monitoring and intervention of the bone healing microenvironment. Bone Res. 2025;13(1):95. doi:10.1038/s41413-025-00466-5
13. Ying C, Wang R, Wang Z, et al. BMSC-exosomes carry mutant HIF-1α for improving angiogenesis and osteogenesis in critical-sized calvarial defects. Front Bioeng Biotechnol. 2020;8:565561. doi:10.3389/fbioe.2020.565561
14. Ma YX, Jiao K, Wan QQ, et al. Silicified collagen scaffold induces semaphorin 3A secretion by sensory nerves to improve in-situ bone regeneration. Bioact Mater. 2022;9:475–490. doi:10.1016/j.bioactmat.2021.07.016
15. Jia B, Yang H, Zhang Z, et al. Biodegradable Zn-Sr alloy for bone regeneration in rat femoral condyle defect model: in vitro and in vivo studies. Bioact Mater. 2021;6(6):1588–1604. doi:10.1016/j.bioactmat.2020.11.007
16. Ji C, Qiu M, Ruan H, et al. Transcriptome analysis revealed the symbiosis niche of 3D scaffolds to accelerate bone defect healing. Adv Sci. 2022;9(8):e2105194. doi:10.1002/advs.202105194
17. Kang Y, Li S. Nanomaterials: breaking through the bottleneck of tumor immunotherapy. Int J Biol Macromol. 2023;230:123159. doi:10.1016/j.ijbiomac.2023.123159
18. Jin GZ. Current nanoparticle-based technologies for osteoarthritis therapy. Nanomaterials. 2020;10(12):2368. doi:10.3390/nano10122368
19. Yu F, Lian R, Liu L, et al. Biomimetic hydroxyapatite nanorods promote bone regeneration via accelerating osteogenesis of BMSCs through T cell-derived IL-22. ACS Nano. 2022;16(1):755–770. doi:10.1021/acsnano.1c08281
20. Guo Y, Wang M, Ge J, et al. Bioactive biodegradable polycitrate nanoclusters enhances the myoblast differentiation and in vivo skeletal muscle regeneration via p38 MAPK signaling pathway. Bioact Mater. 2020;5(3):486–495. doi:10.1016/j.bioactmat.2020.04.004
21. Wei R, Jiang G, Lv M, et al. TMTP1-modified indocyanine green-loaded polymeric micelles for targeted imaging of cervical cancer and metastasis sentinel lymph node in vivo. Theranostics. 2019;9(24):7325–7344. doi:10.7150/thno.35346
22. Sun L, Liu H, Ye Y, et al. Smart nanoparticles for cancer therapy. Signal Transduct Target Ther. 2023;8(1):418. doi:10.1038/s41392-023-01642-x
23. Wang S, Duan Y, Zhang Q, et al. Drug targeting via platelet membrane-coated nanoparticles. Small Struct. 2020;1(1):2000018. doi:10.1002/sstr.202000018
24. Chou WC, Lin Z. Impact of protein coronas on nanoparticle interactions with tissues and targeted delivery. Curr Opin Biotechnol. 2024;85:103046. doi:10.1016/j.copbio.2023.103046
25. Luo Z, Lu Y, Shi Y, et al. Neutrophil hitchhiking for drug delivery to the bone marrow. Nat Nanotechnol. 2023;18(6):647–656. doi:10.1038/s41565-023-01374-7
26. Chi S, Zhang L, Cheng H, et al. Biomimetic nanocomposites camouflaged with hybrid cell membranes for accurate therapy of early-stage glioma. Angew Chem Int Ed Engl. 2023;62(29):e202304419. doi:10.1002/anie.202304419
27. Wang Y, Zhang K, Li T, et al. Macrophage membrane functionalized biomimetic nanoparticles for targeted anti-atherosclerosis applications. Theranostics. 2021;11(1):164–180. doi:10.7150/thno.47841
28. Ji Y, Zhang Z, Hou W, et al. Enhanced antitumor effect of icariin nanoparticles coated with iRGD functionalized erythrocyte membrane. Eur J Pharmacol. 2022;931:175225. doi:10.1016/j.ejphar.2022.175225
29. Lu C, Zheng J, Ding Y, et al. Cepharanthine loaded nanoparticles coated with macrophage membranes for lung inflammation therapy. Drug Deliv. 2021;28(1):2582–2593. doi:10.1080/10717544.2021.2009936
30. Yuyama K, Sun H, Sakai S, et al. Decreased amyloid-β pathologies by intracerebral loading of glycosphingolipid-enriched exosomes in Alzheimer model mice. J Biol Chem. 2014;289(35):24488–24498. doi:10.1074/jbc.M114.577213
31. Liu W, Huang J, Chen F, et al. MSC-derived small extracellular vesicles overexpressing miR-20a promoted the osteointegration of porous titanium alloy by enhancing osteogenesis via targeting BAMBI. Stem Cell Res Ther. 2021;12(1):348. doi:10.1186/s13287-021-02303-y
32. Hanske C, Tebbe M, Kuttner C, et al. Strongly coupled plasmonic modes on macroscopic areas via template-assisted colloidal self-assembly. Nano Lett. 2014;14(12):6863–6871. doi:10.1021/nl502776s
33. Cui K, Ren F, Yu J, Pan H. Bioinspired nanomedicines for the management of osteosarcoma: recent progress and perspectives. Mater Today Bio. 2025;32:101607. doi:10.1016/j.mtbio.2025.101607
34. Zong L, Wang Q, Sun H, et al. Intra-articular injection of PLGA/polydopamine core-shell nanoparticle attenuates osteoarthritis progression. ACS Appl Mater Interfaces. 2024;16(17):21450–21462. doi:10.1021/acsami.3c18464
35. Diomede F, D’Aurora M, Gugliandolo A, et al. A novel role in skeletal segment regeneration of extracellular vesicles released from periodontal-ligament stem cells. Int J Nanomed. 2018;13:3805–3825. doi:10.2147/IJN.S162836
36. Alsulaiman M, Bais MV, Trackman PC. Lysyl oxidase propeptide stimulates osteoblast and osteoclast differentiation and enhances PC3 and DU145 prostate cancer cell effects on bone in vivo. J Cell Commun Signal. 2016;10(1):17–31. doi:10.1007/s12079-015-0311-9
37. Rott J, Toepfer ET, Bartosova M, et al. Nutritional calcium supply dependent calcium balance, bone calcification and calcium isotope ratios in rats. Int J Mol Sci. 2022;23(14):7796. doi:10.3390/ijms23147796
38. Gajjeraman S, He G, Narayanan K, George A. Biological assemblies provide novel templates for the synthesis of hierarchical structures and facilitate cell adhesion. Adv Funct Mater. 2008;18(24):3972–3980. doi:10.1002/adfm.200801215
39. Huang YM, Huang CC, Tsai PI, et al. Three-dimensional printed porous titanium screw with bioactive surface modification for bone-tendon healing: a rabbit animal model. Int J Mol Sci. 2020;21(10):3628. doi:10.3390/ijms21103628
40. Wang D, Zhang P, Mei X, Chen Z. Repair calvarial defect of osteoporotic rats by berberine functionalized porous calcium phosphate scaffold. Regen Biomater. 2021;8(3):rbab022. doi:10.1093/rb/rbab022
41. Klíma K, Ulmann D, Bartoš M, et al. A complex evaluation of the in-vivo biocompatibility and degradation of an extruded ZnMgSr absorbable alloy implanted into rabbit bones for 360 days. Int J Mol Sci. 2021;22(24):13444. doi:10.3390/ijms222413444
42. Kim SJ, Hwang YH, Mun SK, et al. Protective Effects of 2,3,5,4’-Tetrahydroxystilbene-2-O-β-d-glucoside on ovariectomy induced osteoporosis mouse model. Int J Mol Sci. 2018;19(9):2554. doi:10.3390/ijms19092554
43. Huang Q, Liu Y, Ouyang Z, Feng Q. Comparing the regeneration potential between PLLA/Aragonite and PLLA/Vaterite pearl composite scaffolds in rabbit radius segmental bone defects. Bioact Mater. 2020;5(4):980–989. doi:10.1016/j.bioactmat.2020.06.018
44. Liu G, Wang X, Zhou X, et al. Modulating the cobalt dose range to manipulate multisystem cooperation in bone environment: a strategy to resolve the controversies about cobalt use for orthopedic applications. Theranostics. 2020;10(3):1074–1089. doi:10.7150/thno.37931
45. Choi K, Kuhn JL, Ciarelli MJ, Goldstein SA. The elastic moduli of human subchondral, trabecular, and cortical bone tissue and the size-dependency of cortical bone modulus. J Biomech. 1990;23(11):1103–1113. doi:10.1016/0021-9290(90)90003-l
46. Xiong Y, He T, Liu WV, et al. Quantitative assessment of lumbar spine bone marrow in patients with different severity of CKD by IDEAL-IQ magnetic resonance sequence. Front Endocrinol. 2022;13:980576. doi:10.3389/fendo.2022.980576
47. Bose S, Banerjee D, Vu AA. Ginger and garlic extracts enhance osteogenesis in 3D printed calcium phosphate bone scaffolds with bimodal pore distribution. ACS Appl Mater Interfaces. 2022;14(11):12964–12975. doi:10.1021/acsami.1c19617
48. Piard C, Baker H, Kamalitdinov T, Fisher J. Bioprinted osteon-like scaffolds enhance in vivo neovascularization. Biofabrication. 2019;11(2):025013. doi:10.1088/1758-5090/ab078a
49. Zhang Q, Lin S, Zhang T, et al. Curved microstructures promote osteogenesis of mesenchymal stem cells via the RhoA/ROCK pathway. Cell Prolif. 2017;50(4):e12356. doi:10.1111/cpr.12356
50. Masters EA, Salminen AT, Begolo S, et al. An in vitro platform for elucidating the molecular genetics of S. aureus invasion of the osteocyte lacuno-canalicular network during chronic osteomyelitis. Nanomedicine. 2019;21:102039. doi:10.1016/j.nano.2019.102039
51. Barati D, Karaman O, Moeinzadeh S, Kader S, Jabbari E. Material and regenerative properties of an osteon-mimetic cortical bone-like scaffold. Regen Biomater. 2019;6(2):89–98. doi:10.1093/rb/rbz008
52. Koushik TM, Miller CM, Antunes E. Bone tissue engineering scaffolds: function of multi-material hierarchically structured scaffolds. Adv Healthc Mater. 2023;12(9):e2202766. doi:10.1002/adhm.202202766
53. Varun D, Srinivasan GR, Tsai YH, et al. A robust vitronectin-derived peptide for the scalable long-term expansion and neuronal differentiation of human pluripotent stem cell (hPSC)-derived neural progenitor cells (hNPCs). Acta Biomater. 2017;48:120–130. doi:10.1016/j.actbio.2016.10.037
54. Liu X, Qu X, Nie T, et al. The beneficial effects of bisphosphonate-enoxacin on cortical bone mass and strength in ovariectomized rats. Front Pharmacol. 2017;8:355. doi:10.3389/fphar.2017.00355
55. Sadeghzadeh H, Mehdipour A, Dianat-Moghadam H, et al. PCL/Col I-based magnetic nanocomposite scaffold provides an osteoinductive environment for ADSCs in osteogenic cues-free media conditions. Stem Cell Res Ther. 2022;13(1):143. doi:10.1186/s13287-022-02816-0
56. Limgala RP, Goker-Alpan O. Effect of substrate reduction therapy in comparison to enzyme replacement therapy on immune aspects and bone involvement in Gaucher disease. Biomolecules. 2020;10(4):526. doi:10.3390/biom10040526
57. Aslankoohi N, Lin S, Mequanint K. Bioactive fluorescent hybrid microparticles as a stand-alone osteogenic differentiation inducer. Mater Today Bio. 2022;13:100187. doi:10.1016/j.mtbio.2021.100187
58. Spoerke ED, Anthony SG, Stupp SI. Enzyme directed templating of artificial bone mineral. Adv Mater. 2009;21(4):425–430. doi:10.1002/adma.200802242
59. Jeon J, Lee MS, Yang HS. Differentiated osteoblasts derived decellularized extracellular matrix to promote osteogenic differentiation. Biomater Res. 2018;22:4. doi:10.1186/s40824-018-0115-0
60. Karbowniczek JE, Kaniuk Ł, Berniak K, Gruszczyński A, Stachewicz U. Enhanced cells anchoring to electrospun hybrid scaffolds with PHBV and HA particles for bone tissue regeneration. Front Bioeng Biotechnol. 2021;9:632029. doi:10.3389/fbioe.2021.632029
61. Gharibeh L, Komati H, Bossé Y, et al. GATA6 regulates aortic valve remodeling, and its haploinsufficiency leads to right-left type bicuspid aortic valve. Circulation. 2018;138(10):1025–1038. doi:10.1161/CIRCULATIONAHA.117.029506
62. Yao J, Guihard PJ, Blazquez-Medela AM, et al. Matrix Gla protein regulates differentiation of endothelial cells derived from mouse embryonic stem cells. Angiogenesis. 2016;19(1):1–7. doi:10.1007/s10456-015-9484-3
63. Gori JL, Butler JM, Chan YY, et al. Vascular niche promotes hematopoietic multipotent progenitor formation from pluripotent stem cells. J Clin Invest. 2015;125(3):1243–1254. doi:10.1172/JCI79328
64. Arron JR, Choi Y. Bone versus immune system. Nature. 2000;408(6812):535–536. doi:10.1038/35046196
65. Walsh MC, Kim N, Kadono Y, et al. Osteoimmunology: interplay between the immune system and bone metabolism. Ann Rev Immunol. 2006;24(24):33–63. doi:10.1146/annurev.immunol.24.021605.090646
66. Pinho S, Frenette PS. Haematopoietic stem cell activity and interactions with the niche. Nat Rev Mol Cell Biol. 2019;20(5):303–320. doi:10.1038/s41580-019-0103-9
67. Owen-Woods C, Kusumbe A. Fundamentals of bone vasculature: specialization, interactions and functions. Semin Cell Dev Biol. 2022;123:36–47. doi:10.1016/j.semcdb.2021.06.025
68. Thai J, Fuller-Jackson JP, Ivanusic JJ. Using tissue clearing and light sheet fluorescence microscopy for the three-dimensional analysis of sensory and sympathetic nerve endings that innervate bone and dental tissue of mice. J Comp Neurol. 2024;532(1):e25582. doi:10.1002/cne.25582
69. Biswas L, Chen J, De Angelis J, et al. Lymphatic vessels in bone support regeneration after injury. Cell. 2023;186(2):382–397.e24. doi:10.1016/j.cell.2022.12.031
70. Ross EA, Flores-Langarica A, Bobat S, et al. Resolving Salmonella infection reveals dynamic and persisting changes in murine bone marrow progenitor cell phenotype and function. Eur J Immunol. 2014;44(8):2318–2330. doi:10.1002/eji.201344350
71. Jiang Y, Zhang Y, Chen W, et al. Achyranthes bidentata extract exerts osteoprotective effects on steroid-induced osteonecrosis of the femoral head in rats by regulating RANKL/RANK/OPG signaling. J Transl Med. 2014;12:334. doi:10.1186/s12967-014-0334-7
72. Bonnet N, Bourgoin L, Biver E, Douni E, Ferrari S. RANKL inhibition improves muscle strength and insulin sensitivity and restores bone mass. J Clin Invest. 2019;129(8):3214–3223. doi:10.1172/JCI125915
73. Tan T, Huang Q, Chu W, et al. Delivery of germacrone (GER) using macrophages-targeted polymeric nanoparticles and its application in rheumatoid arthritis. Drug Deliv. 2022;29(1):692–701. doi:10.1080/10717544.2022.2044936
74. Yin C, Zhao Q, Li W, et al. Biomimetic anti-inflammatory nano-capsule serves as a cytokine blocker and M2 polarization inducer for bone tissue repair. Acta Biomater. 2020;102:416–426. doi:10.1016/j.actbio.2019.11.025
75. Liu W, Dan X, Lu WW, et al. Spatial distribution of biomaterial microenvironment pH and its modulatory effect on osteoclasts at the early stage of bone defect regeneration. ACS Appl Mater Interfaces. 2019;11(9):9557–9572. doi:10.1021/acsami.8b20580
76. Chen Z, Chen L, Liu R, et al. The osteoimmunomodulatory property of a barrier collagen membrane and its manipulation via coating nanometer-sized bioactive glass to improve guided bone regeneration. Biomater Sci. 2018;6(5):1007–1019. doi:10.1039/c7bm00869d
77. Chen M, Li Y, Huang X, et al. Skeleton-vasculature chain reaction: a novel insight into the mystery of homeostasis. Bone Res. 2021;9(1):21. doi:10.1038/s41413-021-00138-0
78. Gu J, Zhang Q, Geng M, et al. Construction of nanofibrous scaffolds with interconnected perfusable microchannel networks for engineering of vascularized bone tissue. Bioact Mater. 2021;6(10):3254–3268. doi:10.1016/j.bioactmat.2021.02.033
79. Liu P, Gao Y, Luo P, et al. Glucocorticoid-induced expansion of classical monocytes contributes to bone loss. Exp Mol Med. 2022;54(6):765–776. doi:10.1038/s12276-022-00764-6
80. Lu J, Hu D, Ma C, et al. Modified Qing’ e Pills exerts anti-osteoporosis effects and prevents bone loss by enhancing type H blood vessel formation. Front Endocrinol. 2022;13:998971. doi:10.3389/fendo.2022.998971
81. Ryu J, Kim HJ, Chang EJ, Huang H, Banno Y, Kim HH. Sphingosine 1-phosphate as a regulator of osteoclast differentiation and osteoclast-osteoblast coupling. EMBO J. 2006;25(24):5840–5851. doi:10.1038/sj.emboj.7601430
82. Wu M, Zhang Y, Wu P, et al. Mussel-inspired multifunctional surface through promoting osteogenesis and inhibiting osteoclastogenesis to facilitate bone regeneration. NPJ Regen Med. 2022;7(1):29. doi:10.1038/s41536-022-00224-9
83. Sayilekshmy M, Hansen RB, Delaissé JM, Rolighed L, Andersen TL, Heegaard AM. Innervation is higher above bone remodeling surfaces and in cortical pores in human bone: lessons from patients with primary hyperparathyroidism. Sci Rep. 2019;9(1):5361. doi:10.1038/s41598-019-41779-w
84. Hohmann EL, Elde RP, Rysavy JA, Einzig S, Gebhard RL. Innervation of periosteum and bone by sympathetic vasoactive intestinal peptide-containing nerve fibers. Science. 1986;232(4752):868–871. doi:10.1126/science.3518059
85. Cooper RR. Nerves in cortical bone. Science. 1968;160(3825):327–328. doi:10.1126/science.160.3825.327
86. Chen H, Hu B, Lv X, et al. Prostaglandin E2 mediates sensory nerve regulation of bone homeostasis. Nat Commun. 2019;10(1):181. doi:10.1038/s41467-018-08097-7
87. Iwaniec UT, Philbrick KA, Wong CP, et al. Room temperature housing results in premature cancellous bone loss in growing female mice: implications for the mouse as a preclinical model for age-related bone loss. Osteoporos Int. 2016;27(10):3091–3101. doi:10.1007/s00198-016-3634-3
88. Cao H, Kou X, Yang R, et al. Force-induced Adrb2 in periodontal ligament cells promotes tooth movement. J Dent Res. 2014;93(11):1163–1169. doi:10.1177/0022034514551769
89. Zhao X, Yao M, Wang Y, et al. Neuroregulation during bone formation and regeneration: mechanisms and strategies. ACS Appl Mater Interfaces. 2025;17(5):7223–7250. doi:10.1021/acsami.4c16786
90. Stritt S, Koltowska K, Mäkinen T. Homeostatic maintenance of the lymphatic vasculature. Trends Mol Med. 2021;27(10):955–970. doi:10.1016/j.molmed.2021.07.003
91. Hu Z, Zhao X, Wu Z, et al. Lymphatic vessel: origin, heterogeneity, biological functions, and therapeutic targets. Signal Transduct Target Ther. 2024;9(1):9. doi:10.1038/s41392-023-01723-x
92. Ma W, Gil HJ, Liu X, et al. Mitochondrial respiration controls the Prox1-Vegfr3 feedback loop during lymphatic endothelial cell fate specification and maintenance. Sci Adv. 2021;7(18):eabe7359. doi:10.1126/sciadv.abe7359
93. Zhao Z, Du Y, Yan K, Zhang L, Guo Q. Exercise and osteoimmunology in bone remodeling. FASEB J. 2024;38(7):e23554. doi:10.1096/fj.202301508RRR
94. Rampichová M, Buzgo M, Míčková A, et al. Platelet-functionalized three-dimensional poly-ε-caprolactone fibrous scaffold prepared using centrifugal spinning for delivery of growth factors. Int J Nanomed. 2017;12:347–361. doi:10.2147/IJN.S120206
95. Bahney CS, Zondervan RL, Allison P, et al. Cellular biology of fracture healing. J Orthop Res. 2019;37(1):35–50. doi:10.1002/jor.24170
96. Hak DJ. The biology of fracture healing in osteoporosis and in the presence of anti-osteoporotic drugs. Injury. 2018;49(8):1461–1465. doi:10.1016/j.injury.2018.04.016
97. Yan Y, Cheng X, Yang RH, et al. Exposure to excess phenobarbital negatively influences the osteogenesis of chick embryos. Front Pharmacol. 2016;7:349. doi:10.3389/fphar.2016.00349
98. Panahipour L, Kargarpour Z, Laggner M, Mildner M, Ankersmit HJ, Gruber R. TGF-β in the secretome of irradiated peripheral blood mononuclear cells supports in vitro osteoclastogenesis. Int J Mol Sci. 2020;21(22):8569. doi:10.3390/ijms21228569
99. Zhu Y, Jia Y, Wang Y, Xu J, Chai Y. Impaired bone regenerative effect of exosomes derived from bone marrow mesenchymal stem cells in type 1 diabetes. Stem Cells Transl Med. 2019;8(6):593–605. doi:10.1002/sctm.18-0199
100. Zhang W, Wang N, Yang M, et al. Periosteum and development of the tissue-engineered periosteum for guided bone regeneration. J Orthop Translat. 2022;33:41–54. doi:10.1016/j.jot.2022.01.002
101. Claes L, Recknagel S, Ignatius A. Fracture healing under healthy and inflammatory conditions. Nat Rev Rheumatol. 2012;8(3):133–143. doi:10.1038/nrrheum.2012.1
102. Loi F, Córdova LA, Pajarinen J, hua LT, Yao Z, Goodman SB. Inflammation, fracture and bone repair. Bone. 2016;86:119–130. doi:10.1016/j.bone.2016.02.020
103. Ma Y, Zhou Y, Wu F, Ji W, Zhang J, Wang X. The bidirectional interactions between inflammation and coagulation in fracture hematoma. Tissue Eng Part B Rev. 2019;25(1):46–54. doi:10.1089/ten.TEB.2018.0157
104. Short AR, Koralla D, Deshmukh A, et al. Hydrogels that allow and facilitate bone repair, remodeling, and regeneration. J Mater Chem B. 2015;3(40):7818–7830. doi:10.1039/C5TB01043H
105. Kim S, Garcia A, Jackson SP, Kunapuli SP. Insulin-like growth factor-1 regulates platelet activation through PI3-Kalpha isoform. Blood. 2007;110(13):4206–4213. doi:10.1182/blood-2007-03-080804
106. Li Y, Hoffman MD, Benoit DSW. Matrix metalloproteinase (MMP)-degradable tissue engineered periosteum coordinates allograft healing via early stage recruitment and support of host neurovasculature. Biomaterials. 2021;268:120535. doi:10.1016/j.biomaterials.2020.120535
107. Frisch J, Venkatesan JK, Rey-Rico A, Schmitt G, Madry H, Cucchiarini M. Influence of insulin-like growth factor I overexpression via recombinant adeno-associated vector gene transfer upon the biological activities and differentiation potential of human bone marrow-derived mesenchymal stem cells. Stem Cell Res Ther. 2014;5(4):103. doi:10.1186/scrt491
108. Yan CP, Wang XK, Jiang K, et al. β-Ecdysterone enhanced bone regeneration through the BMP-2/SMAD/RUNX2/Osterix signaling pathway. Front Cell Dev Biol. 2022;10:883228. doi:10.3389/fcell.2022.883228
109. Spiller KL, Anfang RR, Spiller KJ, et al. The role of macrophage phenotype in vascularization of tissue engineering scaffolds. Biomaterials. 2014;35(15):4477–4488. doi:10.1016/j.biomaterials.2014.02.012
110. Einhorn TA. The science of fracture healing. J Orthop Trauma. 2005;19(10 Suppl):S4–6. doi:10.1097/00005131-200511101-00002
111. Wang T, Zhang X, Bikle DD. Osteogenic differentiation of periosteal cells during fracture healing. J Cell Physiol. 2017;232(5):913–921. doi:10.1002/jcp.25641
112. Wang Y, Zhang H, Hu Y, Jing Y, Geng Z, Su J. Bone repair biomaterials: a perspective from immunomodulation. Adv Funct Mater. 2022;32(51):2208639. doi:10.1002/adfm.202208639
113. Chang B, Osteon LX. Structure, turnover, and regeneration. Tissue Eng Part B Rev. 2022;28(2):261–278. doi:10.1089/ten.TEB.2020.0322
114. Saeki C, Saito M, Oikawa T, et al. Effects of denosumab treatment in chronic liver disease patients with osteoporosis. World J Gastroenterol. 2020;26(33):4960–4971. doi:10.3748/wjg.v26.i33.4960
115. Schindeler A, McDonald MM, Bokko P, Little DG. Bone remodeling during fracture repair: the cellular picture. Semin Cell Dev Biol. 2008;19(5):459–466. doi:10.1016/j.semcdb.2008.07.004
116. Qiao K, Xu L, Tang J, et al. The advances in nanomedicine for bone and cartilage repair. J Nanobiotechnology. 2022;20(1):141. doi:10.1186/s12951-022-01342-8
117. Meng X, Wang J, Zhou J, et al. Tumor cell membrane-based peptide delivery system targeting the tumor microenvironment for cancer immunotherapy and diagnosis. Acta Biomater. 2021;127:266–275. doi:10.1016/j.actbio.2021.03.056
118. Elsharkasy OM, Nordin JZ, Hagey DW, et al. Extracellular vesicles as drug delivery systems: why and how? Adv Drug Deliv Rev. 2020;159:332–343. doi:10.1016/j.addr.2020.04.004
119. Jhan YY, Prasca-Chamorro D, Palou Zuniga G, et al. Engineered extracellular vesicles with synthetic lipids via membrane fusion to establish efficient gene delivery. Int J Pharm. 2020;573:118802. doi:10.1016/j.ijpharm.2019.118802
120. Martínez-López AL, Pangua C, Reboredo C, Campión R, Morales-Gracia J, Irache JM. Protein-based nanoparticles for drug delivery purposes. Int J Pharm. 2020;581:119289. doi:10.1016/j.ijpharm.2020.119289
121. Pirhaghi M, Mamashli F, Moosavi-Movahedi F, et al. Cell-penetrating peptides: promising therapeutics and drug-delivery systems for neurodegenerative diseases. Mol Pharm. 2024;21(5):2097–2117. doi:10.1021/acs.molpharmaceut.3c01167
122. Hu CMJ, Fang RH, Wang KC, et al. Nanoparticle biointerfacing by platelet membrane cloaking. Nature. 2015;526(7571):118–121. doi:10.1038/nature15373
123. Piffoux M, Volatron J, Cherukula K, et al. Engineering and loading therapeutic extracellular vesicles for clinical translation: a data reporting frame for comparability. Adv Drug Deliv Rev. 2021;178:113972. doi:10.1016/j.addr.2021.113972
124. Nguyen B, Tolia NH. Protein-based antigen presentation platforms for nanoparticle vaccines. NPJ Vaccines. 2021;6(1):70. doi:10.1038/s41541-021-00330-7
125. Lagziel-Simis S, Cohen-Hadar N, Moscovich-Dagan H, Wine Y, Freeman A. Protein-mediated nanoscale biotemplating. Curr Opin Biotechnol. 2006;17(6):569–573. doi:10.1016/j.copbio.2006.10.005
126. Lee HJ, Ehsan M, Zhang X, et al. Development of 1,3-acetonedicarboxylate-derived glucoside amphiphiles (ACAs) for membrane protein study. Chem Sci. 2022;13(19):5750–5759. doi:10.1039/d2sc00539e
127. Wu Y, Wan S, Yang S, et al. Macrophage cell membrane-based nanoparticles: a new promising biomimetic platform for targeted delivery and treatment. J Nanobiotechnology. 2022;20(1):542. doi:10.1186/s12951-022-01746-6
128. Kuhn V, Diederich L, Keller TCS, et al. Red blood cell function and dysfunction: redox regulation, nitric oxide metabolism, anemia. Antioxid Redox Signal. 2017;26(13):718–742. doi:10.1089/ars.2016.6954
129. Zheng BD, Xiao MT. Red blood cell membrane nanoparticles for tumor phototherapy. Colloids Surf B Biointerfaces. 2022;220:112895. doi:10.1016/j.colsurfb.2022.112895
130. Kundu M, Lindsten T, Yang CY, et al. Ulk1 plays a critical role in the autophagic clearance of mitochondria and ribosomes during reticulocyte maturation. Blood. 2008;112(4):1493–1502. doi:10.1182/blood-2008-02-137398
131. Li Q, Lin B, Li Y, Lu N. Erythrocyte-camouflaged mesoporous titanium dioxide nanoplatform for an ultrasound-mediated sequential therapies of breast cancer. Int J Nanomed. 2021;16:3875–3887. doi:10.2147/IJN.S301855
132. Thon JN, Italiano JE. Platelets: production, morphology and ultrastructure. Handb Exp Pharmacol. 2012;210:3–22. doi:10.1007/978-3-642-29423-5_1
133. Zhu X, Li J, Deng S, et al. Genome-wide analysis of DNA methylation and cigarette smoking in a Chinese population. Environ Health Perspect. 2016;124(7):966–973. doi:10.1289/ehp.1509834
134. Lannan KL, Spinelli SL, Blumberg N, Phipps RP. Maresin 1 induces a novel pro-resolving phenotype in human platelets. J Thromb Haemost. 2017;15(4):802–813. doi:10.1111/jth.13620
135. Tang J, Su T, Huang K, et al. Targeted repair of heart injury by stem cells fused with platelet nanovesicles. Nat Biomed Eng. 2018;2:17–26. doi:10.1038/s41551-017-0182-x
136. Wang H, Wang K, He L, Liu Y, Dong H, Li Y. Engineering antigen as photosensitiser nanocarrier to facilitate ROS triggered immune cascade for photodynamic immunotherapy. Biomaterials. 2020;244:119964. doi:10.1016/j.biomaterials.2020.119964
137. Vasan N, Baselga J, Hyman DM. A view on drug resistance in cancer. Nature. 2019;575(7782):299–309. doi:10.1038/s41586-019-1730-1
138. Russo M, Crisafulli G, Sogari A, et al. Adaptive mutability of colorectal cancers in response to targeted therapies. Science. 2019;366(6472):1473–1480. doi:10.1126/science.aav4474
139. Shi Y, Lin G, Zheng H, et al. Biomimetic nanoparticles blocking autophagy for enhanced chemotherapy and metastasis inhibition via reversing focal adhesion disassembly. J Nanobiotechnology. 2021;19(1):447. doi:10.1186/s12951-021-01189-5
140. Cao X, Tan T, Zhu D, et al. Paclitaxel-loaded macrophage membrane camouflaged albumin nanoparticles for targeted cancer therapy. Int J Nanomed. 2020;15:1915–1928. doi:10.2147/IJN.S244849
141. Sun L, Ouyang J, Zeng Z, et al. Targeted and activatable nanosystem for fluorescent and optoacoustic imaging of immune-mediated inflammatory diseases and therapy via inhibiting NF-κB/NLRP3 pathways. Bioact Mater. 2022;10:79–92. doi:10.1016/j.bioactmat.2021.08.010
142. Li Z, Zhang X, Liu C, et al. Macrophage-biomimetic nanoparticles ameliorate ulcerative colitis through reducing inflammatory factors expression. J Innate Immun. 2022;14(4):380–392. doi:10.1159/000519363
143. Dong J, Dong W, Ran H, et al. BMSCs-targeting piezoelectric stimulation and immunomodulatory dual-functional hydrogel for promoting diabetic bone regeneration. Mater Today Bio. 2025;33:102015. doi:10.1016/j.mtbio.2025.102015
144. Fan L, Wei A, Gao Z, Mu X. Current progress of mesenchymal stem cell membrane-camouflaged nanoparticles for targeted therapy. Biomed Pharmacother. 2023;161:114451. doi:10.1016/j.biopha.2023.114451
145. Liao J, Lu L, Chu X, et al. Cell membrane coated nanoparticles: cutting-edge drug delivery systems for osteoporosis therapy. Nanoscale. 2024;16(17):8236–8255. doi:10.1039/d3nr06264c
146. Li Q, Liu S, Sun T, et al. Nanoparticles coated with osteoblast-like cell membranes as curcumin delivery vehicles for targeted postmenopausal osteoporosis therapy. Mater Today Bio. 2026;36:102636. doi:10.1016/j.mtbio.2025.102636
147. Kaiser S, Frase S, Selzner L, et al. Neuroprotection after hemorrhagic stroke depends on cerebral heme Oxygenase-1. Antioxidants. 2019;8(10):496. doi:10.3390/antiox8100496
148. Li M, Liu Y, Chen J, et al. Platelet bio-nanobubbles as microvascular recanalization nanoformulation for acute ischemic stroke lesion theranostics. Theranostics. 2018;8(18):4870–4883. doi:10.7150/thno.27466
149. Mathieu M, Martin-Jaular L, Lavieu G, Théry C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol. 2019;21(1):9–17. doi:10.1038/s41556-018-0250-9
150. Shao H, Im H, Castro CM, Breakefield X, Weissleder R, Lee H. New technologies for analysis of extracellular vesicles. Chem Rev. 2018;118(4):1917–1950. doi:10.1021/acs.chemrev.7b00534
151. Sansone R, Stanske B, Keymel S, et al. Macrovascular and microvascular function after implantation of left ventricular assist devices in end-stage heart failure: role of microparticles. J Heart Lung Transplant. 2015;34(7):921–932. doi:10.1016/j.healun.2015.03.004
152. Alexandru N, Andrei E, Niculescu L, Dragan E, Ristoiu V, Georgescu A. Microparticles of healthy origins improve endothelial progenitor cell dysfunction via microRNA transfer in an atherosclerotic hamster model. Acta Physiol. 2017;221(4):230–249. doi:10.1111/apha.12896
153. Bao H, Chen YX, Huang K, et al. Platelet-derived microparticles promote endothelial cell proliferation in hypertension via miR-142-3p. FASEB J. 2018;32(7):3912–3923. doi:10.1096/fj.201701073R
154. Zhang Q, Jeppesen DK, Higginbotham JN, et al. Supermeres are functional extracellular nanoparticles replete with disease biomarkers and therapeutic targets. Nat Cell Biol. 2021;23(12):1240–1254. doi:10.1038/s41556-021-00805-8
155. Santos A, Domingues C, Jarak I, Veiga F, Figueiras A. Osteosarcoma from the unknown to the use of exosomes as a versatile and dynamic therapeutic approach. Eur J Pharm Biopharm. 2022;170:91–111. doi:10.1016/j.ejpb.2021.12.003
156. Wang J, Yeung BZ, Cui M, et al. Exosome is a mechanism of intercellular drug transfer: application of quantitative pharmacology. J Control Release. 2017;268:147–158. doi:10.1016/j.jconrel.2017.10.020
157. Ke W, Afonin KA. Exosomes as natural delivery carriers for programmable therapeutic nucleic acid nanoparticles (NANPs). Adv Drug Deliv Rev. 2021;176:113835. doi:10.1016/j.addr.2021.113835
158. Osteikoetxea X, Silva A, Lázaro-Ibáñez E, et al. Engineered Cas9 extracellular vesicles as a novel gene editing tool. J Extracell Vesicles. 2022;11(5):e12225. doi:10.1002/jev2.12225
159. Wei H, Chen J, Wang S, et al. A nanodrug consisting of doxorubicin and exosome derived from mesenchymal stem cells for osteosarcoma treatment in vitro. Int J Nanomed. 2019;14:8603–8610. doi:10.2147/IJN.S218988
160. Mukherjee P, Berns EJ, Patino CA, et al. Temporal sampling of enzymes from live cells by localized electroporation and quantification of activity by SAMDI mass spectrometry. Small. 2020;16(26):e2000584. doi:10.1002/smll.202000584
161. Faruqu FN, Xu L, Al-Jamal KT. Preparation of exosomes for siRNA delivery to cancer cells. J Vis Exp. 2018;142:10–3791. doi:10.3791/58814
162. Hutapea TP, Madurani KA, Syahputra MY, et al. Albumin: source, preparation, determination, applications, and prospects. J Sci. 2023;8(2):100549. doi:10.1016/j.jsamd.2023.100549
163. Zhu YW, Wei YW, Ma JY, Chen W, Shen Z, Qiu J. Bioactive deproteinized bovine bone mineral based on self-assembled albumin nanoparticles promoted bone regeneration via activation of Wnt/β-catenin pathway. Mater Today Bio. 2025;32:101730. doi:10.1016/j.mtbio.2025.101730
164. Kim M, Schöbel L, Geske M, Boccaccini AR, Ghorbani F. Bovine serum albumin-modified 3D printed alginate dialdehyde-gelatin scaffolds incorporating polydopamine/SiO2-CaO nanoparticles for bone regeneration. Int J Biol Macromol. 2024;264(Pt 2):130666. doi:10.1016/j.ijbiomac.2024.130666
165. Zheng J, Ren Y, Ke J, et al. Engineered extracellular vesicles derived from juvenile mice enhance mitochondrial function in the aging bone microenvironment and achieve rejuvenation. ACS Nano. 2025;19(14):13952–13967. doi:10.1021/acsnano.4c17989
166. Haqqani AS, Thom G, Burrell M, et al. Intracellular sorting and transcytosis of the rat transferrin receptor antibody OX26 across the blood-brain barrier in vitro is dependent on its binding affinity. J Neurochem. 2018;146(6):735–752. doi:10.1111/jnc.14482
167. Fu M, He Q, Guo Z, et al. Therapeutic bispecific T-cell engager antibody targeting the transferrin receptor. Front Immunol. 2019;10:1396. doi:10.3389/fimmu.2019.01396
168. Peng J, Liu K, Cao L, et al. Adenoviral vector for enhanced prostate cancer specific transferrin conjugated drug targeted therapy. Nano Lett. 2022;22(10):4168–4175. doi:10.1021/acs.nanolett.2c00931
169. Lyu X, Zhang Q, Liang D, Huang Y. Interaction between human serum albumin and cholesterol-grafted polyglutamate as the potential carriers of protein drugs. Acta Pharm Sin B. 2019;9(1):186–193. doi:10.1016/j.apsb.2018.08.001
170. Verma D, Gulati N, Kaul S, Mukherjee S, Nagaich U. Protein based nanostructures for drug delivery. J Pharm. 2018;2018:9285854. doi:10.1155/2018/9285854
171. Mousavizadeh A, Jabbari A, Akrami M, Bardania H. Cell targeting peptides as smart ligands for targeting of therapeutic or diagnostic agents: a systematic review. Colloids Surf B Biointerfaces. 2017;158:507–517. doi:10.1016/j.colsurfb.2017.07.012
172. Wang F, Wang Y, Zhang X, Zhang W, Guo S, Jin F. Recent progress of cell-penetrating peptides as new carriers for intracellular cargo delivery. J Control Release. 2014;174:126–136. doi:10.1016/j.jconrel.2013.11.020
173. Zhong C, Li N, Wang S, et al. Targeting osteoblastic 11β-HSD1 to combat high-fat diet-induced bone loss and obesity. Nat Commun. 2024;15(1):8588. doi:10.1038/s41467-024-52965-4
174. Xu H, Wang W, Liu X, et al. Targeting strategies for bone diseases: signaling pathways and clinical studies. Signal Transduct Target Ther. 2023;8(1):202. doi:10.1038/s41392-023-01467-8
175. Wu Y, Sun B, Tang Y, et al. Bone targeted nano-drug and nano-delivery. Bone Res. 2024;12(1):51. doi:10.1038/s41413-024-00356-2
176. Lu Y, Jiang W, Wu X, et al. Peptide T7-modified polypeptide with disulfide bonds for targeted delivery of plasmid DNA for gene therapy of prostate cancer. Int J Nanomed. 2018;13:6913–6927. doi:10.2147/IJN.S180957
177. Zhao Z, Chen C, Xie C, Zhao Y. Design, synthesis and evaluation of liposomes modified with dendritic aspartic acid for bone-specific targeting. Chem Phys Lipids. 2020;226:104832. doi:10.1016/j.chemphyslip.2019.104832
178. Low SA, Galliford CV, Yang J, Low PS, Kopeček J. Biodistribution of fracture-targeted GSK3β inhibitor-loaded micelles for improved fracture healing. Biomacromolecules. 2015;16(10):3145–3153. doi:10.1021/acs.biomac.5b00777
179. Wang Y, Yang J, Liu H, et al. Osteotropic peptide-mediated bone targeting for photothermal treatment of bone tumors. Biomaterials. 2017;114:97–105. doi:10.1016/j.biomaterials.2016.11.010
180. Wang J, Li X, Wang S, Cui J, Ren X, Su J. Bone-targeted exosomes: strategies and applications. Adv Healthc Mater. 2023;12(18):e2203361. doi:10.1002/adhm.202203361
181. Zhang G, Guo B, Wu H, et al. A delivery system targeting bone formation surfaces to facilitate RNAi-based anabolic therapy. Nat Med. 2012;18(2):307–314. doi:10.1038/nm.2617
182. Anselmino N, Starbuck M, Labanca E, et al. Heme Oxygenase-1 is a pivotal modulator of bone turnover and remodeling: molecular implications for prostate cancer bone metastasis. Antioxid Redox Signal. 2020;32(17):1243–1258. doi:10.1089/ars.2019.7879
183. Wang S, Gu R, Wang F, et al. 3D-Printed PCL/Zn scaffolds for bone regeneration with a dose-dependent effect on osteogenesis and osteoclastogenesis. Mater Today Bio. 2022;13:100202. doi:10.1016/j.mtbio.2021.100202
184. Mediero A, Wilder T, Reddy VSR, et al. Ticagrelor regulates osteoblast and osteoclast function and promotes bone formation in vivo via an adenosine-dependent mechanism. FASEB J. 2016;30(11):3887–3900. doi:10.1096/fj.201600616R
185. Gu W, Wu C, Chen J, Xiao Y. Nanotechnology in the targeted drug delivery for bone diseases and bone regeneration. Int J Nanomed. 2013;8:2305–2317. doi:10.2147/IJN.S44393
186. Zheng K, Torre E, Bari A, et al. Antioxidant mesoporous Ce-doped bioactive glass nanoparticles with anti-inflammatory and pro-osteogenic activities. Mater Today Bio. 2020;5:100041. doi:10.1016/j.mtbio.2020.100041
187. Wang J, Tang Y, Cao Q, et al. Fabrication and biological evaluation of 3D-printed calcium phosphate ceramic scaffolds with distinct macroporous geometries through digital light processing technology. Regen Biomater. 2022;9:rbac005. doi:10.1093/rb/rbac005
188. Sun C, Yuan H, Wang L, et al. FAK promotes osteoblast progenitor cell proliferation and differentiation by enhancing Wnt signaling. J Bone Miner Res. 2016;31(12):2227–2238. doi:10.1002/jbmr.2908
189. Jiang T, Xia C, Chen X, et al. Melatonin promotes the BMP9-induced osteogenic differentiation of mesenchymal stem cells by activating the AMPK/β-catenin signalling pathway. Stem Cell Res Ther. 2019;10(1):408. doi:10.1186/s13287-019-1511-7
190. Thiel WH, Esposito CL, Dickey DD, et al. Smooth muscle cell-targeted RNA aptamer inhibits neointimal formation. Mol Ther. 2016;24(4):779–787. doi:10.1038/mt.2015.235
191. Ha J, Kang JS, Lee M, et al. Simplified brain organoids for rapid and robust modeling of brain disease. Front Cell Dev Biol. 2020;8:594090. doi:10.3389/fcell.2020.594090
192. Biasoli D, Sobrinho MF, da Fonseca ACC, et al. Glioblastoma cells inhibit astrocytic p53-expression favoring cancer malignancy. Oncogenesis. 2014;3(10):e123. doi:10.1038/oncsis.2014.36
193. Zhang W, Zhang Y, Hao Z, et al. Synthetic nanoparticles functionalized with cell membrane-mimicking, bone-targeting, and ROS-controlled release agents for osteoporosis treatment. J Control Release. 2025;378:306–319. doi:10.1016/j.jconrel.2024.12.017
194. Shi Y, Massagué J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell. 2003;113(6):685–700. doi:10.1016/s0092-8674(03)00432-x
195. Chen J, Liu M, Luo X, et al. Exosomal miRNA-486-5p derived from rheumatoid arthritis fibroblast-like synoviocytes induces osteoblast differentiation through the Tob1/BMP/Smad pathway. Biomater Sci. 2020;8(12):3430–3442. doi:10.1039/c9bm01761e
196. Liu A, Lin D, Zhao H, et al. Optimized BMSC-derived osteoinductive exosomes immobilized in hierarchical scaffold via lyophilization for bone repair through Bmpr2/Acvr2b competitive receptor-activated Smad pathway. Biomaterials. 2021;272:120718. doi:10.1016/j.biomaterials.2021.120718
197. Ji X, Yuan X, Ma L, et al. Mesenchymal stem cell-loaded thermosensitive hydroxypropyl chitin hydrogel combined with a three-dimensional-printed poly(ε-caprolactone) /nano-hydroxyapatite scaffold to repair bone defects via osteogenesis, angiogenesis and immunomodulation. Theranostics. 2020;10(2):725–740. doi:10.7150/thno.39167
198. Shan Z, Bi H, Suonan A, et al. Tobacco mosaic viral nanoparticle inhibited osteoclastogenesis through inhibiting mTOR/AKT signaling. Int J Nanomed. 2020;15:7143–7153. doi:10.2147/IJN.S245870
199. Kim SE, Choi S, Hong JY, et al. Accelerated osteogenic differentiation of MC3T3-E1 cells by lactoferrin-conjugated nanodiamonds through enhanced anti-oxidant and anti-inflammatory effects. Nanomaterials. 2019;10(1):50. doi:10.3390/nano10010050
200. Wang J, Meng F, Song W, et al. Nanostructured titanium regulates osseointegration via influencing macrophage polarization in the osteogenic environment. Int J Nanomed. 2018;13:4029–4043. doi:10.2147/IJN.S163956
201. Fan L, Guan P, Xiao C, et al. Exosome-functionalized polyetheretherketone-based implant with immunomodulatory property for enhancing osseointegration. Bioact Mater. 2021;6(9):2754–2766. doi:10.1016/j.bioactmat.2021.02.005
202. Hao Z, Ren L, Zhang Z, et al. A multifunctional neuromodulation platform utilizing Schwann cell-derived exosomes orchestrates bone microenvironment via immunomodulation, angiogenesis and osteogenesis. Bioact Mater. 2023;23:206–222. doi:10.1016/j.bioactmat.2022.10.018
203. Wu H, Chen C, Li J, et al. Engineered magneto-piezoelectric nanoparticles-enhanced scaffolds disrupt biofilms and activate oxidative phosphorylation in Icam1+ macrophages for infectious bone defect regeneration. ACS Nano. 2024;18(52):35575–35594. doi:10.1021/acsnano.4c13562
204. Gao A, Liao Q, Xie L, et al. Tuning the surface immunomodulatory functions of polyetheretherketone for enhanced osseointegration. Biomaterials. 2020;230:119642. doi:10.1016/j.biomaterials.2019.119642
205. Chen L, Wang D, Peng F, et al. Nanostructural surfaces with different elastic moduli regulate the immune response by stretching macrophages. Nano Lett. 2019;19(6):3480–3489. doi:10.1021/acs.nanolett.9b00237
206. Sadowska JM, Wei F, Guo J, et al. The effect of biomimetic calcium deficient hydroxyapatite and sintered β-tricalcium phosphate on osteoimmune reaction and osteogenesis. Acta Biomater. 2019;96:605–618. doi:10.1016/j.actbio.2019.06.057
207. Zhang Z, Hao Z, Xian C, Zhang J, Wu J. Triple functional magnesium ascorbyl phosphate encapsulated hydrogel: a cosmetic ingredient promotes bone repair via anti-oxidation, calcium uptake and blood vessel remodeling. Chem Eng J. 2023;472:145061. doi:10.1016/j.cej.2023.145061
208. Ma B, Wang T, Li J, Wang Q. Extracellular matrix derived from Wharton’s Jelly-derived mesenchymal stem cells promotes angiogenesis via integrin αVβ3/c-Myc/P300/VEGF. Stem Cell Res Ther. 2022;13(1):327. doi:10.1186/s13287-022-03009-5
209. Marrella A, Lee TY, Lee DH, et al. Engineering vascularized and innervated bone biomaterials for improved skeletal tissue regeneration. Mater Today. 2018;21(4):362–376. doi:10.1016/j.mattod.2017.10.005
210. Qin Q, Lee S, Patel N, et al. Neurovascular coupling in bone regeneration. Exp Mol Med. 2022;54(11):1844–1849. doi:10.1038/s12276-022-00899-6
211. Jiang H, Xia W, Xia T, et al. Chemotactic recruitment of genetically engineered cell membrane-camouflaged metal-organic framework nanoparticles for ischemic osteonecrosis treatment. Acta Biomater. 2024;185:410–428. doi:10.1016/j.actbio.2024.07.024
212. Yang W, Pan Q, Peng Y, et al. Dual-target nanotherapy for vascular endothelium and bone mesenchymal stem cells halt steroid-induced osteonecrosis of the femoral head progression. J Control Release. 2025;380:219–239. doi:10.1016/j.jconrel.2024.12.081
213. Yu T, Xu Y, Ahmad MA, Javed R, Hagiwara H, Tian X. Exosomes as a promising therapeutic strategy for peripheral nerve injury. Curr Neuropharmacol. 2021;19(12):2141–2151. doi:10.2174/1570159X19666210203161559
214. Todorova D, Simoncini S, Lacroix R, Sabatier F, Dignat-George F. Extracellular vesicles in angiogenesis. Circ Res. 2017;120(10):1658–1673. doi:10.1161/CIRCRESAHA.117.309681
215. Zha Y, Li Y, Lin T, Chen J, Zhang S, Wang J. Progenitor cell-derived exosomes endowed with VEGF plasmids enhance osteogenic induction and vascular remodeling in large segmental bone defects. Theranostics. 2021;11(1):397–409. doi:10.7150/thno.50741
216. Su Y, Gao Q, Deng R, et al. Aptamer engineering exosomes loaded on biomimetic periosteum to promote angiogenesis and bone regeneration by targeting injured nerves via JNK3 MAPK pathway. Mater Today Bio. 2022;16:100434. doi:10.1016/j.mtbio.2022.100434
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Exosomes as Emerging Therapeutic Strategies in Primary Osteoporosis: A Narrative Review
Chen G, Li H, Chen R, Chen G
Drug Design, Development and Therapy 2025, 19:9099-9115
Published Date: 7 October 2025