Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 19
Beer Intake is Positively, While Cheese and Dried Fruit Intake Are Negatively Genetically Correlated with Psoriasis: A GWAS Summary-Statistics Study Using condFDR/conjFDR
Authors Liu G ![]() , Gong F
, Gong F ![]() , Yang G, Luo Q
, Yang G, Luo Q ![]()
Received 4 November 2025
Accepted for publication 20 January 2026
Published 29 January 2026 Volume 2026:19 578871
DOI https://doi.org/10.2147/CCID.S578871
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Monica K. Li
Guo Liu,1,2 Fengjuan Gong,1 Guanhu Yang,2 Qinghua Luo2,3
1Department of Dermatology, Qionglai Hospital of Traditional Chinese Medicine, Chengdu, 611530, People’s Republic of China; 2Faculty of Chinese Medicine, Macau University of Science and Technology, Macau, 999078, People’s Republic of China; 3Clinical Medical College, Jiangxi University of Chinese Medicine, Nanchang, 330004, People’s Republic of China
Correspondence: Guo Liu, Department of Dermatology, Qionglai Hospital of Traditional Chinese Medicine, Chengdu, 611530, People’s Republic of China, Email [email protected]
Background: Previous epidemiological investigations have reported associations between beer, cheese, and dried fruit intake and the incidence of psoriasis (PS). Nevertheless, the extent of shared genetic architecture underlying these associations remains insufficiently characterized. Therefore, it is essential to examine their molecular relationships from a genomics perspective.
Methods: A stratified, stepwise analytical framework was applied, utilizing genome-wide association study data for PS and three dietary intake patterns (beer, cheese, and dried fruit intake). In the initial phase, linkage disequilibrium score regression, genetic covariance analysis, and high-definition likelihood approaches were employed to quantify genome-wide genetic correlations. Subsequently, local genetic overlap analyses were conducted to delineate chromosomal regions harboring relevant associations. Finally, the conditional/conjunctional false discovery rate (condFDR/conjFDR) methodology was implemented to identify potential shared genetic loci (genetic overlap) across the phenotypes.
Results: The analysis revealed consistent genome-wide genetic correlations between psoriasis (PS) and the three dietary intake traits across LDSC, GNOVA, and HDL. PS showed a significant positive genetic correlation with beer intake (Rg = 0.1082 [LDSC], 0.0796 [GNOVA], and 0.144 [HDL]), whereas negative genetic correlations were observed for cheese intake (Rg = − 0.1523, − 0.1089, and − 0.1866) and dried fruit intake (Rg = − 0.2187, − 0.1639, and − 0.2623). Local analyses further identified multiple chromosomal regions with shared genetic signals. Finally, condFDR/conjFDR analyses supported these overlaps and enabled the identification of several significant shared loci between PS and the dietary traits.
Conclusion: This study systematically investigated the genetic associations between PS and three dietary intake patterns from a genomic perspective and characterized their genome-wide and local genetic overlap. The shared loci identified enhance the understanding of nutrition–disease molecular mechanisms and provide genetic evidence to inform future mechanistic research and risk-modification strategies.
Keywords: psoriasis, dietary intake patterns, genetic correlation, genetic loci, genetic overlap
Introduction
Psoriasis (PS) is a chronic immune-mediated inflammatory dermatosis characterized by aberrant proliferation and differentiation of epidermal keratinocytes, leading to sharply demarcated erythematous plaques surmounted by silvery-white scales on the skin surface.1 According to the most recent Global Burden of Disease Study, the worldwide prevalence of PS ranges from 0.7% to 11.4%, affecting more than 125 million individuals, with an adult prevalence of approximately 2.3%. Marked geographical disparities have been documented, with the highest prevalence reported in Europe and North America.2,3 Over the past three decades, epidemiological surveys have indicated a progressive increase in PS prevalence, accompanied by rising age-standardized incidence rates and disability-adjusted life years between 1990 and 2019.4 Etiological investigations have demonstrated that, in addition to non-modifiable intrinsic factors such as genetic susceptibility, immune dysregulation, and environmental triggers,5 various modifiable external factors substantially influence disease development, primarily involving dietary patterns, lifestyle behaviors, and environmental exposures.6,7 Accumulating epidemiological evidence suggests that specific traditional dietary components, particularly beer, cheese, and dried fruit, may affect the onset of PS owing to their unique nutritional profiles and bioactive constituents.8–11 Given that confounding interactions between dietary factors in conventional observational studies may bias results, and despite growing interest in diet–PS links, an important research gap remains: integrative analyses that jointly leverage genome-wide (global) and region-specific (local) genetic information to systematically characterize shared genetic architecture between PS and dietary intake traits are still limited. Therefore, applying genomic methodologies to elucidate intrinsic associations between PS risk and these three dietary intake patterns may provide more robust and accurate evidence.
Genome-wide association studies (GWAS) provide essential information on genetic variation related to PS and three dietary intake patterns (beer, cheese, and dried fruit intake), thereby enabling the investigation of their genetic associations. The development of novel genetic statistical methodologies has rendered feasible the analysis of genetic relationships among complex traits. These approaches have yielded promising results in multiple studies, facilitating the identification of genetic connections across diverse complex traits. For instance, shared biological mechanisms were revealed between attention deficit hyperactivity disorder and cortisol alterations associated with externalizing behaviors;12 shared genetic loci were identified between inflammatory bowel disease and major depressive disorder;13 and genetic structural relationships were clarified between alcohol and cheese intake and inflammatory bowel disease.14 Building upon these established analytical strategies, this study undertakes a comprehensive examination of the genetic interactions between PS susceptibility and the three dietary intake patterns mentioned above.
This study established an integrated genetic statistical framework to delineate correlation patterns among target traits. At the genome-wide scale, linkage disequilibrium score regression (LDSC),15 genetic covariance analysis (GNOVA),16 and high-definition likelihood (HDL)17 were applied to evaluate global genetic correlations, whereas local analysis of variant association (LAVA)18 was employed to characterize genetic interaction features within specific chromosomal regions. To precisely identify shared genetic variant loci, a conditional/conjunctional false discovery rate (condFDR/conjFDR) approach19 was adopted, enabling both quantification of polygenic interaction strength and effective detection of shared genetic susceptibility loci.19 We aimed to characterize shared genetic architecture between psoriasis (PS) and three dietary intake patterns (beer, cheese, and dried fruit intake) by assessing genome-wide and local genetic overlap and identifying shared pleiotropic loci.
Methods and Materials
GWAS Data
The genomic data on dietary behaviors analyzed in this study were retrieved from the IEU OpenGWAS platform. GWAS summary statistics for three dietary intake patterns—beer intake (dataset ID: “ukb-b-5174”), cheese intake (dataset ID: “ukb-b-1489”), and dried fruit intake (dataset ID: “ukb-b-16576”)—were derived from the UK Biobank cohort, in which participants were recruited between 2006 and 2010. Phenotypic information for PS was obtained from the “L12_PSORIASIS” dataset of the FinnGen project (release 12).20 FinnGen was launched in 2017 and comprises both legacy samples collected from earlier Finnish cohorts (spanning from the late 1980s to 2017) and additional samples collected thereafter through Finnish biobanks. Given the binary (PS) versus continuous/ordinal nature of dietary intake traits, statistical power depends on case–control numbers, trait prevalence, measurement error, and GWAS sample size; thus, we leveraged large, well-powered GWAS datasets and applied multiple complementary methods to enhance the detection of shared genetic signals. The population characteristics of each cohort are summarized in Table 1.
 |
Table 1 Date Sources |
To ensure analytical rigor, stringent quality control procedures were implemented. Genetic variants were required to satisfy the following criteria:1 inclusion in the 1000 Genomes Project Phase 3 reference panel; (2) exclusion of variants lacking rsID identifiers or containing duplicate rsIDs; and (3) restriction to biallelic polymorphisms; and (4) exclusion of variants within the extended MHC region (chr6: ~25–34 Mb) to minimize potential bias arising from long-range linkage disequilibrium (LD). To minimize potential bias due to population stratification, analyses were restricted to individuals of European ancestry, consistent with the underlying GWAS datasets. A detailed workflow of the study is illustrated in Figure 1.
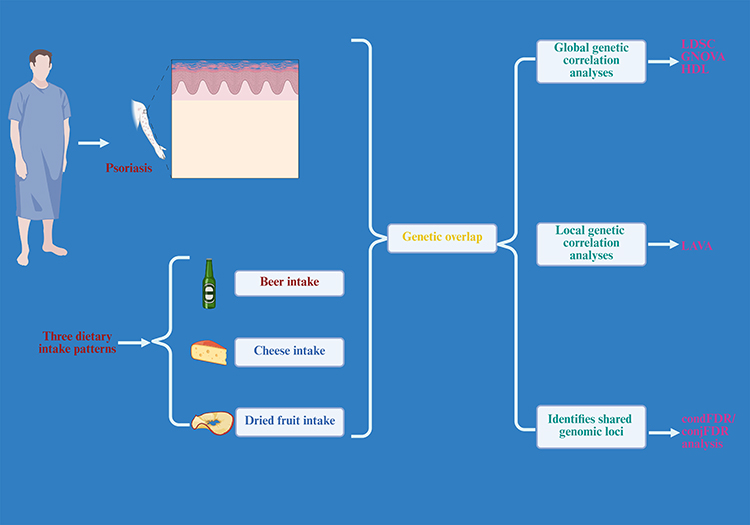 |
Figure 1 Flowchart of the study. This figure is created by biorender. |
Global Genetic Correlation Analyses
The LDSC workflow for estimating global genetic correlations between two traits consists of several sequential steps: data preprocessing, linkage disequilibrium (LD) score computation, regression modeling, genetic correlation estimation, error adjustment, and result interpretation.15 Initially, LD scores are calculated for single-nucleotide polymorphisms (SNPs), with each SNP assigned a score representing the extent of genotypic association with other SNPs. Weighted least squares regression is subsequently applied to examine the relationship between SNP effects (Z-scores) and LD scores, thereby yielding an estimate of genetic covariance (ρg) between traits. To enhance estimation precision, the Jackknife method is used to evaluate standard errors and correct potential estimation biases. Normalization of genetic covariance then allows calculation of the genetic correlation (rg), which represents the shared genetic foundation of the traits. Throughout this process, LDSC effectively mitigates the influence of sample overlap by incorporating the constrained intercept method to adjust bias, while integrating population structure adjustments (eg, principal component analysis) to reduce false-positive signals. Moreover, because LDSC does not require individual-level genotype data, its application can be extended broadly to diverse genetic epidemiological investigations.
GNOVA is a genetic covariance estimation approach based on GWAS summary statistics that enhances estimation precision and statistical power by incorporating functional genomic annotation stratification analysis.16 In this framework, high-quality SNPs are screened and LD information is calculated, with the assumption that SNP effects vary across distinct functional regions. The method of moments is applied to construct linear equations, which are solved for genetic covariance through weighted matrices, while block jackknife procedures are used to correct for sample overlap. GNOVA also enables stratification analyses according to functional annotations or minor allele frequency, thereby uncovering shared genetic architecture among complex traits and yielding precise covariance estimates. This approach is particularly well-suited for large-scale GWAS data analyses.
HDL is a full-likelihood genetic correlation estimation method that achieves greater precision than LDSC by fully exploiting LD information contained in GWAS summary statistics.17 In this approach, GWAS Z-statistics for SNPs are computed and genetic covariance models are constructed based on LD matrices. Maximum likelihood estimation is then applied to derive genetic correlations, while eigenvalue decomposition is employed to regularize LD matrices and alleviate computational complexity. Unlike LDSC, which relies solely on diagonal elements of LD matrices, HDL incorporates the entire matrix structure to generate more accurate genetic correlation estimates, offering distinct advantages, particularly in analyses of low-heritability or binary traits. Both simulation and empirical evidence have demonstrated that HDL substantially improves the precision and statistical power of genetic correlation estimation.
Local Genetic Correlation Analyses
LAVA estimates local genetic correlations between two traits using localized genetic correlation modeling approaches.18 In this framework, genetic effects of individual SNPs within target regions are first estimated through linear regression models to compute local genetic signals. To ensure consistency of effect direction, GWAS summary statistics from different traits are aligned prior to analysis. The local genetic covariance matrix (Ω) is then estimated, from which the local genetic correlation (ρ) for each trait pair is derived. Moreover, LAVA accommodates multi-trait inference by incorporating partial correlations and multiple regression analyses, thereby enabling the characterization of complex genetic relationships. Both simulation experiments and empirical validations have demonstrated that LAVA accurately controls Type I error rates and exhibits superior statistical power in the analysis of multi-trait local genetic correlations, establishing it as an effective tool for disentangling genetic relationships among complex traits.
condFDR/conjFDR Analysis
condFDR and conjFDR are statistical approaches for GWAS data analysis designed to improve genetic signal detection and identify cross-trait shared genetic loci.19 condFDR is grounded in an empirical Bayesian framework that corrects GWAS results for target traits by incorporating SNP association information from auxiliary phenotypes, thereby reducing the false discovery rate (FDR) through optimized ranking of statistical tests. The analytical workflow involves preprocessing independent GWAS datasets, standardizing SNP effect directions, and applying quality control procedures; constructing conditional quantile plots to illustrate SNP enrichment patterns; calculating FDR based on p-value stratification from auxiliary phenotypes; and generating two-dimensional FDR lookup tables to enhance detection sensitivity.
conjFDR extends condFDR and is specifically designed to identify SNPs that exert simultaneous effects on two traits. Its calculation process involves estimating condFDR values separately for both traits and adopting the larger value as the final conjFDR, thereby providing conservative estimates of shared genetic signals. This method is capable of identifying shared variants even when the overall genetic correlation is modest. To mitigate bias from population stratification or sample overlap, conjFDR applies genomic inflation factor correction and reduces LD interference through decorrelation procedures (r2 < 0.1).
In summary, condFDR enhances the discovery potential of GWAS, whereas conjFDR facilitates the identification of shared genetic architecture across traits. These approaches are of considerable value in complex trait genetics, particularly for investigating comorbidity mechanisms and functional genomic annotation. For loci identified using condFDR/conjFDR, systematic gene annotation was conducted through the SNP2GENE functional module of the FUMA platform.21
Results
Global Genetic Correlation
Comprehensive genetic epidemiological analyses identified association patterns between PS and three specific dietary habits. LDSC indicated a positive genetic correlation between PS and alcohol intake (Rg = 0.1082, P = 0.0108). In contrast, negative correlations were observed for cheese intake (Rg = −0.1523, P = 1.2584e-05) and dried fruit intake (Rg = −0.2187, P = 1.3801e-10) (Table 2).
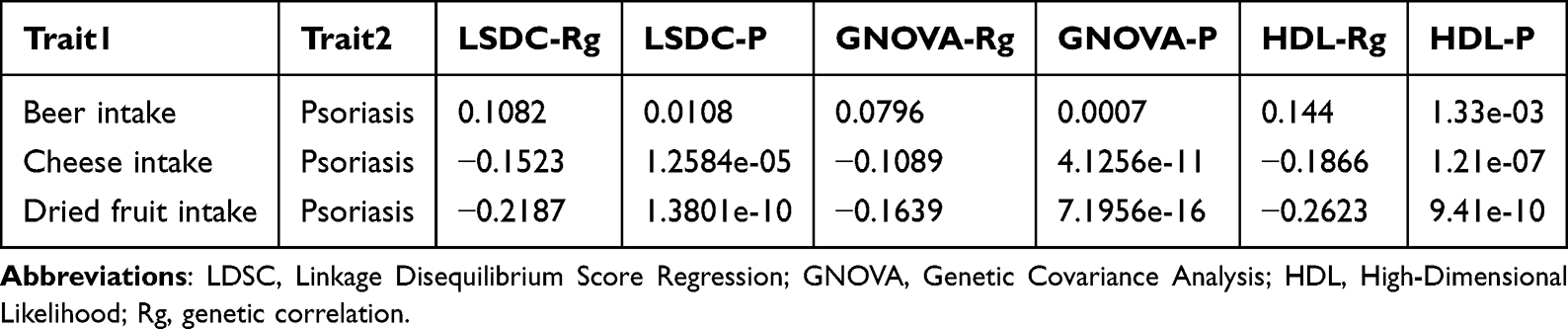 |
Table 2 Genetic Correlation of Psoriasis and Three Food Intake Patterns |
Independent validation with GNOVA and HDL algorithms corroborated these findings. The concordant results across multiple methods reinforced the authenticity of genetic interactions between PS and the dietary patterns under investigation (Table 2). Collectively, these observations broaden the understanding of shared genetic determinants linking PS with dietary intake behaviors.
Local Genetic Correlation
Local genetic correlation analyses across chromosomal regions uncovered multilayered association characteristics between PS and the three dietary intake patterns. Multiple statistically significant loci were identified throughout the genome, reflecting substantial heterogeneity in the genetic basis of PS relative to different dietary behaviors.
For the association between PS and beer intake, a genomic segment with significant positive correlation was detected on chromosome 2 (Figure 2A and Supplementary Table 1).
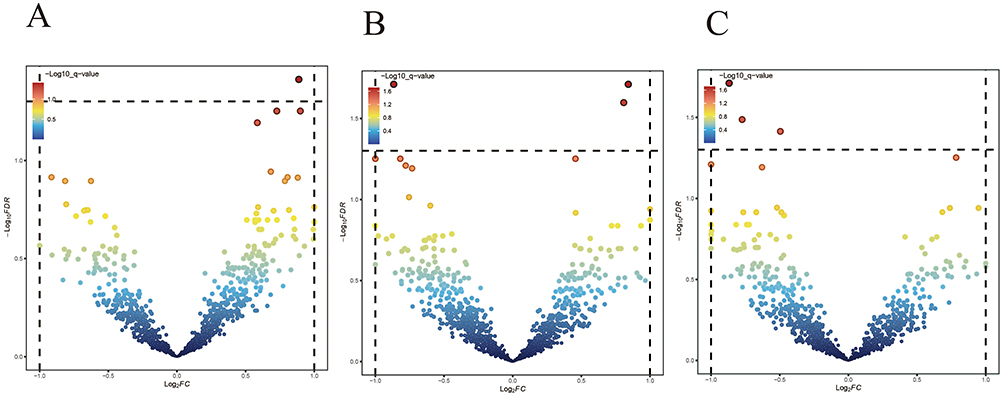 |
Figure 2 LAVA analysis of psoriasis and three dietary intake patterns. The dashed line indicates the expected line with a correction P of 0.05. (A) Local genetic correlation between psoriasis-beer intake. (B) Local genetic correlation between psoriasis-cheese intake. (C) Local genetic correlation between psoriasis-dried fruit intake. LAVA, local variant association. |
For cheese intake, three loci reaching significance were identified: two positively correlated regions on chromosomes 10 and 5, and one negatively correlated region on chromosome 11 (Figure 2B and Supplementary Table 2).
For dried fruit intake, five regions, all showing negative correlations, were identified, located on chromosomes 11, 12, 16, and 17 (Figure 2C and Supplementary Table 3).
Overall, local genetic correlation analysis revealed complex association patterns between PS and dietary intake at the chromosomal level, demonstrating multiple significantly correlated loci across distinct dietary factors.
conjFDR Analysis Identifies Shared Genomic Loci Between Two Traits
Quantile-quantile (Q-Q) plot visualization confirmed genome-wide associations between PS and the three dietary intake patterns (Figure 3). The analyses demonstrated that as the association signal strength of PS increased, the corresponding association strength of dietary behaviors exhibited a distinct leftward shift. This shift not only validated the genetic correlations among these traits but also suggested the existence of shared susceptibility loci, thereby providing a basis for further exploration of their underlying biological mechanisms.
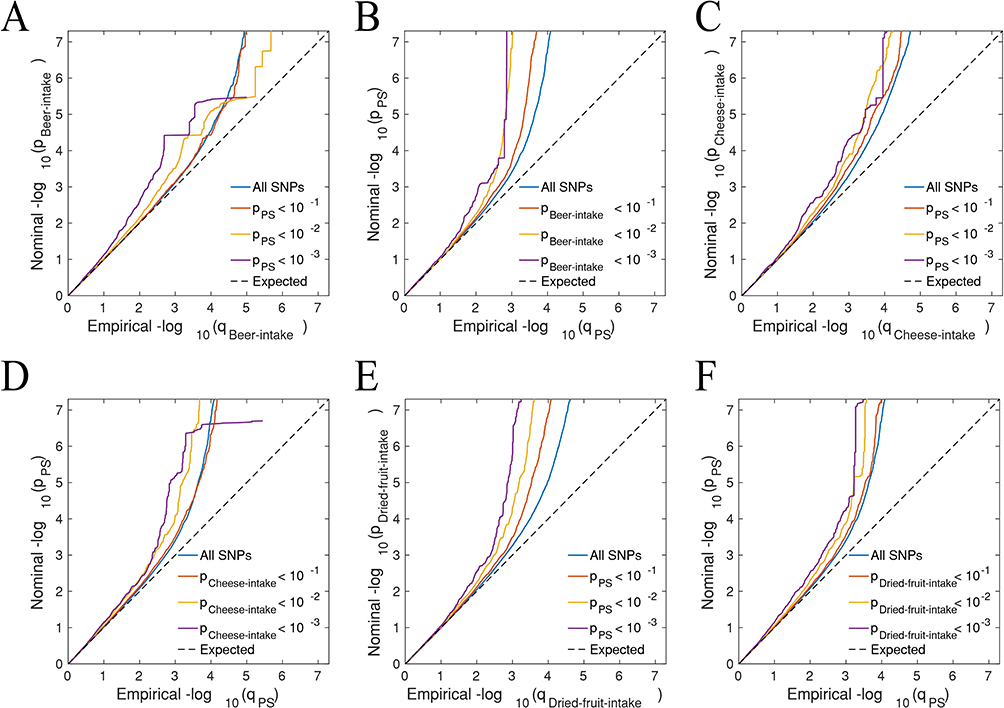 |
Figure 3 Conditional quantile-quantile plot. The dashed line indicates the expected line under the null hypothesis, and the deflection to the left indicates the degree of pleiotropic enrichment. (A) Beer intake-Psoriasis. (B) Psoriasis-Beer intake. (C) Cheese intake-Psoriasis. (D) Psoriasis-cheese intake. (E) Dried fruit intake-Psoriasis. (F) Psoriasis-Dried fruit intake. |
Application of the condFDR/conjFDR approach to genome-wide analysis identified four pleiotropic variants shared between PS and beer intake at the significance threshold of conjFDR < 0.05 (involving RP11-84D1.2, RAPGEF6, C5orf56, and DNAJC28 genes) (Figure 4A and Supplementary Table 4). Notably, RAPGEF6 has been implicated in immune-cell signaling and inflammatory regulation, supporting the biological plausibility of its involvement in PS-related pathways.22
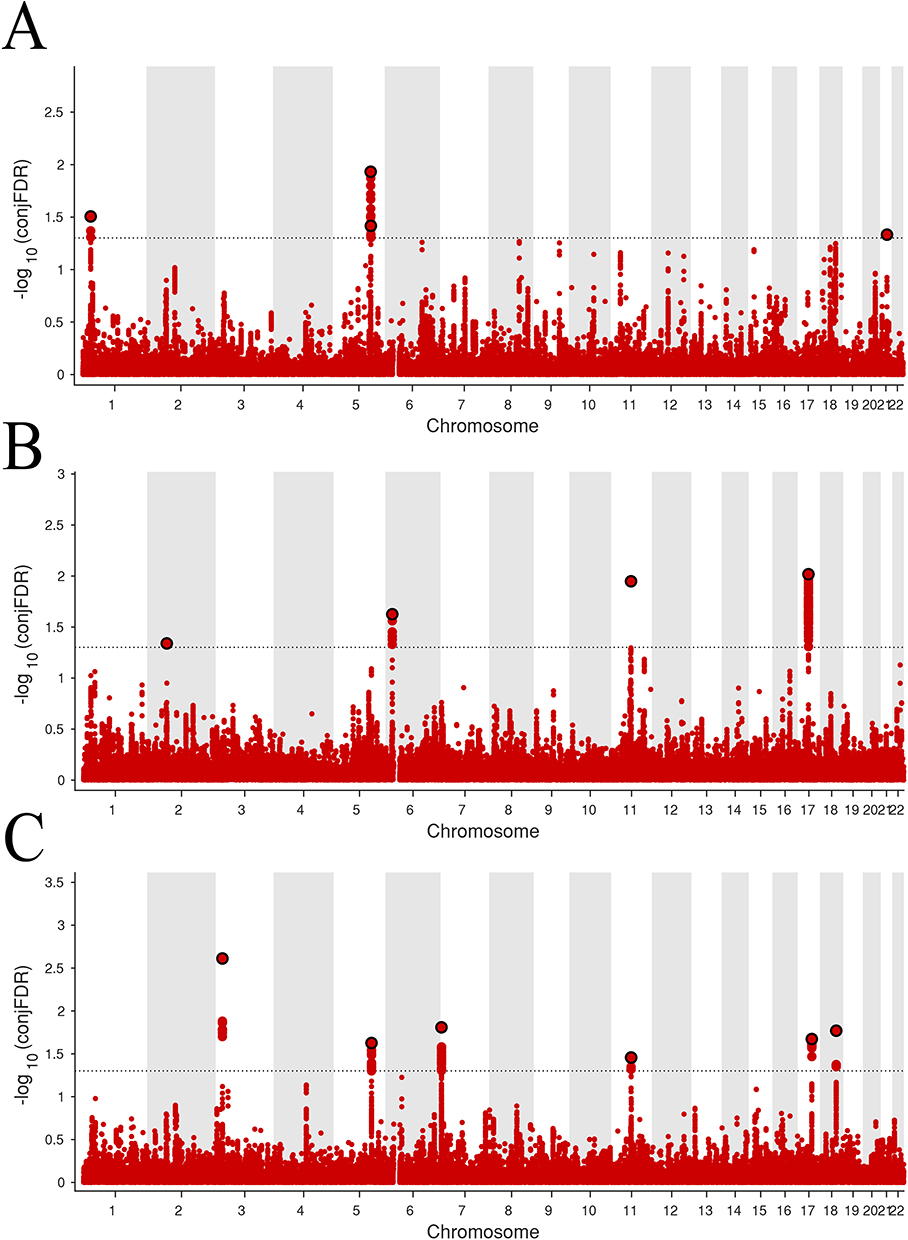 |
Figure 4 ConjFDR Manhattan plot of psoriasis and three dietary intake patterns. (A) PS-Beer intake. (B) PS-Cheese intake. (C) PS-Dried fruit intake. The statistically significant causality is defined to be conjFDR <0.05. |
Extended analyses revealed that PS and cheese intake shared four loci (located within AC007381.2, CDKAL1, FOSL1, and ATP6V0A1 gene regions) (Figure 4B and Supplementary Table 5), while seven shared variants were associated with dried fruit intake (distributed across AC144521.1, KIF3A, MAD1L1, SF3B2, PC, STXBP4, and RP11-795H16.2 genes) (Figure 4C and Supplementary Table 6). Importantly, to our knowledge, this study is among the first to apply the conjFDR framework to identify shared loci between PS and dietary traits—particularly for cheese and dried fruit intake—thereby highlighting the novelty of these pleiotropic signals. Several of the identified loci show biological relevance to PS-related mechanisms; for example, FOSL1 is linked to inflammatory transcriptional programs,23 ATP6V0A1 is involved in cellular acidification processes relevant to immune function,24 and KIF3A has been associated with epithelial/skin biology.25 Collectively, these genome-wide results provide genetic evidence for a shared molecular basis linking PS occurrence with specific dietary intake patterns.
Discussion
This study systematically investigated the genetic associations between PS and three dietary intake patterns using GWAS. Genome-wide analyses demonstrated a significant positive genetic correlation between PS and beer intake, whereas significant negative correlations were observed with both cheese and dried fruit intake. Local genetic correlation analyses further identified distinct association signals across multiple chromosomal segments. Validation through Q-Q plots confirmed the presence of overlapping genetic patterns among these phenotypes. In conjunction with the conjFDR analytical framework, multiple shared genetic variant loci were identified. From a clinical perspective, these results primarily support risk stratification and hypothesis generation by highlighting shared genetic architecture between PS and dietary intake traits, rather than providing a basis for dietary prescriptions. Collectively, these findings provide novel genetic-level insights into shared genetic mechanisms underlying the relationships between beer, cheese, and dried fruit intake and PS, thereby broadening the conceptual framework of nutrition–disease genetic association networks.
Recent studies suggest that beer intake may be associated with a higher risk of psoriasis.26 Large-scale prospective cohort studies reported that women consuming ≥5 servings of non-light beer weekly exhibited a 1.83-fold higher incidence of PS,27 whereas observational studies demonstrated a risk ratio of 1.30 for PS occurrence in high alcohol consumers compared with low consumers.28 From a mechanistic perspective, alcohol and its metabolites activate hepatic Kupffer cells and circulating monocytes to release pro-inflammatory mediators, thereby initiating persistent systemic inflammatory responses.26 Ethanol and reactive oxygen species generated during metabolism further induce pro-inflammatory cytokines in lymphocytes, macrophages, and keratinocytes through modulation of diverse signal transduction pathways, creating an inflammatory milieu that contributes to PS onset.29 Among the shared genetic variants identified, RAPGEF6 warrants particular attention, as it may serve a critical regulatory role in beer intake–induced PS. RAPGEF6 encodes Rap guanine nucleotide exchange factor 6, a member of the small GTPase regulatory protein family, which activates key signaling molecules including Rap1A, Rap2A, and M-Ras, thereby exerting central influence on immune cell regulation and inflammatory processes.22 Animal studies have shown that RAPGEF6-deficient mice display pronounced immune dysfunction, including splenomegaly, abnormally elevated serum immunoglobulin E and G levels, and altered pro-inflammatory cytokine secretion profiles, highlighting its indispensable role in immune homeostasis.30 In the context of PS pathogenesis, aberrantly activated T cells infiltrate lesional sites and secrete inflammatory mediators such as IL-17 and IFN-γ, whereas RAPGEF6 dysfunction may amplify this pathological immune cascade.31 Additionally, genetic variants in RAPGEF6 may influence susceptibility to alcohol intake patterns, indirectly modulating PS onset risk, as supported by prior evidence of ethanol-induced enhancement of pro-inflammatory factor secretion in PS cell co-culture systems and the proposed role of RAPGEF6 in psychiatric behavioral regulation.32 Furthermore, C5orf56 has been identified as a shared susceptibility gene between PS and cardiovascular diseases in recent GWAS reports.33
Observational epidemiological studies have reported an inverse association between cheese intake and psoriasis risk, although the underlying nutritional and genetic mechanisms remain unclear. Large-scale US national survey data indicate that dairy and calcium intake among PS patients are markedly lower than in healthy controls,34 implying that moderate cheese intake may confer preventive benefits. Pathophysiologically, PS patients frequently exhibit hypocalcemia, with more than one-third presenting abnormally reduced serum calcium levels. As a rich source of calcium, cheese can compensate for this critical nutritional deficiency.35 At the molecular level, calcium abundant in cheese interacts synergistically with Vitamin D to optimize intracellular calcium regulatory networks, thereby ameliorating the abnormal hyperproliferation of PS keratinocytes.36 Moreover, dairy intake can alleviate sodium-induced cutaneous microvascular dysfunction by reducing ascorbate-sensitive oxidative substances, providing evidence for the vascular protective role of cheese in PS prevention.37 As a key member of the tRNA methylthiotransferase family, CDKAL1 not only regulates translational fidelity but also plays a central role in cellular energy metabolism and calcium homeostasis.38 CDKAL1 deficiency markedly impairs adipose tissue mitochondrial function, leading to morphological abnormalities, respiratory dysfunction, and ATP synthesis defects.38 As PS is characterized as a metabolic inflammatory disorder associated with mitochondrial dysfunction and oxidative stress, calcium homeostasis disruption represents a pivotal feature. This includes impaired function of calcium-regulating proteins such as SERCA within the endoplasmic reticulum.39 Importantly, our GWAS-based results indicate shared genetic architecture between cheese intake and PS and do not establish that cheese intake causally prevents PS. The high calcium content of cheese may be relevant to calcium homeostasis in PS, but these mechanistic links remain speculative and require direct experimental and interventional validation. FOSL1 expression is markedly elevated in PS patients and shows positive correlations with PASI scores, serum TNF-α, and IL-6 levels,23 highlighting its pro-inflammatory role in PS pathogenesis. The proposed calcium–inflammation network concept offers new perspectives on the role of calcium ions in inflammatory regulation, suggesting that adequate calcium levels may attenuate systemic inflammation by reducing lipopolysaccharide aggregation and suppressing excessive macrophage activation.40 Therefore, cheese intake may mitigate FOSL1 overexpression through calcium-dependent signaling pathways, thereby reducing PS-associated inflammatory cascades and exerting potential preventive effects. Accordingly, any clinical implications should be framed as hypothesis-generating and primarily relevant to prevention/risk modification rather than therapeutic recommendations.
Recent observational evidence suggests that dried fruit intake may be inversely associated with psoriasis risk, although the underlying mechanisms remain to be fully established. Dried fruits are generally rich in dietary fiber and polyphenolic antioxidants, which may help modulate inflammatory pathways and oxidative stress. These bioactive components could potentially influence immune responses and keratinocyte proliferation and differentiation; however, causal effects cannot be inferred from the available epidemiological data.41 In the APPLE project, a cross-sectional study by Zanesco et al42 reported an inverse correlation between dried fruit intake and PS severity, supporting a protective effect. Furthermore, a systematic review and meta-analysis of 560 participants confirmed that omega-3 fatty acid supplementation significantly reduced psoriasis area and severity index scores by 1.58 points and improved clinical manifestations such as erythema and scaling.43 A recent Mendelian randomization analysis also established an inverse causal relationship between dried fruit intake and PS.10 Among the identified genetic loci, KIF3A has been extensively investigated for its role in dried fruit–mediated PS prevention. A large-scale GWAS by Leppers-van de Straat et al demonstrated that KIF3A variants rs11740584 and rs2299007 were significantly associated with immune dermatitis susceptibility. These alleles generated novel CpG sites and increased DNA methylation levels, resulting in downregulation of KIF3A expression and compromised skin barrier integrity.25 Mechanistic studies further showed that KIF3A-deficient mice exhibited elevated transepidermal water loss, disrupted claudin-1 expression in tight junctions, and heightened susceptibility to allergen-induced dermatitis.44 Notably, omega-3 fatty acids abundant in dried fruits may regulate KIF3A expression through epigenetic mechanisms. A comprehensive review by Balić et al indicated that omega-3 fatty acids indirectly enhance transcription of skin barrier–related genes, including KIF3A, by suppressing activation of the pro-inflammatory transcription factor NF-κB, thereby sustaining keratinocyte differentiation and intercellular junction stability.41 The remaining genes identified have been less extensively studied in the context of PS and require further investigation. Overall, our conclusions regarding dried fruit intake should be interpreted primarily as evidence of shared genetic architecture and hypothesis-generating clinical relevance (eg, prevention/risk modification), rather than as direct therapeutic recommendations.
Several limitations should be acknowledged. First, the dietary intake phenotypes were largely derived from self-reported questionnaire data (eg, UK Biobank) and are therefore susceptible to recall bias, social desirability bias, and measurement error/misclassification; consequently, non-differential misreporting may attenuate true genetic associations and increase uncertainty in effect estimates. Second, lifestyle factors correlated with dietary habits (eg, physical activity and smoking) were not fully accounted for, which may have introduced residual confounding. Third, potential sample overlap between the exposure and outcome GWAS datasets cannot be entirely excluded, which may bias correlation-based estimates and inflate statistical significance. Fourth, although multiple complementary statistical approaches were applied, the influence of LD cannot be completely eliminated, and we did not conduct sensitivity analyses using alternative LD reference panels, which should be considered in future studies to further assess robustness. Fifth, these summary-statistic methods assume polygenicity, additive effects, and well-calibrated GWAS with appropriate LD references; our conclusions also depend on the quality of the original public GWAS summary statistics. Sixth, condFDR/conjFDR identifies genetic overlap (pleiotropic enrichment) and does not imply causality. Seventh, the analyses were restricted to individuals of European ancestry, limiting generalizability; replication in more diverse populations is warranted. Finally, because dedicated studies directly interrogating diet–gene–disease interactions remain limited, our interpretation of the implicated genes relied partly on evidence from other diseases, and further functional validation is needed.
Conclusion
In conclusion, this study identified genome-wide genetic correlations between PS and three dietary intake patterns (beer, cheese, and dried fruit intake), with genetic correlation coefficients ranging from 0.1082 to 0.2187, providing genomic evidence of shared genetic architecture between these traits. Through multi-level genomic integration analyses, we further identified several core shared loci, including RAPGEF6, CDKAL1, and KIF3A, which may implicate immune- and inflammation-related biological pathways relevant to PS. Notably, our findings reflect genetic overlap and correlation rather than causal dietary effects; therefore, any discussion of “dietary intervention” should be framed as genetically informed, hypothesis-generating evidence that warrants further causal inference and interventional validation, rather than as actionable clinical recommendations. Given that the analyses were restricted to individuals of European ancestry, validation in more diverse populations is warranted to improve generalizability. These findings can inform future Mendelian randomization, functional genomics, and personalized nutrition studies.
Data Sharing Statement
All the GWAS data and statistical software used in this study were publicly available (which can be accessed through the following URLs), and all the generated results in this study were provided in the main text and supplemental data.
IEU database: https://gwas.mrcieu.ac.uk/
LDSC:https://github.com/bulik/ldsc
GNOVA: https://github.com/xtonyjiang/GNOVA
HDL: https://github.com/zhenin/HDL
LAVA: https://github.com/josefin-werme/LAVA
ConjFDR: https://github.com/precimed/pleiofdr
FUMA: https://fuma.ctglab.nl
FinnGen: https://r12.finngen.fi/.
Ethics Statement
According to Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Beings issued by the National Science and Technology Ethics Committee of the People’s Republic of China, this study qualified for an exemption from ethical review because it used only open, legally accessible data, posed no risk to participants, and did not involve sensitive personal information or commercial interests.
Acknowledgments
The authors thank Bullet Edits Limited for the linguistic editing and proofreading of the manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study did not receive any funding in any form.
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construe as a potential conflict of interest.
References
1. Sieminska I, Pieniawska M, Grzywa TM. The immunology of psoriasis-current concepts in pathogenesis. Clin Rev Allergy Immunol. 2024;66:164–12. doi:10.1007/s12016-024-08991-7
2. Parisi R, Iskandar IYK, Kontopantelis E, Augustin M, Griffiths CEM, Ashcroft DM. Global Psoriasis Atlas. National, regional, and worldwide epidemiology of psoriasis: systematic analysis and modelling study. BMJ. 2020;369:m1590. doi:10.1136/bmj.m1590
3. Zhang Y, Dong S, Ma Y, Mou Y. Burden of psoriasis in young adults worldwide from the global burden of disease study 2019. Front Endocrinol. 2024;15:1308822. doi:10.3389/fendo.2024.1308822
4. AlQassimi S, AlBrashdi S, Galadari H, Hashim MJ. Global burden of psoriasis - comparison of regional and global epidemiology, 1990 to 2017. Int J Dermatol. 2020;59:566–571. doi:10.1111/ijd.14864
5. Hawkes JE, Chan TC, Krueger JG. Psoriasis pathogenesis and the development of novel targeted immune therapies. J Allergy Clin Immunol. 2017;140:645–653. doi:10.1016/j.jaci.2017.07.004
6. Zeng J, Luo S, Huang Y, Lu Q. Critical role of environmental factors in the pathogenesis of psoriasis. J Dermatol. 2017;44:863–872. doi:10.1111/1346-8138.13806
7. Musumeci ML, Nasca MR, Boscaglia S, Micali G. The role of lifestyle and nutrition in psoriasis: current status of knowledge and interventions. Dermatol Ther. 2022;35:e15685. doi:10.1111/dth.15685
8. Choi J, Han I, Min J, et al. Dose-response analysis between alcohol consumption and psoriasis: a systematic review and meta-analysis. JDDG: J Dtsch Dermatol Ges. 2024;22:641–652. doi:10.1111/ddg.15380
9. Xu Y, Zanesco S, Dalrymple KV, et al. Associations between food intake and psoriasis severity and comorbidity: a cross-sectional analysis using UK biobank data. Proc Nutr Soc. 2025;84:E226. doi:10.1017/S0029665125100955
10. Minghui L, Changyong G, Runtian Z, Jianhong L, Lingling Y, Xi C. The association between dietary consumption habits and psoriasis: a two-sample mendelian randomization study. Front Nutr. 2024;11:1405663. doi:10.3389/fnut.2024.1405663
11. Xu Y, Zanesco S, Zhang R, et al. Are psoriasis severity and comorbidities associated with diet quality? A cross-sectional analysis using UK biobank. Clin Nutr. 2025;50:198–209. doi:10.1016/j.clnu.2025.05.014
12. Ramos JKN, Gravet EH, Junger-Santos I, et al. Shared biological pathways linking ADHD and cortisol variability are related to externalizing behaviors. Psychoneuroendocrinology. 2025;181:107587. doi:10.1016/j.psyneuen.2025.107587
13. Chang S, Luo Q. Identification of overlapping genetic loci between inflammatory bowel disease and major depressive disorder. Eur Arch Psychiatry Clin Neurosci. 2025. doi:10.1007/s00406-025-02047-3
14. Huang Z, Yuan W. Exploring genetic structures and shared sites between alcohol, cheese intake, and inflammatory bowel disease. Front Nutr. 2025;12:1468457. doi:10.3389/fnut.2025.1468457
15. Bulik-Sullivan B, Finucane HK, Anttila V, Gusev A, Day FR, Loh P-R. ReproGen consortium, psychiatric genomics consortium, genetic consortium for anorexia nervosa of the wellcome trust case control consortium 3, Duncan l, et al. an atlas of genetic correlations across human diseases and traits. Nat Genet. 2015;47:1236–1241. doi:10.1038/ng.3406
16. Lu Q, Li B, Ou D, et al. A powerful approach to estimating annotation-stratified genetic covariance via GWAS summary statistics. Am J Hum Genet. 2017;101:939–964. doi:10.1016/j.ajhg.2017.11.001
17. Ning Z, Pawitan Y, Shen X. High-definition likelihood inference of genetic correlations across human complex traits. Nat Genet. 2020;52:859–864. doi:10.1038/s41588-020-0653-y
18. Werme J, Van der sluis S, Posthuma D, De Leeuw CA. An integrated framework for local genetic correlation analysis. Nat Genet. 2022;54:274–282. doi:10.1038/s41588-022-01017-y
19. Smeland OB, Frei O, Shadrin A, et al. Discovery of shared genomic loci using the conditional false discovery rate approach. Hum Genet. 2020;139:85–94. doi:10.1007/s00439-019-02060-2
20. Kurki MI, Karjalainen J, Palta P, et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature. 2023;613:508–518. doi:10.1038/s41586-022-05473-8
21. Watanabe K, Taskesen E, van Bochoven A, Posthuma D. Functional mapping and annotation of genetic associations with FUMA. Nat Commun. 2017;8:1826. doi:10.1038/s41467-017-01261-5
22. Maeta K, Edamatsu H, Nishihara K, Ikutomo J, Bilasy SE, Kataoka T. Crucial role of Rapgef2 and Rapgef6, a family of guanine nucleotide exchange factors for Rap1 small GTPase, in formation of apical surface adherens junctions and neural progenitor development in the mouse cerebral cortex. Eneuro. 2016;3:
23. Liang Y, Han D, Zhang S, Sun L. FOSL1 regulates hyperproliferation and NLRP3-mediated inflammation of psoriatic keratinocytes through the NF-kB signaling via transcriptionally activating TRAF3. Biochim Biophys Acta. 2024;1871:119689. doi:10.1016/j.bbamcr.2024.119689
24. Chen F, Zhu S, Kang R, Tang D, Liu J. ATP6V0D1 promotes alkaliptosis by blocking STAT3-mediated lysosomal pH homeostasis. Cell Reports. 2023;42. doi:10.1016/j.celrep.2022.111911
25. Stevens ML, Zhang Z, Johansson E, et al. Disease-associated KIF3A variants alter gene methylation and expression impacting skin barrier and atopic dermatitis risk. Nat Commun. 2020;11:4092. doi:10.1038/s41467-020-17895-x
26. Svanström C, Lonne-Rahm S-B, Nordlind K. Psoriasis and alcohol. Psoriasis. 2019;9:75–79. doi:10.2147/PTT.S164104
27. Qureshi AA, Dominguez PL, Choi HK, Han J, Curhan G. Alcohol intake and risk of incident psoriasis in US women: a prospective study. Arch Dermatol. 2010;146:1364–1369. doi:10.1001/archdermatol.2010.204
28. Jordan A, Näslund-Koch C, Vedel-Krogh S, Egil Bojesen S, Skov L. Alcohol consumption and risk of psoriasis: results from observational and genetic analyses in more than 100,000 individuals from the Danish general population. JAAD Int. 2024;15:197–205. doi:10.1016/j.jdin.2024.03.003
29. Szentkereszty-Kovács Z, Gáspár K, Szegedi A, Kemény L, Kovács D, Törőcsik D. Alcohol in psoriasis—from bench to bedside. Int J Mol Sci. 2021;22:4987. doi:10.3390/ijms22094987
30. Ishihara S, Nishikimi A, Umemoto E, Miyasaka M, Saegusa M, Katagiri K. Dual functions of Rap1 are crucial for T-cell homeostasis and prevention of spontaneous colitis. Nat Commun. 2015;6:8982. doi:10.1038/ncomms9982
31. Chiricozzi A, Guttman-Yassky E, Suárez-Fariñas M, et al. Integrative responses to IL-17 and TNF-α in human keratinocytes account for key inflammatory pathogenic circuits in psoriasis. J Invest Dermatol. 2011;131:677–687. doi:10.1038/jid.2010.340
32. Levy RJ, Kvajo M, Li Y, et al. Deletion of Rapgef6, a candidate schizophrenia susceptibility gene, disrupts amygdala function in mice. Transl Psychiatry. 2015;5:e577–e577. doi:10.1038/tp.2015.75
33. Li X, Yan Z, Lan H, et al. Genetic comorbidity of psoriasis and four cardiovascular diseases: uncovering shared mechanisms and potential therapeutic targets. Exp Dermatol. 2025;34:e70158. doi:10.1111/exd.70158
34. Afifi L, Danesh MJ, Lee KM, et al. Dietary behaviors in psoriasis: patient-reported outcomes from a U.S. national survey. Dermatol Ther. 2017;7:227–242. doi:10.1007/s13555-017-0183-4
35. Zheng X, Su Q, Wang Y, Geng X. Impact of serum calcium levels on the occurrence of sepsis and prognosis in hospitalized patients with concomitant psoriasis: a retrospective study based on the MIMIC-IV database. Front Immunol. 2025;16. doi:10.3389/fimmu.2025.1621231
36. Stanescu AMA, Simionescu AA, Diaconu CC. Oral vitamin D therapy in patients with psoriasis. Nutrients. 2021;13:163. doi:10.3390/nu13010163
37. Formisano E, Proietti E, Borgarelli C, Pisciotta L. Psoriasis and vitamin D: a systematic review and meta-analysis. Nutrients. 2023;15:3387. doi:10.3390/nu15153387
38. Palmer CJ, Bruckner RJ, Paulo JA, et al. Cdkal1, a type 2 diabetes susceptibility gene, regulates mitochondrial function in adipose tissue. Mol Metab. 2017;6:1212–1225. doi:10.1016/j.molmet.2017.07.013
39. Arruda AP, Hotamisligil GS. Calcium homeostasis and organelle function in the pathogenesis of obesity and diabetes. Cell Metab. 2015;22:381–397. doi:10.1016/j.cmet.2015.06.010
40. Ametaj BN. The calci-inflammatory network: a paradigm shift in understanding milk fever. Dairy. 2025;6:22. doi:10.3390/dairy6030022
41. Balić A, Vlašić D, Žužul K, Marinović B, Bukvić Mokos Z. Omega-3 versus omega-6 polyunsaturated fatty acids in the prevention and treatment of inflammatory skin diseases. Int J Mol Sci. 2020;21:741. doi:10.3390/ijms21030741
42. Zanesco S, Maruthappu T, Griffiths CEM, Dalrymple KV, Gibson R, Hall WL. Associations between diet quality indices and psoriasis severity: results from the asking people with psoriasis about lifestyle and eating (APPLE) cross-sectional study. Br J Nutr. 2025;133:546–557. doi:10.1017/S0007114525000340
43. Clark CCT, Taghizadeh M, Nahavandi M, Jafarnejad S. Efficacy of ω-3 supplementation in patients with psoriasis: a meta-analysis of randomized controlled trials. Clin Rheumatol. 2019;38:977–988. doi:10.1007/s10067-019-04456-x
44. Stevens ML, Mersha TB, Zhang Z, Kothari A, Khurana Hershey GK. Skin depletion of Kif3a resembles the pediatric atopic dermatitis transcriptome profile. Hum Mol Genet. 2022;31:1588–1598. doi:10.1093/hmg/ddab342
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.