Back to Journals » Drug Design, Development and Therapy » Volume 14
Baicalin Ameliorates Dexamethasone-Induced Osteoporosis by Regulation of the RANK/RANKL/OPG Signaling Pathway
Authors Zhao Y, Wang HL, Li TT, Yang F, Tzeng CM
Received 31 July 2019
Accepted for publication 2 December 2019
Published 15 January 2020 Volume 2020:14 Pages 195—206
DOI https://doi.org/10.2147/DDDT.S225516
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Qiongyu Guo
Ye Zhao, 1, 2 Hui-Ling Wang, 1 Tong-Tong Li, 1 Fei Yang, 1 Chi-Meng Tzeng 1
1School of Pharmaceutical Sciences, Nanjing Tech University, Nanjing 211800, People’s Republic of China; 2Jiangsu Synergetic Innovation Center for Advanced Bio-Manufacture, Nanjing Tech University, Nanjing 211800, People’s Republic of China
Correspondence: Ye Zhao; Chi-Meng Tzeng Email [email protected]; [email protected]
Background: Osteoporosis is a chronic bone metabolism disorder affecting millions of the world population. The RANKL/RANK/OPG signaling pathway has been confirmed to be the main regulator of osteoporosis. It is of great interest to identify appropriate therapeutic agents that can regulate the RANKL/RANK/OPG pathway. Baicalin (BA) is a well-known traditional Chinese medicine formula against various inflammatory diseases with a proven role of the RANKL/RANK/OPG pathway regulation. However, the potential effect of BA on osteoporosis and the mechanisms underlying this remain unclear. In the present study, we aimed to evaluate the efficacy of BA in the prevention of dexamethasone (DEX)-induced osteoporosis in zebrafish.
Methods: In this study, growth and development changes of zebrafish and calcein staining were assessed with a micrograph. The expression levels of RANKL and OPG and transcription factors in response to DEX induction and BA administration were evaluated by Western blotting and qRT-PCR. In addition, the intermolecular interactions of BA and RANKL were investigated by molecular docking.
Results: Results show that BA enhances the growth and development of dexamethasone (DEX)-induced osteoporosis in zebrafish larvae. Calcein staining and calcium and phosphorus determination revealed that BA ameliorates mineralization of DEX-induced osteoporosis zebrafish larvae. BA also regulates the expression of RANKL and OPG and hampers the changes in gene expression related to bone formation and resorption under the induction of DEX in zebrafish. It can be inferred by molecular docking that BA may interact directly with the extracellular domain of RANKL.
Conclusion: The findings, herein, reveal that BA ameliorates DEX-induced osteoporosis by regulation of the RANK/RANKL/OPG signaling pathway.
Keywords: Baicalin, osteoporosis, RANKL, RANK, OPG
Introduction
Osteoporosis, a chronic bone metabolism disorder characterized by bone loss and structural deterioration of bone tissue, affects the life quality of millions of the world population.1 Several interacting factors contribute to the risk of osteoporosis and fracture, including aging, hormonal factors, low physical activity, lack of calcium or vitamin D, medical treatments such as glucocorticosteroids and genetic variables.2 As known, bone mass is maintained by a dynamic balance between bone resorption and bone formation. In the bone homeostasis, osteoclasts, osteoblasts and osteocytes play prominent roles. Osteoclasts are the dominated cells performing bone resorption while osteoblasts and osteocytes function in bone formation.3 Generally, imbalances occur when osteoclastic bone resorption outstrips osteoblastic bone synthesis. Hyper‐activity of osteoclasts differentiation that resorbs bone mass leads to the pathogenesis of osteoporosis.3 Conversely, osteoclasts with hypo‐activity increase bone mass and result in osteosclerosis. Therefore, identification of the dual-direction of physiological and pathological regulations of osteoclasts is critical for treating bone diseases.
It has been found that the receptor activator of nuclear factor-B ligand (RANKL), a member of the tumor necrosis factor (TNF) ligand family, can bind to its receptor RANK on the surface of osteoclasts and stimulate osteoclast maturation and differentiation.4,5 The action of RANKL is blocked by osteoprotegerin (OPG), which prevents the binding of RANKL to the RANK receptor and subsequently inhibits bone resorption.6,7 Overall, RANKL is the key component of the RANK/RANKL/OPG pathway, which is critical in the regulation of bone remodeling.8,9 Substantial evidences have revealed the potential of RANKL-targeted therapies to treat osteoclast-mediated bone loss diseases.10 For example, clinical trials indicated that RANKL inhibitor denosumab can reduce the rate of bone remodeling, thereby increasing bone mineral density and strength, and reducing the risk of fractures in women with postmenopausal osteoporosis.11 Studies have proved that melatonin suppresses bone loss in the inhibition of RANKL pathway.7,12–14 OPG was also used as a therapy candidate to block RANKL which was proved to be successful.15 Estrogen has been reported to increase OPG levels, suggesting that estrogen can be a possible therapy solution for modulation of bone resorption.16 By binding to RANK receptor, RANKL drives downstream NF‐κB pathway, and mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (ERK)/p38/c‐Jun N‐terminal kinase (JNK) pathways to stimulate the expression of key osteoclastic transcriptional factors such as c‐Fos and nuclear factor of activated T‐cells 1 (NFATc1), and consequently impels the formation of functional mature osteoclasts.17–20
In recent decades, a great deal of effort has been made toward developing drugs to treat and prevent osteoporosis. Flavonoids derived from natural herbs have aroused growing favored owning to its beneficial effects and minimal side effects.21 Baicalin (BA), also known as 7-glucuronic acid-5,6-dihydroxy-flavone, is a flavonoid isolated from Scutellaria baicalensis Georgi (Lamiaceae) and is broadly used as a herbal treatment for various inflammatory diseases in traditional Chinese medicine.22,23 Previous studies revealed that BA exhibits widely potent biological activities, including anti-inflammatory,24 antiviral25 and antitumor effects,26 etc. BA has been reported to suppress inflammation by inhibiting NF‐κB,27,28 which plays an essential role in the differentiation and function of osteoclasts. In human periodontal ligament cells, BA upregulates OPG expression and downregulates RANKL expression.29,30 Additionally, BA has also been found to enhance the mRNA expression of OPG and promote osteoblastic differentiation via the Wnt/β-catenin signaling pathway.31 These encourage findings hint that BA might regulate the activity of osteoclasts. Herein, we investigated the effect of BA on osteoporosis with a focus on the regulation of the RANK/RANKL/OPG signaling pathway.
Materials and Methods
Animals and Chemical Treatments
Zebrafish were maintained according to the compliance with Institutional Animal Care and Use Committee (IACUC) guidelines of Nanjing Tech University (Nanjing, China) for laboratory animal use. The study was approved by the ethics committee of Nanjing Tech University. Adult wild-type zebrafish (AB line) were obtained from Model Animal Research Center of Nanjing University (Nanjing, China). Fish were maintained in a closed flow-through system under the standardized conditions of temperature (28 ± 0.5 °C), pH (7.0 ± 1.0), and in a 14-hr light/10-hr dark rotation as reported.32 Fish were fed twice daily with live brine. Fish spawning was stimulated by the onset of light artificially. Normally fertilized embryos were selected for culturing and for experiment.
Baicalin (BA), etidronate disodium (ED) and dexamethasone (DEX) were purchased from the Chinese National Institute for the Control of Pharmaceutical and Biological Products. The final concentration of DMSO in experimental solutions did not exceed 0.01%. Chemical treatments were conducted in 6-well plates for zebrafish embryos from 0 to 8 or 9 dpf (day post fertilization) and in 1000 mL-tanks for larvae fish after 8 dpf. For exposure experiments, six groups were divided, including vehicle control (DEX-), DEX-induced model group (DEX+) with 25 μM/L DEX treatments, co-administration of DEX and ED with a concentration of 0.12 uM/mL, BA treatments with low concentration of 0.22 μM/L (DEX-BA(L)+) and high concentration of 11 μM/L (DEX-BA(H)+), co-administration of DEX and both concentrations of BA (DEX+BA(L)+), (DEX+BA(H)+). The dose levels were determined based on preliminary experiments. In the preliminary experiment, zebrafish larvae were treated by BA with concentrations of 0.1μg/mL (0.22 μM/L), 1μg/mL (2.2 μM/L), 5μg/mL (11μM/L). BA with the concentration of 0.22 μM/L begun to take effect and the concentration of 11 μM/L did not show higher toxic than lower doses.
During the chemical treatment, incubation solutions were renewal daily for zebrafish embryos. For zebrafish larvae, exposure solutions were renewed every 3 days, and they were fed with commercial nutritionally balanced artificial fish meal (Suzhou Green Blue Pet Products Co., Ltd., China).
Mineral Analyses
For detection of calcium and phosphorus contents, zebrafish embryos at 9 dpf were collected and immobilizated by submersion in ice water (5 parts ice/1 part water, 0–4º C) for 20 mins after cessation of all movement to ensure death by hypoxia. The method of zebrafish euthanasia was following the Guidelines for Use of Zebrafish in the NIH Intramural Research Program. After the fish dead, the carcasses were dried and subsequently dissolved in concentrated nitric acid (70%, Sigma-Aldrich) following with dilution 100-fold with demineralized water. Each group contains 10 fish. Calcium and phosphorus contents were analyzed by inductively coupled plasma optical emission spectrophotometry.
Calcein Staining
Calcein staining is a fluorescent dye used for detection of calcium.33 Zebrafish embryos at 9 dpf were collected and carried out euthanasia with the same method above and immersed in 1% calcein (CAS Number: C0875, Sigma USA) working solution for 15 mins and later washed by water three times (each time 5 mins), until the eluent was colorless. Fish were taken out and kept in glycerin. The fluorescent signal of the calcein-stained vertebrate column was observed under fluorescence microscopy (Nikon eclipse Ti, Tokyo, Japan) (Magnification 10×, excitation 480 ± 40 nm; emission 510 nm).
Western Blotting
Protein samples of zebrafish larvae were extracted with lysis buffer and separated in the 12% SDS-polyacrylamide gel electrophoresis (PAGE). The samples were then transferred onto a nitrocellulose membrane and blocked by TBS containing 10% skim milk. The membranes were incubated with mouse anti-RANKL antibody (1:1000 dilutions, ab198609; Abcam, USA) or rabbit anti-OPG antibody (ab183910; Abcam, USA) at 1:2000 dilutions in TBS containing 1% skim milk; then the membrane was washed with TBS and incubated with secondary antibodies, HRP-conjugated goat anti-mouse or goat anti-rabbit IgGs (1:2000 dilutions, Zhongshanjinqiao Co. China) for 2 hrs at RT. The experiments were repeated three times with different samples. The densities of the bands were determined using image analysis software Gel-pro Analyzer 4.5.
RNA Isolation and qRT-PCR
Ten larvae were randomly pooled as one replicate at the termination of experiments and were maintained in Trizol reagent (Invitrogen, USA) at −80 °C. Total RNA was isolated using Trizol reagent and digested using RNase-free DNaseI (Promega, USA). The purity of RNA samples was verified by the A260/A280 ratio detection and confirmed by 1% agarose-formaldehyde gel electrophoresis. The purified RNA was immediately reverse-transcribed using the SuperScript II cDNA Synthesis Kit (Invitrogen). QRT-PCR was performed through LightCycler ® 480 SYBR Green I Master system (Roche, USA). Each sample was analyzed with triplicate. β-Actin mRNA was used as the internal control for calibration of gene expression levels between samples. Log2 fold changes between treatment and un-treatment control samples were calculated by the -∆∆CT method. Primers used for PCR are shown in Table 1.
 |
Table 1 PCR Primer Sequences |
Molecular Docking
The Flexible Docking tool of Discovery Studio 3.5 software (DS 3.5, Accelrys, San Diego, CA, USA) was used to perform the automated molecular docking. This 3D crystal structure of RANKL, human RANKL and four cysteine-rich regions on human OPG (cysteine-rich domain, CRD) combined with the formed complex (RANKL/OPG- CDR) is obtained from PDB (http://www.pdb.org/, PDB ID: 3URF). Structures of RANKL and BA were performed optimization (hydrogenation, addition of CHAEMm force field and Momany-Rone charge, and structural optimization); Evaluation of the molecular docking was performed according to the -CDOCKER_INTERACTION_ENERGY. According to the binding-energy, the best pose for each peptide was obtained. DS 3.5 was used to identify the hydrogen bonds, the hydrophobic, hydrophilic, electrostatic, van der Waals forces and coordination interactions between BA and residues at the RANKL-active site.
Statistical Analyses
Statistical analyses were performed by one-way analysis of variance (ANOVA) with Dunnett’s post hoc test using InStat version 162 5.0 for Windows (GraphPad, San Diego, CA). Gene transcription levels were expressed as fold changes relative to the reference gene. The means and standard deviations are derived from at least triplicates. Data in bars represent mean ± SD in histograms and line chart. Zebrafish larvae for measurement were randomly collected in all experiments and differences were considered statistically significant at P < 0.05.
Results
BA Enhances Growth and Development of Dexamethasone (DEX)-Induced Osteoporosis Zebrafish Larvae
Dexamethasone (DEX)-induced osteopenia zebrafish larvae has been proved to be a well-established animal model for study of osteoporosis.34 In this study, zebrafish larvae at 96 hpf (hours post fertilization) were exposed to DEX solutions with a concentration of 25 μM/L (DEX+), and 0.01% DMSO (DEX-) was selected as the vehicle control. Etidronate disodium (ED) is known to reduce bone resorption through the inhibition of osteoclastic activity.35 Therefore, ED-treated group with a concentration of 0.12 uM/mL was chosen as the positive control (DEX+ED+). BA of 0.22 μM/L and 11 μM/L were respectively used as low and high concentrations-treated groups, BA(L)+ and BA(H)+. All groups were incubated in 6-well plates until 8 dpf or 60 dpf. To evaluate the potential positive effect of BA on embryonic development of DEX-induced osteopenia zebrafish, we analyzed the common growth and developmental phenotype including body length, hatching rate, mortality rate for 8 days and percent survival for 60 days. Figure 1A shows that body length at 96 hpf of DEX-treated embryos (DEX+) was significantly shortened comparing with untreated ones (DEX-) (P<0.05). Meanwhile, BA(H) significantly rescued the body length changes comparing with DEX- (P<0.05). The tendency is consistent with ED-treated group (ED+). Under the treatment of DEX, the hatching rate of embryos at 72 hpf was decreased, while BA(L) and BA(H) enhanced the hatching rate significantly comparing with DEX+ (P<0.05) (Figure 1B). Figure 1C demonstrates that the mortality rate of BA (L) and BA (H), as well as positive control (ED)-treated osteoporosis model group was reduced at 8 dpf comparing with DEX+. Additionally, we observed that the mortality rate of BA (L)- and BA (H)-treated normal embryos was higher than untreated ones, suggesting the slight toxicity effect of BA on embryos. Therefore, percent survival for 60 days was further evaluated to detect the chronic toxicity of BA, and to investigate the long-term effect of BA on DEX-induced osteoporosis. Results shown that the percent survival for 60 days was improved in BA (L)- and BA (H)-treated groups than DEX+ (Figure 1D). Additionally, BA has slight and chronic toxicity t for development of embryos. In all, BA enhances the growth and development of DEX-induced osteoporosis zebrafish larvae.
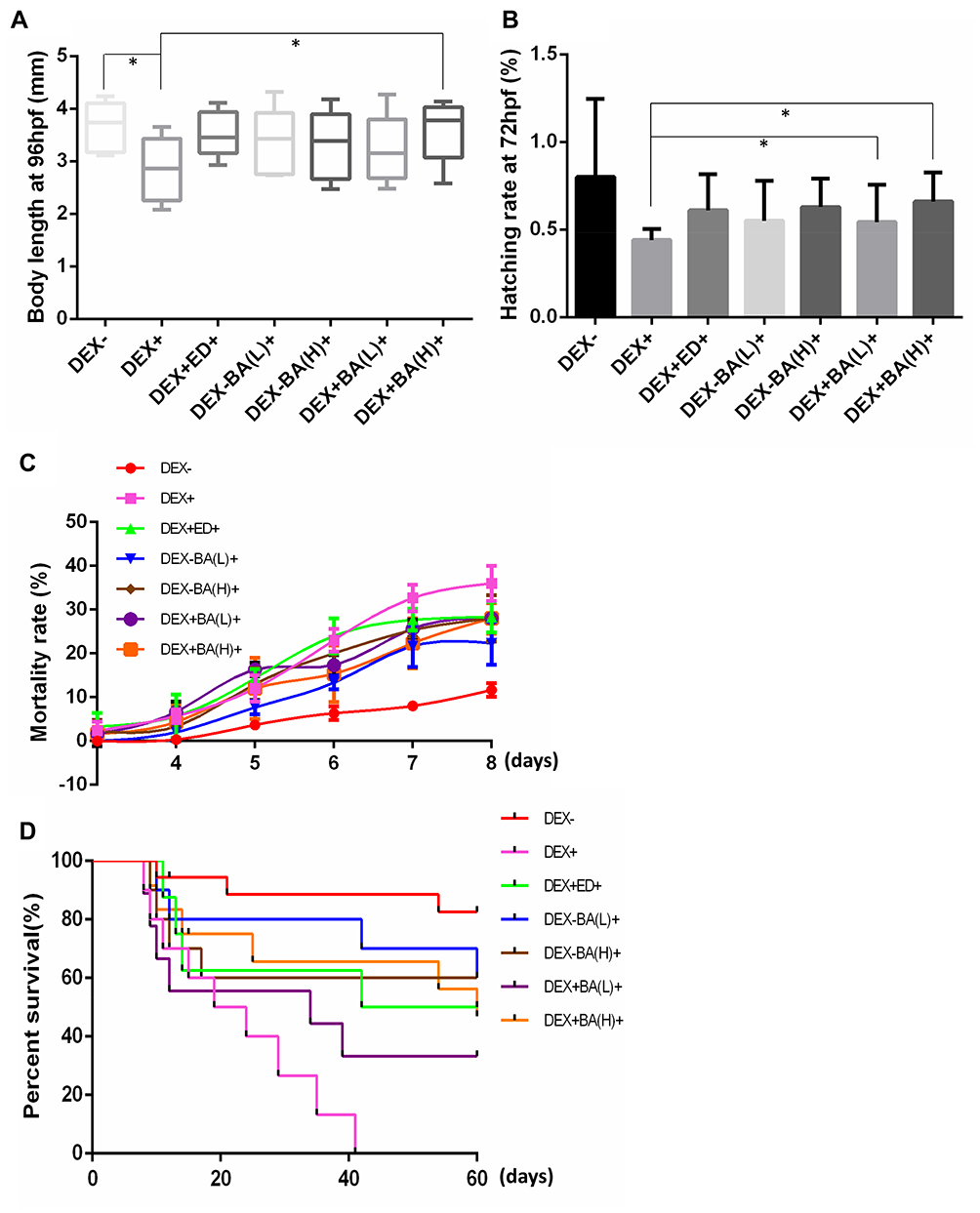 |
Figure 1 Effects of BA on growth and development of dexamethasone (DEX)-induced osteoporosis zebrafish larvae. (A) Body length of zebrafish embryos at 96 hpf (hours post fertilization); (B) Hatching rate of embryos at 72 hpf; (C) Mortality rate for 8 days after fertilization. (D) Percent survival for 60 days after fertilization. The quantitative data were based on 30 samples per group with three biological duplications. Data are represented as mean ± SDs. *P < 0.05 as compared to the respective vehicle group. (Dunnett’s post hoc test following one-way analysis of variance (ANOVA)). |
BA Ameliorates Mineralization of DEX-Induced Osteoporosis Zebrafish Larvae
Bone tissue mineralization plays an essential role in improving strength of the skeleton.36 The content of calcium and phosphorus were assayed to identify the effect of BA on mineral level in DEX-induced osteoporosis. In Figure 2A and B, significant reductions of both mineralized matrix (calcium and phosphorus) were observed in DEX-induced osteoporosis zebrafish larvae in contrast with normal fish (DEX-) (P<0.05). The remarked elevation of calcium and phosphorus contents was detected after treatments of BA and positive control ED.
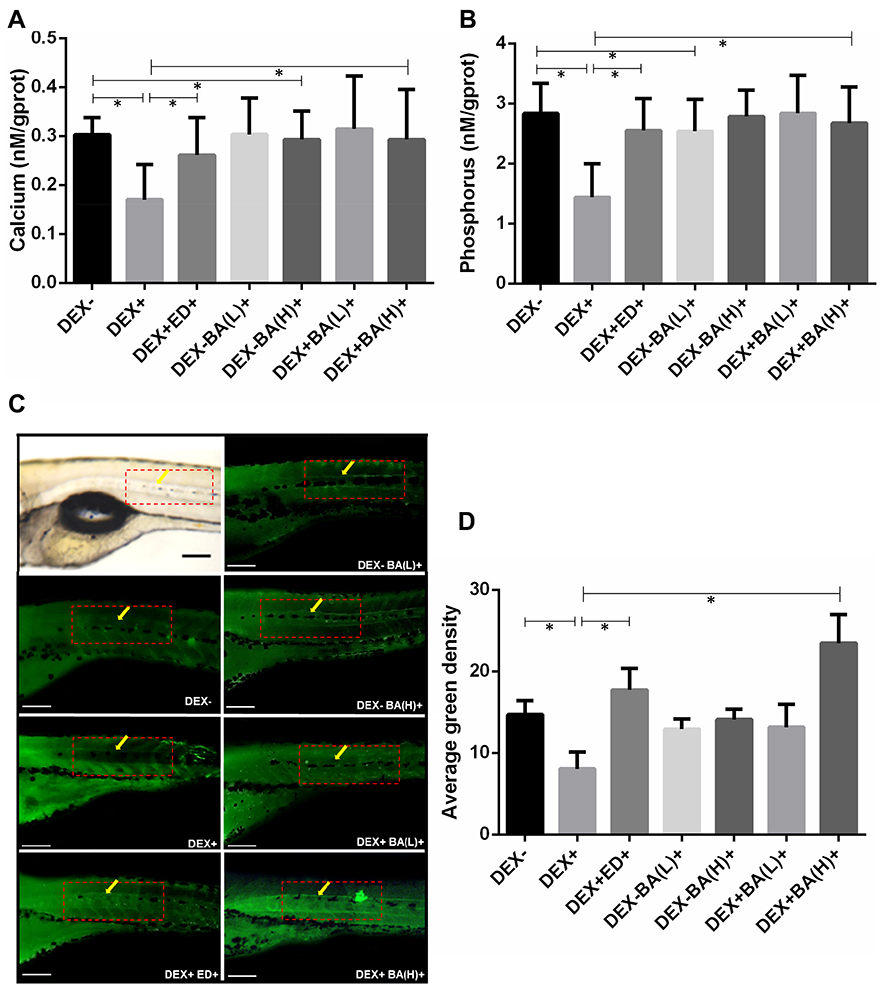 |
Figure 2 Effect of BA on bone tissue mineralization of dexamethasone (DEX)-induced osteoporosis zebrafish larvae. (A) Effect of BA on calcium content (n = 10); (B) Effect of BA on phosphorus content (n = 10); (C) Zebrafish larvae were collected stained with 1% calcein. The fluorescent signal of the calcein-stained vertebrate column was observed under fluorescence microscopy (n=10; magnification 10×, excitation 480 ± 40 nm; emission 510 nm). (D) Quantifications of average green density are defined by the ratio of bone green intensity (green) over the total boxed area (from the 21 to 26 notochords counted from the tail, as demarcated by red dotted box) and measured by online ImageJ software. All values are expressed as mean ± SDs. *P < 0.05 as compared to respective vehicle group. (Dunnett’s post hoc test following one-way analysis of variance (ANOVA)). |
To investigate the direct effects of BA on bone formation in DEX-induced osteoporosis, the bone mineralization was also evaluated by calcein staining using 9 dpf embryos. The stained average green fluorescence density was quantitated by microscopic inspection and digital imaging methods to evaluate the impact of treated compounds. Figure 3C and D show that comparing with untreated fish, DEX distinctly induced a decrease of bone mineral density in the larval zebrafish. On the contrary, we observed that bone mineralization was strikingly enhanced in BA-treated osteoporosis zebrafish. The effect of BA(H)- and positive ED-treated groups were significant (P<0.05). These results combined confirmed that BA might play a positive role in bone mineralization in DEX-induced osteoporosis in zebrafish larvae.
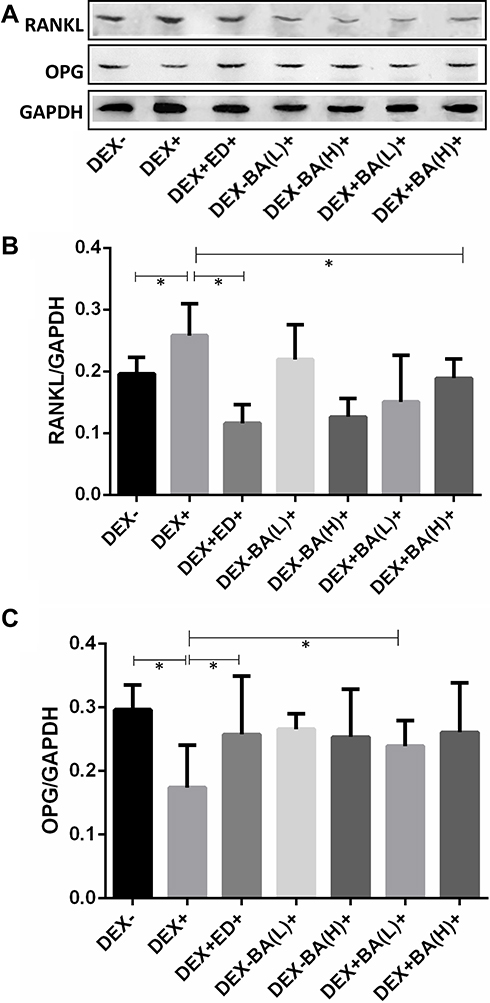 |
Figure 3 Western blotting analysis of the RANKL and OPG expression from dexamethasone (DEX)-induced osteoporosis zebrafish larvae. (A) RANKL and OPG protein level detected via Western blotting. (B) RANKL expression analysis result. (C) OPG protein expression analysis result. Data are shown as the mean ± SD, n = 10, *P < 0.05. |
BA Regulates the Expression of RANKL and OPG in Response to DEX Induction
To further investigate the regulation effect of BA on RANK/RANKL/OPG signaling pathway, the expression levels of RANKL and OPG were examined by Western blotting. Results in Figure 3A and B show that RANKL was significantly increased in DEX-treated osteoporosis zebrafish model group comparing with the untreated group (P < 0.05). However, BA-treated DEX-treated osteoporosis model group with high concentration significantly decreased the RANKL expression with contrast to the DEX-treated osteoporosis group (P < 0.05). The effect is identical with the positive ED-treated group (P < 0.05). Simultaneously, the expression of OPG was inhibited by DEX-induction, but this effect was ameliorated by BA treatment (Figure 3C). Together, these results suggest that BA in low concentration plays an effective role in regulation of the expression of OPG and the RANKL protein.
BA Affects the Expression Levels of Transcription Factors Involving in Bone Remodeling
RANKL-induced osteoclast genesis is associated with changes in expression of osteoclast-specific genes. To verify the role of BA in bone remodeling, several osteoclast-specific transcription factors involving in this regulation process were examined by qRT-PCR. As shown in Figure 4, it was found that DEX dramatically enhanced the expression of nuclear factor κB2 (nfκb2), TNF receptor-associated factor 6 (traf6), forkhead box O1 a2 (foxo1a2) and mitogen-activated protein kinase 8b (mapk8b), whereas it significantly decreased the expression of TNF superfamily member 13b (tnfsl3b) (Figure 4A–E). In addition, BA with both low and high concentrations restored the DEX-induced upregulation of nf-κb2 (Figure 4A), traf6 (Figure 4B), mapk8b (Figure 4C), foxo1a2 (Figure 4D), and downregulation of tnfsl3b (Figure 4E). The results were verified by PCR (Figure 4F). As genes detected are downstream transcriptional factors of RANKL, these data further demonstrate that BA ameliorates DEX-induced osteoporosis by regulation of the RANK/RANKL/OPG signaling pathway.
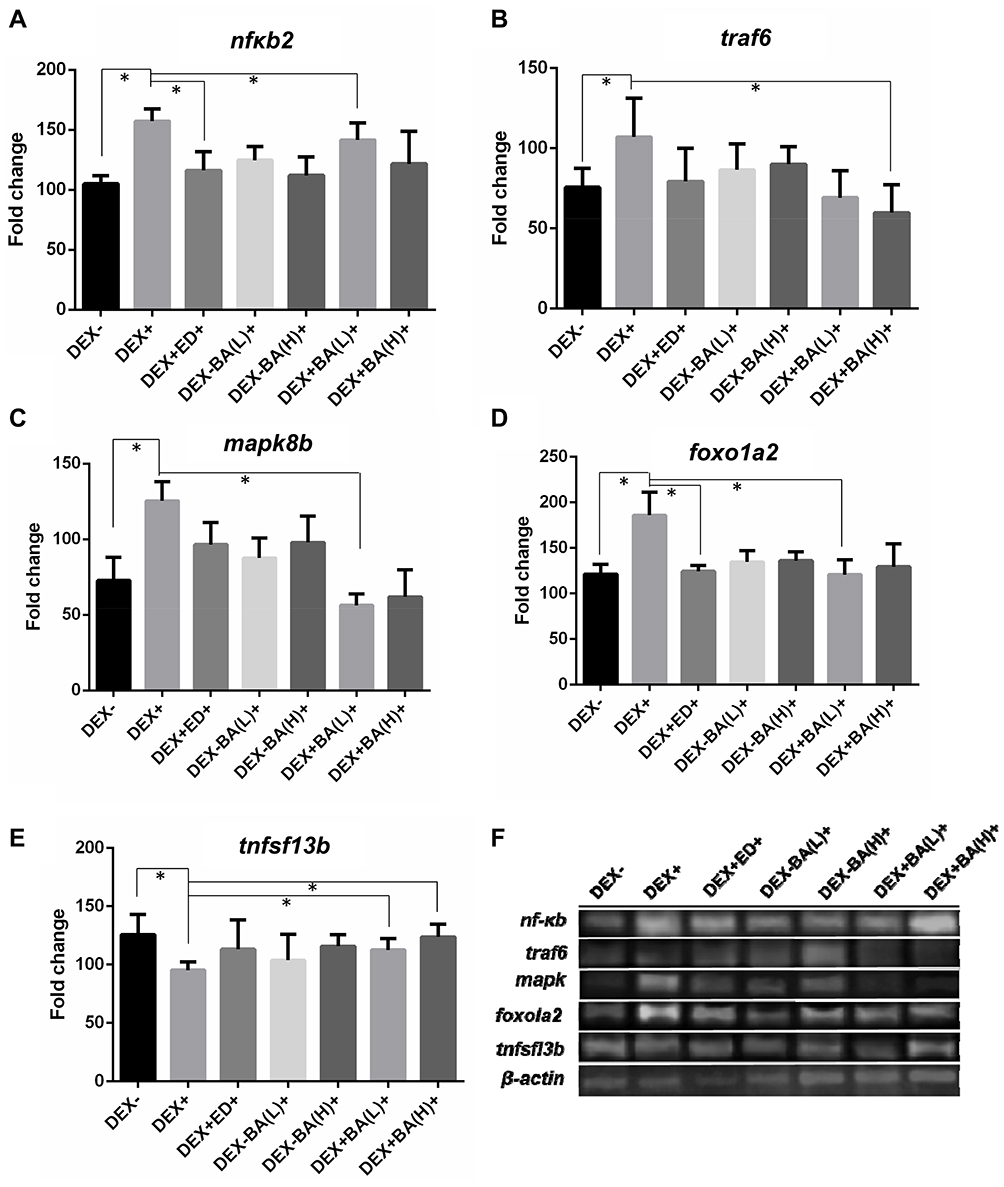 |
Figure 4 Assessing the effect of BA on the expression levels of transcription factors involving in bone remodeling. Zebrafish larvae samples of 9 dpf were collected for RT-qPCR detection (n=10). (A–E) Fold changes shown are the ratio of the values from the treatment of DEX, BA or ED over untreated wild-type fish after calibration with β-actin mRNA as an internal control. Asterisks indicate significant difference with *P < 0.05 by one-way analysis of variance (ANOVA) among the three biological replicates; (F) PCR verification test. |
The Specific Interactions Between BA and RANKL
To explore the detail interactions between BA and RANKL, the Flexible Docking module of DS 3.5 was applied to simulate the binding situation. It was analyzed that the docking interaction energy of BA and RANKL was 45.11kcal/mol. Figure 5A shows the simulation of three-dimensional (3D) structure of BA. Figure 5B exhibits the hydrogen bonding interactions between BA and RANKL in 3D. It was found that after the peptide automatic docked at the RANKL-active site, BA was wrapped by the inner central cavity of the RANKL composite. The detail interactions of BA and amino acid residues of RANKL in 3D schematic diagram (Fig. 5C) and 2D diagram of interactions (Figure 5D) show that there were five hydrogen bonds in BA-RANKL interaction, which indicates that BA and human source RANKL complex is comparatively stable. The molecular simulation results provide a theoretical basis for the interaction of BA and RANKL.
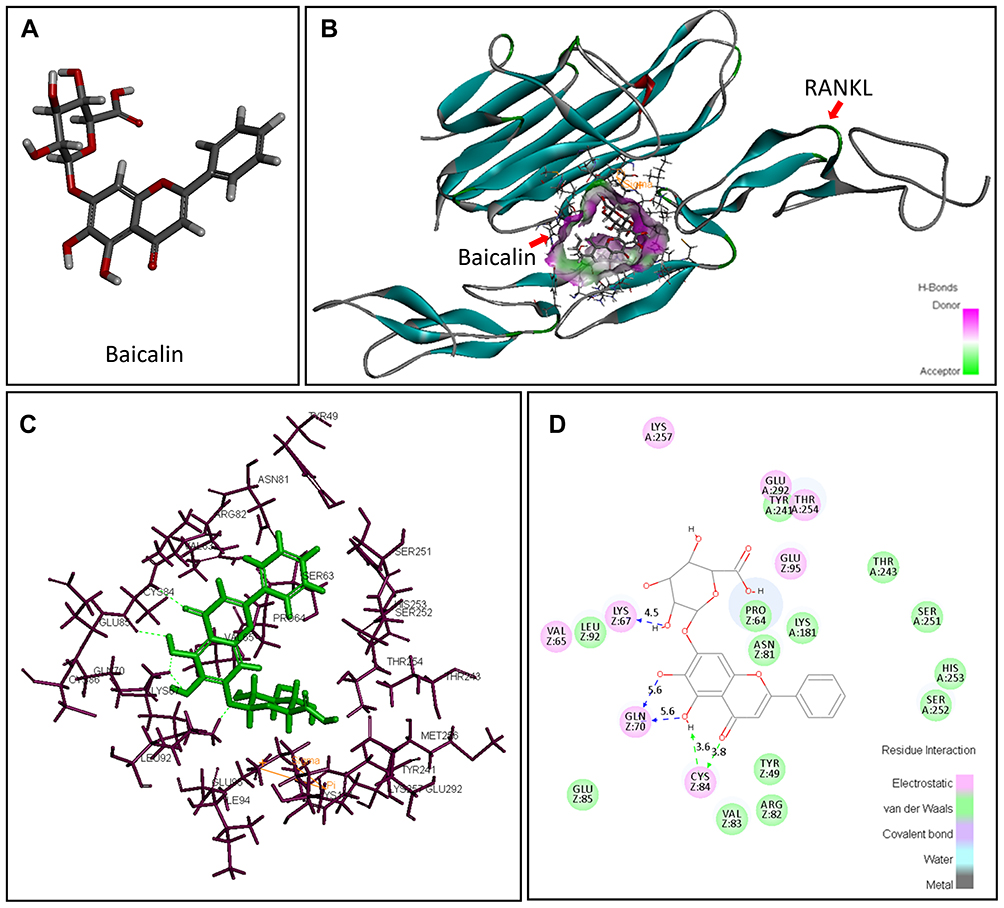 |
Figure 5 The specific interactions between RANKL and BA after the peptide automatic docked at the RANKL-active site. (A) Three-dimensional (3D) structure of baicalin; (B) Details hydrogen bonding interactions of RANKL-BA in the 3D schematic diagram; (C) Interactions of BA and amino acid residues of RANKL in 3D schematic diagram; (D) The 2D diagram of interactions between RANKL and BA. H-bonds with main chains of amino acids are represented by a green dashed arrow directed towards the electron donor. H-bonds with side-chains of amino acids are represented by a blue dashed arrow directed towards the electron donor (image obtained with Accelrys DS Visualizer software). |
Discussion
With the growth of the elderly population, osteoporosis has become a major public health problem worldwide.37 Since osteoporosis is a chronic disease, natural flavonoids have certain advantages because of their fewer side effects and adaptation for suitability for long-term use.38 Among natural flavonoids, BA has been used in herbal formulas in traditional Chinese medicine to treat inflammatory diseases for thousands of years.24 Extensive studies have shown that BA has an effect on scavenging oxygen free radicals,39 alleviating ischemic reperfusion40 of the organizations, regulation of the immune,24 suppressing autophagy,41 accelerating the apoptosis,42 etc. Recently, studies point out that the anti-inflammatory properties of baicalin may result from the inhibition of signaling NF-κB pathway.27,28,30 Previous evidence has demonstrated that BA showed desirable effects against periodontitis and alveolar bone resorption and it exerts these effects by inhibition of the RANKL mRNA expression and upregulation of the NF-κB activation.5,29,30,43 Other than, BA was proved to increase the osteoblastic mineralization and promote osteoblastic differentiation via Wnt/β-catenin signaling.31 However, the effects of BA on osteoporosis and the mechanisms by which it exerts these effects remain unclear.
Zebrafish is an ideal animal model in vivo for studying bone deformations for the high similarity of the structure and genetics of bone with human beings.34,44 The bone of zebrafish larval contains the sufficient and necessary cells for both bone formation and resorption activity.33 In the present study, we examined the bone mineralization by calcein staining, which is a vital dye staining and the biological mineralization process of zebrafish at phenotypic can be observed directly. Based on the lines of advantages above, we established the DEX-induced zebrafish osteoporosis model to investigate the effects of BA on the dysfunction of bone formation.
In this work, our evidence demonstrates that BA exerts positive effects on DEX-induced growth retardation and mortality of zebrafish larvae. As known, bone mineral density is an important marker for bone strength, serving as a basis to predict the risk of fracture. Our results show that the BA treatment significantly improved the contents of mineralized matrix (calcium and phosphorus), as well as the mineralization detected by calcein staining, suggesting that BA prevents the DEX-induced osteoporosis changes. Other than this, BA rescues impaired osteogenesis elicited by DEX, involved in enhancing the bone mineralization and hampering the gene expression changes related to bone formation and resorption under the conditions of DEX in zebrafish. It can be inferred by molecular docking that BA may interact directly with the extracellular domain of RANKL. The findings reveal that BA ameliorates DEX-induced osteoporosis by regulation of the RANK/RANKL/OPG signaling pathway.
It has been well recognized that the proposed theory of the RANKL/RANK/OPG pathway opened up a new therapeutic avenue of osteoporosis.9 In our study, the expression of RANKL and OPG was detected by Western blotting. From the results, it was concluded that through BA administration, the expression of RANKL is inhibited while OPG is upregulated. Alternatively, the ratio of OPG/RANKL is decreased in the DEX-treated group; however, the reduction is turned over by BA. This conclusion is consistent with reported.45 During the process of osteoporosis, RANKL recognizes and binds to its receptor RANK on the surface of osteoclast precursors, and then recruits the adapter protein TRAF6, thereby leading to sequential activation of NF-κB, MAPK, c-Fos and nuclear factor of activated T cells c1 (NFATc1), which mediate the completion of the osteoclast differentiation process.46,47 TRAF6 triggers the self-amplification and gene transcription of NFATc1 and induces the differentiation process.47 Mature osteoclast can produce tumor necrosis factor superfamily 13b (TNFSF13b), which acts on myeloma cells to produce DNA polymerase γ (MIP-1) and Interleukin (IL)-3, thereby stimulating the osteoclast precursor cells to form a circulation (Figure 6).48 In this study, the results of gene expression detection verified the signaling pathway in DEX-induced zebrafish. Moreover, BA treatments rescue the DEX-induced gene expression changes to a normal level, which suggests a possible mechanism for BA action.
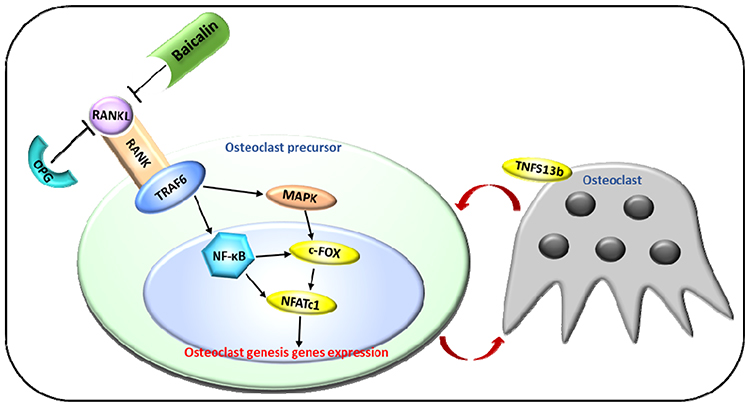 |
Figure 6 Regulation of the RANK/RANKL/OPG Signaling Pathway in osteoporosis process. RANKL recognizes and binds to its receptor RANK on the surface of osteoclast precursors, and then recruits the adapter protein TRAF6, thereby leading to sequential activation of NF-κB, MAPK, c-Fos and NFATc1, which mediate the completion of the osteoclast differentiation process.46,47 Mature osteoclast produces TNFSF13b, which stimulates the osteoclast precursor cells to form a circulation.48 |
To further explore the interactions of BA and RANKL, molecular simulation was performed. Results show that there was a strong interaction between BA and human RANKL/OPG-CRD complex, and the interaction locates in the inner cavity of the complex center, which suggests the close combination of RANKL and BA. The potential directly binding reveals that BA might be beneficial in preventing the formation of the RANKL-RANK complexes; however, further experimental evidence is still needed. In all, we propose that baicalin ameliorates dexamethasone-induced osteoporosis by regulation of the RANK/RANKL/OPG signaling pathway. This conclusion provides a novel candidate drug for osteoporosis therapy with few adverse side effects such as gastrointestinal complaints comparing with ED49 and proposes a new perspective idea for BA anti-osteoporosis mechanism study.
Acknowledgements
This work was supported by grants from National Natural Science Foundation of China (No. 31701279) and Jiangsu Synergetic Innovation Center for Advanced Bio-Manufacture (No. XTE1845).
Disclosure
The authors declare that they have no conflict of interest.
References
1. Cornelius C, Koverech G, Crupi R, et al. Osteoporosis and alzheimer pathology: role of cellular stress response and hormetic redox signaling in aging and bone remodeling. Front Pharmacol. 2014;5:120. doi:10.3389/fphar.2014.00120
2. Genant HK, Cooper C, Poor G, et al. Interim report and recommendations of the World Health Organization task-force for osteoporosis. Osteoporos Int. 1999;10:259–264. doi:10.1007/s001980050224
3. Sato AY, Cregor M, Delgado-Calle J, et al. Protection from glucocorticoid-induced osteoporosis by anti-catabolic signaling in the absence of sost/sclerostin. J Bone Miner Res. 2016;31:1791–1802. doi:10.1002/jbmr.2869
4. Theoleyre S, Wittrant Y, Tat SK, et al. The molecular triad OPG/RANK/RANKL: involvement in the orchestration of pathophysiological bone remodeling. Cytokine Growth Factor Rev. 2004;15:457–475. doi:10.1016/j.cytogfr.2004.06.004
5. Jiang L, Song J, Hu X, et al. The proteasome inhibitor bortezomib inhibits inflammatory response of periodontal ligament cells and ameliorates experimental periodontitis in rats. J Periodontol. 2017;88:473–483. doi:10.1902/jop.2016.160396
6. Kurinami H, Shimamura M, Nakagami H, et al. A novel therapeutic peptide as a partial agonist of RANKL in ischemic stroke. Sci Rep. 2016;6:38062. doi:10.1038/srep38062
7. Maria S, Samsonraj RM, Munmun F, et al. Biological effects of melatonin on osteoblast/osteoclast cocultures, bone, and quality of life: implications of a role for MT2 melatonin receptors, MEK1/2, and MEK5 in melatonin-mediated osteoblastogenesis. J Pineal Res. 2018;64:e12465. doi:10.1111/jpi.12465
8. Holloway WR, Collier FM, Aitken CJ, et al. Leptin inhibits osteoclast generation. J Bone Miner Res. 2002;17:200–209. doi:10.1359/jbmr.2002.17.2.200
9. Yuan Y, Zhang L, Tong X, et al. Mechanical stress regulates bone metabolism through MicroRNAs. J Cell Physiol. 2017;232:1239–1245. doi:10.1002/jcp.25688
10. Montalcini T, Romeo S, Ferro Y, et al. Osteoporosis in chronic inflammatory disease: the role of malnutrition. Endocrine. 2013;43:59–64. doi:10.1007/s12020-012-9813-x
11. Lewiecki EM. Safety and tolerability of denosumab for the treatment of postmenopausal osteoporosis. Drug Healthc Patient Saf. 2011;3:79–91. doi:10.2147/dhps.s7727
12. Lian C, Wu Z, Gao B, et al. Melatonin reversed tumor necrosis factor-alpha-inhibited osteogenesis of human mesenchymal stem cells by stabilizing SMAD1 protein. J Pineal Res. 2016;61:317–327. doi:10.1111/jpi.12349
13. Sharan K, Lewis K, Furukawa T, Yadav VK. Regulation of bone mass through pineal-derived melatonin-MT2 receptor pathway. J Pineal Res. 2017;63:e12423. doi:10.1111/jpi.12423
14. Li T, Jiang S, Lu C, et al. Melatonin: another avenue for treating osteoporosis? J Pineal Res. 2019;66:e12548. doi:10.1111/jpi.12548
15. Tella SH, Gallagher JC. Prevention and treatment of postmenopausal osteoporosis. J Steroid Biochem Mol Biol. 2014;142:155–170. doi:10.1016/j.jsbmb.2013.09.008
16. Mitani M, Miura Y, Saura R, et al. Estrogen specifically stimulates expression and production of osteoprotegerin from rheumatoid synovial fibroblasts. Int J Mol Med. 2005;15:827–832.
17. Yeon JT, Kim K-J, Choi S-W, et al. Anti-osteoclastogenic activity of praeruptorin A via inhibition of p38/Akt-c-Fos-NFATc1 signaling and PLCgamma-independent Ca2+ oscillation. PLoS One. 2014;9:e88974. doi:10.1371/journal.pone.0088974
18. He L, Lee J, Jang JH, et al. Ginsenoside Rh2 inhibits osteoclastogenesis through down-regulation of NF-kappaB, NFATc1 and c-Fos. Bone. 2012;50:1207–1213. doi:10.1016/j.bone.2012.03.022
19. Park SH, Kim J-Y, Cheon Y-H, et al. Protocatechuic acid attenuates osteoclastogenesis by downregulating JNK/c-Fos/NFATc1 signaling and prevents inflammatory bone loss in mice. Phytother Res. 2016;30:604–612. doi:10.1002/ptr.5565
20. Qu B, Xia X, Yan M, et al. miR-218 is involved in the negative regulation of osteoclastogenesis and bone resorption by partial suppression of p38MAPK-c-Fos-NFATc1 signaling: potential role for osteopenic diseases. Exp Cell Res. 2015;338:89–96. doi:10.1016/j.yexcr.2015.07.023
21. Li H, Wang M, Liang Q, et al. Urinary metabolomics revealed arsenic exposure related to metabolic alterations in general Chinese pregnant women. J Chromatogr A. 2017;1479:145–152. doi:10.1016/j.chroma.2016.12.007
22. Ming J, Zhuoneng L, Guangxun Z. Protective role of flavonoid baicalin from scutellaria baicalensis in periodontal disease pathogenesis: a literature review. Complement Ther Med. 2018;38:11–18. doi:10.1016/j.ctim.2018.03.010
23. Cai LL, Liu GY, Tzeng CM. Genome-wide DNA methylation profiling and its involved molecular pathways from one individual with thyroid malignant/benign tumor and hyperplasia: a case report. Medicine. 2016;95:e4695. doi:10.1097/md.0000000000004695
24. Luan Y, Chao S, Ju Z-Y, et al. Therapeutic effects of baicalin on monocrotaline-induced pulmonary arterial hypertension by inhibiting inflammatory response. Int Immunopharmacol. 2015;26:188–193. doi:10.1016/j.intimp.2015.01.009
25. Jin J, Chen Y, Wang D, et al. The inhibitory effect of sodium baicalin on oseltamivir-resistant influenza A virus via reduction of neuraminidase activity. Arch Pharm Res. 2018;41:664–676. doi:10.1007/s12272-018-1022-6
26. Wang Q, Xu H, Zhao X. Baicalin inhibits human cervical cancer cells by suppressing protein kinase C/Signal Transducer and Activator of Transcription (PKC/STAT3) signaling pathway. Med Sci Monit. 2018;24:1955–1961. doi:10.12659/MSM.909640
27. Dong SJ, Zhong Y-Q, Lu W-T, et al. Baicalin inhibits lipopolysaccharide-induced inflammation through signaling NF-kappaB pathway in HBE16 airway epithelial cells. Inflammation. 2015;38:1493–1501. doi:10.1007/s10753-015-0124-2
28. Liu J, Wei Y, Luo Q, et al. Baicalin attenuates inflammation in mice with OVA-induced asthma by inhibiting NF-kappaB and suppressing CCR7/CCL19/CCL21. Int J Mol Med. 2016;38:1541–1548. doi:10.3892/ijmm.2016.2743
29. Pei Z, Wang B, Zhang F, et al. Response of human periodontal ligament cells to baicalin. J Periodontol. 2014;85:1283–1290. doi:10.1902/jop.2014.130635
30. Wang GF, Wu ZF, Wan L, Wang QT, Chen FM. Influence of baicalin on the expression of receptor activator of nuclear factor-kappaB ligand in cultured human periodontal ligament cells. Pharmacology. 2006;77:71–77. doi:10.1159/000092853
31. Guo AJ, Choi RCY, Cheung AWH, et al. Baicalin, a flavone, induces the differentiation of cultured osteoblasts: an action via the Wnt/beta-catenin signaling pathway. J Biol Chem. 2011;286:27882–27893. doi:10.1074/jbc.M111.236281
32. Sprague J, Bayraktaroglu L, Bradford Y, et al. The zebrafish information network: the zebrafish model organism database provides expanded support for genotypes and phenotypes. Nucleic Acids Res. 2007;36:D768–D772. doi:10.1093/nar/gkm956
33. Chen JR, Lai YH, Tsai JJ, Hsiao CD. Live fluorescent staining platform for drug-screening and mechanism-analysis in zebrafish for bone mineralization. Molecules. 2017;22. doi:10.3390/molecules22122068
34. Luo S, Yang Y, Chen J, et al. Tanshinol stimulates bone formation and attenuates dexamethasone-induced inhibition of osteogenesis in larval zebrafish. J Orthop Translat. 2016;4:35–45. doi:10.1016/j.jot.2015.07.002
35. Hanley DA, Ioannidis G, Adachi JD. Etridronate therapy in the treatment and prevention of osteoporosis. J Clin Densitom. 2000;3:79–95. doi:10.1385/JCD:3:1:079
36. Martin RB, Ishida J. The relative effects of collagen fiber orientation, porosity, density, and mineralization on bone strength. J Biomech. 1989;22:419–426. doi:10.1016/0021-9290(89)90202-9
37. Anbinder AL, Moraes RM, Lima GMG, et al. Periodontal disease exacerbates systemic ovariectomy-induced bone loss in mice. Bone. 2016;83:241–247. doi:10.1016/j.bone.2015.11.014
38. Feize W, Meng L, Yanni L, et al. A randomized controlled study to observe the efficacy of external treatment with a traditional chinese medicine herbal ointment on malignant plural effusion: outcome report and design review. Integr Cancer Ther. 2017;16:473–478. doi:10.1177/1534735416660193
39. Wang S, Zang W, Yang Y, et al. Tanshinone IIA and baicalin inhibiting the formation of benzo[a]pyrene and benzo[a]pyrene induced cytotoxicity: correlation with scavenging free radical. Environ Toxicol Pharmacol. 2013;36:403–410. doi:10.1016/j.etap.2013.05.003
40. Dinda B, Dinda S, DasSharma S, et al. Therapeutic potentials of baicalin and its aglycone, baicalein against inflammatory disorders. Eur J Med Chem. 2017;131:68–80. doi:10.1016/j.ejmech.2017.03.004
41. Wang G, Liang J, Gao L-R, et al. Baicalin administration attenuates hyperglycemia-induced malformation of cardiovascular system. Cell Death Dis. 2018;9:234. doi:10.1038/s41419-018-0318-2
42. Shu YJ, Bao R-F, Wu X-S, et al. Baicalin induces apoptosis of gallbladder carcinoma cells in vitro via a mitochondrial-mediated pathway and suppresses tumor growth in vivo. Anticancer Agents Med Chem. 2014;14:1136–1145. doi:10.2174/1871520614666140223191626
43. Sun JY, Li D-L, Dong Y, et al. Baicalin inhibits toll-like receptor 2/4 expression and downstream signaling in rat experimental periodontitis. Int Immunopharmacol. 2016;36:86–93. doi:10.1016/j.intimp.2016.04.012
44. Li N, Felber K, Elks P, Croucher P, Roehl HH. Tracking gene expression during zebrafish osteoblast differentiation. Dev Dyn. 2009;238:459–466. doi:10.1002/dvdy.21838
45. Wu H, Zhong Q, Wang J, et al. Beneficial effects and toxicity studies of xian-ling-gu-bao on bone metabolism in ovariectomized rats. Front Pharmacol. 2017;8:273. doi:10.3389/fphar.2017.00273
46. Aoyama E, Kubota S, Khattab HM, Nishida T, Takigawa M. CCN2 enhances RANKL-induced osteoclast differentiation via direct binding to RANK and OPG. Bone. 2015;73:242–248. doi:10.1016/j.bone.2014.12.058
47. Boyce BF, Xing L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch Biochem Biophys. 2008;473:139–146. doi:10.1016/j.abb.2008.03.018
48. Breitkreutz I, Raab MS, Vallet S, et al. Lenalidomide inhibits osteoclastogenesis, survival factors and bone-remodeling markers in multiple myeloma. Leukemia. 2008;22:1925–1932. doi:10.1038/leu.2008.174
49. KRANE SM. Drugs five years later: etidronate disodium in the treatment of Paget’s disease of bone. Ann Intern Med. 1982;96:619–625. doi:10.7326/0003-4819-96-5-619
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.