Back to Journals » Drug Design, Development and Therapy » Volume 20
Beyond Symptom Suppression: The Multitargeted Reversal of Chronic Pain by Maresin1
Authors Wang F, Yang Y, Wang W, Shi Q, Fan B
Received 30 November 2025
Accepted for publication 24 March 2026
Published 25 April 2026 Volume 2026:20 585597
DOI https://doi.org/10.2147/DDDT.S585597
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Tin Wui Wong
Fuquan Wang,* Yang Yang,* Wen Wang, Qing Shi, Bifa Fan
Department of Pain Medicine, China-Japan Friendship Hospital, Beijing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Fuquan Wang, Department of Pain Management, China-Japan Friendship Hospital, Beijing, People’s Republic of China, Email [email protected]
Abstract: The transition from acute inflammation to chronic pain represents a significant clinical challenge, often driven by a failure of endogenous resolution programs. Specialized pro-resolving mediators (SPMs), derived from polyunsaturated fatty acids, are crucial for actively terminating inflammation and restoring tissue homeostasis. Maresin 1 (MaR1), a prototypical SPM biosynthesized from docosahexaenoic acid (DHA), has emerged as a powerful modulator of pain. This review comprehensively synthesizes the current preclinical evidence, primarily derived from preclinical animal models, detailing the analgesic effects of MaR1 across a spectrum of pain models, including inflammatory, neuropathic, postoperative, osteoarthritis-related pain, etc. We dissect the multifaceted mechanisms underlying its efficacy, which extend beyond simple anti-inflammation. MaR1 exerts its effects by: (1) attenuating neuroinflammation including the suppression of glial (microglia and astrocyte) activation and reprogramming macrophage phenotypes; (2) directly modulating neuronal function by inhibiting nociceptive ion channels (e.g. TRPV1) and reversing central synaptic plasticity; and (3) promoting robust tissue repair, including peripheral nerve regeneration. These actions are mediated through potential specific receptors, notably G-protein-coupled receptor 37-like 1 (GPR37L1) on glial cells and retinoic acid–related orphan receptor α (RORA) on neurons. While MaR1 demonstrates significant therapeutic potential, challenges related to its pharmacokinetic instability and observed sex-dependent analgesic effects must be addressed for successful clinical translation. This review provides a comprehensive mechanistic framework supporting MaR1 as a next-generation therapeutic candidate for pain management, and outlines the core research directions to overcome key translational barriers for MaR1-based therapies.
Keywords: Maresin1, pain modulation mechanism, neuroinflammation resolution, nociceptor modulation, tissue repair
Introduction
Chronic pain remains a pervasive and unmet clinical challenge across multiple clinical disorders, affecting hundreds of millions of people worldwide and imposing a heavy burden on individuals and healthcare systems. Conventional analgesic therapies, including non-steroidal anti-inflammatory drugs (NSAIDs), opioids and ion channel blockers, primarily exert passive symptomatic suppression by blocking pro-inflammatory signaling pathways or nociceptive transmission. However, these therapies fail to target the root causes of chronic pain, such as impaired endogenous inflammation resolution, neural damage, glial dysregulation, and maladaptive synaptic plasticity, are often accompanied by severe side effects such as immunosuppression, opioid tolerance, addiction, and gastrointestinal damage.1,2 The transition from acute inflammation to chronic pain remains a central therapeutic challenge across multiple clinical disorders, with dysregulation of endogenous resolution programs emerging as a key driver of persistent pain.3–5
Specialized pro-resolving mediators (SPMs) are a class of endogenous lipid autacoids biosynthesized from polyunsaturated fatty acids (PUFAs), including ω-3 PUFAs such as docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA).6 The SPM family includes four major subclasses: resolvins, protectins, lipoxins, and maresins, each with distinct biosynthesis pathways and biological functions. Resolvins are divided into E-series (from EPA) and D-series (from DHA), which exert anti-inflammatory effects by inhibiting neutrophil infiltration and promoting macrophage efferocytosis; protectins (from DHA) are predominantly involved in neuroprotection and tissue repair; lipoxins (from arachidonic acid) are the first discovered SPMs and regulate early inflammation resolution; maresins (from DHA) are newly identified SPMs with unique pro-regenerative and analgesic properties.7,8 All SPMs share a core biological function: actively orchestrating the termination of acute inflammation, restoring tissue homeostasis and maintaining immune balance, which is fundamentally different from traditional anti-inflammatory agents that only passively inhibit pro-inflammatory signals and cannot promote the return to physiological immune and tissue homeostasis. Developing therapeutics based on SPMs—resolution pharmacology—has become a novel and promising direction for chronic pain treatment.9,10
Among these, maresin-1 (MaR1), the prototypical member of the maresin family, has attracted increasing attention for its potent anti-inflammatory, pro-regenerative, and neuroprotective properties, positioning it as a promising endogenous modulator of pain.11 MaR1 as an endogenous specialized pro-resolving mediator (SPM), is biosynthesized from the ω-3 polyunsaturated fatty acid docosahexaenoic acid (DHA) via the lipoxygenase (LOX) pathway. First isolated and characterized from activated human macrophages in 2009,12 MaR1 is chemically defined as 7R,14S-dihydroxydocosa-4Z,8E,10E,12Z,16Z,19Z-hexaenoic acid. Rather than merely blocking the initiation of inflammation—an action typical of conventional anti-inflammatory agents—MaR1 actively drives resolution, thereby restoring tissue homeostasis.13 MaR1 exerts potent, nanomolar-range bioactivity through potential specific receptors, including leucine-rich repeat-containing G-protein-coupled receptor 6 (LGR6),14 retinoic acid-related orphan receptor α (RORA),15 and G-protein-coupled receptor 37-like 1 (GPR37L1).16 However, the evidence for these receptor–ligand relationships remains limited and controversial. The cell-type–restricted expression of these receptors across immune, neural, and parenchymal cells underpins MaR1’s capacity for precise, multi-tissue modulation without overt immunosuppressive side effects.
Pain initiation and persistence are intimately coupled to inflammatory cascades, neural damage, glial activation, and non-inflammatory pathological processes such as maladaptive synaptic plasticity and central sensitization.17 Maresin-1 (MaR1) selectively engages these critical pathophysiological nodes, thereby exerting pronounced analgesic efficacy across diverse pre-clinical pain models.18,19 Beyond directly modulating nociceptive signaling in neuropathic states, MaR1 simultaneously remodels the inflammatory microenvironment and repairs tissue damage, achieving a synergistic “symptom-plus-cause” analgesia.20 This multi-level mechanistic coverage positions MaR1 as a novel therapeutic strategy for chronic pain management. However, MaR1 faces the general challenges of developing lipid-based therapeutics, including rapid enzymatic metabolism, short biological half-life, poor water solubility, and delivery difficulties, which limit its direct clinical application.
While previous reviews have focused on the lipidomics of SPMs or the single mechanism of MaR1 in inflammation, this review uniquely synthesizes the tripartite interaction between neurons, glia, and immune cells underlying MaR1’s analgesic effects, and systematically integrates its analgesic spectrum, multi-target mechanisms, and translational strategies. Accordingly, this review will systematically collate the evidence for MaR1 across diverse pain phenotypes, dissect the molecular mechanisms by which it achieves analgesia via “modulation of neuronal excitability, correction of immune–inflammatory dysregulation, and facilitation of tissue repair,” and critically evaluate the distinct signaling cascades involved. Emphasis will be placed on receptor- and cell-type-specific pathways with cautious discussion of mechanistic uncertainties, translational pharmacokinetics, formulation strategies, and sex-dependent efficacy—key determinants for clinical translation. The review aims to provide a mechanistic framework supporting MaR1 as a next-generation therapeutic for chronic pain and to chart actionable directions for mitigating its persistence and refractoriness in the clinic.
Analgesic Spectrum of MaR1: Beneficial Effects Across Diverse Pain Types
As a key endogenous molecule that promotes inflammation resolution and tissue repair, MaR1 has demonstrated significant analgesic potential across various preclinical pain models.13,21 Its analgesic spectrum encompasses inflammation-dominant pain, pain resulting from nerve injury, and other mixed or specific pain states. All preclinical studies on MaR1’s analgesic effects are systematically summarized in Table 1.
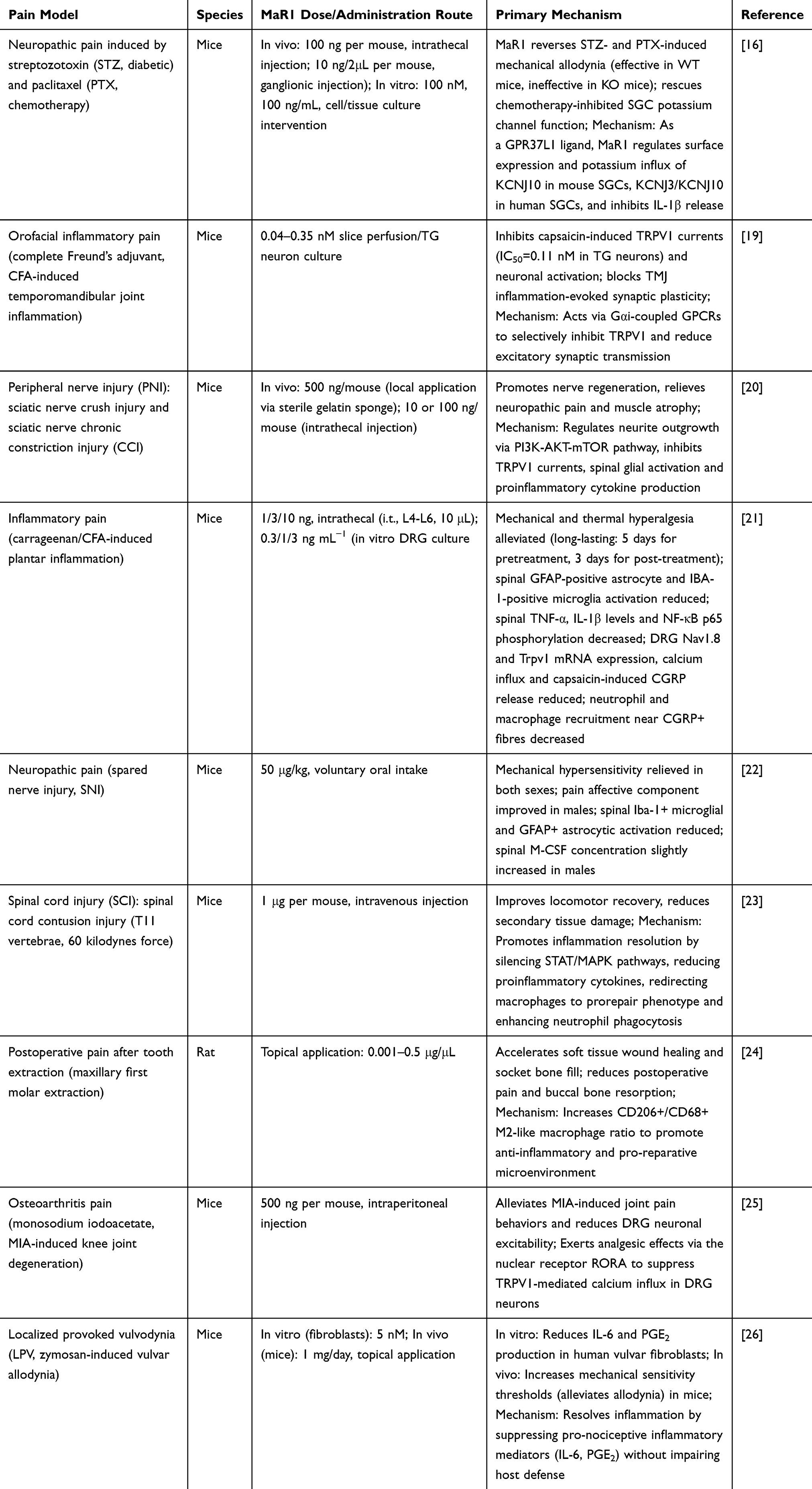 |
Table 1 The Summary of Core Experimental Details, Outcomes, and Mechanisms of MaR1 in Various Pain Models |
Inflammatory Pain
MaR1 exhibits potent analgesic effects in various acute and chronic inflammatory pain models. In mouse plantar inflammation models induced by carrageenan or complete Freund’s adjuvant (CFA), local or intrathecal administration of low-dose (nanogram level) MaR1 effectively alleviates mechanical allodynia and thermal hyperalgesia.21,27,28 Its analgesic mechanism is directly associated with MaR1’s robust pro-resolving properties, including inhibiting excessive neutrophil infiltration into inflammatory sites, inducing macrophage polarization toward the M2 phenotype, reducing the sustained sensitization of peripheral nociceptors by inflammatory factors, and downregulating pro-inflammatory cytokines (TNF-α, IL-1β) and NF-κB activation in the spinal cord,20,22 etc.
Neuropathic Pain (NP)
Neuropathic pain is a chronic pain directly caused by damage to or disease of the somatosensory nervous system, with mechanisms involving the sensitization of both peripheral and central nervous systems.29,30 MaR1 has demonstrated significant efficacy in various NP models:
Traumatic Nerve Injury Models
In models such as spinal nerve ligation (SNL), chronic constriction injury (CCI), and sciatic nerve crush (SNC), intrathecal injection or local application of MaR1 not only prevents but also effectively reverses established mechanical allodynia and thermal hyperalgesia.20 The analgesic effect of MaR1 in traumatic nerve injury models is sustained for several days, which is superior to traditional anti-inflammatory agents with short-term effects and rapid tolerance.
Neuroprotective and Regenerative Effects
Beyond alleviating pain symptoms, MaR1 also exhibits potential in promoting nerve regeneration.20,23 In the sciatic nerve crush model, MaR1 was more effective than nerve growth factor (NGF) in facilitating the recovery of sensory and motor functions, reducing dorsal root ganglion (DRG) neuron damage (evidenced by decreased ATF3 expression), promoting neural pathway reconstruction (increased DiI-labeled neurons) and axonal regeneration (elevated NF200 expression), as well as inhibiting muscle atrophy following nerve injury.20 In vitro experiments confirmed that MaR1 can directly promote axonal growth of DRG neurons. Its regenerative mechanism may involve the activation of the PI3K-AKT-mTOR signaling pathway.20
Chemotherapy-Induced Peripheral Neuropathy (CIPN)
MaR1also shows analgesic effects in chemotherapy-induced pain models, such as those induced by paclitaxel or oxaliplatin.31
Other Types of Pain
Postoperative Pain (POP)
In the mouse femoral fracture postoperative pain (fPOP) model, perioperative intravenous injection of MaR1 effectively delays the development of mechanical allodynia and cold allodynia.32 Intrathecal injection of MaR1 also efficiently alleviates established fPOP. Notably, MaR1 is far more potent than its precursor docosahexaenoic acid (DHA) in preventing fPOP (requiring a 1000-fold lower dose). In the fPOP model, the analgesic effect of MaR1 is superior to that of Resolvin D1 (RvD1). In a rat model of tooth extraction, local application of MaR1 not only accelerates soft tissue healing and alveolar socket bone filling, and preserves alveolar ridge morphology, but also significantly reduces immediate postoperative pain scores.24 This indicates that MaR1 possesses dual potential for promoting healing and relieving pain after oral surgery.13,24 MaR1 also exhibits analgesic effects in other types of postoperative pain models, such as thoracotomy-induced pain and muscle stretch pain.33
Osteoarthritis Pain (OA)
In the monosodium iodoacetate (MIA)-induced mouse OA model, intraperitoneal injection of MaR1 (500 ng) significantly alleviates established OA-like pain behaviors (evidenced by increased von Frey threshold and improved dynamic weight-bearing). Its analgesic effect can last for several days. MaR1 also reduces the expression of pain-related calcitonin gene-related peptide (CGRP) and F4/80+ macrophage infiltration in the dorsal root ganglion (DRG) of OA mice, and decreases the proportion of CD68+ activated macrophages.25,34
Orofacial Pain
In TMJ-related trigeminal nociceptors, MaR1 potently inhibits capsaicin-induced TRPV1 channel currents and neuronal activity. Its half-maximal inhibitory concentration (IC50) for TRPV1 inhibition is lower in trigeminal ganglion (TG) neurons than in dorsal root ganglion (DRG) neurons (0.11 nM vs 0.17 nM), indicating a tissue-specific and cell-type-specific inhibitory effect of MaR1 on nociceptive ion channels. On brain slices of the spinal trigeminal caudal nucleus (Sp5C), MaR1 blocks capsaicin-induced enhancement of excitatory synaptic transmission (increased frequency of spontaneous excitatory postsynaptic currents, sEPSCs) and reverses synaptic plasticity changes caused by CFA-induced TMJ inflammation.19,35 These results suggest that MaR1 has potential as a novel endogenous inhibitor for the treatment of TMJ inflammatory pain.
Local Provoked Pain
In the zymosan-induced mouse vulvodynia model, topical application of MaR1 increases the mechanical sensitivity threshold (ie, relieves pain) and reduces the level of prostaglandin E2. In vitro, MaR1 also significantly decreases the production of pronociceptive inflammatory mediators interleukin-6 (IL-6) and prostaglandin E2 (PGE2) by vulvar fibroblasts derived from patients with localized provoked vulvodynia (LPV).26 Pre-treatment with MaR1 yields more pronounced suppression of pro-inflammatory mediator release than post-treatment, suggesting that early intervention with MaR1 may be a more effective therapeutic strategy for local provoked pain associated with fibroblast activation and chronic tissue inflammation. This suggests that MaR1 may be effective in the treatment of local provoked pain such as LPV.
The Core Mechanisms of Maresin-Mediated Pain Regulation
Maresin 1 (MaR1) exerts potent analgesic effects by coordinately regulating three core pathophysiological nodes: neuroinflammation, neuronal function, and tissue repair (as shown in Figure 1). The actions of MaR1 are mediated through potential-specific receptors, whose receptor–ligand relationships and downstream intracellular signaling pathways remain under active investigation and validation in the SPM field. A critical and objective discussion of the evidence for MaR1 receptor assignments is necessary for an unbiased review: supporting evidence mainly comes from in vitro binding assays, cell function experiments, and knockout mouse models, while major limitations include lack of human sample validation, cross-reactivity with other lipid mediators and orphan GPCRs, and inconsistent results in different pain models and species. Below is a detailed elaboration of the cell-type-specific functional consequences and downstream intracellular signaling pathways of MaR1’s action on different immune and neural cells, which is the core mechanistic basis of its analgesic effect.
 |
Figure 1 Schematic diagram of the core mechanisms underlying MaR1-mediated analgesia and tissue repair in chronic pain. Upward arrows (↑) represent an increase in expression, activity or cellular function; downward arrows (↓) represent a decrease in expression, activity or cellular function. |
Attenuation of Glial Reactivity
The analgesic effect of MaR1 is attributed to its potent anti-inflammatory and pro-resolving properties.36 Chronic pain is increasingly recognized as a neuroinflammatory disorder, characterized by the sustained activation of spinal and supraspinal glial cells (microglia and astrocytes) and the infiltration of peripheral immune cells into neural structures (eg, DRG, spinal cord).37 Maresins target these processes at multiple levels. By remodeling the inflammatory microenvironment, MaR1 reduces the sustained stimulation of nociceptors caused by pro-inflammatory mediators, thereby facilitating analgesia.38
In neuropathic pain states, microglia and astrocytes in the spinal dorsal horn become reactive, releasing a barrage of pro-nociceptive mediators that enhance central sensitization.39,40 MaR1 acts as a potent “off-switch” for this glial activation. Neuro-inflammation modulation by MaR1 is characterized by a marked suppression of spinal glial activation after spared-nerve injury (SNI). Quantitative immunofluorescence revealed that MaR1 treatment significantly decreased Iba-1 (microglia) and GFAP (astrocyte) signal intensities in the spinal dorsal horn, thereby attenuating local neuro-inflammation, MaR1 can effectively reduce the concentration of macrophage colony-stimulating factor (M-CSF) in the spinal cord; however, this reduction exhibits a gender bias, with male subjects deriving greater benefit from MaR1 intervention,22 which is the key cellular and molecular reason for the sex-dependent analgesic efficacy of MaR1 in neuropathic pain models.
The analgesic effect of MaR1 is not just suppressive; it is homeostatic. Single-nucleus RNA sequencing (snRNA-seq) identifies GPR37L1 as a GPCR highly enriched in spinal dorsal horn (SDH) astrocytes.41 Following peripheral nerve injury, astrocytic GPR37L1 expression is reduced. MaR1 binding to GPR37L1 on astrocytes specifically enhances the activity and expression of the glutamate transporter GLT-1 (EAAT2)—the core downstream functional consequence of this receptor–ligand interaction. This accelerates glutamate clearance from the synaptic cleft, preventing extrasynaptic glutamate accumulation and subsequent excitotoxicity.41 This results in a marked reduction in both the frequency and amplitude of excitatory postsynaptic currents (EPSCs) in dorsal horn neurons, dampening central sensitization. The analgesic effect of intrathecal MaR1 is abolished in Gpr37l1-knockout mice, confirming this astrocyte-specific, GPR37L1-dependent mechanism.41 However, the direct binding affinity between MaR1 and GPR37L1 is only validated in mouse astrocyte models, and no direct binding data and functional validation in human astrocyte and neural tissue samples are available to date.
Reprogramming of Macrophage Phenotypes
Macrophages infiltrating the DRG and injured nerves are key determinants of pain persistence. MaR1 exerts antinociceptive effects in persistent pain states by orchestrating macrophage-mediated inflammation.42 In the K/BxN serum-transfer arthritis model, resolution-phase joint swelling is accompanied by sustained hypersensitivity that coincides with markedly reduced MaR1 levels in dorsal-root-ganglion (DRG) tissue and a concomitant increase in pro-inflammatory (M1) macrophage infiltration. In vitro, MaR1 suppresses lipopolysaccharide-stimulated peritoneal macrophage polarization toward an M1 phenotype, as evidenced by decreased expression of TNF-α, IL-6, inducible nitric oxide synthase (NOS2), and miR-155. The downstream intracellular signaling mechanism by which MaR1 inhibits macrophage M1 polarization and exerts anti-inflammatory effects may involve the regulation of miR-155, which is known to modulate TNF-α and NOS2 production and NF-κB signaling, thereby blocking the release of pro-inflammatory cytokines. In vivo, repeated systemic administration of MaR1 diminishes the abundance of M1 macrophages in both DRG and joint compartments; this macrophage re-programming correlates with reversal of persistent hypersensitivity. The sustained analgesic effect appears to be mediated primarily by long-term modulation of macrophage infiltration and function rather than by acute anti-inflammatory actions.
Similarly, in an osteoarthritis model, maresin-1 (MaR1) exerts multi-level control over pain signaling and inflammation within dorsal-root-ganglion (DRG) neurons.25 It down-regulates the nociceptive marker calcitonin gene-related peptide (CGRP) in the DRG, thereby attenuating nociceptive transmission. MaR1 also markedly reduces the number of activated macrophages—both F4/80⁺ and CD68⁺ subsets—in the DRG, alleviating localized neuroinflammation. Additionally, systemic administration of MaR1 lowers circulating levels of interferon-γ (IFN-γ), eotaxin-2, and interleukin-2 (IL-2), further diminishing the impact of systemic inflammation on DRG nociceptive signaling.
Maintenance of Satellite Glial Cell Homeostasis
G-protein-coupled receptor 37-like 1 (GPR37L1) is classified as an orphan GPCR.43 Prosaposin and its derived peptides, such as TX-14, have been suggested as potential ligands for both GPR37 and GPR37L1, demonstrating neuroprotective and glioprotective effects.44,45 Studies have demonstrated that GPR37L1 is highly expressed in DRGs of both mice and humans and is selectively localized to satellite glial cells (SGCs).16 In human SGCs, the receptor is enriched at the soma-neurite interface that faces the neuron, providing an anatomical substrate for neuron–glia communication. In streptozotocin (STZ)-induced diabetic neuropathy and paclitaxel (PTX)-induced chemotherapy-related neuropathy models, membrane-associated GPR37L1 is markedly reduced, whereas maresin-1 (MaR1) has been identified as a selective ligand that exerts its effects through direct binding to GPR37L1.
Mechanistically, MaR1 modulates potassium channel function via GPR37L1: in murine SGCs it rescues PTX-impaired plasma-membrane localization of KCNJ10 (Kir4.1) and restores inwardly rectifying K⁺ currents, whereas in human SGCs it enhances the activity of KCNJ3 (Kir3.1). Simultaneously, MaR1 suppresses PTX-evoked release of the pro-inflammatory cytokine IL-1β from SGCs via GPR37L1-mediated signaling, thereby preserving potassium homeostasis and attenuating neuroinflammation. Intrathecal or DRG-restricted administration of MaR1 significantly alleviates neuropathic pain in both STZ and PTX models, an effect that is abolished in Gpr37L1-deficient mice, confirming that the analgesic action is GPR37L1-dependent. Moreover, the human GPR37L1-E296K variant, which disrupts receptor function, increases susceptibility to chronic pain, further underscoring the importance of GPR37L1 in nociceptive processing.16
Regulation of Non-Immune Cells
Pain is also driven by non-immune, non-neuronal cells, such as fibroblasts, which release pro-nociceptive mediators.46 Maresin-1 (MaR1) could attenuate localized provoked vulvodynia (LPV) by reprogramming the inflammatory secretome of vulvar fibroblasts.26 In vitro, MaR1 applied either before (pre-treatment) or after (post-treatment) inflammatory challenge markedly reduced IL-6 and prostaglandin E2 (PGE2) release from LPV-patient-derived vulvar fibroblasts, with pre-treatment yielding the more pronounced suppression. In a mouse model of vulvar pain, topical MaR1 decreased local PGE2 concentrations and simultaneously elevated mechanical pain thresholds, reversing hyperalgesia more effectively than vehicle. These effects are attributable to MaR1-mediated restriction of pro-nociceptive mediator secretion by vulvar fibroblasts, underscoring its therapeutic relevance in LPV.
Direct Modulation of Nociceptor Function and Synaptic Plasticity
Beyond its actions on glia and immune cells, MaR1 directly modulates the excitability of primary sensory neurons and the plasticity of central pain circuits.
Inhibition of Nociceptive Ion Channels
Peripheral sensitization is driven by the post-translational modification and sensitization of nociceptive ion channels, particularly TRPV1 (Transient Receptor Potential Vanilloid 1).47 A mechanistic study of MaR1 in the trigeminal system demonstrated potent, dose-dependent suppression of capsaicin-evoked TRPV1 currents in trigeminal ganglion (TG) neurons, with an IC50 = 0.11 nM.19 MaR1 completely abolished capsaicin-triggered action-potential firing but had no effect on TRPA1 currents, confirming its high selectivity for TRPV1 among nociceptive ion channels. This rapid, acute inhibition of TRPV1 is Gαi-coupled GPCR-dependent, as it was abolished by pertussis toxin (PTX).19 This Gαi signaling likely inhibits adenylyl cyclase, reduces cAMP levels, and decreases PKA-mediated phosphorylation of TRPV1, thereby reducing channel sensitization.
This neuronal inhibition is also RORA-dependent, involving a slow, genomic regulatory mechanism that complements the rapid non-genomic Gαi signaling pathway, forming a dual regulatory mechanism of MaR1 on nociceptive ion channels. In an OA model, MaR1 significantly suppresses DRG neuronal excitability.25 Calcium-imaging experiments revealed that MaR1 dose-dependently attenuates capsaicin-evoked Ca2⁺ influx in DRG neurons. This effect is contingent upon the nuclear receptor RORA: co-application of the RORA inverse agonist SR3335 completely reversed MaR1-mediated suppression. MaR1 regulates the functional response of DRG neurons via RORA, and reduces the calcium response of DRG neurons induced by the TRPV1 agonist capsaicin, thereby exerting an analgesic effect in osteoarthritis. This suggests a dual mechanism: rapid Gi/o coupled inhibition and slower, RORA-mediated transcriptional regulation of the nociceptive machinery.
However, the direct inhibitory effects may be context-dependent. Some studies note that MaR1, compared with gabapentin, exerts modest direct inhibitory effects on DRG neurons and produces no acute systemic analgesia, implying its primary antinociceptive action arises from long-term modulation of the inflammatory milieu rather than rapid neuronal block.42
Attenuation of Synaptic Plasticity
The transition to chronic pain involves robust synaptic plasticity in the spinal dorsal horn, manifesting as long-term potentiation (LTP) of C-fiber inputs.48 MaR1 can reverse this central sensitization. At the synaptic level, MaR1 did not alter basal spontaneous excitatory postsynaptic currents (sEPSCs) in superficial lamina neurons of the caudal subnucleus of the spinal trigeminal tract (Sp5C).19 However, it fully blocked capsaicin-induced increases in sEPSC frequency. In a CFA-induced TMJ inflammation model, MaR1 reversed inflammation-evoked elevations in sEPSC frequency and amplitude, indicating central antinociceptive actions. This effect is likely secondary to its actions on both presynaptic TRPV1-expressing terminals (reducing glutamate release) and postsynaptic glial regulation (via GPR37L1-mediated glutamate uptake).
Promotion of Tissue Repair and Regeneration
Peripheral Nerve Regeneration
Maresin-1 (MaR1) exerts robust pro-regenerative effects after peripheral nerve injury (PNI). In vitro, MaR1 stimulates neurite outgrowth from dorsal-root-ganglion (DRG) neurons in a dose-dependent manner, with a half-maximal effective concentration (EC50) of ≈1 ng/mL—substantially lower than that of nerve growth factor (NGF, 7–9 ng/mL). At 10 ng/mL, MaR1 outperforms the same concentration of NGF in extending neurite length.4
In vivo, local application of MaR1 in a mouse sciatic-nerve crush model reduces neuronal injury (fewer ATF-3⁺ DRG neurons), accelerates pathway re-establishment as verified by Dil tracing, and mitigates gastrocnemius atrophy (increased muscle mass and volume). Functionally, MaR1 accelerates both motor and sensory recovery. Rotarod assays show superior and faster restoration of complex motor performance compared with NGF, while footprint analysis and nociceptive tests further demonstrate its advantage in basic locomotion and sensory recovery.
Accelerating Healing of Other Tissues
Maresin-1 (MaR1) plays a pivotal role in promoting wound healing and alveolar-bone regeneration of rat extraction sockets.24 Following extraction of the maxillary first molar, gelfoam sponges pre-loaded with graded concentrations of MaR1 or saline were placed in the sockets and locally administered twice weekly until wound closure. Comprehensive assessment was conducted through macroscopic inspection, histomorphometry, micro-CT, immunohistochemistry, and pain behaviour.
Macroscopically, MaR1 accelerated soft-tissue healing; at an optimal concentration, the wound closure rate on post-operative day 10 was significantly higher than that of controls, accompanied by smaller residual openings and more rapid epithelial regeneration. In the osseous compartment, MaR1 enhanced socket bone fill, attenuated buccal cortical resorption, increased alveolar-ridge width, and reduced vertical bone loss, thereby effectively preserving alveolar architecture.
At the cellular level, MaR1 skewed macrophage polarization toward the M2 phenotype, as evidenced by an elevated M2/M1 ratio, thereby promoting an anti-inflammatory, pro-reparative milieu. Behaviourally, MaR1 alleviated post-operative pain, with treated animals displaying lower pain-related scores as early as one day after extraction. Collectively, topical MaR1 application improves extraction-socket healing by simultaneously accelerating soft-tissue closure, stimulating bone regeneration, modulating macrophage phenotype, and reducing post-operative pain, offering a promising therapeutic strategy for clinical alveolar-ridge preservation and subsequent implant rehabilitation.
Previous work has demonstrated that maresin-1 (MaR1) markedly accelerates tissue regeneration in planarians.13 Following head amputation, exposure to 1–100 nM MaR1 dose-dependently expedites cephalic restoration; pronounced regenerative effects are evident within 3–4 days post-injury, with 100 nM MaR1 producing the most robust response. Mechanistically, injured planarians convert deuterium-labeled DHA into endogenous MaR1, and this biosynthesis is blocked by lipoxygenase (LOX) inhibitors, which concomitantly delay regeneration. Administration of exogenous MaR1 fully rescues the regenerative delay caused by LOX inhibition, confirming an obligate role for MaR1 in the process.
Translational Considerations and Therapeutic Potential
Despite the overwhelming preclinical evidence, the translation of maresins from bench to bedside faces significant hurdles, including pharmacokinetic instability, delivery challenges, and biological variability such as sex differences.
Pharmacokinetics, Stability, and Delivery
Like most lipid mediators, MaR1 has a very short biological half-life, as it is rapidly metabolized and inactivated by enzymatic processes (eg, ω-oxidation and β-oxidation).49 This rapid enzymatic metabolism and short biological half-life mean that simple systemic injection of free MaR1 is unlikely to achieve sustained therapeutic concentrations at the target site (eg, the DRG, spinal cord, or peripheral injured nerves)—which is the single biggest and most fundamental hurdle for MaR1’s clinical translation and application. In addition, the poor water solubility of MaR1 as a lipid mediator further limits its clinical application, as it cannot be formulated into conventional aqueous injections and is difficult to achieve targeted delivery to pain-related tissues and cells. Notably, MaR1’s dose–response curve is bell-shaped,50 which adds another challenge: it exerts optimal analgesic effects only within a narrow dose range, complicating the determination of a safe and effective clinical dosage.
To overcome this, two primary strategies are being explored:
Stable Analogs
Medicinal chemistry efforts have yielded synthetic MaR1 analogs that are resistant to metabolic degradation (eg, modifications at the carboxyl terminus or hydroxyl groups). These stable analogs often retain the biological activity of the parent compound and exhibit superior pharmacokinetic profiles and in vivo efficacy.51 However, potential off-target effects and non-specific receptor binding of MaR1 analogs are key safety concerns that need to be addressed: structural modifications may alter the receptor-binding specificity of MaR1, leading to unintended effects on immune and neural cells and potential side effects. In addition, long-term efficacy and systematic safety evaluation of MaR1 analogs in large animal models (eg, non-human primates) are still lacking, which is a necessary step for clinical translation.
Advanced Delivery Systems
Encapsulating MaR1 within nanocarriers (eg, liposomes, polymeric nanoparticles, or hydrogels) can protect it from degradation, increase its solubility, and provide controlled, localized release at the site of injury or inflammation.52,53 For example, MaR1-loaded hydrogels applied locally to an injured nerve could provide sustained release, promoting regeneration and long-term analgesia.20,54 These delivery systems not only solve the core pharmacokinetic problems of MaR1 (degradation, solubility) but also minimize systemic side effects via local and targeted delivery, which is particularly suitable for the treatment of localized chronic pain (eg, OA, peripheral nerve injury, oral pain, post-surgical pain). However, the feasibility of large-scale industrial production and clinical application of these delivery systems remains to be verified, and the biocompatibility, long-term degradation, and potential immune response of nanocarriers/hydrogels need further systematic evaluation in animal models and human clinical trials.
Sex-Specific Analgesic Effects
The field of pain research has increasingly recognized profound sex dimorphism in both pain mechanisms and analgesic efficacy. SPMs are no exception. The analgesic effect of MaR1 in the SNI model exhibits clear sexual dimorphism.22 MaR1-mediated suppression of spinal gliosis was accompanied by reduced post-operative mechanical hypersensitivity in both sexes, but by diminished pain-related escape/avoidance behaviors (an affective-motivational component of pain) only in male mice. Cytokine profiling revealed that MaR1 did not appreciably alter spinal levels of IL-1-β, IL-6, or IL-10; however, a modest, sex-restricted elevation of macrophage colony-stimulating factor (M-CSF) was observed exclusively in male mice. This suggests that M-CSF may contribute to the male-biased analgesic effect of MaR1, and that different mechanisms may be at play in females. Such findings are critical, as they imply that maresin-based therapies may require sex-specific dosing or combinatorial approaches.
Clinical Evidence and Future Directions
To date, direct clinical trials of MaR1 for chronic pain treatment are lacking, and no clinical data on the safety, efficacy, and optimal dosage of MaR1 in human patients are available. However, correlational human studies strongly support the “resolution failure” hypothesis in chronic pain. Patients with chronic post-surgical pain,10 neuropathic pain,55 and chronic headaches10,56 have been found to have significantly lower circulating or tissue-level concentrations of MaR1 and other SPMs compared to healthy controls. This “SPM deficiency” suggests that replenishing the endogenous pro-resolving milieu—either by administering exogenous MaR1 (or stable analogs) or by providing nutritional precursors like DHA—represents a viable therapeutic strategy.
Based on the current preclinical evidence and existing translational hurdles, the core and actionable future research directions for MaR1-based chronic pain therapy are clearly defined as follows: (1) Validate the druggability of MaR1 receptors: Further confirm the receptor–ligand relationships of GPR37L1/RORA/LGR6 with MaR1 in human neural and immune cells, and develop specific agonists and modulators of these receptors for pain treatment;57,58 (2) Clarify sex-dependent mechanisms: Elucidate the molecular and hormonal mechanisms of the sex-dependent analgesic efficacy of MaR1, and develop sex-specific therapeutic strategies; (3) Conduct rigorous clinical trials: Design and conduct Phase I/II clinical trials of MaR1 analogs/delivery systems in chronic pain patients, with sex stratification and strict evaluation of safety, efficacy, and optimal dosage.
Discussion
Despite critical translational hurdles for MaR1-based therapy—including poor pharmacokinetic stability of endogenous MaR1, sex-dependent efficacy, unclear receptor signaling mechanisms, lack of stable analogs and targeted delivery systems, and absence of human clinical data—MaR1 represents a paradigm shift in pain therapeutics by focusing on active inflammation resolution rather than mere symptomatic blockade. Robust preclinical evidence shows MaR1 acts as a tripartite modulator of persistent pain pathologies: it calms neuroinflammation via glial cell deactivation and macrophage reprogramming (via GPR37L1), normalizes neuronal hyperexcitability by inhibiting nociceptive channels (eg, TRPV1 via RORA), and promotes tissue repair such as peripheral nerve regeneration. These unique pro-resolving, neuroprotective, and regenerative properties make MaR1 a disease-modifying candidate targeting the root cause of chronic pain, with “resolution pharmacology” offering a novel strategy for unmet clinical needs.
The primary translational barrier is MaR1’s poor stability and rapid degradation, so future research should prioritize developing metabolically stable analogs and targeted delivery systems with high bioavailability. Complemented by sex-stratified trials to validate safety and efficacy, and exploration of combination therapies with classic analgesics, MaR1-mediated resolution pathways hold great promise. In summary, MaR1 is a promising next-generation candidate for chronic pain management, and resolution pharmacology based on MaR1 and other SPMs is expected to become a core direction in future pain research and clinical practice, offering new hope to global chronic pain patients.
Data Sharing Statement
All raw data can be obtained by contacting the corresponding author.
Consent for Publication
All listed authors consent to the submission.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The study was supported by the National High Level Hospital Clinical Research Funding (NO.2025-NHLHCRF-PY-08); Elite Medical Professionals Initiative of China–Japan Friendship Hospital (NO.ZRJY2025-QM15); National Key Research and Development Program of China (2022YFC3602201). National High-level Medical Talents Program (2025-YXGCCRC-01).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Volkow ND, McLellan TA. Curtailing diversion and abuse of opioid analgesics without jeopardizing pain treatment. JAMA. 2011;305(13):1346–13. doi:10.1001/jama.2011.369
2. Atkinson TJ, Fudin J. Nonsteroidal Antiinflammatory Drugs for Acute and Chronic Pain. Phys Med Rehabil Clin N Am. 2020;31(2):219–231. doi:10.1016/j.pmr.2020.01.002
3. Serhan CN. Pro-resolving lipid mediators are leads for resolution physiology. Nature. 2014;510(7503):92–101. doi:10.1038/nature13479
4. Ji RR, Xu ZZ, Gao YJ. Emerging targets in neuroinflammation-driven chronic pain. Nat Rev Drug Discov. 2014;13(7):533–548. doi:10.1038/nrd4334
5. Parisien M, Lima LV, Dagostino C, et al. Acute inflammatory response via neutrophil activation protects against the development of chronic pain. Sci Transl Med. 2022;14(644):eabj9954. doi:10.1126/scitranslmed.abj9954
6. Dobson EP, Barrow CJ, Kralovec JA, et al. Controlled formation of mono- and dihydroxy-resolvins from EPA and DHA using soybean 15-lipoxygenase. J Lipid Res. 2013;54(5):1439–1447. doi:10.1194/jlr.M036186
7. Moro K, Nagahashi M, Ramanathan R, et al. Resolvins and omega three polyunsaturated fatty acids: clinical implications in inflammatory diseases and cancer. World J Clin Cases. 2016;4(7):155–164. doi:10.12998/wjcc.v4.i7.155
8. Serhan CN, Yacoubian S, Yang R. Anti-inflammatory and proresolving lipid mediators. Annu Rev Pathol. 2008;3(1):279–312. doi:10.1146/annurev.pathmechdis.3.121806.151409
9. Serhan CN, Chiang N, Dalli J. The resolution code of acute inflammation: novel pro-resolving lipid mediators in resolution. Semin Immunol. 2015;27(3):200–215. doi:10.1016/j.smim.2015.03.004
10. Chávez-Castillo M, Ortega Á, Cudris-Torres L, et al. Specialized Pro-Resolving Lipid Mediators: the Future of Chronic Pain Therapy? Int J Mol Sci. 2021;22(19):10370. doi:10.3390/ijms221910370
11. Serhan CN. Discovery of specialized pro-resolving mediators marks the Dawn of resolution physiology and pharmacology. Mol Aspects Med. 2017;58:1–11. doi:10.1016/j.mam.2017.03.001
12. Serhan CN, Yang R, Martinod K, et al. Maresins: novel macrophage mediators with potent antiinflammatory and proresolving actions. J Exp Med. 2009;206(1):15–23. doi:10.1084/jem.20081880
13. Serhan CN, Dalli J, Karamnov S, et al. Macrophage proresolving mediator maresin 1 stimulates tissue regeneration and controls pain. FASEB J. 2012;26(4):1755–1765. doi:10.1096/fj.11-201442
14. Im DS. Maresin-1 resolution with RORα and LGR6. Prog Lipid Res. 2020;78:101034. doi:10.1016/j.plipres.2020.101034
15. Li T, Zeng H, Xian W, et al. Maresin1 alleviates liver ischemia/reperfusion injury by reducing liver macrophage pyroptosis. J Transl Med. 2023;21(1):472. doi:10.1186/s12967-023-04327-9
16. Bang S, Jiang C, Xu J, et al. Satellite glial GPR37L1 and its ligand maresin 1 regulate potassium channel signaling and pain homeostasis. J Clin Invest. 2024;134(9). doi:10.1172/JCI173537.
17. Costigan M, Scholz J, Woolf CJ. Neuropathic pain: a maladaptive response of the nervous system to damage. Annu Rev Neurosci. 2009;32(1):1–32. doi:10.1146/annurev.neuro.051508.135531
18. Leite CBG, Fricke HP, Tavares LP, et al. Maresin 1-LGR6 axis mitigates inflammation and posttraumatic osteoarthritis after transection of the anterior cruciate ligament in mice. Osteoarthritis Cartilage. 2025;33(7):861–873. doi:10.1016/j.joca.2025.03.005
19. Park CK, Hwang SW. Maresin 1 Inhibits TRPV1 in Temporomandibular Joint-Related Trigeminal Nociceptive Neurons and TMJ Inflammation-Induced Synaptic Plasticity in the Trigeminal Nucleus. Mediators Inflamm. 2015;2015:275126. doi:10.1155/2015/275126
20. Wei J, Su W, Zhao Y, et al. Maresin 1 promotes nerve regeneration and alleviates neuropathic pain after nerve injury. J Neuroinflammation. 2022;19(1):32. doi:10.1186/s12974-022-02405-1
21. Fattori V, Pinho-Ribeiro FA, Staurengo-Ferrari L, et al. The specialised pro-resolving lipid mediator maresin 1 reduces inflammatory pain with a long-lasting analgesic effect. Br J Pharmacol. 2019;176(11):1728–1744. doi:10.1111/bph.14647
22. Teixeira-Santos L, Martins S, Sousa T, et al. The pro-resolving lipid mediator Maresin 1 ameliorates pain responses and neuroinflammation in the spared nerve injury-induced neuropathic pain: a study in male and female mice. PLoS One. 2023;18(6):e0287392. doi:10.1371/journal.pone.0287392
23. Francos-Quijorna I, Santos-Nogueira E, Gronert K, et al. Maresin 1 Promotes Inflammatory Resolution, Neuroprotection, and Functional Neurological Recovery After Spinal Cord Injury. J Neurosci. 2017;37(48):11731–11743. doi:10.1523/JNEUROSCI.1395-17.2017
24. Wang CW, Yu SH, Fretwurst T, et al. Maresin 1 Promotes Wound Healing and Socket Bone Regeneration for Alveolar Ridge Preservation. J Dent Res. 2020;99(8):930–937. doi:10.1177/0022034520917903
25. Shih YV, Tao H, Gilpin A, et al. Specialized pro-resolving mediator Maresin 1 attenuates pain in a mouse model of osteoarthritis. Osteoarthritis Cartilage. 2025;33(3):341–350. doi:10.1016/j.joca.2024.10.018
26. Falsetta ML, Wood RW, Linder MA, et al. Specialized Pro-resolving Mediators Reduce Pro-nociceptive Inflammatory Mediator Production in Models of Localized Provoked Vulvodynia. J Pain. 2021;22(10):1195–1209. doi:10.1016/j.jpain.2021.03.144
27. Wang YH, Li Y, Wang J-N, et al. Maresin 1 Attenuates Radicular Pain Through the Inhibition of NLRP3 Inflammasome-Induced Pyroptosis via NF-κB Signaling. Front Neurosci. 2020;14:831. doi:10.3389/fnins.2020.00831
28. Xu ZZ, Zhang L, Liu T, et al. Resolvins RvE1 and RvD1 attenuate inflammatory pain via central and peripheral actions. Nat Med. 2010;16(5):592–597. doi:10.1038/nm.2123
29. Ji RR, Berta T, Nedergaard M. Glia and pain: is chronic pain a gliopathy? Pain. 2013;154(1):S10–s28. doi:10.1016/j.pain.2013.06.022
30. von Hehn CA, Baron R, Woolf CJ. Deconstructing the neuropathic pain phenotype to reveal neural mechanisms. Neuron. 2012;73(4):638–652. doi:10.1016/j.neuron.2012.02.008
31. Southall MD, Vasko MR. Prostaglandin E(2)-mediated sensitization of rat sensory neurons is not altered by nerve growth factor. Neurosci Lett. 2000;287(1):33–36. doi:10.1016/S0304-3940(00)01158-7
32. Zhang L, Terrando N, Xu -Z-Z, et al. Distinct Analgesic Actions of DHA and DHA-Derived Specialized Pro-Resolving Mediators on Post-operative Pain After Bone Fracture in Mice. Front Pharmacol. 2018;9:412. doi:10.3389/fphar.2018.00412
33. Serhan CN. Systems approach to inflammation resolution: identification of novel anti-inflammatory and pro-resolving mediators. J Thromb Haemost. 2009;7(1):44–48. doi:10.1111/j.1538-7836.2009.03396.x
34. Moulin D, Sellam J, Berenbaum F, et al. The role of the immune system in osteoarthritis: mechanisms, challenges and future directions. Nat Rev Rheumatol. 2025;21(4):221–236. doi:10.1038/s41584-025-01223-y
35. Largent-Milnes TM, Hegarty DM, Aicher SA, et al. Physiological temperatures drive glutamate release onto trigeminal superficial dorsal horn neurons. J Neurophysiol. 2014;111(11):2222–2231. doi:10.1152/jn.00912.2013
36. Ji RR, Nackley A, Huh Y, et al. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology. 2018;129(2):343–366. doi:10.1097/ALN.0000000000002130
37. Inoue K, Tsuda M. Microglia in neuropathic pain: cellular and molecular mechanisms and therapeutic potential. Nat Rev Neurosci. 2018;19(3):138–152. doi:10.1038/nrn.2018.2
38. Serhan CN. Novel omega -- 3-derived local mediators in anti-inflammation and resolution. Pharmacol Ther. 2005;105(1):7–21. doi:10.1016/j.pharmthera.2004.09.002
39. Scholz J, Woolf CJ. The neuropathic pain triad: neurons, immune cells and glia. Nat Neurosci. 2007;10(11):1361–1368. doi:10.1038/nn1992
40. Zhang ZJ, Cao D-L, Zhang X, et al. Chemokine contribution to neuropathic pain: respective induction of CXCL1 and CXCR2 in spinal cord astrocytes and neurons. Pain. 2013;154(10):2185–2197. doi:10.1016/j.pain.2013.07.002
41. Xu J, Yan Z, Bang S, et al. GPR37L1 identifies spinal cord astrocytes and protects neuropathic pain after nerve injury. Neuron. 2025;113(8):1206–1222.e6. doi:10.1016/j.neuron.2025.01.012
42. Allen BL, Montague-Cardoso K, Simeoli R, et al. Imbalance of proresolving lipid mediators in persistent allodynia dissociated from signs of clinical arthritis. Pain. 2020;161(9):2155–2166. doi:10.1097/j.pain.0000000000001908
43. O’Dowd BF, Heiber M, Chan A, et al. A human gene that shows identity with the gene encoding the angiotensin receptor is located on chromosome 11. Gene. 1993;136(1–2):355–360. doi:10.1016/0378-1119(93)90495-O
44. Meyer RC, Giddens MM, Schaefer SA, et al. GPR37 and GPR37L1 are receptors for the neuroprotective and glioprotective factors prosaptide and prosaposin. Proc Natl Acad Sci U S A. 2013;110(23):9529–9534. doi:10.1073/pnas.1219004110
45. Meyer RC, Giddens MM, Coleman BM, et al. The protective role of prosaposin and its receptors in the nervous system. Brain Res. 2014;1585:1–12. doi:10.1016/j.brainres.2014.08.022
46. Shinotsuka N, Denk F. Fibroblasts: the neglected cell type in peripheral sensitisation and chronic pain? A review based on a systematic search of the literature. BMJ Open Sci. 2022;6(1):e100235. doi:10.1136/bmjos-2021-100235
47. Szallasi A, Blumberg PM. Vanilloid (Capsaicin) receptors and mechanisms. Pharmacol Rev. 1999;51(2):159–212.
48. Li L, Ru Q, Lu Y, et al. Tiam1 coordinates synaptic structural and functional plasticity underpinning the pathophysiology of neuropathic pain. Neuron. 2023;111(13):2038–2050.e6. doi:10.1016/j.neuron.2023.04.010
49. Colas RA, Dalli J, Chiang N, et al. Identification and Actions of the Maresin 1 Metabolome in Infectious Inflammation. J Immunol. 2016;197(11):4444–4452. doi:10.4049/jimmunol.1600837
50. Oh SF, Pillai PS, Recchiuti A, et al. Pro-resolving actions and stereoselective biosynthesis of 18S E-series resolvins in human leukocytes and murine inflammation. J Clin Invest. 2011;121(2):569–581. doi:10.1172/JCI42545
51. Byrne L, Guiry PJ. Advances in the Chemistry and Biology of Specialised Pro-Resolving Mediators (SPMs). Molecules. 2024;29(10):2233. doi:10.3390/molecules29102233
52. Sharma A, Sharma D, Panda PK, et al. Solid Lipid Nanoparticles and Nanostructured Lipid Particles: a Comparative Review on Lipid-Based Nanocarriers. Pharm Nanotechnol. 2024;13. doi:10.2174/0122117385318686241003103005
53. Sousa AB, Martins C, Sarmento B, et al. Zein nanocarriers for controlled maresin-1 delivery: a novel approach in biomaterial-based immunomodulation. Biomater Adv. 2025;172:214238. doi:10.1016/j.bioadv.2025.214238
54. Zeng J, Su P, Li F, et al. An Injectable Hydrogel for Treatment of Chronic Neuropathic Pain. Macromol Biosci. 2022;22(6):e2100529. doi:10.1002/mabi.202100529
55. Sánchez-Fernández A, Zandee S, Mastrogiovanni M, et al. Administration of Maresin-1 ameliorates the physiopathology of experimental autoimmune encephalomyelitis. J Neuroinflammation. 2022;19(1):27. doi:10.1186/s12974-022-02386-1
56. Chen TB, Yang -C-C, Tsai I-J, et al. Neuroimmunological effects of omega-3 fatty acids on migraine: a review. Front Neurol. 2024;15:1366372. doi:10.3389/fneur.2024.1366372
57. Serhan CN. Treating inflammation and infection in the 21st century: new hints from decoding resolution mediators and mechanisms. FASEB J. 2017;31(4):1273–1288. doi:10.1096/fj.201601222R
58. Dalli J, Serhan CN. Specific lipid mediator signatures of human phagocytes: microparticles stimulate macrophage efferocytosis and pro-resolving mediators. Blood. 2012;120(15):e60–e72. doi:10.1182/blood-2012-04-423525
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.