Back to Journals » Drug Design, Development and Therapy » Volume 20
Beyond Inhibition: Rebalancing the Hsp90 Chaperone Network as a Therapeutic Strategy for Alzheimer’s Disease
Authors Tang Z, Xie H, Xu Y, Zhou Z, Lu F, Wu Q, Liang W ![]() , Zhong M, Li S
, Zhong M, Li S ![]()
Received 9 February 2026
Accepted for publication 20 May 2026
Published 26 May 2026 Volume 2026:20 602287
DOI https://doi.org/10.2147/DDDT.S602287
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Manfred Ogris
Zhikang Tang,1,* Haiyu Xie,1,2,* Yongxing Xu,1 Zhenhong Zhou,1 Feng Lu,1,2 Qiaozhen Wu,1,2 Weidong Liang,1,2 Maolin Zhong,1,2 Shihong Li1,2
1The First Clinical Medical College of Gannan Medical University, Ganzhou, Jiangxi, People’s Republic of China; 2Department of Anesthesiology, Ganzhou Key Laboratory of Anesthesiology, The First Affiliated Hospital of Gannan Medical University, Ganzhou, Jiangxi, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Maolin Zhong, Email [email protected] Shihong Li, Email [email protected]
Abstract: Alzheimer’s disease (AD) is characterized by the pathological accumulation of misfolded amyloid-β (Aβ) and tau, driven by dysfunction of the proteostasis network in which the molecular chaperone heat shock protein 90 (Hsp90) is a central regulator. Here, we propose a shift from pan-inhibition of Hsp90 to rebalancing: selectively normalizing pathological co-chaperone assemblies while preserving homeostatic chaperone functions. We first summarize how specific co-chaperones (eg, FK506-binding protein 51 (FKBP51), activator of Hsp90 ATPase homolog 1 (Aha1), cell division cycle 37(Cdc37)) shift Hsp90 toward tau stabilization, whereas others (eg, C-terminus of Hsc70-interacting protein (CHIP), FK506-binding protein 52 (FKBP52)) promote tau clearance, and we outline Hsp90’s context-dependent effects on Aβ. We then trace the evolution of therapeutic strategies from N-terminal ATPase inhibitors, which have shown limited clinical efficacy, to precision approaches including allosteric C-terminal modulators, co-chaperone-selective protein–protein interaction (PPI) inhibitors, induction-based microglial clearance strategies, and epichaperome disruptors. We conclude by critically discussing translational challenges, including blood-brain barrier (BBB) penetration, isoform selectivity, biomarker development, and long-term safety, noting that most current evidence remains preclinical. Rebalancing the Hsp90 chaperone network, rather than inhibiting it indiscriminately, offers a mechanistically grounded yet experimentally early avenue to disease-modifying therapy for AD.
Keywords: heat shock protein 90, Alzheimer’s disease, tauopathy, proteostasis, molecular chaperones
Introduction
Alzheimer’s disease (AD), the most common form of dementia, is characterized by a progressive decline in cognitive function, memory, and personal identity.1 At the molecular level, AD represents a disorder of protein misfolding, a profound disruption of cellular protein homeostasis, or proteostasis.2 The neuropathological hallmarks of AD include two principal lesions: the extracellular deposition of amyloid-β (Aβ) peptides into senile plaques and the intracellular aggregation of hyperphosphorylated tau protein into neurofibrillary tangles.3,4 These toxic protein aggregates disrupt synaptic function, provoke neuroinflammation, and trigger downstream pathological processes that ultimately lead to neuronal loss.1,5 Despite decades of research, therapeutic interventions have shown limited success, with recent Aβ-targeting antibodies yielding modest clinical efficacy and significant side effects. These limitations highlight the need for novel therapeutic strategies that address the underlying cellular dysfunction driving the disease.6 Increasingly, research has shifted focus to the cellular machinery responsible for maintaining proteostasis, with molecular chaperones emerging as critical regulators and promising therapeutic targets.
At the core of the cellular proteostasis network lies heat shock protein 90 (Hsp90), a highly conserved, ATP-dependent molecular chaperone.7,8 Rather than functioning solely as a housekeeping protein, Hsp90 regulates the folding, stability, activation, and degradation of a diverse set of client proteins.9 These clients are predominantly metastable signaling proteins, including kinases, transcription factors, and steroid hormone receptors, which are essential for cellular growth, differentiation, and stress responses.5,10 Hsp90’s activity is powered by an ATP-driven conformational cycle and is tightly regulated by an array of over 20 co-chaperones.11,12 These co-chaperones modulate client specificity, regulate ATPase activity, and link Hsp90 to downstream pathways such as proteasomal degradation, forming a highly dynamic and adaptable chaperone network.13 Given its central role in regulating proteins involved in pathological signaling, Hsp90 has emerged as a key node in disease biology, particularly in oncology and neurodegeneration.14,15
The initial therapeutic approach to targeting Hsp90 in disease, largely inspired by its success in oncology, focused on inhibition.15,16 Small molecules such as geldanamycin and its derivatives block the N‑terminal ATP‑binding pocket to promote degradation of Hsp90‑dependent clients, including oncogenic drivers.17 This concept was extended to AD with the aim of reducing pathogenic tau and the kinases that promote its hyperphosphorylation.18,19 However, broad Hsp90 inhibition risks disrupting essential cellular functions and can trigger a detrimental heat shock response.12 These limitations have driven a paradigm shift, supported by a deeper understanding of the Hsp90 machinery. Recent efforts have moved beyond inhibition toward strategies that rebalance the chaperone system. Current approaches include allosteric modulators that fine‑tune Hsp90 activity (eg, LA1011),20,21 precision inhibitors targeting specific protein-protein interactions (PPIs) with pathological co-chaperones such as FK506-binding protein 51 (FKBP51),6,22,23 and strategies that induce Hsp90 expression in a context-dependent manner to enhance the clearance of toxic proteins.24,25
The conceptual novelty of this review is to reframe Hsp90-targeted therapy for AD as a rebalancing strategy, shifting from indiscriminate inhibition to selective normalization of pathological co-chaperone assemblies while preserving homeostatic chaperone functions. Rather than cataloging Hsp90 inhibitors or co-chaperone biology in isolation, we integrate tauopathy, amyloidopathy, epichaperome biology, and emerging drug-design paradigms within a single unifying framework, and we critically evaluate each strategy against AD-relevant development criteria BBB penetration, isoform selectivity, biomarker translatability, and long-term safety.
Literature Search Strategy
This narrative review synthesizes mechanistic, preclinical, and early clinical literature on Hsp90 in AD. Primary studies were identified via PubMed, Web of Science, and Scopus (coverage through December 2025) using combinations of the terms “Hsp90”, “HSP90AA1”, “HSP90AB1”, “TRAP1”, “GRP94”, “epichaperome”, “co-chaperone”, “FKBP51”, “FKBP52”, “CHIP/STUB1”, “Aha1”, “Cdc37”, and “Alzheimer”, “tau”, “amyloid-β”, and “neurodegeneration”. We prioritized mechanistic and structural studies, seminal primary reports, and publications from the past five years; older foundational references were retained for historical context. ClinicalTrials.gov was queried for active and completed trials of Hsp90/epichaperome-directed agents. As a narrative (rather than systematic) review, the synthesis emphasizes conceptual framing over exhaustive enumeration; where opinion or hypothesis predominates over validated data, we indicate this explicitly in the text.
The Double-Edged Sword: Hsp90’s Role in AD Pathogenesis
Hsp90 plays a central yet paradoxical role in AD pathogenesis. Rather than acting as a simple accomplice, Hsp90 functions as a master regulator, exerting a profound and dichotomous influence on the two hallmark pathologies of AD: tauopathies and amyloidopathies. Hsp90 operates at the intersection of these pathological processes, acting both as a potential driver of neurodegeneration and as a component of the cell’s defense system. Its function is highly dependent on the cellular context, particularly the cohort of co-chaperones with which it partners. This intricate network of interactions ultimately determines whether Hsp90 promotes the degradation of pathogenic proteins or, conversely, facilitates their stabilization in toxic forms. Understanding this duality is crucial for the development of effective therapeutic strategies (Figure 1).
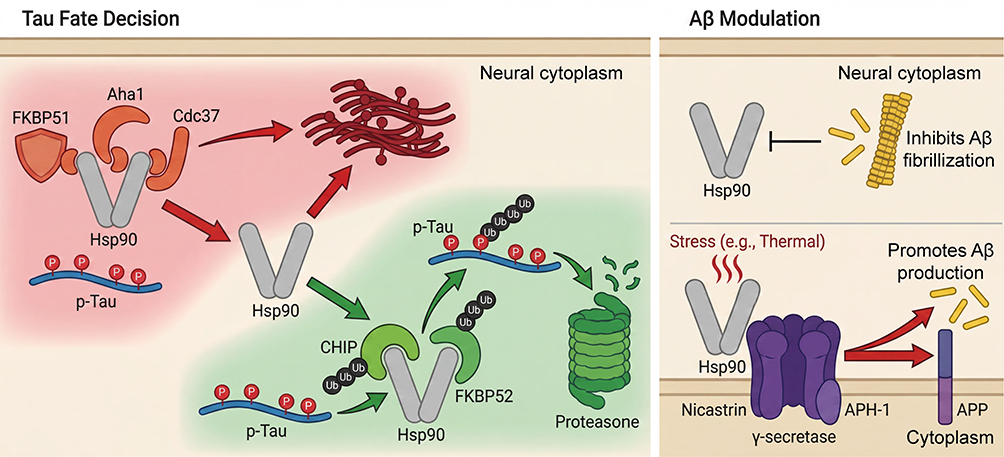 |
Figure 1 Dichotomous role of the Hsp90 chaperone machinery in Alzheimer’s disease pathology. Tau Fate Decision: Hsp90 functions as a central regulatory hub for Tau. Association with pro-pathogenic co-chaperones FKBP51, Aha1, or Cdc37 (red pathway) stabilizes hyperphosphorylated Tau (p-Tau) and promotes its oligomerization. In contrast, recruitment of protective co-chaperones CHIP or FKBP52 (green pathway) targets p-Tau for ubiquitination and proteasomal degradation. Aβ Modulation: Hsp90 exert context-dependent effects on Aβ. Under physiological conditions, Hsp90 can inhibit Aβ fibrillization. Under stress (eg, thermal stress), Hsp90 facilitates assembly of the γ-secretase complex (via Nicastrin and APH-1), thereby enhancing APP cleavage and Aβ production. Abbreviations: Aβ, amyloid-β; Aha1, activator of Hsp90 ATPase homolog 1; APH-1, anterior pharynx-defective 1; APP, amyloid precursor protein; Cdc37, cell division cycle 37; CHIP, C-terminus of Hsc70-interacting protein; FKBP51, FK506-binding protein 51; FKBP52, FK506-binding protein 52; Hsp90, heat shock protein 90; p-tau, hyperphosphorylated tau. |
The apparently contradictory roles of Hsp90 in AD are best reconciled by recognizing that its function is contextual. Disease stage appears critical: in PS19 tauopathy models, epichaperome formation precedes overt tau pathology, emerging in the hippocampus and amygdala by 3 months and progressing to widespread distribution with increasing intensity by 11 months.26 Brain-region specificity is evident in AD-vulnerable areas, where there is preferential loss of Hsp90 binding alongside epichaperome enrichment; correspondingly, [11C]HSP990 positron emission tomography (PET) tracer binding is significantly reduced in hippocampal gray matter from AD patients that exhibits tau pathology.27 Cell-type also matters. In neurons, Hsp90/FKBP51 complexes stabilize toxic tau species, whereas in microglia, Hsp90-dependent pathways (eg, Axl/HSP90β/PPARγ) promote Aβ phagocytosis and clearance, a distinction central to the rationale for induction-based strategies25 Moreover, Hsp90 intersects with neuroinflammation and proteostasis collapse: FKBP51 is upregulated by damage- and inflammation-associated signaling and exacerbates tau pathology, while age-related shifts in chaperone/co-chaperone stoichiometry favor epichaperome assembly and pathogenic rewiring of the interactome, motivating small molecules approaches to restore balances.27 Together, these contextual axes, stage, region, cell type, and the proteostatic/inflammatory milieu determine whether Hsp90 acts as a guardian or accomplice and directly inform the design of stage- and compartment-selective interventions.
Hsp90 and the Fate of Tau: A Co-Chaperone-Dictated Balance
One of the most well-documented roles of Hsp90 in AD is its regulation of the microtubule-associated protein tau. In healthy neurons, Hsp90 and its co-chaperones maintain tau homeostasis, ensuring proper folding and preventing aggregation.2 However, in the diseased state, this regulation becomes profoundly dysfunctional, transforming Hsp90’s influence from protective to pathological. The fate of tau whether it is refolded, cleared, or driven into aggregation is dictated by the specific co-chaperones recruited into the Hsp90 machinery. In the AD brain, this delicate balance is catastrophically disrupted.13,28
Stabilizing Pathological Tau: The Role of FKBP51, Aha1, and Cdc37
A growing body of evidence highlights a subset of Hsp90 co-chaperones that actively promote tau pathology. Foremost among these is FKBP51, the levels of which are significantly elevated in the aging and AD brain.6,29,30 Research by Oroz et al demonstrated that the Hsp90/FKBP51 complex acts as a pro-toxic scaffold, stabilizing tau in conformations prone to aggregation.31 This interaction prevents tau clearance and fosters the accumulation of neurotoxic oligomeric tau species.29 Jinwal et al further showed that FKBP51 directly increases tau stability, with recent structural studies elucidating the molecular mechanisms behind this detrimental complex.6,30
Another key player is the activator of Hsp90 ATPase homolog 1 (Aha1), which accelerates Hsp90’s ATPase activity, paradoxically increasing the production of pathological tau. Shelton et al provided compelling evidence that Aha1 drives the accumulation of insoluble, aggregated tau, with its expression correlating with AD progression.32 Criado-Marrero et al extended these findings, showing that Aha1 overexpression in the hippocampus of aged wild-type mice leads to tau phosphorylation and cognitive deficit.33 Similarly, the kinase-recruiting co-chaperone cell division cycle 37 (Cdc37) indirectly promotes tau pathology by stabilizing tau kinases, thereby enhancing tau hyperphosphorylation. By partnering with Hsp90, Cdc37 contributes to the maintenance and pathological modification of tau.10,21 Collectively, these “pro-pathology” co-chaperones create a cellular environment where Hsp90, instead of mitigating tauopathy, facilitates the stabilization and accumulation of toxic tau species.
Promoting Tau Degradation: The Protective Functions of CHIP and FKBP52
In contrast to the deleterious effects of FKBP51 and Aha1, a separate group of co-chaperones directs Hsp90 toward a protective role, promoting the degradation of pathological tau. Chief among these is the C-terminus of Hsc70-interacting protein (CHIP), an E3 ubiquitin ligase that partners with Hsp90 to target hyperphosphorylated tau for degradation. Dickey et al demonstrated that the Hsp90-CHIP complex selectively recognizes aberrant tau, ubiquitinating it for proteasomal clearance.18 This quality control mechanism serves as a critical defense against tau accumulation. Salminen et al further underscored the importance of Hsp90-CHIP interactions in regulating tau pathology, highlighting their disruption as a contributing factor in AD progression.13
Complementing CHIP’s protective role is FK506-binding protein 52 (FKBP52), a homolog of FKBP51 with opposing effects on tau. Unlike FKBP51, which is upregulated in AD, FKBP52 levels are often reduced.11 FKBP52 competes with FKBP51 for binding to Hsp90, and its presence is associated with reduced tau aggregation and the preservation of physiological tau states.28 The imbalance between elevated FKBP51 and diminished FKBP52 levels in the aging brain represents a crucial tipping point, shifting Hsp90’s activity from promoting tau clearance to facilitating its toxic aggregation.6,28
It is essential to consider Hsp90 co-chaperone dysregulation within the broader, age-related collapse of the proteostasis network.34 Aging drives a systemic decline in chaperone capacity, the ubiquitin-proteasome system, and autophagy, collectively diminishing the cell’s ability to manage misfolded proteins. The imbalances described above including the upregulation of FKBP51 and Aha1 with concomitant downregulation of protective factors such as cyclophilin 40 and protein phosphatase 5 should therefore be viewed as a critical facet of this larger proteostatic deterioration. As the network weakens, cells become increasingly reliant on residual chaperone capacity, rendering the specific composition of the Hsp90 co-chaperone ensemble even more consequential for protein fate decisions.
Hsp90’s Complex Relationship with Aβ
Hsp90’s involvement in tau pathology is primarily mediated through its co-chaperone network, but its relationship with Aβ is significantly more intricate. This complexity arises from Hsp90’s direct and indirect roles in Aβ production, aggregation, and clearance. The literature presents a dichotomy, wherein Hsp90 is portrayed as either a protective factor or a contributor to amyloid pathology, depending on the specific cellular context and mechanisms under investigation.
This dual role is exemplified by contrasting lines of evidence. On one hand, Hsp90 has shown a distinct ability to suppress Aβ fibrillization. In vitro studies, as reviewed by Iqbal et al, demonstrate that Hsp90 interacts with Aβ monomers, inhibiting their aggregation into mature fibrils and stabilizing Aβ in a less toxic, monomeric state.35,36 This activity highlights Hsp90’s direct anti-aggregation function. Moreover, Hsp90 activation has been linked to enhanced Aβ clearance. For instance, Zhang et al revealed that activation of the Axl/HSP90/PPARγ pathway in microglia stimulates the phagocytosis and degradation of Aβ, thereby reducing amyloid burden.25 In this context, Hsp90 operates as a key player in the brain’s innate immune response to amyloid plaques.
Conversely, other studies suggest a pathogenic role for Hsp90 in Aβ metabolism. A notable study by Noorani et al demonstrated that under conditions of thermal stress relevant given the associations between sleep disruption, elevated core body temperature, and AD Hsp90 facilitates the assembly and activation of the γ-secretase complex.37 Specifically, Hsp90 binds to immature components of the γ-secretase complex, APH-1 and Nicastrin, promoting the maturation of the enzyme responsible for the final cleavage of amyloid precursor protein to produce Aβ peptides. Remarkably, Hsp90 knockdown completely abolished the thermal stress-induced increase in Aβ production, identifying Hsp90 as a positive regulator of amyloidogenesis under these conditions. These findings raise the possibility that global inhibition of Hsp90 could mitigate Aβ production. Further adding to the complexity, Hsp90 has been implicated in Aβ-induced vascular pathology. By interacting with endothelial nitric oxide synthase, Hsp90 contributes to impaired nitric oxide production and endothelial dysfunction, which are hallmarks of Aβ-related vascular damage.38 It is important to note, however, that much of the evidence for Hsp90’s protective effects on Aβ (eg, direct suppression of fibrillization) arises from in vitro or cell-based systems, whereas reported pathogenic contributions (eg, γ-secretase maturation, endothelial dysfunction) have been demonstrated under specific stress paradigms Caution is therefore warranted when extrapolating these mechanistic observations to the chronic human AD brain, and direct evidence in disease-relevant human tissue remains limited. Collectively, these findings underscore the multifaceted and context-dependent roles of Hsp90 in Aβ pathology, ranging from protective chaperone-mediated functions to its involvement in pathogenic mechanisms.
A New Layer of Complexity: The “Epichaperome” as a Pathological Scaffold in Neurodegeneration
Recent advances by Chiosis et al have introduced the concept of the “epichaperome,” a paradigm-shifting hypothesis that adds a crucial layer of complexity to the role of Hsp90 in chronic diseases such as cancer and neurodegeneration.8 This theory postulates that Hsp90 undergoes a structural and functional transformation in disease states. Rather than existing as dynamic, transient dimers, Hsp90 assembles into stable, high-molecular-weight hetero-oligomeric complexes, termed epichaperomes. These complexes represent a distinct pathological entity, characterized by a gain-of-function mechanism that diverges from normal chaperone activity.
In the context of neurodegeneration, the epichaperome is hypothesized to act as a maladaptive scaffolding platform. Unlike the typical foldase activity of Hsp90, the epichaperome functions as a “holdase,” aberrantly sequestering and organizing already folded proteins. This pathological scaffolding rewires cellular PPI networks, triggering widespread dysfunction.8 In AD models, the formation of epichaperomes disrupts the cellular interactome, with over a thousand proteins losing their normal interaction partners while simultaneously forming new, non-native interactions. This extensive rewiring perturbs critical pathways involved in synaptic plasticity, brain energetics, and inflammatory signaling.8
The epichaperome concept provides a compelling framework for understanding how dysfunction in a single chaperone can produce pleiotropic and devastating effects across the cellular landscape in AD. Importantly, it suggests that therapeutic strategies should go beyond the inhibition of Hsp90’s ATPase activity to specifically target and dismantle these pathological, high-order epichaperome structures. This novel approach holds significant promise for drug discovery, offering a potential means to correct the proteostasis imbalance that underlies neurodegenerative diseases (Figure 2). Although the epichaperome offers a compelling unifying framework, it remains a relatively recent concept whose clinical validation is still in progress.8 Independent replication across laboratories, standardized biochemical criteria for epichaperome identification, and prospective clinical data (eg, from the ongoing PU-AD Phase 2 trial)39 will be required to establish the epichaperome as a clinically actionable entity rather than primarily an experimental construct.
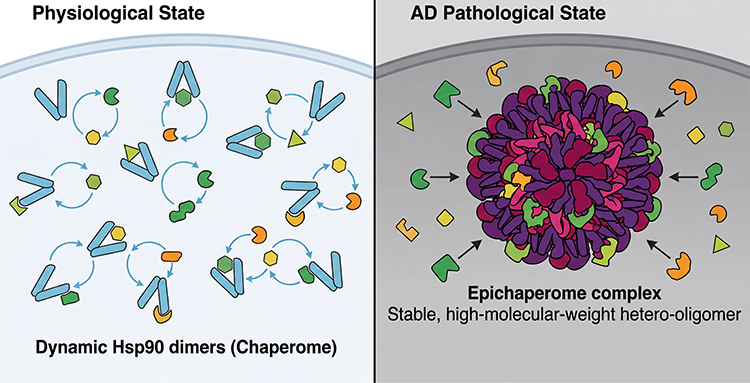 |
Figure 2 The epichaperome as a pathological scaffold that rewires the cellular interactome. Physiological State: In healthy neurons, Hsp90 exists predominantly as dynamic dimers (the Chaperome) that transiently interact with clients to facilitate folding, thereby maintaining a flexible and functional proteome. AD Pathological State: Under chronic stress and neurodegenerative conditions, Hsp90 assembles into stable, high-molecular-weight hetero-oligomeric complexes termed “Epichaperomes.” These structures act as maladaptive scaffolds (“holdases”), aberrantly sequestering folded proteins and rewiring PPI networks, leading to synaptic dysfunction and metabolic failure. The “epichaperome” model depicted here is a hypothesis-generating framework primarily advanced by Chiosis and Neckers. Although supported by chemical biology and proteomics data in AD mouse models, the precise composition and stoichiometry of epichaperomes in the human AD brain remain to be fully validated. The transition from the dynamic chaperome to stable epichaperome is shown schematically and is based on native PAGE analyses and chemical pulldown experiments. Abbreviations: AD, Alzheimer’s disease; Hsp90, heat shock protein 90; PPI, protein–protein interaction. |
The Evolution of Hsp90-Targeted Therapeutic Strategies for AD
The recognition of Hsp90’s multifaceted and often contradictory roles in AD pathology has necessitated an evolution in therapeutic strategies. The development of Hsp90-targeted drugs for AD represents a remarkable journey of scientific refinement, transitioning from broad-spectrum, oncology-inspired approaches to more sophisticated, mechanism-driven interventions. This progression reflects a deepening understanding of Hsp90’s chaperone machinery and a paradigm shift from indiscriminate inhibition to precise, context-sensitive modulation (Figure 3).
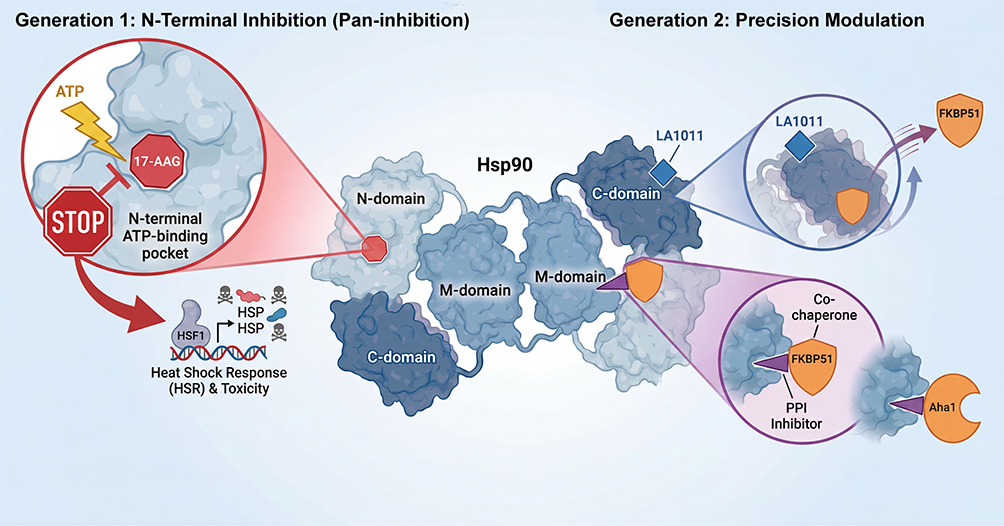 |
Figure 3 Evolution of Hsp90-targeted therapeutic strategies: from pan-inhibition to precision rebalancing. Generation 1: N-terminal inhibition (Pan-inhibition). Agents such as 17-AAG competitively bind the N-terminal ATP pocket, blocking ATP hydrolysis. Although this promotes degradation of pathogenic clients (eg, p-Tau), it indiscriminately inhibits all Hsp90 functions, triggering the HSR and causing on-target toxicity Generation 2: Precision modulation. (1) Allosteric Modulation (eg, LA1011): binds the C-terminal domain, inducing a conformational change that selectively displaces the pro-toxic co-chaperone FKBP51 while sparing other interactions (2) PPI inhibition: Small molecules designed to block the Hsp90 co-chaperones interface (eg, Hsp90-FKBP51 or Hsp90-Aha1 inhibitors), preventing formation of pathogenic complexes without abolishing Hsp90 ATPase activity. Status and evidence: Generation 1 (17-AAG): mechanism well-established; preclinical efficacy demonstrated; clinical development for AD limited by pharmacokinetic challenges. Generation 2 – Allosteric Modulation (LA1011): structural basis for FKBP51 displacement confirmed by X-ray crystallography; preclinical efficacy demonstrated in the APPxPS1 mouse model; clinical development is ongoing. Generation 2 – PPI inhibition: proof of concept demonstrated in cellular assays; in vivo validation and translational studies remain at early stages. Abbreviations: 17-AAG, 17-allylamino-17-demethoxygeldanamycin; Aha1, activator of Hsp90 ATPase homolog 1; ATP, adenosine triphosphate; FKBP51, FK506-binding protein 51; Hsp90, heat shock protein 90; HSR, heat shock response; PPI, protein–protein interaction; p-tau, hyperphosphorylated tau. |
First Generation: Pan-Inhibition of the Hsp90 ATPase
The initial efforts to target Hsp90 in neurodegeneration were inspired by its success in oncology, where Hsp90 inhibitors demonstrated significant therapeutic potential.15,16 The underlying rationale was straightforward: cancer cells are highly reliant on Hsp90 to stabilize and buffer the stress of mutated and overexpressed oncoproteins. Inhibiting Hsp90’s ATPase function in these cells induces degradation of its client proteins, leading to cellular apoptosis. Translating this concept to AD, researchers hypothesized that Hsp90 inhibition could similarly destabilize and promote the clearance of pathogenic clients, such as hyperphosphorylated tau and the kinases responsible for its abnormal modification.17,40
Mechanism of N-Terminal Domain Inhibitors
First-generation Hsp90 inhibitors, including the benzoquinone ansamycin geldanamycin and its less toxic derivative 17-allylamino-17-demethoxygeldanamycin (17-AAG), function by competitively binding to the conserved N-terminal ATP-binding pocket of Hsp90.9 This binding locks the chaperone in a non-productive conformation, thereby preventing ATP hydrolysis, which is essential for driving Hsp90’s conformational cycle. Without ATPase activity, Hsp90 is unable to stabilize its client proteins, resulting in their release, ubiquitination via ligases such as CHIP, and subsequent degradation via the proteasome.13,18 Seminal studies by Luo et al and Dickey et al provided critical proof-of-concept for this approach, demonstrating that N-terminal domain (NTD) inhibitors could effectively reduce both soluble and insoluble phosphorylated tau species in cellular and animal models of tauopathy.17,18 This strategy offered a dual advantage: it not only facilitated the clearance of toxic tau but also triggered a beneficial heat shock response. Inhibition of Hsp90 activates Heat Shock Factor 1 (HSF1), leading to the upregulation of protective chaperones, such as Hsp70, that further mitigate proteotoxic stress.14,41
Preclinical Success and Inherent Limitations
Despite promising preclinical results in clearing pathological proteins, the development of pan-Hsp90 inhibitors for clinical use in AD has faced substantial challenges. A significant limitation of early-generation compounds was their poor pharmacological properties, including low solubility, metabolic instability, and, most crucially, limited penetration of the blood-brain barrier (BBB), which restricted their access to the central nervous system (CNS).12,41 While later-generation inhibitors, such as OS47720, were specifically designed to improve CNS permeability and demonstrated efficacy in rescuing memory deficits in AD mouse models,41 fundamental issues with the pan-inhibition strategy persisted. The very mechanism of pan-inhibition blocking the function of a ubiquitous and essential cellular chaperone carries a significant risk of on-target toxicity.9
The key limitation lies in the broad, indiscriminate mechanism of these inhibitors. Hsp90 is a ubiquitous and essential chaperone with a clientele of over 400 proteins, many of which are critical for maintaining cellular homeostasis. These inherent limitations underscored the need for a more nuanced and selective approach to Hsp90-targeted therapy, driving the field toward the development of second-generation, mechanism-specific strategies.
The translational failure of first-generation N-terminal Hsp90 inhibitors in AD can be attributed to three convergent factors: (i) limited CNS exposure due to efflux transport and suboptimal physicochemical properties (eg, 17-AAG), (ii) on-target toxicity from discriminate inhibition of homeostatic Hsp90 in non-neuronal tissues (hepatotoxicity, ocular and cardiac effects observed in oncology programs) compounded by poor isoform selectivity and induction of the heat‑shock response;42 and (iii) robust HSF1 activation that rewires proteostasis in a cytoprotective manner, potentially blunting pro-degradation efforts Notably, to date only pimitespib has received regulatory approval (for gastrointestinal stromal tumors), agents such as ganetespib and onalespib have yielded variable outcomes in clinical trials, and no N‑terminal Hsp90 inhibitor has progressed beyond early clinical evaluation in AD.42
The New Wave: Precision Modulation of the Hsp90 Machinery
In response to the limitations of pan-inhibition, recent research has shifted toward a new wave of therapeutic design centered on precision and subtlety. Rather than globally silencing the Hsp90 system, these next-generation strategies aim to selectively modulate its pathological activities while preserving its essential physiological functions. This is achieved by targeting alternative sites outside the N-terminal ATP pocket, enabling a more balanced approach to reconfiguring the chaperone machinery.
Allosteric Modulation: Rebalancing the System without Full Inhibition
A key example of this refined approach lies in the development of allosteric modulators, which do not directly inhibit Hsp90’s ATPase activity but instead influence its interactions with co-chaperones. The dihydropyridine derivative LA1011 exemplifies this strategy. Structural studies by Roe et al revealed that LA1011 binds to a distinct hydrophobic pocket at the C-terminus of the Hsp90 dimer.20 This binding site overlaps with the interaction region for the tetratricopeptide repeat (TPR) domain of FKBP51, a pro-toxic co-chaperone implicated in tau pathology. By competitively displacing FKBP51 from the Hsp90 complex, LA1011 effectively disrupts the pathological Hsp90-FKBP51 interaction, thereby “rebalancing” the chaperone system toward a more protective state.6,20 This elegant mechanism, which modulates Hsp90 function allosterically rather than shutting it down entirely, has shown promising preclinical efficacy. LA1011 treatment significantly reduced both tau and Aβ pathology and prevented neurodegeneration, suggesting potentially transformative directions, contingent on clinical validation.6
The principal advantage of allosteric C-terminal modulation over N-terminal inhibition is preservation of baseline ATPase activity and HSF1 homeostasis, which may widen the therapeutic window. LA1011 is the most advanced exemplar but remain preclinical; its developers explicitly acknowledged that, because numerous co-chaperones and clients engage Hsp90, adverse effects from broader perturbation cannot be excluded and are under active evaluation across a range of Hsp90 complexes.6
Targeting Protein-Protein Interactions (PPIs): A More Specific Approach
Building on the principle of disrupting specific chaperone complexes, a more refined approach involves the rational design of small molecules that directly target PPI interfaces between Hsp90 and its pathogenic partners. This strategy offers unparalleled specificity by selectively severing pathological interactions within the Hsp90 interactome. A notable breakthrough by Wang et al utilized structure-based virtual screening to identify highly potent and selective small-molecule inhibitors of the FKBP51-Hsp90 interaction.22 These compounds bind to FKBP51, preventing it from docking with Hsp90 and thereby disrupting the complex in cellular assays. This disruption modulates downstream biological processes linked to tau pathology.22,23 The same principle is being applied to other key co-chaperones. For example, inhibitors targeting the Hsp90-Aha1 interface have demonstrated efficacy in reducing tau phosphorylation in neuronal models.10,43 By selectively disrupting the Aha1-driven hyperactivation of Hsp90’s ATPase cycle, these compounds show promise in halting tau aggregation.32 Such hyper-specific targeting of PPIs within the Hsp90 machinery represents a precise and potentially safer therapeutic strategy for AD.
PPI inhibitors offer the highest theoretical selectivity but carry distinct translational liabilities. Most reported compounds remain at the tool or lead-optimization stages, with pharmacokinetic properties, CNS penetration, and selectivity across the ~20 Hsp90 co-chaperones still incompletely characterized. Moreover, recent structural work indicates that co-chaperones such as FKBP51 and p23 can modulate tau via Hsp90-independent mechanisms, eg, p23 alone or a p23-FKBP51 complex, influencing tau aggregation, suggesting that disrupting a single Hsp90–co-chaperone interface may be insufficient.44
Strategic Induction: Leveraging Hsp90 for Proteolytic Clearance
In a striking departure from the inhibition paradigm, another innovative approach involves leveraging Hsp90’s functions to enhance proteolytic clearance of pathogenic proteins. This counterintuitive strategy recognizes that Hsp90’s role is not universally pathogenic and, in specific contexts, can be harnessed to promote protein clearance. A proof-of-concept study by Nguyen et al demonstrated that inducing Hsp90 expression on the surface of microglia with all-trans retinoic acid (ATRA) significantly enhanced the internalization and degradation of extracellular tau via lysosomal and proteasomal pathways.24 In this context, Hsp90 functions as a receptor or facilitator for tau clearance, transforming it into a therapeutic tool. Similarly, Zhang et al identified Jujuboside A as a compound that ameliorates cognitive deficits by activating the Axl/HSP90β/PPARγ pathway, ultimately enhancing Aβ clearance by microglia.25 Although conceptually innovative, this approach is confounded by the pleiotropic actions of all‑trans retinoic acid and natural products such as jujuboside A, making it difficult to ascribe the observed benefits solely to Hsp90‑mediated mechanisms. Another example comes from Liu et al, who found that deer antler uridine improves cognitive function by modulating microglia via an HSP90/HIF-1α pathway.40 These findings underscore that, depending on the cellular context and pathological process, boosting specific Hsp90-mediated pathways can serve as a powerful therapeutic strategy for AD.
Induction strategies, although mechanistically attractive, carry the converse risk: sustained upregulation of Hsp90 in non-target cells could in principle, stabilize pro-oncogenic or pro-inflammatory clients. Rigorous cell type- and tissue-specific delivery, coupled with long-term safety monitoring, will therefore be indispensable.
The Table 1 summary of Hsp90 rebalancing strategies for AD.
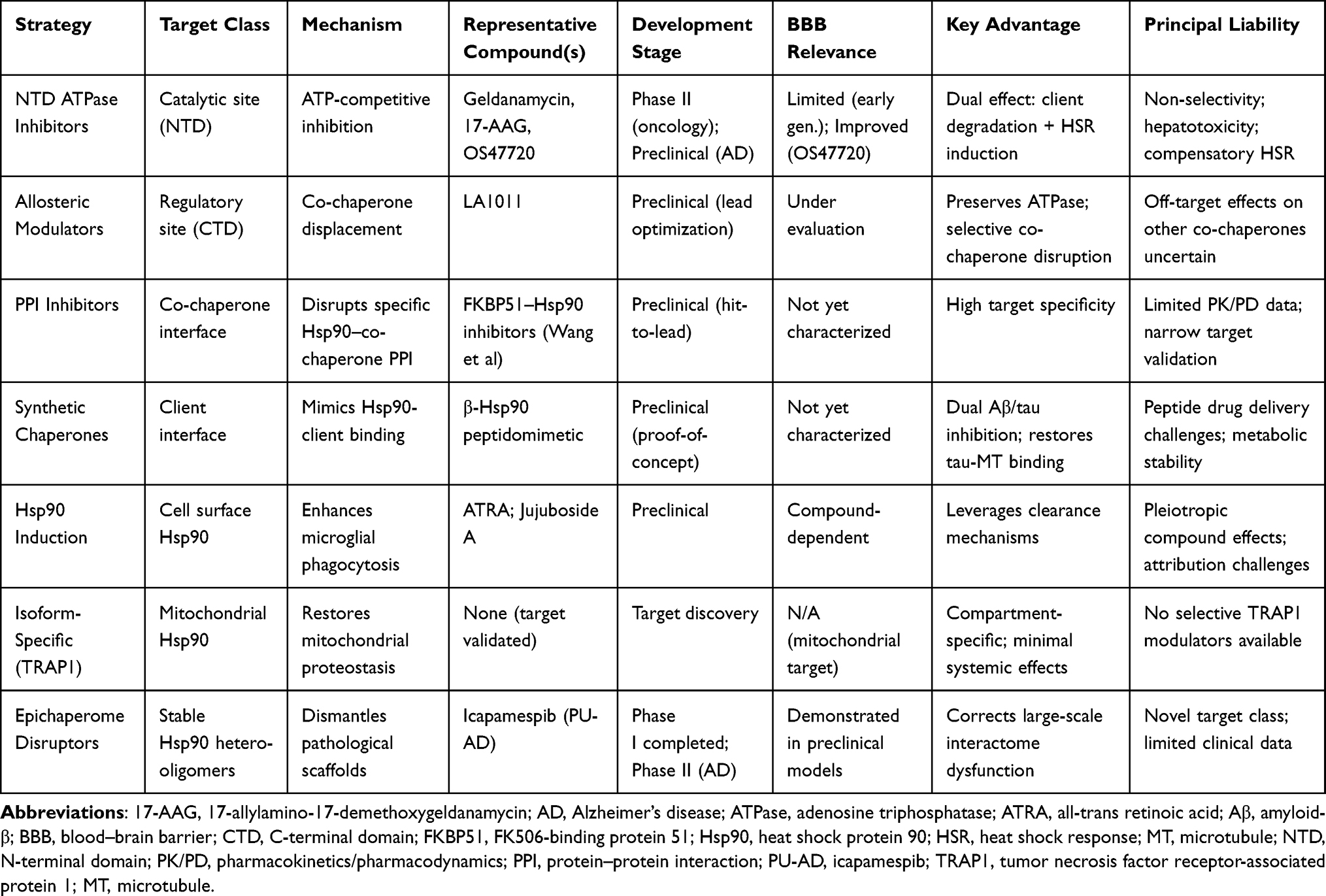 |
Table 1 Comparative Overview of Hsp90-Targeted Therapeutic Strategies for Alzheimer’s Disease |
Emerging Frontiers and Future Perspectives
The ongoing evolution of Hsp90-targeted therapeutics from broad-spectrum inhibition to precision modulation is opening up new frontiers that could redefine the treatment landscape for AD. These innovative approaches, which include pharmacological mimicry, isoform-specific targeting, and the development of sophisticated clinical tools, represent a significant departure from conventional strategies. However, their transition from preclinical concept to clinical application is fraught with challenges that the field must systematically address (Figure 4).
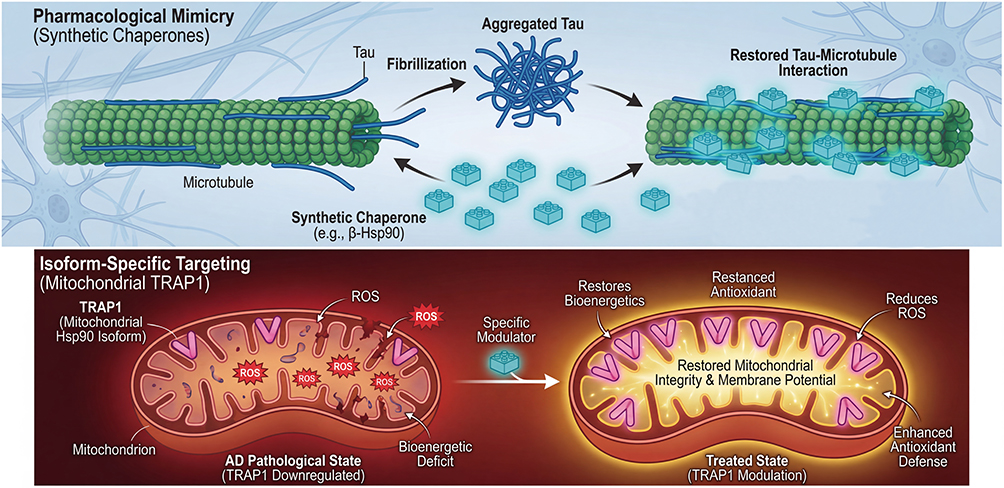 |
Figure 4 Emerging frontiers in chaperone-based therapeutics. Pharmacological mimicry (synthetic Chaperones): Synthetic peptides (eg, β-Hsp90) mimic the Hsp90 binding interface and function as “decoys,” binding directly to aggregation-prone regions of Tau, thereby inhibiting fibrillization and restoring physiological Tau-microtubule interactions. Isoform-specific targeting (mitochondrial TRAP1): Targeting the mitochondrial Hsp90 isoform TRAP1, which is downregulated in AD aims to restore mitochondrial integrity, membrane potential, and antioxidant defenses, addressing bioenergetic deficits in AD neurons. Evidence status: Pharmacological mimicry (β-Hsp90): demonstrated in vitro and in cellular models; in vivo efficacy data are not yet available. Isoform-specific targeting (TRAP1): Based on observational data showing TRAP1 downregulation in AD brain tissue; therapeutic strategies targeting TRAP1 in AD remain theoretical and require experimental validation. These approaches are presented as emerging concepts rather than validated therapeutic strategies. Abbreviations: AD, Alzheimer’s disease; Hsp90, heat shock protein 90; TRAP1, tumor necrosis factor receptor-associated protein 1. |
Novel Therapeutic Modalities
Beyond the established strategies of allosteric modulation and targeted PPI inhibition, researchers are exploring advanced modalities that seek even greater specificity and unique mechanisms of action. These pioneering efforts aim to engage the Hsp90 machinery with refined precision, targeting its pathological roles at their source.
Synthetic Chaperones Based on Hsp90-Client Interfaces
One particularly elegant strategy involves the rational design of synthetic molecules that mimic the interaction interfaces between Hsp90 and its clients. Instead of targeting Hsp90 itself, this approach creates a “synthetic chaperone” that competitively inhibits pathological PPIs driving aggregation. A notable example is the work by Di Lorenzo et al, who developed a peptidomimetic termed β-Hsp90, inspired by specific Hsp90 sequences known to bind tau.45 This synthetic molecule effectively interacts with tau’s aggregation-prone region, potently inhibiting its fibrillization. Crucially, by targeting tau directly rather than its microtubule-binding region, β-Hsp90 successfully restored the physiological interaction between tau and microtubules a function often disrupted in tauopathies. Additionally, the approach demonstrated efficacy against Aβ aggregation, highlighting its potential as a dual-inhibitor for AD. This innovative strategy of pharmacological mimicry underscores the power of mechanism-based drug design and offers a pathway to creating highly specific therapeutics tailored to AD pathology.45
Targeting Isoform-Specific Hsp90 (eg, Mitochondrial TRAP1)
Another promising avenue lies in exploiting the isoform diversity of the Hsp90 family In mammals, Hsp90 exists as four main paralogues: the cytosolic Hsp90α (inducible) and Hsp90β (constitutive), the endoplasmic reticulum-resident glucose-regulated protein 94 (GRP94), and the mitochondrial tumor necrosis factor receptor-associated protein 1 (TRAP1).8,46 This diversity offers an opportunity to develop isoform-specific drugs that confine therapeutic effects to specific cellular compartments, minimizing the systemic toxicity associated with inhibiting the ubiquitously expressed cytosolic isoforms. Among these paralogues, the mitochondrial Hsp90 isoform TRAP1 has emerged as an intriguing target. Dekker and Rüdiger have highlighted compelling evidence that TRAP1 is significantly downregulated in AD brains.46 Given TRAP1’s critical role in maintaining mitochondrial function including preserving the electron transport chain, regulating mitochondrial membrane potential, and mitigating oxidative stress, its modulation represents a logical therapeutic goal. As mitochondrial dysfunction is an early and central feature of AD pathogenesis, restoring TRAP1 activity or selectively targeting its pathological interactions could provide a viable neuroprotective strategy. Drugs designed with selectivity for TRAP1 could directly address mitochondrial health, a key vulnerability in AD.46
The Potential of Epichaperome Disruptors
Perhaps the most conceptually advanced frontier in Hsp90-targeted therapeutics is the development of drugs designed to dismantle pathological “epichaperome” complexes. As hypothesized by Chiosis and Neckers, these stable, high-molecular-weight Hsp90 hetero-oligomers act as pathological scaffolds, driving widespread proteomic disruption in AD.8 Unlike conventional approaches that inhibit Hsp90’s ATPase activity or disrupt specific PPIs, targeting epichaperomes would aim to dismantle their unique architecture, releasing sequestered proteins and restoring the cellular interactome. The therapeutic potential of such a strategy lies in its ability to correct large-scale proteostasis disruptions at their root. By breaking apart these pathological scaffolds, epichaperome disruptors could normalize the interactome, alleviating the pleiotropic downstream effects of protein misfolding and aggregation. While this field remains in its infancy, the concept represents a logical and exciting progression in Hsp90-targeted therapy, moving beyond functional inhibition to structural correction. As tools for studying and targeting epichaperomes become more sophisticated, this approach could herald a new era of therapeutics for AD and other neurodegenerative diseases.
Challenges and Clinical Translation
Despite the growing innovation in therapeutic design, significant translational hurdles remain. The journey from a promising compound in preclinical models to an effective therapy for human patients is especially arduous for neurodegenerative diseases. Key challenges in the clinical development of Hsp90 modulators include overcoming drug delivery barriers, validating targets in humans, and ensuring a safe and effective therapeutic window.
Overcoming the Blood-Brain Barrier
A persistent challenge in CNS drug development is delivering therapeutics across the highly selective BBB. Early-generation Hsp90 inhibitors often suffered from poor CNS penetration, severely limiting their utility for treating brain disorders.12 Newer compounds, such as the Hsp90 inhibitor OS47720, have been specifically optimized for BBB permeability and demonstrated efficacy in mouse models of AD.41 However, achieving adequate drug concentrations in the brain remains a critical parameter that must be rigorously evaluated for every clinical candidate. To address this, continued innovation in medicinal chemistry and advancements in drug delivery systems, such as nanoparticle-based carriers or prodrug strategies, will be essential. These approaches aim to enhance delivery efficiency while minimizing off-target effects, ensuring that novel Hsp90 modulators reach their intended targets in the brain at therapeutically relevant levels.
Developing Biomarkers for Target Engagement and Efficacy
For translational application, Hsp90-related biomarkers can be grouped into three categories, each aligned with a distinct drug-development decision point:
- Disease-state biomarkers quantify pathological Hsp90 biology. [11C]HSP990 PET shows reduced tracer binding in AD hippocampus, particularly in tau-pathology-enriched gray matter, and was developed to quantify brain Hsp90 expression in vivo to support Hsp90-targeted drug development.27 Radiolabeled PU-AD selectively labels epichaperomes in AD patients and can serve as a combined diagnostic and therapeutic tool.47 These biomarkers enable patient stratification and population enrichment in clinical trials.
- Target-engagement biomarkers verify in vivo target binding. The same Hsp90-directed tracers can be used in displacement or occupancy paradigms (eg, with onalespib, SNX-0723, PU-AD, or HSP990 pretreatment), hereby informing dose selection and first-in-human proof of mechanism.27
- Pharmacodynamic biomarkers reflect downstream biological response Hsp70 induction (measurable in blood or CSF) has been quantitatively linked to Hsp90 target occupancy in preclinical models and serves as a surrogate of chaperone-network perturbation. For AD-relevant efficacy readouts, fluid biomarkers such as plasma/CSF p-tau217, p-tau181, Aβ42/40, GFAP, and NfL are appropriate secondary endpoints for early proof-of-pharmacodynamic-effect studies.
A pragmatic trial design would combine category (i) for enrollment, category (ii) for Phase 1 dose selection, and category (iii) for early Phase 2 proof-of-mechanism evaluation.
Selectivity: Targeting Pathological vs Homeostatic Hsp90 Function
A fundamental challenge in targeting Hsp90, a protein essential for maintaining cellular homeostasis, is achieving therapeutic selectivity. Specifically, therapies must inhibit Hsp90’s pathological activities without disrupting its vital physiological functions. Striking this balance is crucial to minimize off-target toxicity, particularly for chronic conditions like AD where long-term treatment is required. A breakthrough study by Dickey et al offered a promising solution to this challenge. They discovered that Hsp90 complexes isolated from diseased AD brain regions exhibited dramatically increased affinity up to 1000-fold higher for Hsp90 inhibitors compared to those from healthy brain tissue.18 This finding highlights a phenomenon similar to the “chaperone addiction” observed in cancer cells, where hyper-activated Hsp90 systems in stressed or diseased cells become uniquely vulnerable to inhibition. This natural therapeutic window suggests that, at appropriate concentrations, inhibitors may preferentially bind to pathological Hsp90 complexes while sparing the lower-affinity, homeostatic chaperone systems. The emerging wave of precision modulation strategies, such as allosteric modulators and PPI inhibitors, is specifically designed to exploit this differential sensitivity. By targeting co-chaperone interactions or pathological client associations unique to disease states, these approaches aim to selectively disrupt the dysregulated Hsp90 machinery while preserving its normal, life-sustaining functions.20,22,23 This refined approach to therapeutic selectivity offers a safer and more effective path forward in Hsp90-targeted drug development.
Conclusion
Recapitulation: Hsp90 as a Central Nexus in AD Pathophysiology
A comprehensive exploration of the literature reveals that Hsp90 is not merely a bystander in AD but a central and deeply influential nexus in its pathophysiology. Its pivotal role arises from its profound and often paradoxical governance over the key protein drivers of the disease, tau and Aβ. The Hsp90 molecular chaperone machinery, through its intricate and context-dependent interactions with a diverse array of co-chaperones, wields considerable power in dictating the fate of these proteins.12,13 However, in the AD brain, this delicate balance is severely disrupted, causing Hsp90’s role to shift from protective to pathological. Co-chaperones such as FKBP51 and Aha1 hijack the Hsp90 system, stabilizing and promoting toxic tau species, while the protective actions of co-chaperones like CHIP are overwhelmed.6,18,32,33 Meanwhile, Hsp90’s interactions with Aβ are strikingly dualistic: it can inhibit its aggregation under normal conditions but, under stress, may paradoxically facilitate its production.35,37 This duality highlights both the immense complexity of Hsp90’s role in AD and the challenges of therapeutic intervention. The recent conceptualization of the “epichaperome” adds yet another dimension, proposing that in diseased states, Hsp90 transforms into a pathological scaffold, regulating widespread disruption of the cellular protein interactome.8 These insights collectively underscore the transition in therapeutic strategy from broad-spectrum inhibition to precise and context-sensitive modulation.
Final Verdict: Rebalancing the Hsp90 Chaperone Network as a Sophisticated and Viable Therapeutic Avenue
Given this evolving understanding, our assessment of the current evidence suggests that targeting the Hsp90 chaperone network remains a highly promising therapeutic avenue for AD but one that demands sophistication and precision. The era of pan-Hsp90 inhibition as a primary approach for chronic neurodegenerative diseases has rightly given way to more nuanced strategies that aim to rebalance, rather than eliminate, Hsp90’s activities. Recent preclinical successes exemplify this shift. Allosteric modulators like LA1011, which selectively disrupt pathological co-chaperone interactions,20 and rationally designed PPI inhibitors,22,23 suggest potentially transformative directions, contingent on clinical validation. Synthetic chaperones that mimic Hsp90-client interaction sites45 and microglia-focused Hsp90 induction strategies24 further illustrate the growing repertoire of innovative approaches. These precision-based strategies hold theoretical potential for improved therapeutic efficacy, though this remains to be demonstrated clinically while minimizing off-target effects, significantly improving the safety profile of Hsp90-targeted therapies. Nonetheless, significant translational challenges remain. Chief among these are the effective delivery of drugs to the brain across the BBB and the development of robust, translatable biomarkers for target engagement and efficacy.41,48 However, the discovery that pathological Hsp90 complexes in AD brains exhibit heightened sensitivity to modulation provides a crucial therapeutic window18 This insight shifts the therapeutic objective: the goal is no longer to inhibit Hsp90 indiscriminately but to skillfully manipulate its intricate network of interactions. By dismantling Hsp90’s pathological functions while preserving or even enhancing its homeostatic roles, we can achieve a sophisticated rebalancing act that offers a mechanistically coherent framework that, if validated clinically, could contribute to disease-modifying therapy for AD. It should be emphasized that with the partial exception of the epichaperome-directed agent PU‑AD, currently in Phase 2 evaluation for AD following a Phase 1 safety study in healthy volunteers, no Hsp90‑rebalancing therapy has yet produced positive pivotal clinical data in AD; accordingly, the claims advanced in this review are mechanistically grounded but clinically unvalidated.39
Looking forward, three actionable priorities should guide the next phase of Hsp90-targeted AD drug development: (i) mechanism-matched compound design, pairing allosteric modulators or PPI inhibitors with defined pathological targets (eg, Hsp90/FKBP51, Hsp90/Aha1, epichaperomes) rather than global ATPase inhibition; (ii) biomarker-anchored trials, embedding target-engagement markers (eg, [11C]HSP990 or [124I]PU-AD PET) and pharmacodynamic readouts (eg, CSF Hsp70, phospho-tau, Aβ) from first-in-human studies onward; and (iii) cell-type-selective delivery, leveraging advances in nanocarriers, prodrugs, and microglia-directed strategies to confine pharmacology to disease-relevant compartments A tempered interpretation is warranted: despite a compelling mechanistic rationale, the field remains predominantly preclinical, and the definitive test of the rebalancing hypothesis will come from prospective, biomarker-anchored clinical evaluation.
Data Sharing Statement
No new data were created or analyzed in this study.
Author Contributions
All authors have made substantial contributions to the work, whether in conception and study design, execution, data acquisition, analysis and interpretation, or in all these areas; have participated in drafting, revising, or critically reviewing the manuscript; have approved the final version for publication; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported in part by the Education Department Science Foundation of Jiangxi Province (GJJ2201452), the Jiangxi Provincial Natural Science Foundation (20232BAB216048, 20244BAB28044), the National Natural Science Foundation of China (82460266), the Science and Technology Project of Ganzhou City (2025YLCE0105).
Disclosure
The authors declare no conflicts of interest.
References
1. Sharma A, Srivastava S, Gupta P, et al. Targeting protein misfolding in Alzheimer’s disease: the emerging role of molecular chaperones. Biomed Pharmacother. 2025;191:118531 doi:10.1016/j.biopha.2025.118531.
2. Lackie RE, Maciejewski A, Ostapchenko VG, et al. The Hsp70/Hsp90 chaperone machinery in neurodegenerative diseases. Front Neurosci. 2017;11:254. doi:10.3389/fnins.2017.00254
3. Ou JR, Tan MS, Xie AM, Yu JT, Tan L. Heat shock protein 90 in Alzheimer’s disease. Biomed Res Int. 2014;2014:796869 doi:10.1155/2014/796869.
4. Muller P, Ruckova E, Halada P, et al. C-terminal phosphorylation of Hsp70 and Hsp90 regulates alternate binding to co-chaperones CHIP and HOP to determine cellular protein folding/degradation balances. Oncogene. 2013;32:3101–15. doi:10.1038/onc.2012.314
5. Campanella C, Pace A, Caruso Bavisotto C, et al. Heat shock proteins in Alzheimer’s disease: role and targeting. Int J Mol Sci. 2018;19:2603. doi:10.3390/ijms19092603
6. Jeanne X, Torok Z, Vigh L, Prodromou C. The role of the FKBP51-Hsp90 complex in Alzheimer’s disease: an emerging new drug target. Cell Stress Chaperones. 2024;29:792–804. doi:10.1016/j.cstres.2024.11.006
7. Wei H, Zhang Y, Jia Y, et al. Heat shock protein 90: biological functions, diseases, and therapeutic targets. MedComm. 2024;5:e470. doi:10.1002/mco2.470
8. Chiosis G, Digwal CS, Trepel JB, Neckers L. Structural and functional complexity of HSP90 in cellular homeostasis and disease. Nat Rev Mol Cell Biol. 2023;24:797–815. doi:10.1038/s41580-023-00640-9
9. Zuehlke AD, Moses MA, Neckers L. Heat shock protein 90: its inhibition and function. Philos Trans R Soc Lond B Biol Sci. 2018;373:20160527. doi:10.1098/rstb.2016.0527
10. Gracia L, Lora G, Blair LJ, Jinwal UK. Therapeutic potential of the Hsp90/Cdc37 interaction in neurodegenerative diseases. Front Neurosci. 2019;13:1263 doi:10.3389/fnins.2019.01263.
11. Bohush A, Bieganowski P, Filipek A. Hsp90 and its co-chaperones in neurodegenerative diseases. Int J Mol Sci. 2019;20:4976. doi:10.3390/ijms20204976
12. Blair LJ, Sabbagh JJ, Dickey CA. Targeting Hsp90 and its co-chaperones to treat Alzheimer’s disease. Expert Opin Ther Targets. 2014;18:1219–1232 doi:10.1517/14728222.2014.943185.
13. Salminen A, Ojala J, Kaarniranta K, Hiltunen M, Soininen H. Hsp90 regulates tau pathology through co-chaperone complexes in Alzheimer’s disease. Prog Neurobiol. 2011;93:99–110 doi:10.1016/j.pneurobio.2010.10.006.
14. Luo W, Sun W, Taldone T, Rodina A, Chiosis G. Heat shock protein 90 in neurodegenerative diseases. Mol Neurodegener. 2010;5:24. doi:10.1186/1750-1326-5-24
15. Luo W, Rodina A, Chiosis G. Heat shock protein 90: translation from cancer to Alzheimer’s disease treatment? BMC Neurosci. 2008;9 Suppl 2:S7. doi:10.1186/1471-2202-9-S2-S7
16. Waza M, Adachi H, Katsuno M, et al. Modulation of Hsp90 function in neurodegenerative disorders: a molecular-targeted therapy against disease-causing protein. J Mol Med. 2006;84:635–646. doi:10.1007/s00109-006-0066-0
17. Luo W, Dou F, Rodina A, et al. Roles of heat-shock protein 90 in maintaining and facilitating the neurodegenerative phenotype in tauopathies. Proc Natl Acad Sci U S A. 2007;104:9511–9516. doi:10.1073/pnas.0701055104
18. Dickey CA, Kamal A, Lundgren K, et al. The high-affinity HSP90-CHIP complex recognizes and selectively degrades phosphorylated tau client proteins. J Clin Invest. 2007;117:648–658. doi:10.1172/JCI29715
19. Petrucelli L, Dickson D, Kehoe K, et al. CHIP and Hsp70 regulate tau ubiquitination, degradation and aggregation. Hum Mol Genet. 2004;13:703–714. doi:10.1093/hmg/ddh083
20. Roe SM, Torok Z, McGown A, et al. The crystal structure of the Hsp90-LA1011 complex and the mechanism by which LA1011 may improve the prognosis of Alzheimer’s disease. Biomolecules. 2023;13:1051. doi:10.3390/biom13071051
21. Roe MS, Wahab B, Torok Z, Horvath I, Vigh L, Prodromou C. Dihydropyridines allosterically modulate Hsp90 providing a novel mechanism for heat shock protein co-induction and neuroprotection. Front Mol Biosci. 2018;5:51. doi:10.3389/fmolb.2018.00051
22. Wang L, Kumar R, Winblad B, Pavlov PF. Structure-based discovery of small molecule inhibitors of FKBP51-Hsp90 protein-protein interaction. Eur J Med Chem. 2024;270:116356. doi:10.1016/j.ejmech.2024.116356
23. Wang L, Bergkvist L, Kumar R, Winblad B, Pavlov PF. Targeting chaperone/co-chaperone interactions with small molecules: a novel approach to tackle neurodegenerative diseases. Cells. 2021;10:2596. doi:10.3390/cells10102596
24. Nguyen NL, Hoang TX, Kim JY. All-trans retinoic acid-induced cell surface heat shock protein 90 mediates tau protein internalization and degradation in human microglia. Mol Neurobiol. 2025;62:742–755 doi:10.1007/s12035-024-04295-1.
25. Zhang M, Qian C, Zheng ZG, et al. Jujuboside A promotes Abeta clearance and ameliorates cognitive deficiency in Alzheimer’s disease through activating Axl/HSP90/PPARgamma pathway. Theranostics. 2018;8:4262–4278. doi:10.7150/thno.26164
26. Inda MC, Joshi S, Wang T, et al. The epichaperome is a mediator of toxic hippocampal stress and leads to protein connectivity-based dysfunction. Nat Commun. 2020;11:319. doi:10.1038/s41467-019-14082-5
27. Cools R, Vermeulen K, Vonck E, et al. In vivo visualization and quantification of brain heat shock protein 90 with [(11)C]HSP990 in healthy aging and neurodegeneration. J Nucl Med. 2025;66:940–947. doi:10.2967/jnumed.124.268961
28. Shelton LB, Koren J, Blair LJ. Imbalances in the Hsp90 chaperone machinery: implications for tauopathies. Front Neurosci. 2017;11:724 doi:10.3389/fnins.2017.00724.
29. Blair LJ, Nordhues BA, Hill SE, et al. Accelerated neurodegeneration through chaperone-mediated oligomerization of tau. J Clin Invest. 2013;123:4158–4169 doi:10.1172/JCI69003.
30. Jinwal UK, Koren J, Borysov SI, et al. The Hsp90 cochaperone, FKBP51, increases Tau stability and polymerizes microtubules. J Neurosci. 2010;30:591–599 doi:10.1523/JNEUROSCI.4815-09.2010.
31. Oroz J, Chang BJ, Wysoczanski P, et al. Structure and pro-toxic mechanism of the human Hsp90/PPIase/Tau complex. Nat Commun. 2018;9:4532 doi:10.1038/s41467-018-06880-0.
32. Shelton LB, Baker JD, Zheng D, et al. Hsp90 activator Aha1 drives production of pathological tau aggregates. Proc Natl Acad Sci U S A. 2017;114:9707–9712. doi:10.1073/pnas.1707039114
33. Criado-Marrero M, Gebru NT, Blazier DM, et al. Hsp90 co-chaperones, FKBP52 and Aha1, promote tau pathogenesis in aged wild-type mice. Acta Neuropathol Commun. 2021;9:65 doi:10.1186/s40478-021-01159-w.
34. Hipp MS, Kasturi P, Hartl FU. The proteostasis network and its decline in ageing. Nat Rev Mol Cell Biol. 2019;20:421–435 doi:10.1038/s41580-019-0101-y.
35. Iqbal M, Lewis SL, Padhye S, Jinwal UK. Updates on abeta processing by Hsp90, BRICHOS, and newly reported distinctive chaperones. Biomolecules. 2023;14:16. doi:10.3390/biom14010016
36. Schirmer C, Lepvrier E, Duchesne L, et al. Hsp90 directly interacts, in vitro, with amyloid structures and modulates their assembly and disassembly. Biochim Biophys Acta. 2016;1860:2598–2609. doi:10.1016/j.bbagen.2016.07.033
37. Noorani AA, Yamashita H, Gao Y, et al. High temperature promotes amyloid beta-protein production and gamma-secretase complex formation via Hsp90. J Biol Chem. 2020;295:18010–18022. doi:10.1074/jbc.RA120.013845
38. Lamoke F, Mazzone V, Persichini T, et al. Amyloid beta peptide-induced inhibition of endothelial nitric oxide production involves oxidative stress-mediated constitutive eNOS/HSP90 interaction and disruption of agonist-mediated Akt activation. J Neuroinflammation. 2015;12:84. doi:10.1186/s12974-015-0304-x
39. Silverman MH, Duggan S, Bardelli G, et al. Safety, tolerability and pharmacokinetics of Icapamespib, a selective epichaperome inhibitor, in healthy adults. J Prev Alzheimers Dis. 2022;9:635–645. doi:10.14283/jpad.2022.71
40. Liu Y, Han C, Guo L, et al. Deer antler uridine regulates glycolysis in microglia via HSP90/HIF-1alpha to improve cognitive impairment in Alzheimer’s disease mice. CNS Neurosci Ther. 2025;31:e70416. doi:10.1111/cns.70416
41. Wang B, Liu Y, Huang L, et al. A CNS-permeable Hsp90 inhibitor rescues synaptic dysfunction and memory loss in APP-overexpressing Alzheimer’s mouse model via an HSF1-mediated mechanism. Mol Psychiatry. 2017;22:990–1001. doi:10.1038/mp.2016.104
42. Goel B, Jaiswal S, Tripathi N. Recent advances in HSP90 inhibitors as targeted cancer therapy: chemical scaffolds, isoform selectivity, and clinical progress. Bioorg Chem. 2025;163:108782. doi:10.1016/j.bioorg.2025.108782
43. Singh JK, Hutt DM, Tait B, et al. Management of Hsp90-dependent protein folding by small molecules targeting the Aha1 co-chaperone. Cell Chem Biol. 2020;27:292–305e296. doi:10.1016/j.chembiol.2020.01.008
44. Chakraborty P, Zweckstetter M. Interplay of p23 with FKBP51 and their chaperone complex in regulating tau aggregation. Nat Commun. 2025;16:669. doi:10.1038/s41467-025-56028-0
45. Di Lorenzo D, Bisi N, Kaffy J, et al. Synthetic chaperone based on Hsp90-Tau interaction inhibits Tau aggregation and rescues physiological Tau-Microtubule interaction. Nat Commun. 2025;16:8756. doi:10.1038/s41467-025-63824-1
46. Dekker FA, Rudiger SGD. The mitochondrial Hsp90 TRAP1 and Alzheimer’s Disease. Front Mol Biosci. 2021;8:697913. doi:10.3389/fmolb.2021.697913
47. Bolaender A, Zatorska D, He H, et al. Chemical tools for epichaperome-mediated interactome dysfunctions of the central nervous system. Nat Commun. 2021;12:4669. doi:10.1038/s41467-021-24821-2
48. Soriano-Herrador C, Ubeda-Banon I, Villanueva-Anguita P, et al. The chaperones HSP90AA1, HSP90AB1 and BAG3 are specifically distributed among human hippocampal subfields during different Alzheimer’s disease stages. Neurobiol Dis. 2025;212:106971. doi:10.1016/j.nbd.2025.106971
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.