Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 15
Association Between SNPs in the One-Carbon Metabolism Pathway and the Risk of Female Breast Cancer in a Chinese Population
Authors Wang X, Xiong M, Pan B, Cho WC ![]() , Zhou J, Wang S, He B
, Zhou J, Wang S, He B ![]()
Received 9 July 2021
Accepted for publication 3 November 2021
Published 11 January 2022 Volume 2022:15 Pages 9—16
DOI https://doi.org/10.2147/PGPM.S328612
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Xuhong Wang,1,2,* Mengqiu Xiong,2,* Bei Pan,1,2 William CS Cho,3 Jin Zhou,4 Shukui Wang,1,2,5 Bangshun He2,5
1School of Medicine, Southeast University, Nanjing, Jiangsu Province, 210096, People’s Republic of China; 2Department of Laboratory Medicine, Nanjing First Hospital, Nanjing Medical University, Nanjing, Jiangsu Province, 210006, People’s Republic of China; 3Department of Clinical Oncology, Queen Elizabeth Hospital, Hong Kong, People’s Republic of China; 4Department of General Surgery, Nanjing First Hospital, Nanjing Medical University, Nanjing, People’s Republic of China; 5Jiangsu Collaborative Innovation Center on Cancer Personalized Medicine, Nanjing Medical University, Nanjing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Shukui Wang
School of Medicine, Southeast University, Nanjing, Jiangsu Province, 210096, People’s Republic of China
Email [email protected]
Bangshun He
Department of Laboratory Medicine, Nanjing First Hospital, Nanjing Medical University, Nanjing, Jiangsu Province, 210006, People’s Republic of China
Email [email protected]
Objective: The aim of this study is to assess the relationship between the single-nucleotide polymorphism (SNP) in the one-carbon metabolism pathway (MTR rs1805087; MTHFR rs1801133; ALDH1L1 rs2002287, rs2276731; DNMT1 rs16999593, rs2228611; DNMT3B rs2424908) and the risk of female breast cancer (BC) in a Chinese population.
Methods: A population-based case–control study was conducted, involving a total of 439 BC patients and 439 age-matched healthy controls. We adopted Sequence MASSarray to identify genotyping, and used immunohistochemistry (IHC) to test the expression of estrogen receptor (ER), progesterone receptor (PR) and human epidermal growth factor receptor-2 (HER-2) in tumor tissue.
Results: We found that rs16999593 (TC/CC vs TT: adjusted OR=1.38, 95% CI: 1.03– 1.84, p=0.030) was associated with an increased risk of BC, while rs2228611 was related to a decreased BC risk (GA/AA vs GG: adjusted OR=0.74, 95% CI: 0.56– 0.97, p=0.030). In addition, stratified analysis revealed that DNMT1 rs16999593, rs2228611 and ALDH1L1 rs2002287 contributed to the risk of BC, with associations with ER, PR and HER-2 expression.
Conclusion: In summary, this study revealed that DNMT1 rs16999593 and rs2228611 were associated with BC risk.
Keywords: one-carbon metabolism, DNA methylation, DNMT1, SNP, breast cancer
Background
According to the Global Cancer Statistics 2020 report, breast cancer (BC) is ranked as the most common malignant tumor in females in the world, and there were 19.3 million new cases of BC and 10.0 million deaths worldwide in 2020. Moreover, the incidence rate of new cases of BC (11.7%) is the highest among female cancers.1 Several risk factors contribute to the occurrence of BC, including environmental and genetic background risk factors. Aging, family history, reproductive factors, estrogen levels and lifestyle are regarded as risk factors for BC.2 In addition, genetic factors, such as mutations in BRCA1/2, HER2, EGFR and c-Myc, are regarded to contribute to the risk of BC, and single-nucleotide polymorphisms (SNPs) are observed to be associated with the risk of BC.3–6
One-carbon metabolism is a necessary process for nucleotide synthesis, and DNA replication, repair and methylation, and it is up-regulated during the pathology and progression of cancer.7 One-carbon metabolism plays a critical role in the establishment and regulation of tissue-specific patterns of methylated cytosine residues. Disordered one-carbon metabolism may lead to the disruption of DNA replication and repair, and to abnormal DNA methylation patterns, which eventually contribute to the occurrence of cancer.8 Previous research has shown that the corresponding genes must be silenced by DNA methylation to maintain the survival of cancer cells.9 Moreover, the one-carbon metabolic pathway has been reported to be associated with the risk of cancers,10–12 including BC.13 A higher intake of vitamin B, which is involed in folate-mediated one-carbon metabolic pathways predicts a higher risk of endometrial cancer.14 However, a high intake of vitamins related to one-carbon metabolism may help prevent BC, especially ER+/PR+ BC,15 as well as hepatocellular cancer16 and lung cancer.17
The SNPs in the genes of one-carbon metabolism, including methionine synthase (MTR), methylenetetrahydrofolate reductase (MTHFR), 10-formyltetrahydrofolate dehydrogenase (ALDH1L1), DNA methyltransferases (DNMTs), thymidylate synthase (TYMS) and reduced folate carrier 1 (RFC1), have been suggested to contribute to the occurrence of cancers.18,19 MTR rs1805087 is associated with the risk of gastric cancer20 and prostate cancer,21 and MTHFR rs1801133 was reported to reduce the susceptibility to hepatocellular cancer,22 while contributing to the risk of cervical cancer.23 In addition, ALDH1L1 rs2276724 was associated with the risk of HBV-associated hepatocellular cancer.24 However, the SNPs occurring in these genes are rarely reported in BC, especially in the Chinese population, in whom the incidence of BC is increasing year by year;25 a study by Sun et al found that DNMT1 rs16999593 was associated with an increased BC risk in Han Chinese women from South China.26 In addition, SNPs occouring in gene may affect the expression or activity of enzymes, contributing to the susceptibility to cancers. Here, to assess the contribution of the SNPs in the genes of one-carbon metabolism (MTR rs1805087; MTHFR rs1801133; ALDH1L1 rs2002287, rs2276731; DNMT1 rs16999593, rs2228611; DNMT3B rs2424908) to BC risk, we conducted this population-based case–control study in a Chinese female population.
Materials and Methods
Study Population
We recruited 439 patients with pathologically confirmed BC as the case group (mean age 52.89±10.78 years) from the Nanjing First Hospital, Nanjing Medical University, and 439 healthy people who came to hospital for a physical examination as the control group (mean age 52.95±10.89 years).27 Information including age, smoking and drinking status was collected by questionnaire. Owing to the small number of individuals with smoking and drinking habits, the inclusion criteria were no history of smoking, no history of drinking, and no personal or family history of cancer.
In addition, we collected the clinical pathological characteristics, including tumor grade, pathological stage and lymph-node metastasis, from clinical records. All participants gave their written informed consent, and this study was approved by the Institutional Review Committee of Nanjing First Hospital. Moreover, this study was conducted in accordance with the Declaration of Helsinki.
SNP Genotyping
Blood samples of patients were collected with anticoagulant tubes containing EDTA for genotyping, and the genotyping was performed based on Sequence MASSarray. Finally, SequenomTyper 4.0 software was used for data management and evaluation, as previously described.27,28
Immunohistochemistry (IHC) Assay
Estrogen receptor (ER), progesterone receptor (PR) and human epidermal growth factor receptor-2 (HER-2) expression in paraffin-embedded tumor tissues was detected by IHC assay, following the instructions for the kit. The monoclonal rabbit ER, PR and HER-2 antibody used in this study were purchased from Spring Bioscience (Pleasanton, CA, USA).
Statistical Analysis
The t-test or fitted chi-squared test was used to evaluate differences in clinical characteristics between the two groups. Evaluation of Hardy–Weinberg equilibrium (HWE) in the healthy control group was assessed by the two-sided chi-squared test. SAS-based logistic regression was used to calculate the odds ratio (OR) and 95% confidence interval (CI) to assess the relationship of SNPs with BC risk. To obtain more rigorous statistical results, we used adjusted ORs to reduce the potential effects of age and menopausal status. The data were analyzed using SPSS 23.0 software (IBM Corp, Armonk, NY, USA), with a p-value <0.05 indicating a statistically significant difference.
Results
The genotypic distributions of all SNPs were not biased in HWE (Table 1). There was no significant difference in age between the case and control groups (p=0.933), as summarized in Table 2.
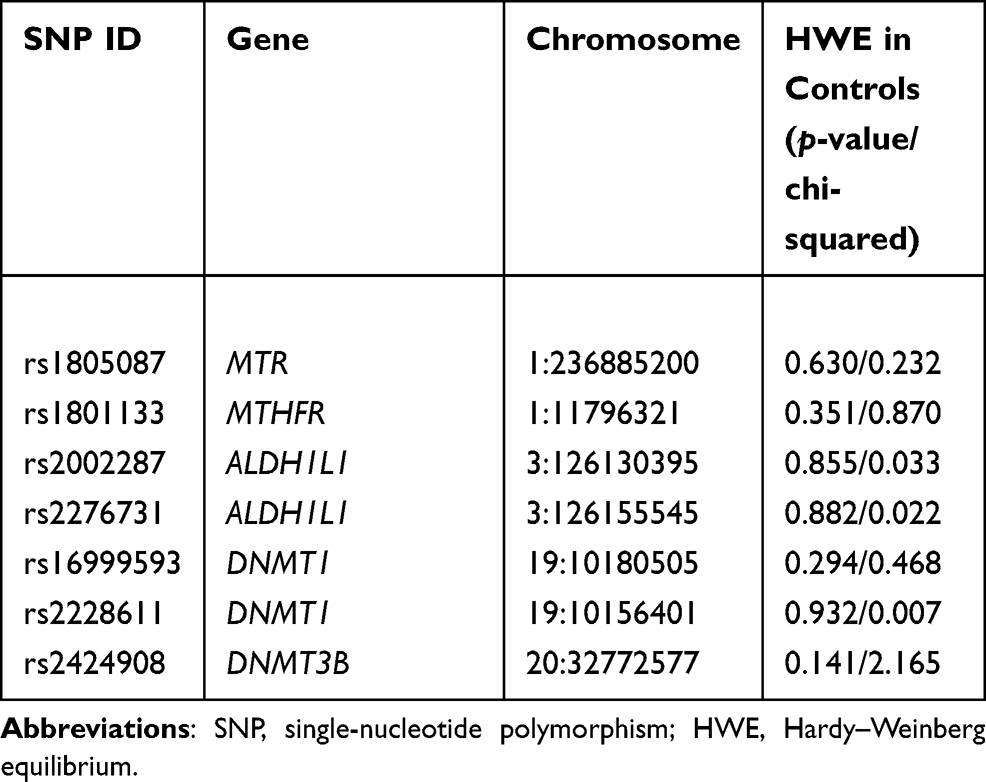 |
Table 1 Information on the Included Genetic Variations |
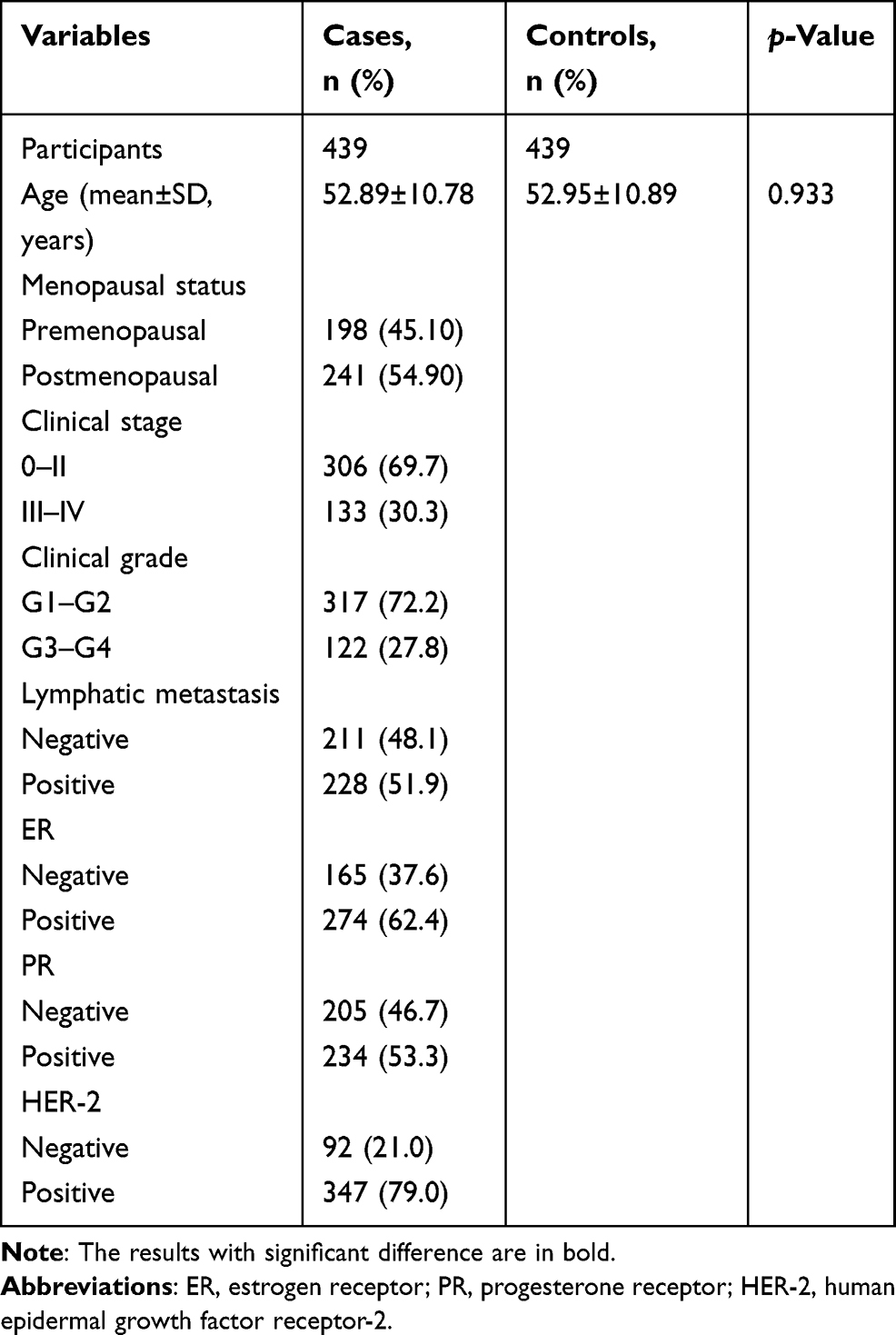 |
Table 2 Clinical Characteristics of the Enrolled Participants |
Logistic regression revealed that DNMT1 rs16999593 and rs2228611 were associated with BC risk. DNMT1 rs16999593 TC genotype (TC vs TT: adjusted OR=1.41, 95% CI: 1.05–1.90, p=0.023) and TC/CC genotype (TC/CC vs TT: adjusted OR=1.38, 95% CI: 1.03–1.84, p=0.030) increased the risk of BC. However, DNMT1 rs2228611 A allele was found to be a protective factor for BC (AA vs GG: adjusted OR=0.55, 95% CI: 0.33–0.94, p=0.030; GA/AA vs GG: adjusted OR=0.74, 95% CI: 0.56–0.97, p=0.030; additive model: adjusted OR=0.76, 95% CI: 0.61–0.94, p=0.012). No other SNPs showed any significant correlation with the risk of BC, as demonstrated in Table 3.
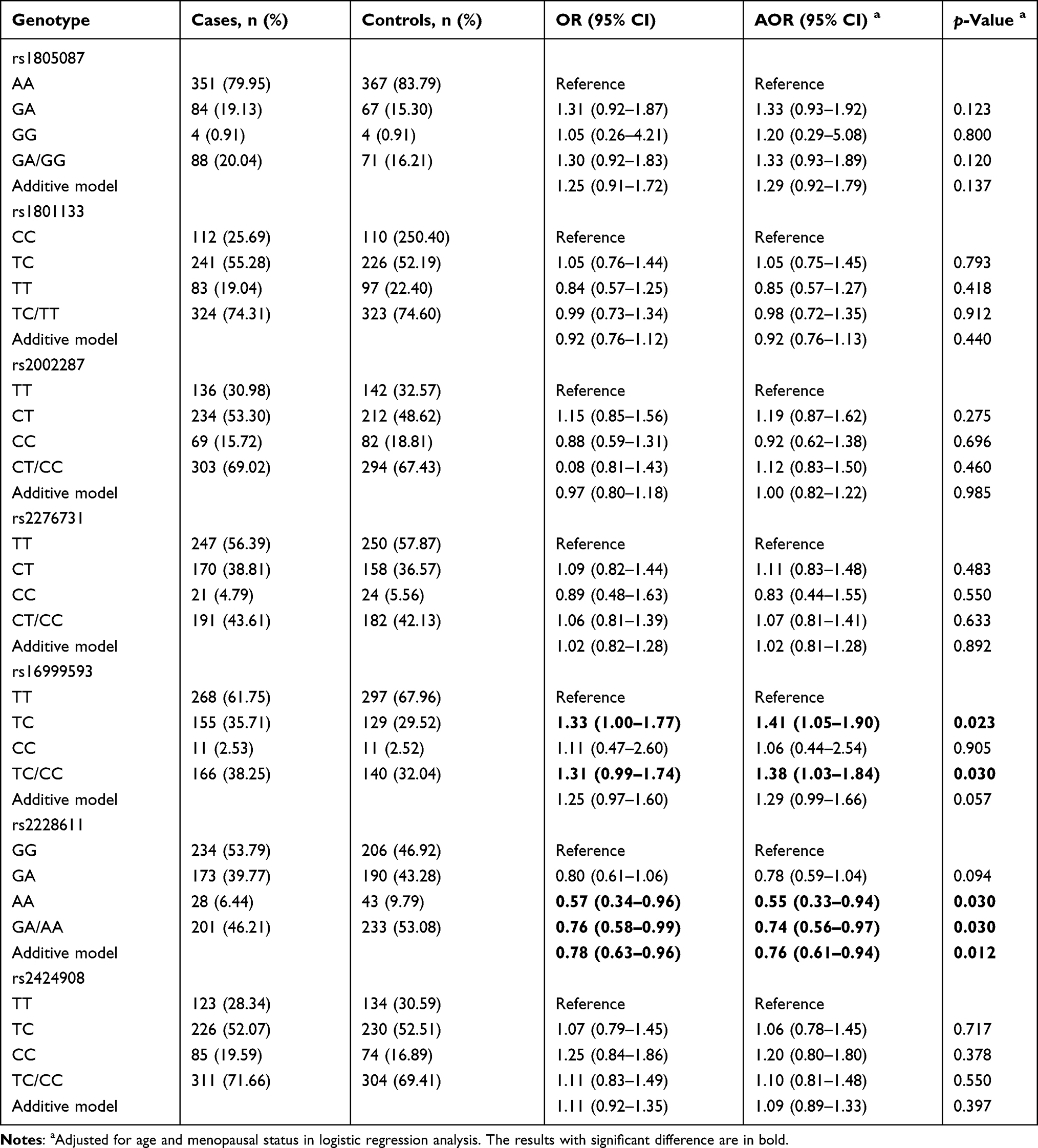 |
Table 3 Association Between Polymorphisms and Risk of Breast Cancer |
In order to explore the impact of menopause on the risk, we stratified the subjects according to menopausal status, and the results showed DNMT1 rs2228611 was associated with a reduced risk of BC in premenopausal females (GA vs AA: adjusted OR=0.62, 95% CI: 0.42–0.90, p=0.012), as shown in Table 4.
 |
Table 4 Stratified Effects of Polymorphisms on Breast Cancer Risk by Menopausal Status |
In addition, a subgroup analysis was performed based on clinical pathological characteristics, and logistic regression analysis showed that DNMT1 rs16999593 TC/CC had a higher BC risk than wild-type TT in the subgroups with TNM stage 0–II (TC/CC vs TT: adjusted OR=1.51, 95% CI: 1.10–2.07, p=0.010) and negative lymph-node metastasis (TC/CC vs TT: adjusted OR=1.46, 95% CI: 1.04–2.07, p=0.031). In contrast, DNMT1 rs2228611 GA/AA was related to a reduced risk of BC in the subgroups of low-rank TNM stage 0–II (GA/AA vs GG: adjusted OR=0.72, 95% CI: 0.52–0.97, p=0.033) and advanced tumor grade (G3–G4) (GA/AA vs GG: adjusted OR=0.55, 95% CI: 0.36–0.84, p=0.005), as demonstrated in Table 5.
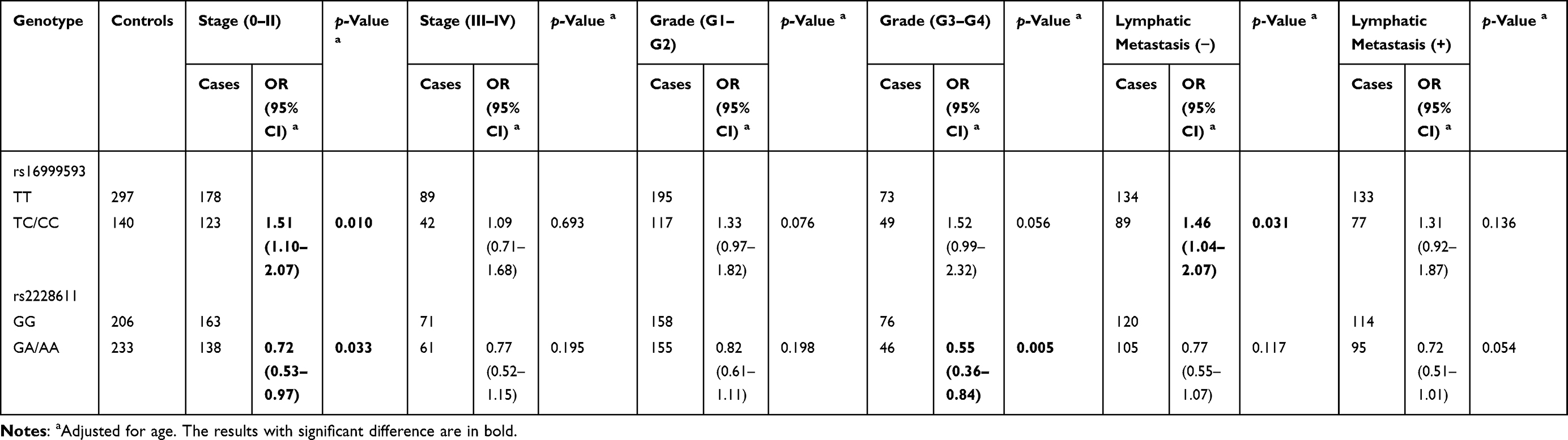 |
Table 5 Stratified Effects of SNPs on Breast Cancer Risk by the Pathological Characteristics of Patients |
Moreover, we performed subgroup analysis according to the expression of ER, PR and HER-2, and found that DNMT1 rs16999593 C allele carriers had an increased risk of HER-2− BC (TC/CC vs TT: adjusted OR=1.40, 95% CI: 1.03–1.90, p=0.031). However, rs2228611 A allele carriers had a decreased risk of ER− BC (GA/AA vs GG: adjusted OR=0.71, 95% CI: 0.52–0.98, p=0.035) and PR− BC (GA/AA vs GG: adjusted OR=0.68, 95% CI: 0.49–0.95, p=0.024). Furthermore, we found that ALDH1L1 rs2002287 C allele carriers had an increased risk of PR+ BC (CT/CC vs TT: adjusted OR=1.54, 95% CI: 1.04–2.26, p=0.030), as shown in Table 6.
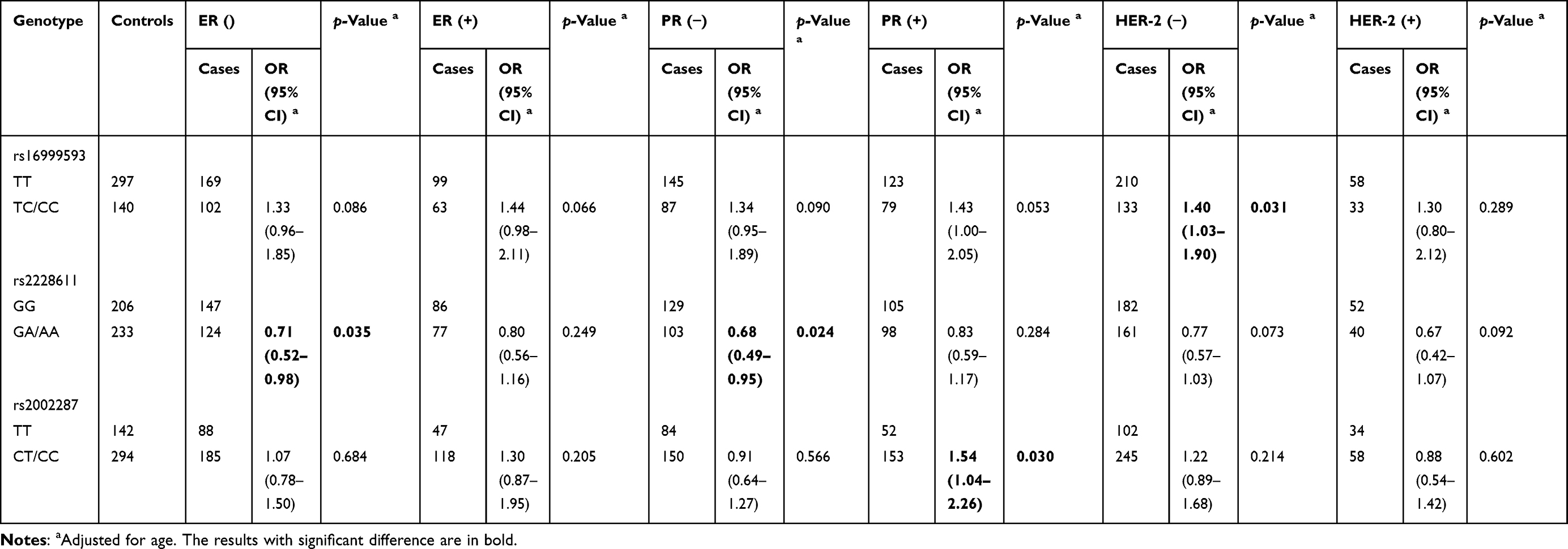 |
Table 6 Stratified Effects of SNPs on Breast Cancer Risk by the Expression of ER, PR and HER-2 |
Discussion
To evaluate the relationship between SNPs in the genes (MTR rs1805087; MTHFR rs1801133; ALDH1L1 rs2002287, rs2276731; DNMT1 rs16999593, rs2228611; DNMT3B rs2424908) of the one-carbon metabolic pathway and BC risk, we recruited 439 pairs of BC patients and healthy controls, and found that DNMT1 rs16999593 and rs2228611 were associated with BC risk.
DNA methylation is closely related to the pathological process of BC,29,30 and methylation genes are potential predictors of survival and prognosis in patients with metastatic BC,31 indicating their clinical application value in risk stratification and disease monitoring. DNMT1 encodes DNA methyltransferase-1, which maintains CpG methylation and participates in embryonic development and somatic cell survival. Knockout of the DNMT1 gene can affect genomic stability and cell viability.32 Our results showed that DNMT1 rs16999593 was associated with an increased risk of BC, which was consistent with the results of Tao et al, who found that the C allele of rs16999593 can increase the risk of triple-negative breast cancer (TNBC), and the rs16999593 TC genotype is a risk factor for TNBC.33 Consistent with this, in another Chinese population, rs16999593 was reported to be highly correlated with the risk of BC.26 Also, a meta-analysis revealed that rs16999593 was associated with gastric cancer risk.34,35 Moreover, we found that DNMT1 rs16999593 was more common in patients with TNM stage 0–II, no lymph-node metastasis and negative expression of HER2, which indicates that DNMT1 could regulate gene expression in an epigenetic manner, resulting in the pathological characteristics of cancers.36–38 Functionally, rs16999593, a missense variant, changes from T to C in amino acid 97 (histidine to arginine), which may disrupt the function of DNMT1, thereby increasing the susceptibility to cancer;35 however, the function of substitution caused by the polymorphism is still unclear. In addition, our results showed that DNMT1 rs2228611 was a protective risk factor for BC patients. A previous study reported that rs2228611 was an independent prognostic factor for gastric cancer.39 In this study, we observed that DNMT1 rs2228611 was associated with BC risk in subtypes of menopausal status (premenopausal women) and pathological characteristics (TNM stage 0–II, grade III–IV, and negative expression of ER and PR), indicating that DNMT1 rs2228611 may contribute to BC through various pathways.40–42 Functionally, DNMT1 rs2228611, a SNP at the exon splicing enhancer region, is a synonymous mutation (proline to proline) that may alter the pre-RNA splicing of DNMT1.39,43
ALDH1L1 is the key enzymes in the metabolic pathway of folic acid, and the intake of folic acid is positively correlated with DNA methylation.18 ALDH1L1 gene expression is down-regulated in cancer, and low ALDH1L1 expression is associated with the occurrence and malignant expression of tumors.44–46 In this study, we observed that ALDH1L1 rs2002287 was not associated with the overall risk of BC, while, in the subgroup analysis, we observed that PR-positive women who carried the C allele have a higher risk of BC, which is consistent with a previous report.47 However, a conflicting conclusion was reported that rs2002287 was associated with a decreased risk of BC,44 which may be attributed to the fact that the effect of SNP on cancer has racial specificity.48 At the same time, this is the first study on the relationship between rs2002287 and BC risk in Chinese people, so large sample sized case-control studies are needed.
This study investigated seven polymorphisms in the one-carbon metabolism pathway and the risk of BC, with the aim of providing a basis for establishing a BC susceptibility gene database in the Chinese population. Our study provided molecular epidemiological data for the basic and clinical study of BC.
Admittedly, this study has some limitations. First of all, the SNPs of many key enzymes in the process of one-carbon metabolism have not been studied. We need to further explore and find other SNPs. Secondly, alcohol is a risk factor for BC, and alcohol can hinder the metabolism of folic acid and change the methylation of genes.47 Other studies have shown that alcohol and folic acid can also affect one-carbon metabolism and disrupt DNA methylation, leading to cancer.18 Therefore, we should take alcohol and dietary factors into account. Thirdly, our research population is only from Nanjing First Hospital in Nanjing, a single center with a small population . In further study, we should increase the sample size as much as possible to expand the scope of the study population.
Conclusion
DNMT1 rs16999593 and rs2228611 were potentially associated with BC risk.
Funding
This study was supported by The National Nature Science Foundation of China (no. 82073288), Jiangsu Youth Medical Talents Training Project to BH (QNRC2016066), and the Innovation Team of Jiangsu Provincial Health-Strengthening Engineering by Science and Education (CXTDB2017008).
Disclosure
The authors report no conflicts of interest for this study.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.21660
2. Sun YS, Zhao Z, Yang ZN, et al. Risk factors and preventions of breast cancer. Int J Biol Sci. 2017;13(11):1387–1397. doi:10.7150/ijbs.21635
3. Yan T, Shen C, Jiang P, et al. Risk SNP-induced lncRNA-SLCC1 drives colorectal cancer through activating glycolysis signaling. Signal Transduct Target Ther. 2021;6(1):70. doi:10.1038/s41392-020-00446-7
4. Hua JT, Ahmed M, Guo H, et al. Risk SNP-mediated promoter-enhancer switching drives prostate cancer through lncRNA PCAT19. Cell. 2018;174(3):564–575 e518. doi:10.1016/j.cell.2018.06.014
5. Cuzick J, Brentnall AR, Segal C, et al. Impact of a panel of 88 single nucleotide polymorphisms on the risk of breast cancer in high-risk women: results from two randomized tamoxifen prevention trials. J Clin Oncol. 2017;35(7):743–750. doi:10.1200/JCO.2016.69.8944
6. Atkinson SR, Way MJ, McQuillin A, et al. Reply to: “The PNPLA3 SNP rs738409: gallele is associated with increased liver disease-associated mortality but reduced overall mortality in a population-based cohort”. J Hepatol. 2018;68(4):860–862. doi:10.1016/j.jhep.2017.12.005
7. Ducker GS, Rabinowitz JD. One-carbon metabolism in health and disease. Cell Metab. 2017;25(1):27–42. doi:10.1016/j.cmet.2016.08.009
8. Cao S, Zhu Z, Zhou J, et al. Associations of one-carbon metabolism-related gene polymorphisms with breast cancer risk are modulated by diet, being higher when adherence to the Mediterranean dietary pattern is low. Breast Cancer Res Treat. 2021;187:793–804. doi:10.1007/s10549-021-06108-8
9. De Carvalho DD, Sharma S, You JS, et al. DNA methylation screening identifies driver epigenetic events of cancer cell survival. Cancer Cell. 2012;21(5):655–667. doi:10.1016/j.ccr.2012.03.045
10. Yang M, Vousden KH. Serine and one-carbon metabolism in cancer. Nat Rev Cancer. 2016;16(10):650–662. doi:10.1038/nrc.2016.81
11. Brasky TM, White E, Chen C-L. Long-Term, Supplemental, One-CarbonMetabolism–Related Vitamin B Use in Relation to Lung Cancer Risk in the Vitamins and Lifestyle (VITAL) Cohort. J Clin Oncol. 2017;36(30). doi:10.1200/JCO
12. Reina-Campos M, Linares JF, Duran A, et al. Increased Serine and One-Carbon Pathway Metabolism by PKClambda/iota Deficiency Promotes Neuroendocrine Prostate Cancer. Cancer Cell. 2019;35(3):385–400 e389. doi:10.1016/j.ccell.2019.01.018
13. Golden E, Rashwan R, Woodward EA, et al. The oncogene AAMDC links PI3K-AKT-mTOR signaling with metabolic reprograming in estrogen receptor-positive breast cancer. Nat Commun. 2021;12(1):1920. doi:10.1038/s41467-021-22101-7
14. Lu J, Trabert B, Liao LM, et al. Dietary intake of nutrients involved in folate-mediated one-carbon metabolism and risk for endometrial cancer. Int J Epidemiol. 2019;48(2):474–488. doi:10.1093/ije/dyy270
15. Zeng J, Gu Y, Fu H, et al. Association between one-carbon metabolism-related vitamins and risk of breast cancer: a systematic review and meta-analysis of prospective studies. Clin Breast Cancer. 2020;20(4):e469–e480. doi:10.1016/j.clbc.2020.02.012
16. Antwi SO, Petrick JL, Campbell PT, et al. One-carbon metabolism-related micronutrients intake and risk for hepatocellular carcinoma: a prospective cohort study. Int J Cancer. 2020;147(8):2075–2090. doi:10.1002/ijc.33007
17. Brasky TM, Ray RM, Navarro SL, et al. Supplemental one-carbon metabolism related B vitamins and lung cancer risk in the Women’s Health Initiative. Int J Cancer. 2020;147(5):1374–1384. doi:10.1002/ijc.32913
18. Song MA, Brasky TM, Marian C, et al. Genetic variation in one-carbon metabolism in relation to genome-wide DNA methylation in breast tissue from heathy women. Carcinogenesis. 2016;37(5):471–480. doi:10.1093/carcin/bgw030
19. Coppede F, Stoccoro A, Tannorella P, et al. Association of Polymorphisms in Genes Involved in One-Carbon Metabolism with MTHFR Methylation Levels. Int J Mol Sci. 2019;20:15. doi:10.3390/ijms20153754
20. Kim W, Woo HD, Lee J, et al. Dietary folate, one-carbon metabolism-related genes, and gastric cancer risk in Korea. Mol Nutr Food Res. 2016;60(2):337–345. doi:10.1002/mnfr.201500384
21. Shao HB, Ren KW, Gao SL, et al. Human methionine synthase A2756G polymorphism increases susceptibility to prostate cancer. AGING. 2018;10:1776–1788. doi:10.18632/aging.101509
22. Zhang S, Jiang J, Tang W, et al. Methylenetetrahydrofolate reductase C677T (Ala>Val, rs1801133 C>T) polymorphism decreases the susceptibility of hepatocellular carcinoma: a meta-analysis involving 12,628 subjects. Biosci Rep. 2020;40(2). doi:10.1042/BSR20194229
23. Yi K, Yang L, Lan Z, et al. The association between MTHFR polymorphisms and cervical cancer risk: a system review and meta analysis. Arch Gynecol Obstet. 2016;294(3):579–588. doi:10.1007/s00404-016-4037-6
24. Zhu G, Liao X, Han C, et al. ALDH1L1 variant rs2276724 and mRNA expression predict post-operative clinical outcomes and are associated with TP53 expression in HBV-related hepatocellular carcinoma. Oncol Rep. 2017;38(3):1451–1463. doi:10.3892/or.2017.5822
25. Wang X, Wang C, Guan J, et al. Progress of Breast Cancer basic research in China. Int J Biol Sci. 2021;17(8):2069–2079. doi:10.7150/ijbs.60631
26. Sun MY, Yang XX, Xu WW, et al. Association of DNMT1 and DNMT3B polymorphisms with breast cancer risk in Han Chinese women from South China. Genet Mol Res. 2012;11(4):4330–4341. doi:10.4238/2012.September.26.1
27. Xu T, Hu XX, Liu XX, et al. Association between SNPs in Long Non-coding RNAs and the Risk of Female Breast Cancer in a Chinese Population. J Cancer. 2017;8(7):1162–1169. doi:10.7150/jca.18055
28. He B, Pan B, Pan Y, et al. IL-4IL-4R and IL-6IL-6R genetic variations and gastric cancer risk in the Chinese population. Am J Transl Res. 2019;11(6):3698–3706.
29. Shukla V, Coumoul X, Lahusen T, et al. BRCA1 affects global DNA methylation through regulation of DNMT1. Cell Res. 2010;20(11):1201–1215. doi:10.1038/cr.2010.128
30. Koo GB, Morgan MJ, Lee DG, et al. Methylation-dependent loss of RIP3 expression in cancer represses programmed necrosis in response to chemotherapeutics. Cell Res. 2015;25(6):707–725. doi:10.1038/cr.2015.56
31. Visvanathan K, Fackler MS, Zhang Z, et al. Monitoring of Serum DNA Methylation as an Early Independent Marker of Response and Survival in Metastatic Breast Cancer: TBCRC 005 Prospective Biomarker Study. J Clin Oncol. 2017;35(7):751–758. doi:10.1200/JCO.2015.66.2080
32. Spada F, Haemmer A, Kuch D, et al. DNMT1 knockout delivers a strong blow to genome stability and cell viability. J Cell Biol. 2007;176(5):565–571. doi:10.1083/jcb.200610062
33. Tao R, Chen Z, Wu P, et al. The possible role of EZH2 and DNMT1 polymorphisms in sporadic triple-negative breast carcinoma in southern Chinese females. Tumour Biol. 2015;36(12):9849–9855. doi:10.1007/s13277-015-3754-y
34. Li H, Li W, Liu S, et al. DNMT1, DNMT3A and DNMT3B Polymorphisms Associated With Gastric Cancer Risk: a Systematic Review and Meta-analysis. EBioMedicine. 2016;13:125–131. doi:10.1016/j.ebiom.2016.10.028
35. Tian J, Liu G, Zuo C, et al. Genetic polymorphisms and gastric cancer risk: a comprehensive review synopsis from meta-analysis and genome-wide association studies. Cancer Biol Med. 2019;16(2):361–389. doi:10.20892/j.issn.2095-3941.2018.0290
36. Tang H, Liu P, Yang L, et al. miR-185 suppresses tumor proliferation by directly targeting E2F6 and DNMT1 and indirectly upregulating BRCA1 in triple-negative breast cancer. Mol Cancer Ther. 2014;13(12):3185–3197. doi:10.1158/1535-7163.MCT-14-0243
37. She S, Zhao Y, Kang B, et al. Combined inhibition of JAK1/2 and DNMT1 by newly identified small-molecule compounds synergistically suppresses the survival and proliferation of cervical cancer cells. Cell Death Dis. 2020;11(9):724. doi:10.1038/s41419-020-02934-8
38. Yu J, Zayas J, Qin B, et al. Targeting DNA methylation for treating triple-negative breast cancer. Pharmacogenomics. 2019;20(16):1151–1157. doi:10.2217/pgs-2019-0078
39. Jia Z, Wu X, Cao D, et al. Polymorphisms of the DNA Methyltransferase 1 Gene Predict Survival of Gastric Cancer Patients Receiving Tumorectomy. Dis Markers. 2016;2016:8578064. doi:10.1155/2016/8578064
40. Azizi M, Fard-Esfahani P, Mahmoodzadeh H, et al. MiR-377 reverses cancerous phenotypes of pancreatic cells via suppressing DNMT1 and demethylating tumor suppressor genes. Epigenomics. 2017;8(8):1–7.
41. Jin X, Li Y, Guo Y, et al. ERalpha is required for suppressing OCT4-induced proliferation of breast cancer cells via DNMT1/ISL1/ERK axis. Cell Prolif. 2019;52(4):e12612. doi:10.1111/cpr.12612
42. Hu Y, Wu F, Liu Y, et al. DNMT1 recruited by EZH2-mediated silencing of miR-484 contributes to the malignancy of cervical cancer cells through MMP14 and HNF1A. Clin Epigenetics. 2019;11(1):186. doi:10.1186/s13148-019-0786-y
43. Maric H, Supic G, Kandolf-Sekulovic L, et al. DNMT1 and DNMT3B genetic polymorphisms affect the clinical course and outcome of melanoma patients. Melanoma Res. 2019;29(6):596–602. doi:10.1097/CMR.0000000000000612
44. Krupenko SA, Horita DA. The Role of Single-Nucleotide Polymorphisms in the Function of Candidate Tumor Suppressor ALDH1L1. Front Genet. 2019;10:1013. doi:10.3389/fgene.2019.01013
45. Krupenko SA, Krupenko NI. Loss of ALDH1L1 folate enzyme confers a selective metabolic advantage for tumor progression. Chem Biol Interact. 2019;302:149–155. doi:10.1016/j.cbi.2019.02.013
46. Krupenko SA, Krupenko NI. ALDH1L1 and ALDH1L2 Folate Regulatory Enzymes in Cancer. Adv Exp Med Biol. 2018;1032:127–143. doi:10.1007/978-3-319-98788-0_10
47. Llanos AA, Dumitrescu RG, Brasky TM, et al. Relationships among folate, alcohol consumption, gene variants in one-carbon metabolism and p16INK4a methylation and expression in healthy breast tissues. Carcinogenesis. 2015;36(1):60–67. doi:10.1093/carcin/bgu219
48. Mo W, Ding Y, Zheng Y, et al. Associations between folate metabolism enzyme polymorphisms and breast cancer: a meta-analysis. Breast J. 2020;26(3):484–487. doi:10.1111/tbj.13527
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.