Back to Journals » Drug Design, Development and Therapy » Volume 19
Astaxanthin Promotes Spermatogenesis in Mice by Reducing Mitochondrial Oxidative Stress Damage and Regulating Fatty Acid Metabolism and Ferroptosis
Authors Liu H ![]() , Chen X, Feng X, Zhu Z, Liao Z, Zhu S, Pang T, Ren X, Yang R
, Chen X, Feng X, Zhu Z, Liao Z, Zhu S, Pang T, Ren X, Yang R
Received 11 April 2025
Accepted for publication 23 August 2025
Published 4 September 2025 Volume 2025:19 Pages 7777—7794
DOI https://doi.org/10.2147/DDDT.S522738
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Yan Zhu
Huang Liu,1,* Xuren Chen,2,* Xin Feng,3,* Zhiyong Zhu,1 Zhiwei Liao,4 Shenghui Zhu,1 Tao Pang,1 Xuejun Ren,5 Ruilin Yang6
1Department of Andrology, National Health Commission (NHC) Key Laboratory of Male Reproduction and Genetics, Guangdong Provincial Reproductive Science Institute (Guangdong Provincial Fertility Hospital), Human Sperm Bank of Guangdong Province, Guangzhou, Guangdong, 510600, People’s Republic of China; 2Department of Andrology, the First Affiliated Hospital of Sun Yat-Sen University, Guangzhou, Guangdong, 510080, People’s Republic of China; 3Department of Urology and Andrology, the First Affiliated Hospital of Sun Yat-sen University., Guangzhou, Guangdong, 510080, People’s Republic of China; 4Radiotherapy Department Area 4, Affiliated Cancer Hospital and Institute of Guangzhou Medical University, Guangzhou, Guangdong, 510095, People’s Republic of China; 5Guangdong Xingding Health Management Co., Ltd., Zhongshan, Guangdong, 528400, People’s Republic of China; 6Department of Andrology, the Affiliated Panyu Central Hospital of Guangzhou Medical University, Guangzhou, 511400, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Huang Liu, Email [email protected] Xuejun Ren, Email [email protected]
Background: Spermatogenesis is a complex process that affects the outcome of fertility. Different types of cellular metabolic processes have both positive and negative effects on sperm production. Exploring new methods to promote spermatogenesis is the best way to improve fertility. This study confirmed the effect of astaxanthin on promoting spermatogenesis through various experiments.
Objective: To explore new activities of astaxanthin and develop the new methods to promote spermatogenesis.
Methods: Network pharmacology, in vitro cell culture and in vivo experiments were used in this research. The targets and potential signaling pathways of astaxanthin in the treatment of spermatogenesis, the effects on the proliferation and apoptosis of spermatogonial stem cells, and the therapeutic effect on oligoasthenozoospermia in mice induced with cyclophosphamide of astaxanthin were all observed. The ACSL3, VDAC, GPX4, FADS2, GLS2, Steap3, MDA, GSH-Px, and iron ions were detected and analyzed to reveal the potential regulatory mechanisms.
Results: Astaxanthin has 52 key targets for treating spermatogenesis, among which the oxidative stress metabolic pathway is one of the most important factors. The sperm concentration and forward motility of the oligoasthenozoospermia model mice fed with astaxanthin were significantly greater than those of the control group. The proliferation rate of spermatogonial stem cells cultured with astaxanthin was also significantly greater than that of quercetin group and the proportion of apoptotic cells was significantly lower. Astaxanthin can reduce ACSL3, MDA, and iron ions in spermatogonial stem cells; increase the expression of Steap3, VDAC, GPX4, GLS2, GSH-Px, and FADS2; and improve ester metabolism to promote spermatogenesis in oligoasthenozoospermia model mice.
Conclusion: Astaxanthin can regulate the metabolism of fatty acid through the ferroptosis pathway, and reduce the mitochondrial oxidative stress damage, and further regulate FADS2, Steap3, and other factors to promote spermatogenesis.
Keywords: astaxanthin, fatty acid metabolism, spermatogenesis, ferroptosis, oxidative stress damage
Introduction
The metabolism of cells affects the composition and function of local microenvironments. The local metabolic dysfunction of Sertoli cells, Leydig cells, and spermatogenic cells in the seminiferous tubules inevitably leads to an imbalance in the spermatogenic microenvironment, and the excessive aggregation or insufficiency of certain metabolites inevitably interferes with the process of spermatogenesis. Excessive accumulation of trace elements such as lead, fluorine, selenium, and cadmium can lead to a decrease in the antioxidant capacity of mouse, leading to a decrease in the vitality and quantity of sperm.1,2 Alcohol, formaldehyde, and other substances can also damage the metabolic function of testicular tissue, causing an imbalance in the internal environment and affecting sperm development.3,4 Therefore, improving the metabolic capacity and the internal environment of the testes, and promoting sperm production are effective measures to enhance fertility.
Lipids are among the three important elements involved in the testicular synthesis of androgens, interference with cell function, and regulation of spermatogenesis. They are involved in spermatogenesis and maturation, characteristic enrichment and differentiation, and the expression of various functions. Studies have shown that sperm membranes are composed of complex lipid mixtures, such as phospholipids, glycolipids, and sterols. These lipid substances have good fluidity, and the metabolism of these unsaturated fatty acids plays an important role in spermatogenesis as sperm fertilization ability changes.5–7 Some studies have also shown that sulfogalactosylglycerides (SGGs) are selectively synthesized in large quantities in mammalian testicular germ cells, and they directly participate in cell adhesion and are degraded into lipid skeletons by lysosomal enzymes in testicular Sertoli cells. When this balance is disrupted, Sertoli cells are prone to lysosomal storage and subdysfunction, which in turn affects their support for germ cell development and leads to a decrease in fertility.8,9
Minguez Alarc ó n et al10 reported that the intake of monounsaturated fatty acids was negatively correlated with the serum levels of free testosterone, total testosterone, and inhibin b. Jarvis et al11 reported that a high-fat diet can affect the expression of 102 important blood‒testis barrier structural proteins, leading to spermatogenesis disorders. Setayesh et al12 reported that excessive fat can lead to DNA damage in multiple organs, including the brain, liver, colon, and testes. The molecular mechanisms involved may include ROS formation, lipid peroxidation, indirect inflammation, and the formation of glycation end products during DNA repair. However, some scholars question this viewpoint. Currently, the results of 39 studies on humans are contradictory, and the significance of the impact of weight gain on genomic instability is unclear. Although abnormal lipid metabolism can lead to a decrease in telomere length, hormonal imbalance, and dysregulation of drug-metabolizing enzyme activity in genotoxic carcinogens, the evidence from these studies is not sufficient, and more data are still needed to draw accurate conclusions.
Astaxanthin and quercetin are two common antioxidants in nature, but their sources, antioxidant effects, and therapeutic effects on diseases differ. Astaxanthin is a ketone carotenoid that has the ability to scavenge free radicals and enhance immunity.13 Quercetin can also improve blood circulation and promote growth. Currently, many studies suggest that quercetin can improve testicular function in animals and promote spermatogenesis.14,15 However, there is relatively little research on astaxanthin. Most studies have shown that astaxanthin has a positive effect on sperm cryopreservation,16,17 but there is not much research on whether taking astaxanthin can promote spermatogenesis.
To further explore the new method to promote spermatogenesis, we conducted experimental research using astaxanthin and found that it is a potential new option with the value of promoting spermatogenesis, it is also the new activity of astaxanthin. We conducted network pharmacology analysis, animal experiments and in vitro cell experiments, by using qtPCR, WB, immunofluorescence, chemiluminescence, and other methods to detect the the new mechanism of astaxanthin regulating spermatogenesis in mice. We discovered the new relationship between lipid metabolism and spermatogenesis, and explored a new reference for astaxanthin to promote spermatogenesis.
Materials and Reagents
Materials
(1) Four-week-old C57BL/6 male mice (SCXK2020 0051, NO.44822700006188, Baishitong Biotechnology, Zhuhai). (2) Mouse Spermatogonial Stem Cells and culture medium (Procell Life Science & Tech, Wuhan). (3) 1 mL and 2 mL syringes (K20210422, Kangdeli, Shanghai). (3) Anatomic forceps, Surgical knife, Anatomic scissors, and Anatomy table (WA3070, J0B020, WA1050, Medical Instruments Group, Shanghai). (4) Gastric gavage tube (TFEP-001, Zhongke Life, Guangdong).
Medications
(1) Quercetin (Shengjing capsule, Z20027672, Liaoyuan Hetang Pharmaceutical, Guizhou). (2) Astaxanthin (Monakang, Q/SSH0226, Xingding Health Management, Guangdong). (3) Physiological saline (A22052402250, Kelun, Sichuan). (4) Cyclophosphamide (H2160467, Bayer, Germany).
Reagents
Please refer to Supplementary Table 1 for details.
Instruments
Please refer to Supplementary Table 2 for details.
Antibodies
Please refer to Supplementary Table 3 for details.
Method
Network Pharmacology Analysis
Materials
(1) Commonly used gene public databases include gene databases and string databases. (2) Network display and analysis software: Cytoscape 3.10.1 software (California, USA).
Target Screening for Spermatogenesis
The keywords “sperm” and “astaxanthin” were used to search for targets related to spermatogenesis in the GeneCards database, and the obtained targets were uploaded to the UniProt database to obtain the standard names of the targets, with the protein species set to “Homo sapiens (Human)”.
Obtaining Key Targets and Building a PPI Network
The genes associated with both astaxanthin and spermatogenesis were uploaded to the STRING database, a drug-targeted PPI network was constructed, and the PPI network was uploaded to Cytoscape 3.10.1 software. The network analyzer function was used for topology analysis, the genes were sorted according to the edge count value, and the key targets of drug action were obtained.
GO Function and KEGG Pathway Enrichment Analysis
The genes associated with both astaxanthin and spermatogenesis were uploaded to the STRING database for GO functional enrichment analysis to obtain the molecular function (MF), cellular component (CC), and biological process (BP) pathways. Cytoscape 3.10.1 software was used to perform KEGG pathway enrichment analysis on significant signaling pathways of key target genes, and a network of astaxanthin target KEGG signaling pathways was constructed.
Animal Modeling
In accordance with the relevant literature, an equivalent drug concentration ratio of 1:9.1 was used to calculate the administration concentration.18,19 A mouse model of severe oligoasthenozoospermia was established by intraperitoneal injection of cyclophosphamide at an equivalent drug concentration of 320 mg/kg for 2 weeks.
Animal in Vivo Experiments
A total of 10 6-week-old normal mice and 20 model mice were used for vivo experiments. Normal mice (n=5) and Models mice (n=5) were euthanized before the intervention to obtain samples as controls, while the remaining normal mice (Group D, n=5) and remaining model mice (Group A, B, C) entered the intervention experiment for 4 weeks of feeding. The remaining model mice were divided into three groups with 5 of each, which contained Group A (n=5) fed with Astaxanthin, Group B (n=5) fed with quercetin, and Group C (n=5) fed with physiological saline. Each group of mice was housed in separate cages and maintained continuously for 4 weeks. The drug concentration fed to the mice of Group A, Group B, and Group C was calculated based on the equivalent concentration formula of human and mouse, with the drug concentration of 5mg/mL and a total daily dose of 0.5mL, administered by gavage at once. The mice of Group D were used as blank controls and fed naturally. The daily mental and dietary status of all mice was observed and recorded, and their weight was measured once a week. The sampling methods were as follows: (1) the mice fasted the night before feeding and could not help but drink; (2) the mice were anesthetized via intraperitoneal injection of 2,2,2-tribromoethanol (150 mg/kg); and (3) the anesthetized mouse was fixed in a supine position on the anatomical table, and its limbs were fixed. (4) After using the eyeball extraction method to extract whole blood, the samples were allowed to stand for approximately 30 minutes, centrifuged, and stored at −80°C for later use. (5) The peritoneum along the midline was cut open, the peritoneum was exposed, the bilateral testicular tissue was located and removed, blood stains were removed with physiological saline, and the tissue was fixed at room temperature with 4% paraformaldehyde for later use. (6) The peritoneum was cut along the midline, the peritoneum was exposed, the epididymal tissue was located and removed, blood was washed away with physiological saline, and the samples were soaked in 2 mL of 4°C physiological saline. (7) The epididymis was cut directly, and the supernatant was semen, which was used for subsequent experiments.
Testing Methods
Semen Detection
After the epididymal fluid was collected, computer-aided sperm analysis (CASA) (Spain SCA) was immediately used to detect the sperm concentration and activity.
Tissue Embedding Sectioning
The mouse testicular tissue was paraffin embedded and sliced via the following steps: (1) dehydration, (2) transparency, (3) soaking wax, (4) embedding, and (5) paraffin sectioning.
Tissue HE Staining
The testicular tissue sections of the mice were stained with HE via the following steps: (1) dewaxing, (2) rehydration, (3) core staining, (4) cytoplasmic staining, (5) sealing, and (6) photography.
Western Blot
WB detection was performed on the testicular tissue samples via the following steps: (1) preparation of protein samples, (2) determination of protein concentration, (3) polyacrylamide gel electrophoresis, (4) membrane transfer, (5) blocking, (6) antibody incubation, (7) development, (8) data analysis, and (9) interpretation of the results.
MTT Detection
MTT assay for detecting the proliferation status of spermatogonial stem cells. The cells were divided into three groups for in vitro culture: the astaxanthin group (Group AA, n=5), quercetin group (Group BB, n=5), and normal control group (Group CC, n=5). Both Group AA and Group BB contained three concentrations of 5 nmol/l, 10 nmol/l, and 15 nmol/l. The logarithmic growth stage cells were collected, resuspended in complete culture medium, the concentration was adjusted to 1×105 cells/mL, and 0.2 mL of suspension was added to each well. Each group had 3 wells and was cultured for 24, 48, or 72 hours before 10 μL of MTT solution was added. The cells were cultured in a cell culture chamber for 4 hours, 200 μL of DMSO was added, and the mixture was placed on a shaking table at 26°C and shaken for 10 minutes to fully dissolve the crystals. The OD490 was subsequently measured via an enzyme-linked immunosorbent assay.
Annexin V-FITC/PI Flow Detection
The apoptosis rate of cultured spermatogonial stem cells was detected via the Annexin V-FITC/PI flow detection method via the following steps: logarithmic growth stage cells were collected, counted, resuspended in complete culture medium, the cell concentration was adjusted to 1×105 cells/mL in a 6-well plate, and 2 mL of cell suspension was added to each well. After the cells adhered to the wall, experimental grouping was carried out. Different concentrations of drug culture media were added to each well, and the samples were cultured for 72 hours. Anexin V-FITC/PI dual-staining flow cytometry was used to detect apoptosis, with a PE excitation wavelength of Ex=488 nm and an emission wavelength of Em=578 nm and a PI excitation wavelength of Ex=536 nm and an emission wavelength of Em=617 nm.
Immunofluorescence
The immunofluorescence method is used to detect specific proteins in testicular tissue via the following steps: (1) dewaxing the paraffin section in water, (2) antigen repair, (3) blocking the antigen with serum, (4) addition of the primary antibody, (5) addition of the secondary antibody, (6) staining the cell nucleus with DAPI, (7) quenching spontaneous fluorescence, (8) sealing, (9) image acquisition, and (10) interpretation of the results.
Determination of Iron Ion, MDA, and GSH-Px
The iron ion, MDA, and GSH-Px concentrations were detected via ELISA via the following steps: (1) According to the instructions of the iron ion colorimetric detection kit, MDA test kit, and GSH-Px test box, the testicular tissue sample was accurately weighed, and 0.9% physiological saline was added as homogenization medium at a weight (mg) to volume (µL) ratio of 1:9. Under ice-water bath conditions, the samples were mechanically homogenized with a tissue grinder to prepare a 10% homogenization solution. (2) Then, the homogenate was placed in a centrifuge and centrifuged at 2500–3000 rpm for 10 minutes. The supernatant was collected after centrifugation for measurement.
Statistical Methods
All the data were statistically analyzed via SPSS 19.0 statistical software (IBM, USA), and the measurement data are expressed as the means ± standard deviations (means ± SD). Homogeneity analysis of variance was performed. Kruskal–Wallis test was used for each categorical variable. When the variance was homogeneous, a t test was used for intergroup comparisons, and when the variance was uneven, Mann–Whitney U-test was used for intergroup comparisons. ANOVA analysis is used for comparison between multiple groups. The count data are expressed in the form of n or percentage (%), chi-square test (χ2) tests were applied for intergroup comparisons, and P<0.05 indicated a statistically significant difference.
Ethics
This study was approved by the Animal Ethics Committee of Guangdong Medical Experimental Animal Center (No. C202107-19), and the Ethics Committee of Guangdong Provincial Reproductive Science Institute (Guangdong Provincial Fertility Hospital) [No. [2021](05)]. Animal experiments are strictly conducted in accordance with the Guidelines for Ethical Review of Experimental Animal Welfare issued by the General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China. The use of the databases in this research is exempt from ethical approval under item 1 and 2 of Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects dated February 18, 2023, China.
Results
Results of Network Pharmacology Analysis
There were 52 genes associated with both spermatogenesis and astaxanthin. GJA1, NFKBIA, SOD2-OT1, TMX2-CTNND1, BDNF-AS, PTGS2, MIR7-3HG, LINC01672, EMSLR, and CERNA3 are the top 10 key targets according to the edge count value (Figure 1A–C). GO functional annotation revealed that “cellular response to chemical stress”, “response to hydrogen peroxide”, and “cellular response to oxidative stress” were the top 10 BP terms (Figure 1D), and the top 10 CC terms included “nuclear memory”, “mitochondrial intermembrane space”, and “nuclear envelope” (Figure 1E). The top 10 pathways in the MF category included ubiquitin protein ligase binding, ubiquitin-like protein ligase binding, NADP binding (Figure 1F), and so on. The KEGG analysis revealed that chemical carcinogenesis-reactive oxygen species, small cell lung cancer, Kaposi sarcoma-associated herpesvirus infection, the AGE-RAGE signaling pathway in diabetic complications, human T-cell leukemia virus 1 infection, hepatitis B, toxoplasmosis, the TNF signaling pathway, the NOD-like receptor signaling pathway, and Epstein‒Barr virus infection were the top 10 pathways, with P values in ascending order (Figure 1G–I).
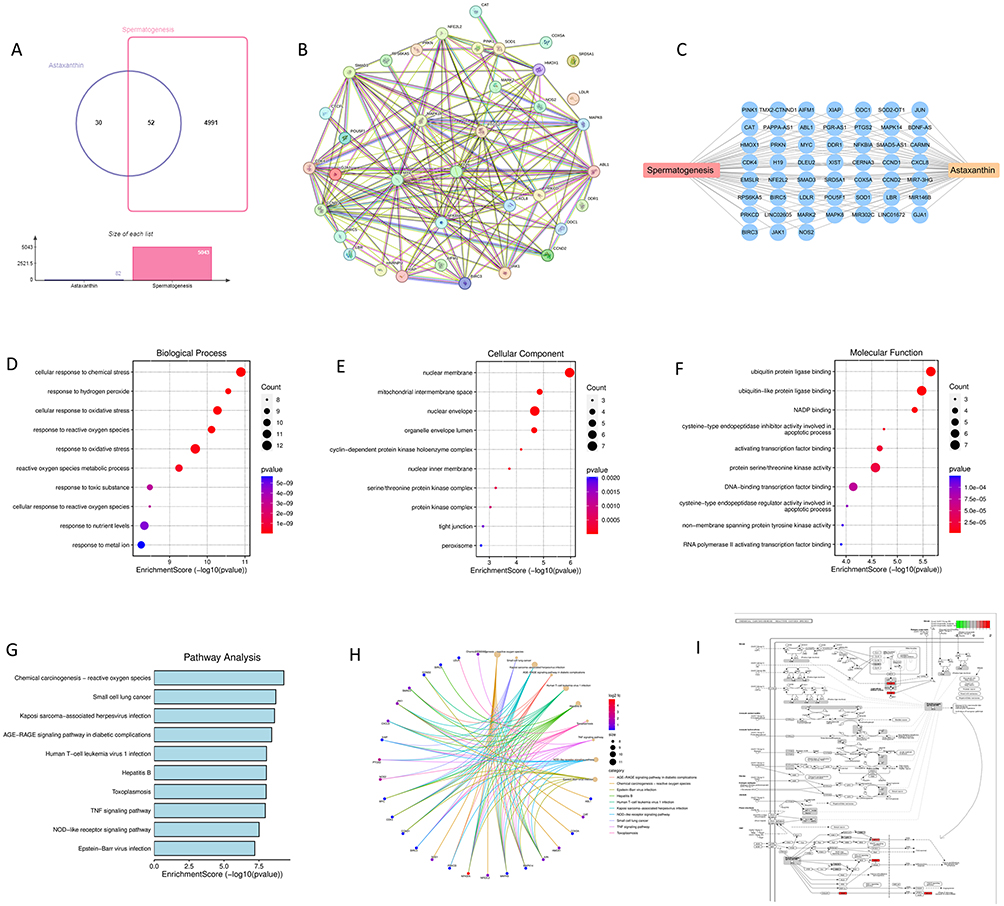 |
Figure 1 Network pharmacology analysis diagram. (A) Wayne diagram of the number of common genes related to spermatogenesis and astaxanthin. (B and C) Key targets of spermatogenesis and astaxanthin. (D) Top 10 pathways in the BP of KEGG analysis. (E) Top 10 pathways in the CC of KEGG analysis. (F) Top 10 pathways in the MF of KEGG analysis. (G–I) top 10 pathways of KEGG analysis. |
Changes in Mouse Body Weight
There was no significant difference in basic body weight among the model mice of Group A (n=5), Group B (n=5), and Group C (n=5) (Group A VS Group B P=0.775, Group A VS Group C P=0.450, Group C VS Group B P=0.635), but the body weight of the model mice was significantly lower than that of normal mice (Group D, n=5) of the same week of age (Group A, Group B, Group C VS Group D P<0.001). After feeding, the weight gain of the mice in Group A and Group B was significantly greater than that of those in Group C and Group D (Group A VS Group C p=0.032, Group A VS Group D P=0.040), but there was no difference between Group A and Group B (Group A VS Group B P=0.915) (Figures 2A–F).
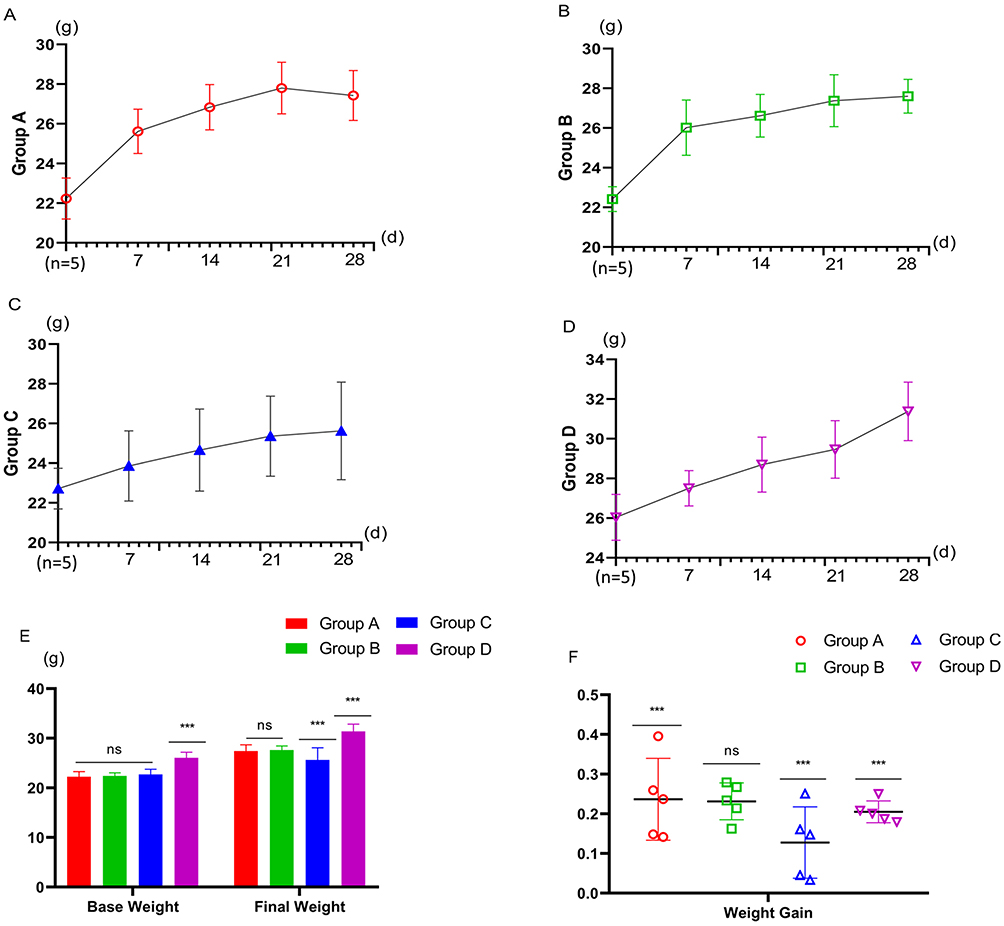 |
Figure 2 Weight change chart. (A) Weight changes in Group A. (B) Weight changes in Group B. (C) Weight changes in Group C. (D) Weight changes in Group D. (E) Comparison of body weight among 4 groups: Base weight: ns: P>0.05, Group A vs Group B, Group C. ***: P<0.01, Group A vs Group D. Final weight: ns: P>0.05, Group A vs Group B. ***: P<0.01, Group A vs Group C, Group D. (F) Comparison of body weight gain: ns: P>0.05, Group A vs Group B. ***: P<0.01, Group A vs Group C, Group D. |
Changes in Sperm Parameters of Mice
After feeding, the sperm concentration of the mice in Group A and Group B were significantly greater than those in Group C (P=0.012), and the forward motility sperm activity of the mice in Group B were significantly greater than those in Group C (P=0.024), but the sperm concentration and forward motility sperm activity of Group A, Group B and Group C were still significantly lower than those in Group D (Group A VS Group D P=0.001, 0.004, Group B VS Group D P<0.001, 0.018, Group C VS Group D P<0.001, <0.001). However, the increase of the forward motility sperm activity of the Group A and Group B was significantly greater than that of Group D (Group A VS Group B P=0.012, Group A VS Group C P=0.002), while there was no difference between the sperm concentration (Figure 3A–C). The CASA and HE results revealed that after feeding, spermatogenesis in Group A was not only significantly greater than that before feeding but also greater than that of Groups B and C, which was similar to that of Group D (Figure 3D–K).
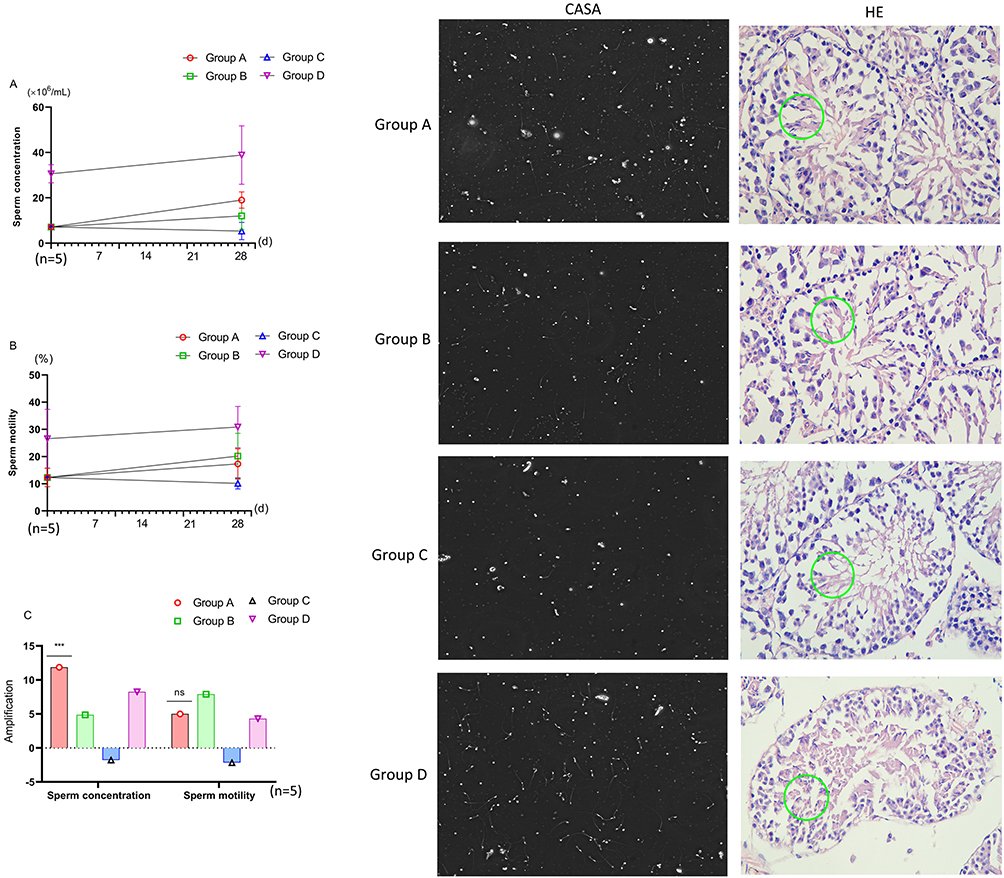 |
Figure 3 Comparison of sperm parameters. Green circle: Mature sperm. (A) Changes in sperm concentration in each group. (B) Changes in sperm motility in each group. (C) Comparison of sperm concentration increment: Group A vs Group B, P<0.01; Group A vs Group C, P<0.01; Group A vs Group D, ***: P<0.01. Comparison of sperm motility increment: Group A vs Group D, ns: P>0.05. CSAS: CASA analysis of sperm parameters. HE: Hematoxylin‒eosin staining of testicular tissue spermatogenesis status. |
Differences in Oxidative Stress Products in Mouse Testes
Before feeding, the MDA and ferrous ion levels of model mice (n=5) were significantly greater than those of normal mice (n=5) (MDA: P=0.044, Fe2+: P=0.006), However, the activity of GSH-Px enzyme showed no difference compared to normal mice (P=0.341). After feeding, the MDA, GSH-Px and ferrous ion concentrations in each group of model mice (Group A, Group B and Group C, n=5) were significantly changed than before, while the normal mice (Group D, n=5) had little change. After feeding, although the MDA of Group A, Group B and Group C showed no difference compared to Group D, but the GSH-Px enzyme activity had significantly difference (Group A VS Group D P<0.001, Group B VS Group D P<0.001, Group C VS Group D P<0.001). While the ferrous ion levels of Group A, Group B and Group C also showed difference change compared to Group D (Group A VS Group D P=0.085, Group B VS Group D P=0.026, Group C VS Group D P=0.010). The oxidative stress damage was effectively alleviated (Figure 4A–F).
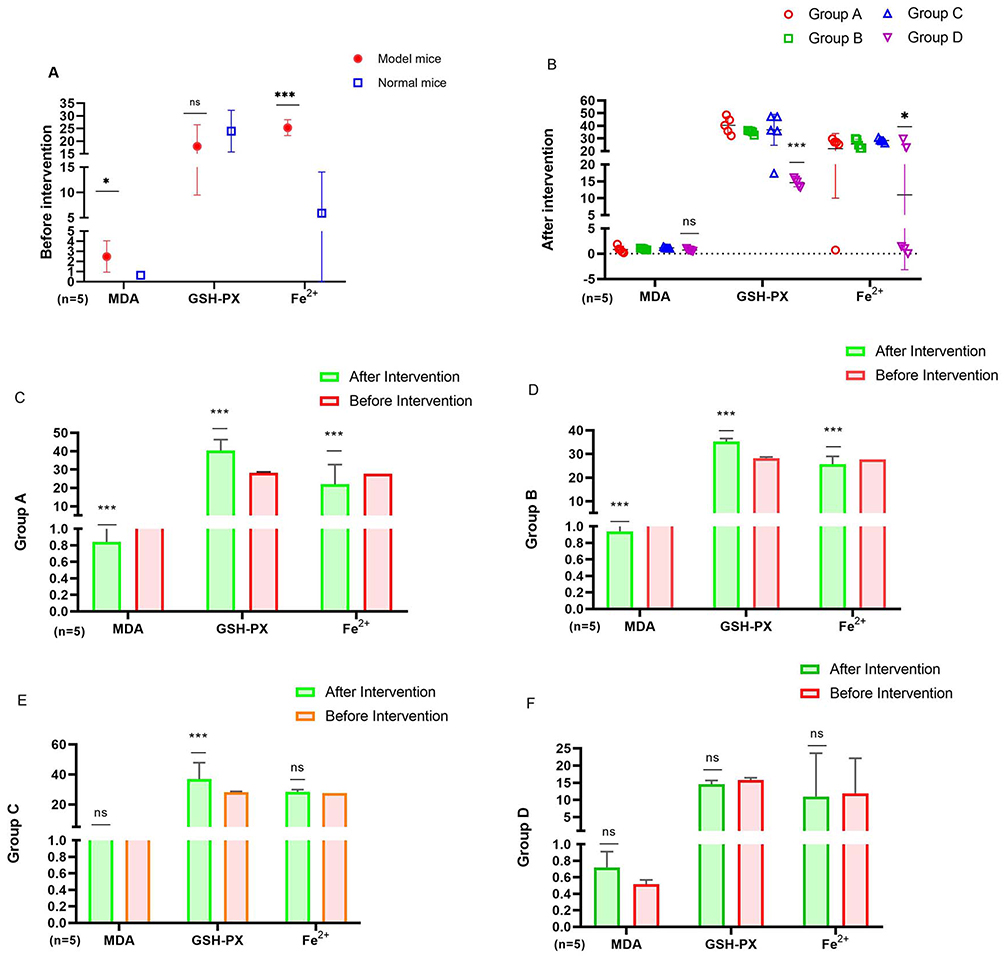 |
Figure 4 Comparison of indicators in mice before and after intervention. (A) Comparison pre intervention between model mice and normal mice: *, P<0.05, ***, P<0.01. (B) After intervention: Group A vs Group D, *, P<0.05, ***: P<0.01. (C) Comparison of Group A before and after intervention: * * * P<0.01, ns: p>0.05. (D) Comparison of Group B before and after intervention: * * * P<0.01, ns: p>0.05. (E) Comparison of Group C before and after intervention: * * * P<0.01, ns: p>0.05. (F) Comparison of Group D before and after intervention: * * * P<0.01, ns: p>0.05. |
Differences in Testicular Regulatory Proteins
Although the GPX4 level of the model mice (n=5) before feeding was significantly lower than that of the normal mice (n=5) (P<0.001), the GPX4 level of the model mice in Group A and Group B after feeding was significantly greater than that before treatment (P<0.001), especially that of Group A, which was even significantly greater than that of Group B (P<0.001). However, there was no significant difference between Group C and Group D (Figure 5A and B). WB revealed that the expression of GPX4 and VADC in the testicular tissue of Group A mice was lower than that in Group D mice after feeding, but the expression of GPX4 was significantly greater than that in Group B mice, while there was no significant difference in the VADC between Groups A and B (Figure 5C). IF also revealed that the deposition of the iron-promoting death protein glutaminase 2 (GLS2) in the seminiferous tubule epithelium of the model mice was significantly lower than that in the normal group, whereas the level of STEAP3 was significantly greater than that in the normal group (Figure 5D and E).
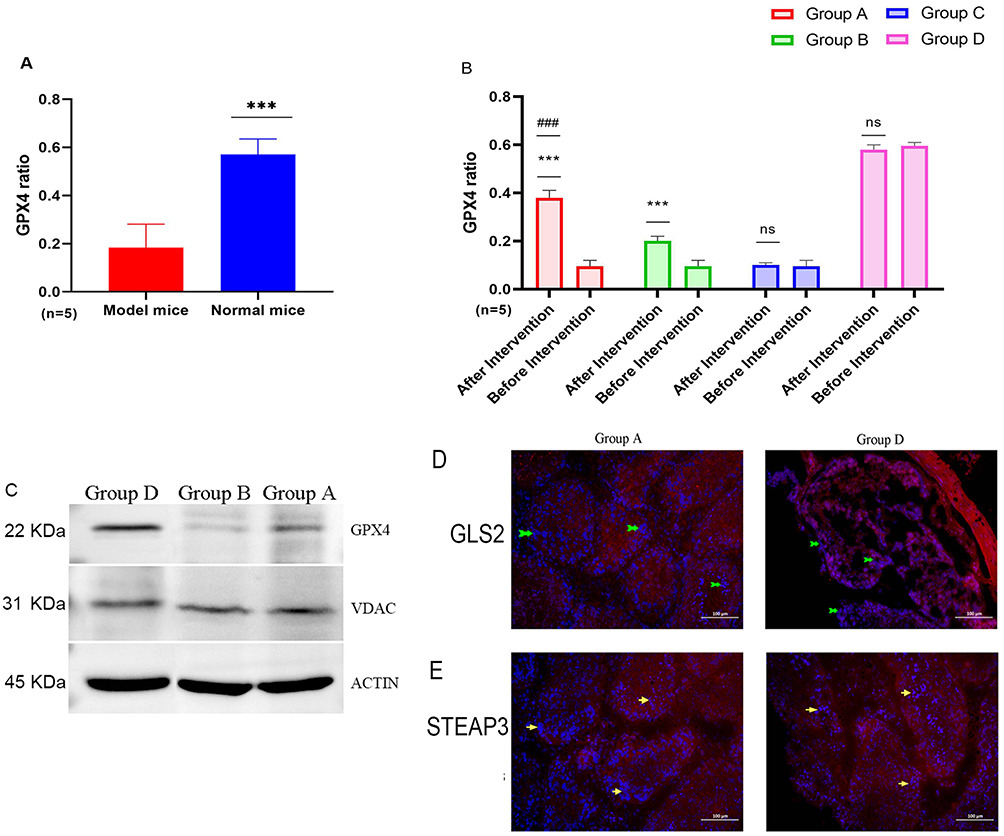 |
Figure 5 Comparison of factors in regulatory pathways. Green arrow: Strong expression of GLS2. Yellow arrow: Strong expression of STEAP3. (A) GPX4 of Model and Normal Mice: * * *: P<0.01. (B) GPX4 Preintervention vs postintervention: * * * P<0.01, ns: p>0.05. ###: Preintervention group A vs preintervention group D, P<0.01. (C) The expression of cytokines in groups. (D) Expression level of each factor: Blue fluorescence: CLS2. (E) Expression level of each factor: Blue fluorescence: STEAP3. |
Differences in the Proliferation of Spermatogonial Stem Cells
MTT assays revealed that the proliferation and inhibition rates of spermatogonial stem cells varied among the different concentrations of drugs cultured. The proliferation rate (PR) of the astaxanthin group (Group AA, n=5) at concentrations of 10 nmol/L and 15 nmol/L was significantly greater than that of the quercetin group (Group BB, n=5) and normal control group (Group CC, n=5) (10 nmol/L: Group AA VS Group BB P=0.003, Group AA VS Group CC P=0.001. 15 nmol/L: Group AA VS Group BB P=0.001, Group AA VS Group CC P=0.005), and the inhibition rate (IR) of Group AA was significantly lower than that of Group CC (10 nmol/L: P=0.025, 15 nmol/L: P=0.015), and the inhibition rate (IR) of Group AA was significantly lower than that of Group BB in the 15 nmol/L group (P=0.003). However, there was no significant difference in the proliferation rate or inhibition rate of the 5 nmol/L group. When the astaxanthin reached 10 nmol/l, it had the highest proliferation rate and the lowest inhibition rate on spermatogonial stem cells (Figure 6A–D).
Figure 6 Continued. Figure 6 MTT Results of Spermatogonial Stem Cells. (A) Inhibition rates of different drug concentrations: Group AA vs Group BB Group CC * * *: P<0.01; Group AA vs Group BB Group CC; ns: p>0.05. (B) Proliferation rates of different drug concentrations: Group AA vs Group BB Group CC * * *: P<0.01; Group AA vs Group BB Group CC; ns: p>0.05. (C) The trend of inhibition rates changes of each group at different drug concentrations. (D) The trend of proliferation rates changes of each group at different drug concentrations. (E) The proliferation changes of spermatogonial stem cells in group BB at different times. (F) The proliferation changes of spermatogonial stem cells in group AA at different times. (G) The proliferation changes of spermatogonial stem cells in group CC at different times.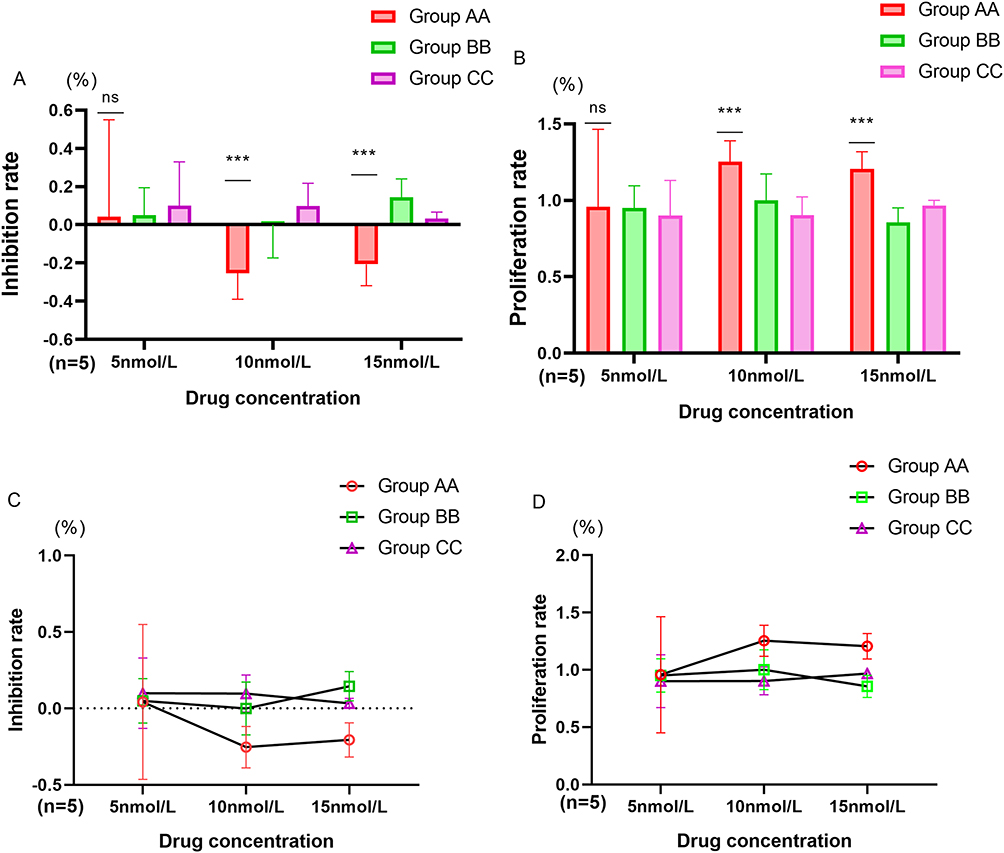

Through microscopic observation at different times, it was found that the growth status of spermatogonial stem cells was inconsistent after different concentrations of drugs were cultured for different durations, and there were significant differences in the degree of differentiation into sperm. Among them, after 8 hours of cultivation, the Group AA with the 10 nmol/l presented the greatest degree of sperm formation, and after 12 h, it was relatively stable (Figure 6E–G).
Differences in the Metabolites of Spermatogonial Stem Cells
The culture medium of cells cultured in vitro was collected, and ELISA was used to detect cell metabolites. The results showed that the metabolic products of spermatogonial stem cells varied with different concentrations of drug. The secretion of the iron-promoting death protein ACSL3 by the spermatogonial stem cells cultured in vitro in Group AA was significantly lower than that in the Group BB (5 nmol/L: P<0.001, 10 nmol/L: P<0.001, 15 nmol/L: P<0.001), but without differences with Group CC (5 nmol/L: P=0.058, 10 nmol/L: P=0.532, 15 nmol/L: P=0.438). The level of the antagonistic iron death protein FADS2 in Group AA was significantly greater than that other two groups (5 nmol/L: Group AA VS Group BB P=0.007, Group AA VS Group CC P=0.001, 10 nmol/L: Group AA VS Group BB P=0.004, Group AA VS Group CC P<0.001, 15 nmol/L: Group AA VS Group BB P<0.001, Group AA VS Group CC P<0.001). Among which the concentration of ACSL3 secreted by spermatogonial stem cells cultured with 10 nmol/L astaxanthin was the lowest, while that of FADSL2 with 15 nmol/L was the highest (Figure 7A–D).
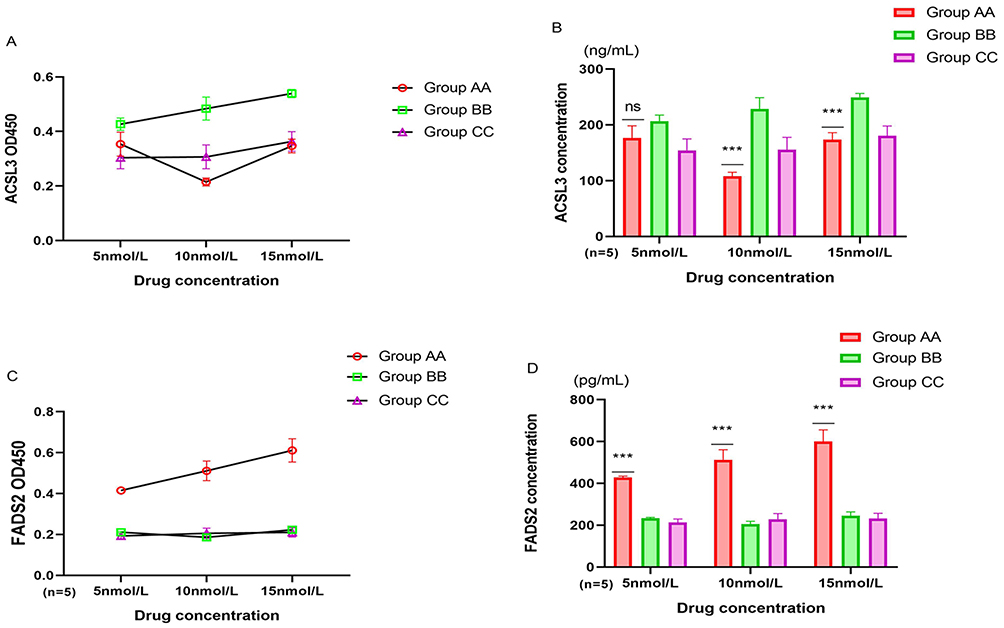 |
Figure 7 Comparison of metabolic products of spermatogonial stem cells. (A) OD450 value of ACSL3. (B) Comparison of ACSL3 concentration: Group AA vs Group BB Group CC * * *: P<0.01, ns: p>0.05. (C) OD450 value of FADS2. (D) Comparison of FADS2 concentration: Group AA vs Group BB Group CC * * *: P<0.01. |
Differences in the Apoptosis of Spermatogonial Stem Cells
Cultured spermatogonial stem cells were collected, and their apoptosis rate was detected via the Annexin V-FITC/PI flow detection method. The results indicate that the apoptosis rate of spermatogonial stem cells cultured with astaxanthin (Group AA) was significantly lower than that of the quercetin group (Group BB) and the physiological saline control group (Group CC) (Figure 8).
 |
Figure 8 Apoptosis of spermatogonial stem cells. Group AA vs Group BB, Group AA vs Group CC: P<0.01. |
Discussion
Fat is crucial for maintaining physiological functions in the human body, but changes in lifestyle can easily lead to excessive accumulation of fat. However, when there is excessive fat, a large amount of free radicals are produced, which can trigger oxidative stress reactions, leading to collective imbalance, cell damage, protein oxidation, DNA oxidation, lipid peroxidation, and the occurrence of various diseases.
We analyzed the keywords astaxanthin and spermatogenesis through network pharmacology and revealed that oxidative stress plays an important role in the process of spermatogenesis and that astaxanthin can act on multiple target genes, including important targets such as SOD and NFKB, during this process. Among the complex regulatory pathways associated with astaxanthin, we found that lipid metabolism is also one of the key pathways affecting spermatogenesis, especially in obese individuals. Lipid metabolism may be more closely related to fat transition. Therefore, we validated this hypothesis through in vivo and in vitro experiments.
The impact of lipid metabolism on spermatogenesis has become a hot topic of international research. Studies have shown that sperm membranes are composed of complex lipid mixtures, such as phospholipids, glycolipids, and sterols.20 These lipid substances have good fluidity, and the metabolism of these unsaturated fatty acids plays an important role in spermatogenesis as sperm fertilization ability changes. Some studies have also shown that mammalian testicular germ cells selectively synthesize large amounts of sulfogalactosylglycerides (SGGs), which directly participate in cell adhesion and are degraded into lipid skeletons by lysosomal enzymes in testicular Sertoli cells.21,22 When this balance is disrupted, Sertoli cells are prone to lysosomal storage and subdysfunction, thereby affecting their support for germ cell development and leading to a decrease in fertility.23 The toxic effect of malondialdehyde on germ cells, ultimately leading to spermatogenesis disorders, has been confirmed. Hammami et al reported that long-term consumption of groundnut grass in rats resulted in a significant increase in malondialdehyde (MDA) in testicular tissue; a decrease in catalase and SOD activity; and inhibition of the entire antioxidant defense system, leading to changes in the sperm concentration, activity, and morphology and disruption of normal spermatogenesis.24 Malondialdehyde (MDA), 4-hydroxynonenal (HNE), and F215 (S)-8-isoprostaglandin F2 α (15 (S)-8-IsoPGF2 α) are products of lipid metabolism and the most commonly measured biomarkers of lipid peroxidation. The degree of oxidative stress can be reliably evaluated by measuring the levels of MDA, HNE, and 15(S)-8-isoPGF.25 In a mouse model of oligoasthenozoospermia induced by cyclophosphamide, the seminiferous tubule skeleton of the testes was severely damaged, a large number of Sertoli cells shed, and sperm development was severely inhibited. Through testing, we also found that the MDA content in the testes of these model mice was significantly greater than that in the testes of normal mice and that their sperm parameters and body weights were significantly lower than those of normal mice. Thus, the testicular lipid metabolism of these oligoasthenozoospermia model mice is definitely abnormal. After intervention with different drugs, the MDA content of the model mice changed. Therefore, our intervention experiment should be able to test the effects of drugs on lipid metabolism.
The glutathione peroxidase (GPx) family is extremely sensitive to oxidative attacks related to lipid peroxidation, DNA damage, and impaired sperm motility and can control the amount of hydrogen peroxide in male gametes. Among them, GPx4 and GPx5 play important roles in determining the fertilization potential of mammalian sperm.26 Ursini et al reported that Px4 plays an important role in inflammation, immunity, and differentiation during spermatogenesis, mainly by acting on mitochondria and nuclei. It catalyzes the oxidation of protein thiols during the stabilization of the middle stage of sperm maturation and chromatin densification.27 Research has shown that by consuming a combination of zinc and folic acid, the antioxidant astaxanthin (Astacalox), or energy containing (acetyl)carnitine (Proxed) as a supplement, the quality and function of sperm can be improved.28 The clinical observation results of Asian men also suggest that the combination of multiple antioxidants (L-carnitine, zinc, astaxanthin, coenzyme Q10, vitamin C, vitamin B12, and vitamin E) can significantly increase the total number of active sperm and promote an increasing trend in semen concentration, semen vitality, and total number of active sperm while not affecting endocrine results.29 Our results also confirmed this conclusion, but although the number of sperm in the model mice fed with astaxanthin and quercetin did not reach the level of normal mice, their growth rate was significantly greater than that of normal mice, especially in the astaxanthin group, which presented the greatest increase in sperm concentration, which may be related to the fact that astaxanthin can promote spermatogenesis. Therefore, we detected and analyzed the relevant links in the process of spermatogenesis to explore the mechanism of this phenomenon. We found that after different types of drug intervention, the weight and sperm parameters of the oligoasthenozoospermia model mice showed varying concentrations, especially in the mice treated with astaxanthin. Although some of these indicators did not reach the level of normal mice, there was a significant increase in GSH-Px and GPX4 in their testicular tissue, which indicates that our astaxanthin can effectively reduce the degree of oxidative stress damage, reduce the risk of lipid metabolism, promote the recovery of the entire antioxidant defense system, maximize the inhibition of MDA damage in the testicular microenvironment, promote the redox protection system, and thus play a role in improving spermatogenesis.
The metabolism of drugs in vivo is extremely complex, with many influencing factors and uncertainties. Therefore, we conducted in vitro cell experiments. Spermatogenic stem cells are a group of reproductive cells with the ability to proliferate and differentiate. After effective cultivation and division, they can progress into sperm. Research has shown that astaxanthin, an effective antioxidant and powerful reactive oxygen species (ROS) scavenger, has a protective effect on BU-induced toxicity in human spermatogonial stem cells. AST can significantly upregulate the expression of the NRF-2 gene and protein, significantly reduce the production of ROS induced by BU, activate the NRF-2/HO-1 pathway, and inhibit BU-induced SSC apoptosis.30 Experiments have shown that quercetin can reduce the increase in superoxide anions in spermatogonial stem cells induced by diethylstilbestrol in vitro and reduce DNA damage in spermatogonial stem cells.31 In vitro, we also cultured spermatogonial stem cells and conducted drug-controlled observation experiments using quercetin. The results showed that the proliferation and inhibition rates of spermatogonial stem cells in different groups varied at different time points after drug cultivation at different concentrations. Among them, 10 nmol/L astaxanthin had the highest proliferation rate and the lowest inhibition rate on spermatogonial stem cells. After 8 h of cultivation, the 10 nmol/L astaxanthin group presented the greatest degree of sperm formation in the mice, and after 12 h, the degree of sperm formation was relatively stable, with a significantly greater effect than that of quercetin. Therefore, we determined that the promotion of the proliferation of spermatogonial stem cells by astaxanthin may be due not only to the repair function of oxidative stress damage but also to the regulation of other metabolic substances and targets.
To further analyze the stronger promoting effect of astaxanthin than quercetin on the proliferation of spermatogonial stem cells, we analyzed and tested the culture media of each drug after cultivation. We found that the secretion of the iron-promoting death protein ACSL3 by the original stem cells of the astaxanthin group after in vitro cultivation was significantly lower than that of the saline control group and quercetin group. The antagonistic iron death protein FADS2 was significantly higher than that of these two groups, among which the concentration of ACSL3 secreted by spermatogonial stem cells cultured with 10 nmol/L astaxanthin was the lowest, and that of FADSL2 was the highest. Previous studies have shown that reversing transcriptional UTP14 gene insertion or mutation in the intron of ACSL3 can hinder spermatogenesis and that reducing the expression of ACSL3 may help promote spermatogenesis.32,33 We also observed this phenomenon via in vitro culture experiments. Interestingly, we found for the first time that after drug intervention, FADSL2 in spermatogonial stem cells also changed, and FADSL2 expression increased in the group with increased cell proliferation. A comparison of astaxanthin, quercetin, and physiological saline revealed that the proliferation rate of spermatogonial stem cells in the astaxanthin group was the highest. We believe that this may suggest that astaxanthin has a better effect on regulating fat metabolism than does quercetin . By detecting the rate of cell apoptosis, we also found that the rate of apoptosis in spermatogonial stem cells cultured with astaxanthin was significantly lower than that in the quercetin group and the saline control group. This finding further confirms our hypothesis; therefore, we believe that astaxanthin can alleviate the damage caused by iron death by regulating fat metabolism, promote the proliferation and differentiation of spermatogonial stem cells, and thus promote spermatogenesis.
Conclusion
In summary, astaxanthin can not only increase the body weight and sperm parameters of cyclophosphamide-induced oligoasthenozoospermia model mice but also regulate the concentrations of metabolic products such as iron ions, MDA, and GSH-Px during iron death in testicular tissue; enhance the expression of GPX4, GLS2, Steap3, and VDAC; and promote sperm development. Its effect is more ideal than that of quercetin, possibly by regulating the lipid metabolites ACSL3 and FADS2.
Shortcomings
Although we discovered through animal experiments and in vitro cell experiments a potential new mechanism by which astaxanthin regulates spermatogenesis by affecting lipid metabolism, our experiments still have several shortcomings, such as (1) the observation cycle of animal experiments is short, and long-term effect analysis is lacking; (2) there is a lack of human experiments to accurately determine the effect of astaxanthin on the human body; and (3) the mechanism we analyzed is not yet fully understood, and the action sites regulating lipid metabolism are still not clear enough. Wait a minute. However, through this preliminary exploration and analysis, we have revealed a new direction for the efficacy of astaxanthin. Therefore, we believe that we will further improve and deepen future research to address the aforementioned shortcomings and confirm the new efficacy of astaxanthin.
Abbreviations
ACSL3, Acyl-CoA synthetase long-chain family member 3; VDAC, Voltage-dependent anion channel; GPX4, glutathione peroxidase 4; FADS2, fatty acid desaturase 2; GLS2, glutaminase 2; MDA, malondialdehyde; GSH-Px, Glutathione peroxidase; SGGs, sulfogalactosylglycerides; SREBP2, sterol regulatory element binding protein 2; APOA1, apolipoprotein A1; ROS, Reactive oxygen species; MF, Molecular function; CC, Cellular component; BP, Biological process; CASA, Computer-aided sperm analysis; DMSO: Dimethylsulfoxide; GJA1, gap junction protein, alpha 1; SOD2, Superoxide Dismutase 2; BDNF, Brain-derived neurotrophin factor; PTGS2, prostaglandin-endoperoxide synthase 2; CERNA3, competing endogenous RNA; NFKB, nuclear factor-k gene binding; HNE, 4-hydroxynonenal; NRF-2, Nuclear factor erythroid 2-related factor 2; HO-1, haem oxygenase 1; PR, proliferation rate; IR, inhibition rate.
Data Sharing Statement
The data of my article has been shared on Mendeley Data, and the link is https://data.mendeley.com/datasets/4x29x6vc3f/1 and https://data.mendeley.com/datasets/hngvv3s3b9/1.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the Guangdong Basic and Applied Basic Research Foundation (No. 2021A1515110921), the General Guidance Project of Guangzhou Health Science and Technology (20241A010111), and the Key Program of Medical and Health Project of Panyu Science and Technology Funds (2023--Z04--109).
Disclosure
The authors declare that they have no competing interests.
References
1. Feng-feng TIAN, Hou-zhao WANG. Effect of occupational lead exposure on male semen quality. Chin J Eugenics Genetics. 2017;25(07):119–120. doi:10.13404/j.cnki.cjbhh.2017.07.046
2. Calvert L, Green MP, De Iuliis GN, et al. Assessment of the emerging threat posed by perfluoroalkyl and polyfluoroalkyl substances to male reproduction in humans. Front Endocrinol. 2022;12:799043. doi:10.3389/fendo.2021.799043
3. Lv MQ, Wang HX, Yang YQ, et al. Semen quality following long-term occupational exposure to formaldehyde in China [published correction appears in JAMA Netw Open. 2022 Oct 3;5(10):e2238925]. JAMA Netw Open. 2022;5(9):e2230359. doi:10.1001/jamanetworkopen.2022.30359
4. Jimenez-Villarreal J, Betancourt-Martinex ND, Carranza-Rosales P, et al. Formaldehyde induces DNA strand breaks on spermatozoa and lymphocytes of Wistar rats. TSitologiia i genetika. 2017;51(1):78–80.
5. Yotbuntueng P, Jiemsup S, Deenarn P, et al. Differential distribution of eicosanoids and polyunsaturated fatty acids in the Penaeus monodon male reproductive tract and their effects on total sperm counts. PLoS One. 2022;17(9):e0275134. doi:10.1371/journal.pone.0275134
6. Signorini C, Moretti E, Noto D, et al. Fatty acid oxidation and pro-resolving lipid mediators are related to male infertility. Antioxidants. 2022;11(1):107. doi:10.3390/antiox11010107
7. Gautier C, Aurich C. “Fine feathers make fine birds” – the mammalian sperm plasma membrane lipid composition and effects on assisted reproduction. Anim Reprod Sci. 2022;246:106884. doi:10.1016/j.anireprosci.2021.106884
8. Fenghua L, Dongzi Y, Yifeng W, et al. Making of the animal model with sterilized testes[J]. Natl J Androl. 2007;02:125–129. doi:10.13263/j.cnki.nja.2007.02.006
9. Tanphaichitr N, Kongmanas K, Faull KF, et al. Properties, metabolism and roles of sulfogalactosylglycerolipid in male reproduction. Prog Lipid Res. 2018;72:18–41. doi:10.1016/j.plipres.2018.08.002
10. MInguez-Alarcón L, Chavarro JE, Mendiola J, et al. Fatty acid intake in relation to reproductive hormones and testicular volume among young healthy men. Asian J Androl. 2017;19(2):184–190. doi:10.4103/1008-682X.190323
11. Jarvis S, Gethings LA, Samanta L, et al. High fat diet causes distinct aberrations in the testicular proteome. Int J Obes Lond. 2020;44(9):1958–1969. doi:10.1038/s41366-020-0595-6
12. Setayesh T, Nersesyan A, Mišík M, et al. Impact of obesity and overweight on DNA stability: few facts and many hypotheses. Mutat Res Rev Mutat Res. 2018;777:64–91. doi:10.1016/j.mrrev.2018.07.001
13. Nishida Y, Berg PC, Shakersain B, et al. Astaxanthin: past, present, and future. Mar Drugs. 2023;21(10):514. doi:10.3390/md21100514
14. Biyik AF, Yulug E, Yenilmez E, et al. Effects of quercetin on gentamicin-induced experimental testicular injury in rats. Rom J Morphol Embryol. 2024;65(1):69–80. doi:10.47162/RJME.65.1.09
15. Panchal H, Bhardwaj JK. Quercetin supplementation alleviates cadmium induced genotoxicity-mediated apoptosis in caprine testicular cells. Biol Trace Elem Res. 2024;202(10):1–14. doi:10.1007/s12011-023-04038-8
16. Wu J-Y, Kang H-Y, Guo Y, et al. Effect of natural astaxanthin on sperm quality and mitochondrial function of breeder rooster semen cryopreservation. Cryobiology. 2024;117:104979. doi:10.1016/j.cryobiol.2024.104979
17. Guo H-T, Wang J-R, Sun L-Z, et al. Effects of astaxanthin on plasma membrane function and fertility of boar sperm during cryopreservation. Theriogenology. 2021;164:58–64. doi:10.1016/j.theriogenology.2021.01.007
18. Nair AB, Jacob S. A simple practice guide for dose conversion between animals and human. J Basic Clin Pharm. 2016;7(2):27–31. doi:10.4103/0976-0105.177703
19. Ji-Han HUANG, Xiao-Hui HUANG, Zhi-Yang CHEN, et al. Dose conversion among different animals and healthy volunteers in pharmacological study. Chin J Clin Pharmacol Ther. 2004;(09):1069–1072.
20. Pakpahan C, Rezano A, Margiana R, et al. The association between lipid serum and semen parameters: a systematic review. Reprod Sci. 2023;30(3):761–771. doi:10.1007/s43032-022-01040-8
21. Serafini S, O’Flaherty C. Sphingolipids modulate redox signalling during human sperm capacitation. Hum Reprod. 2025;40(2):210–225. doi:10.1093/humrep/deae268
22. Serafini S, O’Flaherty C. Novel insights into the lipid signalling in human spermatozoa. Hum Reprod. 2025;deaf085. doi:10.1093/humrep/deaf085
23. Do Nascimento CC, Aguiar O Jr, Viana GM, et al. Morphological damage in Sertoli, myoid and interstitial cells in a mouse model of mucopolysaccharidosis type I (MPS I). Mol Biol Rep. 2021;48(1):363–370. doi:10.1007/s11033-020-06055-5
24. Hammami I, Ali RB, Nahdi A, et al. Chronic consumption of Hypericum humifusum leaf extracts impairs epididymis spermatozoa characters in association with oxidative stress in adult male Wistar rats. Biomed Pharmacother. 2017;93:616–625. doi:10.1016/j.biopha.2017.06.069
25. Tsikas D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: analytical and biological challenges. Anal Biochem. 2017;524:13–30. doi:10.1016/j.ab.2016.10.021
26. Chabory E, Damon C, Lenoir A, et al. Mammalian glutathione peroxidases control acquisition and maintenance of spermatozoa integrity 1. J Anim Sci. 2010;88(4):1321–1331. doi:10.2527/jas.2009-2583
27. Ursini F, Bosello Travain V, Cozza G, et al. A white paper on phospholipid hydroperoxide glutathione peroxidase (GPx4) forty years later. Free Radic Biol Med. 2022;188:117–133. doi:10.1016/j.freeradbiomed.2022.06.227
28. Comhaire FH, Mahmoud A. The role of food supplements in the treatment of the infertile man. Reprod Biomed Online. 2003;7(4):385–391. doi:10.1016/s1472-6483(10)61881-0
29. Terai K, Horie S, Fukuhara S, et al. Combination therapy with antioxidants improves total motile sperm counts: a Preliminary Study. Reprod Med Biolog. 2020;19(1):89–94. doi:10.1002/rmb2.12308
30. Afzali A, Amidi F, Koruji M, et al. Astaxanthin relieves busulfan-induced oxidative apoptosis in cultured human spermatogonial stem cells by activating the Nrf-2/HO-1 pathway. Reprod Sci. 2022;29(2):374–394. doi:10.1007/s43032-021-00651-x
31. Habas K, Brinkworth MH, Anderson D. Diethylstilbestrol induces oxidative DNA damage, resulting in apoptosis of spermatogonial stem cells in vitro. Toxicology. 2017;382:117–121. doi:10.1016/j.tox.2017.03.013
32. Rohozinski J, Lamb DJ, Bishop CE. UTP14c is a recently acquired retrogene associated with spermatogenesis and fertility in man. Biol Reprod. 2006;74(4):644–651. doi:10.1095/biolreprod.105.046698
33. Zhao M, Rohozinski J, Sharma M, et al. Utp14b: a unique retrogene within a gene that has acquired multiple promoters and a specific function in spermatogenesis. Dev Biol. 2007;304(2):848–859. doi:10.1016/j.ydbio.2007.01.005
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.