Back to Journals » Drug, Healthcare and Patient Safety » Volume 16
Baloxavir Resistance Markers in Influenza A and B Viruses in the Americas
Authors Acocal-Juárez E ![]() , Márquez-Domínguez L
, Márquez-Domínguez L ![]() , Vallejo-Ruíz V
, Vallejo-Ruíz V ![]() , Cedillo L, Santos-López G
, Cedillo L, Santos-López G ![]()
Received 30 May 2024
Accepted for publication 20 July 2024
Published 14 September 2024 Volume 2024:16 Pages 105—113
DOI https://doi.org/10.2147/DHPS.S470868
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Hemalkumar B Mehta
Erick Acocal-Juárez,1– 3 Luis Márquez-Domínguez,2 Verónica Vallejo-Ruíz,2 Lilia Cedillo,3 Gerardo Santos-López2
1Centro de Investigaciones en Ciencias Microbiológicas, Benemérita Universidad Autónoma de Puebla, Puebla Pue, Mexico; 2Laboratorio de Biología Molecular y Virología, Centro de Investigación Biomédica de Oriente, Instituto Mexicano del Seguro Social, Puebla, Mexico; 3Centro de Detección Biomolecular, Benemérita Universidad Autónoma de Puebla México, Puebla Pue, Mexico
Correspondence: Lilia Cedillo; Gerardo Santos-López, Email [email protected]; [email protected]
Aim: Influenza control demands multifaceted strategies, including antiviral drugs. Baloxavir, a recent addition to influenza treatment, acts as an inhibitor of the Polymerase acid (PA) component of the viral polymerase. However, mutations associated with resistance have been identified.
Purpose: This study analyzed PA gene sequences of influenza A and B viruses (IAV and IBV, respectively) reported in the Americas, retrieved from databases published until May 2023, to identify primary markers of resistance to baloxavir.
Patients and Methods: PA gene sequences were obtained from the GISAID and NCBI databases, focusing on countries in the Americas with 500 or more sequences for IAV, and 50 or more sequences for IBV.
Results: Of the 58,816 PA sequences analyzed for IAV, only 55 (0.1%) harbored resistance markers, representing approximately 1 in 1000 occurrence. The most frequent markers were I38V (21 cases) and I38M (7 cases) at position 38 of PA, followed by E199G (9 cases) at position 199. For IBV, 14,684 sequences were analyzed, of which only eight presented a resistance marker (0.05%). Five sequences had the M34I marker, while the remaining three had the I38V marker. While frequency of resistance markers in PA is comparable to other regions, these results highlight the need for enhanced sequencing efforts, particularly in Latin America. Such efforts would serve to intensify influenza surveillance and inform public health interventions.
Conclusion: While baloxavir demonstrates efficacy against influenza, resistance markers have been identified, including pre-existing ones. Our study adds eight (IAV: six and IBV: two) new spontaneously occurring substitutions to the existing literature, highlighting the need for continued surveillance. Among these, I38M stands out due to its significant tenfold reduction in drug susceptibility. Therefore, vigilant monitoring of these resistance markers in IAV and IBV remains crucial for maintaining baloxavir’s effectiveness and informing future public health interventions.
Keywords: antiviral resistance, substitutions, PA protein, polymerase inhibitor
Introduction
Influenza viruses are classified into four types: A, B, C and D, based on variations in their internal proteins, the nucleoprotein and matrix protein 1. The genome of these viruses consists of segmented negative-sense RNA, comprising 8 segments in influenza A and B viruses, and 7 segments in influenza C and D viruses.1
Among these, Influenza A virus (IAV) holds the greatest significance for human health. It can be further classified into subtypes based on the genetic and antigenic properties of its surface glycoproteins, hemagglutinin (HA), and neuraminidase (NA). As of now, 18 HA types and 11 NA types have been identified.2
IAV is prone to genetic changes that can alter its antigenic and virulence characteristics, contributing to recurrent epidemics, sporadic pandemics, vaccine efficacy issues, and the emergence of antiviral resistance.3 The most devastating historical pandemic occurred in 1918, caused by an H1N1 subtype of Influenza A virus, resulting in over 50–100 million global fatalities.4,5 Presently, two major subtypes of the virus circulate: H3N2, responsible for the 1968 pandemic, and the pandemic H1N1 subtype (H1N1|pdm09), originating from the 2009 pandemic.5,6
On the other hand, influenza B viruses (IBV) have long been considered less problematic than influenza A due to their lower genetic variation, their status as an exclusive human pathogen with no known animal reservoir, and their lack of pandemic potential. However, in recent years, the relevance of influenza B has been recognized, especially in infants. Additionally, neuraminidase inhibitors are less effective against IBV infections compared to IAV infections.7,8 Two lineages of IBV are recognized: Victoria and Yamagata. These lineages differ in their transmission dynamics, evolution, cellular tropism, and susceptibility patterns, which may vary depending on the age of the host.9
Neuraminidase inhibitors, such as oseltamivir and zanamivir, are the most widely used treatment for IAV infections due to their ability to reduce disease duration and complications. Nevertheless, resistance to neuraminidase inhibitors has been reported in rates of 0.2–2.6%.10–12 Earlier antivirals like amantadine and rimantadine are no longer recommended due to widespread resistance among circulating influenza viruses.13
Recently, the new antiviral baloxavir marboxil was introduced. This prodrug, designed through molecular techniques, gained approval in Japan and the United States in 2018 for combating influenza A and B viruses. Baloxavir targets the endonuclease domain found in the polymerase acidic (PA) protein of influenza A and B viruses. Interestingly, this domain also exists in the P3 protein of influenza C and D viruses, and baloxavir has shown efficacy against these influenza types as well.14,15
Clinical studies involving baloxavir-based treatments for IAV infections have indicated that the virus can develop resistance by mutating amino acids I38 and E23 within the PA protein. Substitutions I38T or I38F significantly decrease baloxavir susceptibility by over tenfold, whereas the E23K substitution has a milder impact. Other substitutions may also influence baloxavir susceptibility.16 Regarding IBV, the I38T and I38M substitutions decrease susceptibility to baloxavir, although the effect is smaller than in IAV, with a reduction of less than 10-fold reported.12,16,17
It is important to identify these markers associated with drug resistance to current antivirals. Therefore, this study focuses on analyzing the PA protein sequences of viruses identified in the Americas over an approximately 43-year period, which spans many years before the original synthesis and authorization of baloxavir. The goal is to pinpoint the primary genetic markers associated with resistance or reduced effectiveness of baloxavir.
Materials and Methods
PA sequences of the IAV from all countries in the Americas were sourced from two databases: GISAID (https://platform.epicov.org/) and NCBI Influenza Virus Resource (https://www.ncbi.nlm.nih.gov/genomes/FLU/Database/nph-select.cgi#mainform). The downloaded PA sequences included all those available until the cutoff date of May 31, 2023. To be included in our analysis, sequences of IAV had to originate from countries with a minimum of 500 entries in the databases. Due to the small number of available sequences for IBV, we included sequences from countries with a minimum of 50 entries. Additionally, the sequences must cover at least the first 200 amino acids of the PA protein, which include all positions related to baloxavir resistance. Duplicate sequences or those failing to meet these criteria were excluded. For IAV, we focused on the following specific amino acid positions associated with resistance to the drug Baloxavir marboxil: E23, K34, A36, A37, E38, E119, E198, and E199. For IBV, we analyzed the positions E23, M34, F36, N37, I38, E120 (equivalent to E119 in IAV), and E194 (equivalent to E199). To analyze these selected sequences, we performed an alignment using ClustalW within the BioEdit software (version 7.2.5).
Results
Sequences
Influenza A virus. Sequences from eight American countries that met the specified criteria were included. In total, 58,816 sequences were recovered, distributed as follows: United States of America (USA): 49,850; Brazil: 2809; Canada: 1678; Chile: 1473; Peru: 886; Nicaragua: 793; Mexico: 742; and Argentina: 585. Sequences from the United States constituted 84.76% of the total (Figure 1A). Regarding viral subtypes, H3N2 had the highest frequency with 39,383 sequences, followed by pandemic H1N1 with 15,989, while H1N1 reported as unknown subtype were 2578 and seasonal H1N1 were only 616. Other subtypes were exceedingly rare (0.43%), with 60 sequences of H1N2 (55 in USA, 3 in Brazil and 2 in Canada), 43 sequences of H2N2 (41 in USA, 1 in Canada and 1 in Chile), 2 sequences of H7N3 (Canada and Mexico), 4 sequences of H5N1 (2 in USA, 1 in Chile and 1 in Brazil), 1 sequence of H7N9 (Canada), 1 sequence of H3N1 (Brazil) and 140 non-subtypable (Figure 1B).
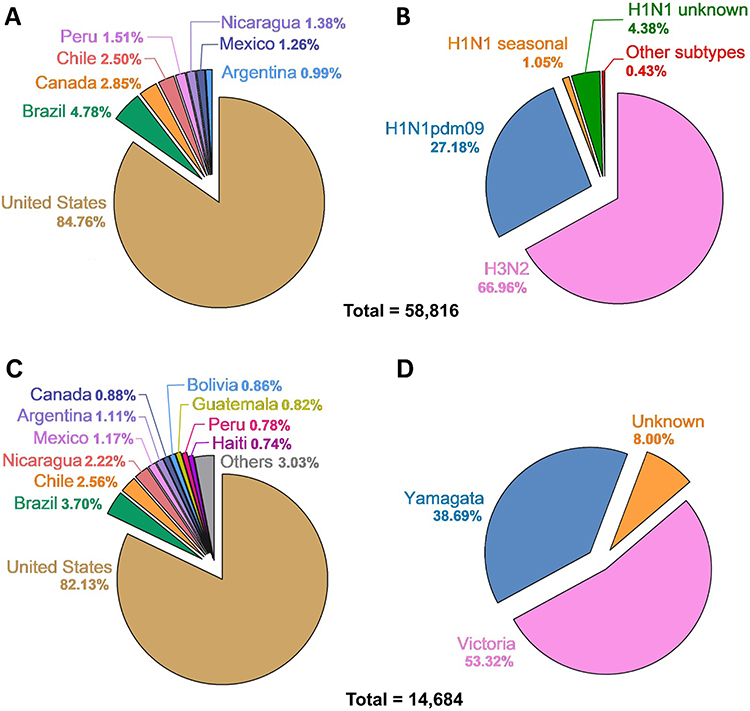 |
Figure 1 Distribution of influenza (A and B) viruses PA protein sequences analyzed through May 2023 (70,500 sequences). (A) shows the percentage distribution of Influenza A virus by country. (B) shows the percentage distribution of Influenza A virus by subtype. (C) illustrates the proportion of Influenza B virus sequences categorized by country. (D) depicts the proportion of Influenza B virus sequences categorized by lineage. |
Influenza B virus. Sequences from 17 American countries that met the specified criteria were included. In total, 14,684 sequences were recovered, distributed as follows: USA: 12,060; Brazil: 544; Chile: 376; Nicaragua: 326; Mexico: 172; Argentina: 163; Canada: 129; Bolivia: 126; Peru: 114; Haiti: 109; Guatemala: 120; El Salvador: 93; Puerto Rico: 93; Jamaica: 74; Paraguay: 68; Honduras: 67; and Colombia: 50. Sequences from the United States constituted 82.14% of the total (Figure 1C). Regarding viral lineages, Victoria had the highest frequency with 7829 sequences, followed by Yamagata with 5681, while Influenza B reported as unknown were 1174 (Figure 1D).
Baloxavir Resistance Markers for Influenza A Virus
Out of the 58,816 sequences, only 55 exhibited resistance markers, representing 0.1% or 1 per 1000 sequences. None of the sequences presented more than one resistance marker. The most frequently affected position was residue 38 of the PA protein, where sensitivity is indicated by the presence of I38. A total of 36 sequences showed a substitution associated with resistance at this position, with the main substitution being I38V found in 21 sequences, followed by I38M in 7 sequences. The second most common marker occurred at position 199, where the amino acid in sensitive viruses is E. This substitution was observed in 9 sequences, with the change noted as E199G (Table 1).
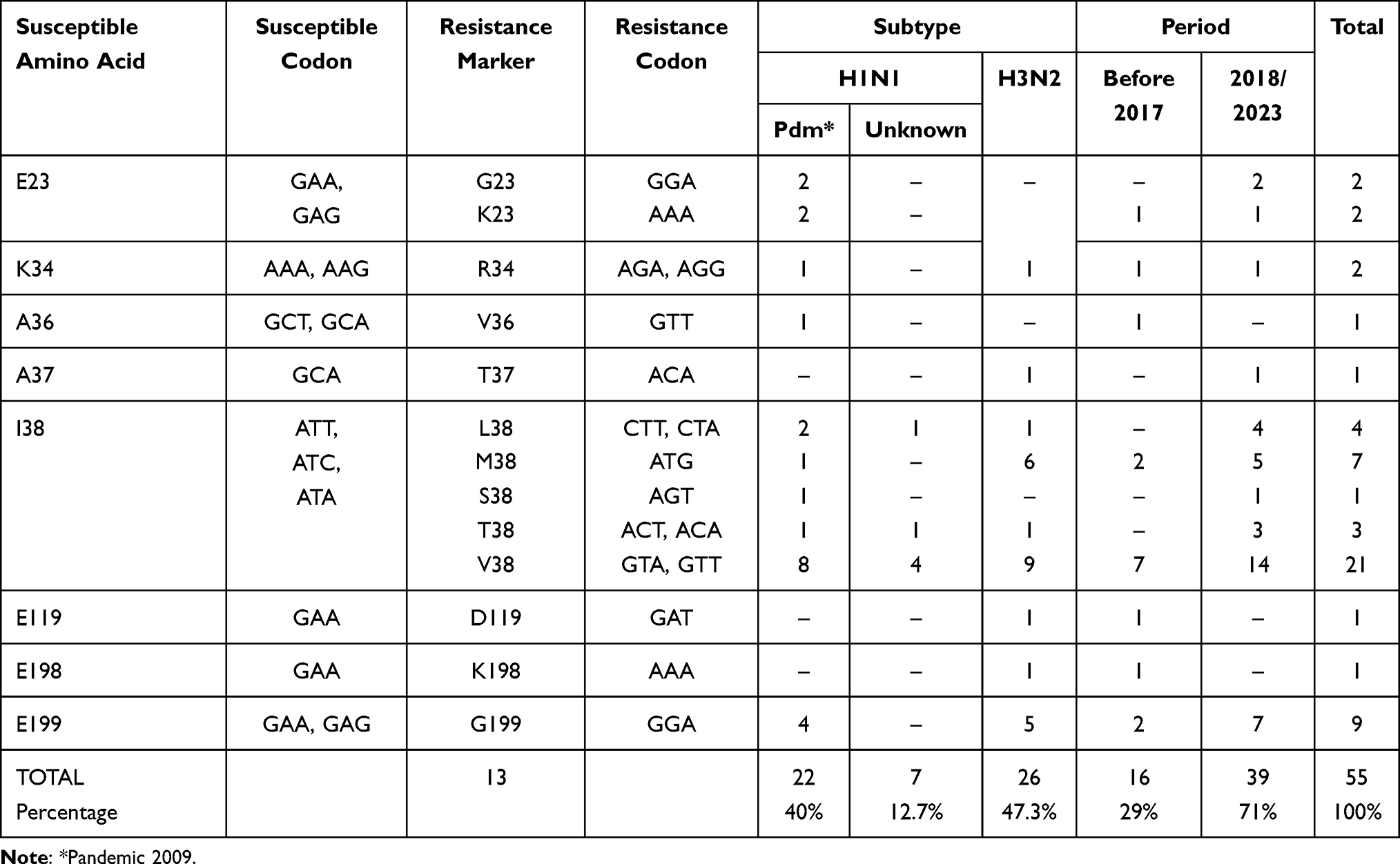 |
Table 1 Amino Acid Substitutions and Codons in PA Protein of Influenza A Viruses Associated with Reduced Susceptibility or Resistance to Baloxavir and Distribution by Subtype and Period |
The distribution of substitutions associated with resistance by country is as follows: USA: 50/49,850 (0.10%); Brazil: 3/2809 (0.10%); Canada: 0/1678 (0.00%); Chile: 1/1473 (0.07%); Peru: 0/886 (0.00%); Nicaragua: 0/793 (0.00%); Mexico: 1/742 (0.14%) and Argentina: 0/585 (0.00%). Resistance markers were more frequently observed in H3N2, with 26 out of 39,383 (0.07%) sequences, compared to pandemic H1N1, which had 22 out of 15,989 sequences (0.14%) and H1N1 unknown with 7 of 2578 (0.27%) sequences reported. In sequences of H1N1 seasonal, H1N2, H2N2, H3N1, H5N1, H7N2, H7N3 and H7N9 any resistance marker was observed (Figure 2).
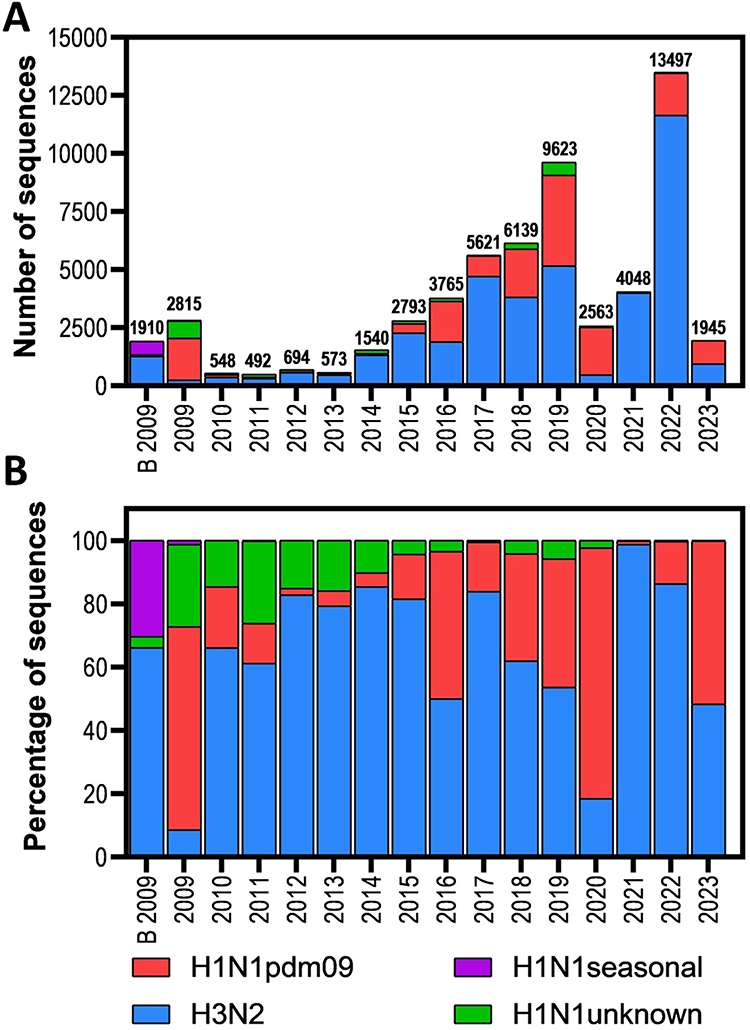 |
Figure 2 Distribution of PA protein sequences of influenza A viruses analyzed in the Americas (through May 2023). (A) Absolute number of sequences per year; (B) Relative number of sequences per year shown as a percentage. B 2009 – before 2009. |
Baloxavir was first licensed in 2018. Consequently, we used this year as a reference point for grouping the presence of resistance markers in the analyzed sequences. We identified thirteen different substitutions associated with resistance in eight positions along the PA protein. These substitutions were categorized as follows: In the period from 1990 to 2017, we detected 16 sequences out of 20,886 (0.08%) with resistance markers, namely E23K, K34R, A36V, I38M, I38V, E119D, E198K, and E199G. In the period from 2018 to 2023, we found 39 sequences out of 37,930 (0.11%) with resistance markers. Notably, five of these markers (E23G, A37T, I38L, I38S, and 138T) appeared exclusively within this period (Table 1 and Figure 3).
 |
Figure 3 Frequency of PA sequences of influenza A viruses with baloxavir resistance markers in the Americas through May 2023. (A) Total PA sequences (Orange) and with resistance markers (blue) per year; (B) Percentage of PA protein sequences containing specific amino acid substitutions associated with baloxavir resistance or reduced susceptibility. B 2009 – before 2009. |
Baloxavir Resistance Markers for Influenza B Virus
Out of the 14,684 sequences analyzed for influenza B, only eight presented a resistance marker, representing 0.05%, or 5 per 10,000 sequences. None of the sequences had more than one resistance marker. The affected positions were M34 and I38. Five sequences had the M34I marker, while the remaining three had the I38V marker (Table 2). The distribution of the resistance-associated substitutions was as follows: six sequences from the USA, one from Argentina, and one from Jamaica; no substitutions were found in other countries. Regarding lineage, seven sequences belonged to the Victoria lineage, one to Yamagata, and none were reported as unknown. In terms of date, we found two substitutions in sequences from 2016 (one with the M34I marker and another with I38V), and the remaining six substitutions were found in sequences from 2018 onwards.
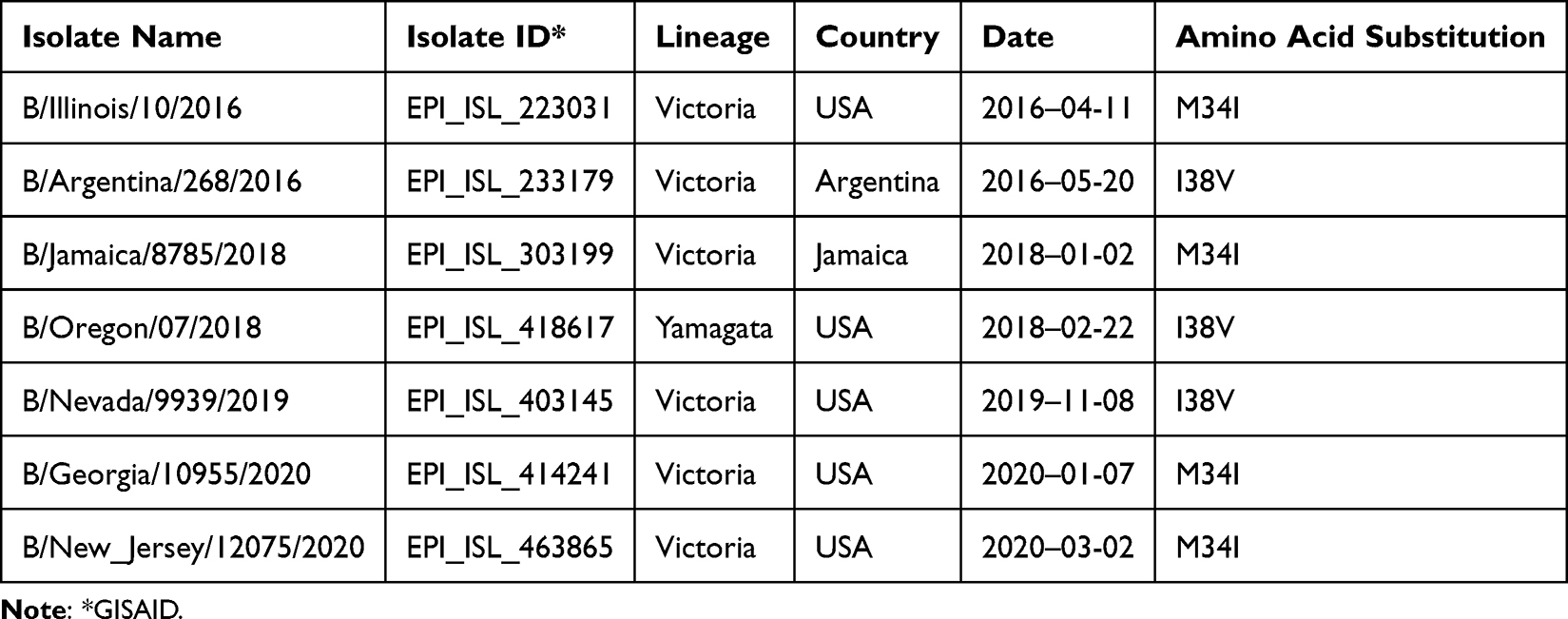 |
Table 2 Amino Acid Substitutions in the PA Protein of the Influenza B Virus Associated with Reduced Susceptibility or Resistance to Baloxavir |
Discussion
Drug resistance poses a significant challenge to public health, necessitating continuous monitoring and identification of its drivers. Baloxavir, a recent addition to influenza treatment, is of particular interest because resistance markers were identified even before its widespread use. However, the global impact of this issue remains incompletely understood, especially in low- and middle-income countries with limited influenza surveillance.
In the Americas, most available sequences (more than 82% for both IAV and IBV) originate from the USA, with limited representation from Latin America. This highlights the need for enhanced data collection efforts in these regions to improve our understanding of baloxavir resistance. Notably, more than 97% of analyzed sequences of IAV and IBV were collected from the 2009 A H1N1 pandemic emerging, highlighting a potential bias that might underestimate the true prevalence of resistance markers before that year.
Our data shows a frequency of baloxavir resistance markers in IAV in the Americas of approximately 1 in 1000 sequences, consistent with other studies worldwide. This low prevalence suggests that baloxavir remains effective against most influenza viruses. For instance, Gubareva et al analyzed 6891 virus sequences from the USA during 2016–2018 and found five sequences with resistance markers (0.7/1000 sequences);17 Takashita et al conducted a comprehensive study, analyzing 13,523 influenza virus sequences reported globally for the 2017–2018 season (from GISAID and NCBI Influenza Virus Resource), and identified 11 sequences with resistance markers (0.81/1000 sequences).18 In a more recent study, Govorkova et al performed a global analysis, including 31,598 sequences from five WHO regions during the 2018–2020 period, and reported 90 sequences carrying resistance markers (2.8 per 1000 sequences). Notably, sequences from Japan, where the administration of baloxavir is highest, showed a higher prevalence of resistance markers, reaching 45 per 1000 sequences. The authors suggest that this elevated level of resistance could be related to the extensive use of baloxavir in Japan, particularly among children older than 12 years.12
Of the resistance markers found in the 55 PA sequences, 16 were reported prior to the introduction of baloxavir in 2018, while other 39 were identified from 2018 onwards (Figure 3A). The percentage of markers identified relative to the total sequences reported in the analyzed periods is 0.08% for the 1990–2017 period and 0.11% for the 2018–2022 period. This finding emphasizes the importance of understanding the evolutionary history of these markers and the potential for their circulation independent of drug pressure.
Among the 16 sequences identified prior to 2018, we observed eight different substitutions: E23K, K34R, A36V, I38M, I38V, E119D, E198K, and E199G. Notably, residues E23, K34, A36, I38, and E119 are situated within the PA endonuclease site.16 Of these markers, it is crucial to note that viruses containing the E23K substitution can be transmitted from person to person.19,20 Position 38M is particularly noteworthy due to reports indicating that it can reduce the viral susceptibility to baloxavir by more than tenfold.12,16,21
E198K has recently been identified as a marker associated with resistance to baloxavir. This marker was identified after passages in MDCK cells in the presence of the drug.22
Trypsin digestion of PA reveals two domains: the N-terminal containing the first 196 amino acids, and the C-terminal containing residues from 277 to 716. In the viral ribonucleoprotein complex, both domains are found on opposite faces, connected by an 81-amino acid linker.19,23,24 The residue E198 is part of the linker region and is not directly involved in the binding of baloxavir to the active site; however, it does contribute to viral polymerase activity.20 Additionally, E199G is located at the outset of the linker site in the PA protein. Its specific role in conferring resistance to baloxavir remains unclear.17,25,26
To our knowledge, the only naturally occurring mutations that have been documented are A36V and I38V.12,16 A36V reduces susceptibility to baloxavir by four times and has been reported to appear in subtype H1N1 in 0.03% of the data deposited in the NCBI database.16 We found this marker in only one H1N1 virus (A/Maryland/NHRC0004/2009), accounting for 0.0018% of the total sequences analyzed and 0.005% of the subtype H1N1 cases. On the other hand, I38V results in a minor alteration in baloxavir susceptibility, ranging from 1 to 4 times.12,16 We identified 21 sequences carrying this mutation, representing 0.04% of the total sequences examined and constituting 38% of all the mutations found. These findings collectively indicate that, while most sequences lack markers associated with resistance to baloxavir, there is a need to enhance molecular surveillance of influenza viruses in extensive geographic regions such as Latin America. This will contribute to a more comprehensive global dataset, enabling vigilance against potential issues of drug resistance.
In the case of IBV, the studied sequences represent 25% of the total analyzed, which aligns with previous studies reporting that IBV infections account for approximately one-fourth of all influenza cases.27 Due to the smaller number of IBV sequences susceptible to analysis, we decided to include countries with at least 50 sequences deposited in banks, unlike IAV sequences, in which we included countries that had at least 500. The number of sequences with any resistance marker represents only 0.05% of the total IBV sequences analyzed, which is half the proportion of sequences with resistance markers compared to IAV. The markers associated with a greater reduction in baloxavir susceptibility IBV are I38T and I38M; however, we did not find any sequences with these markers. We found sequences with the M34 and I38 markers, but these changes do not appear to significantly reduce baloxavir susceptibility by in vitro assays.12,16,17 Since these sequences were present before the introduction of baloxavir, we can assume that these mutations occur spontaneously. Of the sequences with markers, only one belongs to the Yamagata lineage (14%) and seven to the Victoria lineage (86%), which is consistent with previous studies indicating that the Yamagata lineage maintains more stable genetic diversity.9
Regarding the fitness of IAV and IBV strains with the I38X mutation, the results are inconclusive. While some studies have observed a reduction in fitness,28 others have shown an improvement compared to the wild type.28,29 Therefore, we should be cautious in drawing conclusions and await further studies.
Conclusion
While baloxavir demonstrates efficacy against influenza, resistance markers have been identified, including pre-existing ones. Our study adds eight additional spontaneously occurring markers to the existing literature, highlighting the need for continued surveillance. Among these, I38M stands out due to its significant tenfold reduction in drug susceptibility. Therefore, vigilant monitoring of these markers in influenza A and B viruses remains crucial for maintaining baloxavir’s effectiveness and informing future public health interventions.
Abbreviations
IAV, Influenza A virus; IBV, Influenza B virus; PA, Polymerase acid; HA, hemagglutinin; NA, neuraminidase; H1N1|pdm09, pandemic H1N1 subtype; USA, United States of America.
Ethical Approval Statement
This study did not involve any experiments with human participants or animals. All data used in this research were obtained from publicly available databases that do not contain any sensitive personal information. As such, no ethical approval was required.
Acknowledgments
Erick Acocal Juárez had a scholarship from CONAHCYT (No. 689191).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Wang M, Veit M. Hemagglutinin-esterase-fusion (HEF) protein of influenza C virus. Protein Cell. 2016;7:28–45. doi:10.1007/s13238-015-0193-x
2. Uyeki TM, Hui DS, Zambon M, et al. Influenza. Lancet. 2022;400:693–706. doi:10.1016/S0140-6736(22)00982-5
3. Xue KS, Bloom JD. Linking influenza virus evolution within and between human hosts. Virus Evol. 2020;6:veaa010. doi:10.1093/ve/veaa010
4. Schrauwen EJ, Graaf M, Herfst S, et al. Determinants of virulence of influenza A virus. Eur J Clin Microbiol Infect Dis. 2014;33(4):479–490. doi:10.1007/s10096-013-1984-8
5. Gilbertson B, Subbarao K. What have we learned by resurrecting the 1918 influenza virus? Annu Rev Virol. 2023;10:25–47. doi:10.1146/annurev-virology-111821-104408
6. Mostafa A, Pleschka S. Influenza H3N2 vaccines: recent challenges. Trends Microbiol. 2018;26:87–89. doi:10.1016/j.tim.2017.12.003
7. Zaraket H, Hurt AC, Clinch B, et al. Burden of influenza B virus infection and considerations for clinical management. Antiviral Res. 2021;185:104970. doi:10.1016/j.antiviral.2020.104970
8. Koutsakos M, Kent SJ. Influenza B viruses: underestimated and overlooked. Microbiol Aust. 2021;42(3):110–115. doi:10.1071/MA21033
9. Vijaykrishna D, Holmes EC, Joseph U, et al. The contrasting phylodynamics of human influenza B viruses. Elife. 2015;4:e05055. doi:10.7554/eLife.05055
10. Thorlund K, Awad T, Boivin G, et al. Systematic review of influenza resistance to the neuraminidase inhibitors. BMC Infect Dis. 2011;11(1):134. doi:10.1186/1471-2334-11-134
11. Lampejo T. Influenza and antiviral resistance: an overview. Eur J Clin Microbiol Infect Dis. 2020;39:1201–1208. doi:10.1007/s10096-020-03840-9
12. Govorkova EA, Takashita E, Daniels RS, et al. Global update on the susceptibilities of human influenza viruses to neuraminidase inhibitors and the cap-dependent endonuclease inhibitor baloxavir, 2018–2020. Antiviral Res. 2022;200:105281. doi:10.1016/j.antiviral.2022.105281
13. Pizzorno A, Abed Y, Boivin G. Influenza drug resistance. Semin Respir Crit Care Med. 2011;32:409–422. doi:10.1055/s-0031-1283281
14. Mishin VP, Patel MC, Chesnokov A, et al. Susceptibility of Influenza A, B, C, and D Viruses to Baloxavir(1). Emerg Infect Dis. 2019;25:1969–1972. doi:10.3201/eid2510.190607
15. Takashita E, Hu S, Zhu Y, et al. Antiviral Susceptibilities of Distinct Lineages of Influenza C and D Viruses. Viruses. 2023;16:15. doi:10.3390/v16010015
16. Omoto S, Speranzini V, Hashimoto T, et al. Characterization of influenza virus variants induced by treatment with the endonuclease inhibitor baloxavir marboxil. Sci Rep. 2018;8:9633. doi:10.1038/s41598-018-27890-4
17. Gubareva LV, et al. Assessing baloxavir susceptibility of influenza viruses circulating in the United States during the 2016/17 and 2017/18 seasons. Euro Surveill. 2019;2019:24.
18. Takashita E, Daniels RS, Fujisaki S, et al. Global update on the susceptibilities of human influenza viruses to neuraminidase inhibitors and the cap-dependent endonuclease inhibitor baloxavir, 2017-2018. Antiviral Res. 2020;175:104718. doi:10.1016/j.antiviral.2020.104718
19. Takashita E, Abe T, Morita H, et al. Influenza A(H1N1)pdm09 virus exhibiting reduced susceptibility to baloxavir due to a PA E23K substitution detected from a child without baloxavir treatment. Antiviral Res. 2020;180:104828. doi:10.1016/j.antiviral.2020.104828
20. Jones JC, Zagribelnyy B, Pascua PNQ, et al. Influenza A virus polymerase acidic protein E23G/K substitutions weaken key baloxavir drug-binding contacts with minimal impact on replication and transmission. PLoS Pathog. 2022;18:1010698. doi:10.1371/journal.ppat.1010698
21. Koszalka P, Tilmanis D, Roe M, et al. Baloxavir marboxil susceptibility of influenza viruses from the Asia-Pacific, 2012-2018. Antiviral Res. 2019;164:91–96. doi:10.1016/j.antiviral.2019.02.007
22. Takizawa N, Momose F. A novel E198K substitution in the PA gene of influenza A virus with reduced susceptibility to baloxavir acid. Arch Virol. 2022;167:1565–1570. doi:10.1007/s00705-022-05456-0
23. Ikematsu H, Hayden FG, Kawaguchi K, et al. Baloxavir marboxil for prophylaxis against influenza in household contacts. N Engl J Med. 2020;383:309–320. doi:10.1056/NEJMoa1915341
24. Guu TS, Dong L, Wittung-Stafshede P, et al. Mapping the domain structure of the influenza A virus polymerase acidic protein (PA) and its interaction with the basic protein 1 (PB1) subunit. Virology. 2008;379:135–142. doi:10.1016/j.virol.2008.06.022
25. Hayden FG, Sugaya N, Hirotsu N, et al. Baloxavir Marboxil for Uncomplicated Influenza in Adults and Adolescents. N Engl J Med. 2018;379:913–923. doi:10.1056/NEJMoa1716197
26. Kiso M, Yamayoshi S, Murakami J, et al. Baloxavir Marboxil Treatment of Nude Mice Infected With Influenza A Virus. J Infect Dis. 2020;221:1699–1702. doi:10.1093/infdis/jiz665
27. Caini S, Kusznierz G, Garate VV, et al. The epidemiological signature of influenza B virus and its B/Victoria and B/Yamagata lineages in the 21st century. PLoS One. 2019;14(9):e0222381. doi:10.1371/journal.pone.0222381
28. Abed Y, Fage C, Checkmahomed L, et al. Characterization of contemporary influenza B recombinant viruses harboring mutations of reduced susceptibility to baloxavir marboxil, in vitro and in mice. Antivir Res. 2020;179:104807. doi:10.1016/j.antiviral.2020.104807
29. Jones JC, Pascua PNQ, Fabrizio TP, et al. Influenza A and B viruses with reduced baloxavir susceptibility display attenuated in vitro fitness but retain ferret transmissibility. Proc Natl Acad Sci U S A. 2020;117:
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.