Back to Journals » OncoTargets and Therapy » Volume 9
Association between APE1 Asp148Glu polymorphism and the risk of urinary cancers: a meta-analysis of 18 case–control studies
Authors Zhong JH, Zhao Z, Liu J, Yu H, Zhou J, Shi R
Received 28 November 2015
Accepted for publication 2 February 2016
Published 15 March 2016 Volume 2016:9 Pages 1499—1510
DOI https://doi.org/10.2147/OTT.S101456
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Min Li
Jie-Hui Zhong,1,2,* Zhen Zhao,3,* Jie Liu,4 Hai-Lang Yu,2 Jue-Yu Zhou,2 Rong Shi2
1Department of Clinical Medicine, The First Clinical Medical College, Southern Medical University, 2Institute of Genetic Engineering, School of Basic Medical Sciences, Southern Medical University, 3Department of Urinary Surgery, 4Department of Obstetrics and Gynecology, Nanfang Hospital, Southern Medical University, Guangzhou, People’s Republic of China
*These authors contributed equally to this work
Background: Several observational studies suggested that APE1 Asp148Glu was significantly associated with urinary cancers; however, the results of published studies are inconsistent.
Materials and methods: The PubMed and EMBASE were searched for case–control studies regarding the association between Asp148Glu and the risk of urinary cancers with a time limit of September 12, 2015. Pooled odds ratios (ORs) and 95% confidence intervals (CIs) were used to assess the strength of the association between Asp148Glu and the risk of developing prostate cancer, kidney cancer, bladder cancer, as well as all urinary cancers combined.
Results: A total of 18 case–control studies were included in the analysis. Our meta-analysis revealed that the inheritance of at least one APE1 148Glu among Asian men was associated with a 1.26-fold increase in the risk of developing urinary cancers. Meanwhile, APE1 Asp148Glu was significantly associated with the risk of prostate cancer. However, there were no significant relationships between the APE1 SNP (single nucleotide polymorphism) and all urinary cancers combined and bladder cancer and kidney cancer among the men of Caucasian/Asian/African descent or all racial/ethnic groups combined. When stratified by the quality score, no significant association was found in high-quality studies (score ≥7), but a significant increased risk of urinary cancers was observed in lower quality studies (score <7) (dominant model: OR=1.27, 95% CI=1.11–1.45).
Conclusion: Our meta-analysis suggests that APE1 Asp148Glu was not associated with the risk of urinary cancers but might increase the risk of urinary cancers among Asians. Stratification by cancer type identified a significant association of Asp148Glu with prostate cancer.
Keywords: APE1, polymorphism, cancer, meta-analysis
Introduction
Urinary cancers, including kidney cancer, prostate cancer and bladder cancer, are common types of malignancies worldwide.1 For example, prostate cancer is the second most frequently diagnosed cancer and the sixth leading cause of cancer death in males, accounting for 14% of the total new cancer cases and 6% of the total cancer deaths in males in 2008.1 Bladder cancer is the seventh most common cancer among men, with ~297,300 new incident cases per year in the world.1 The estimated probability of developing urinary cancers is based on the average experience of the general population and may over- or underestimate individual risk because of differences in exposure, medical history, and/or genetic susceptibility.2
DNA repair genes play a major role in the DNA mismatch repair pathway, which includes base excision repair (BER), nucleotide excision repair, mismatch repair, and double strand break repair.3 Genetic variations in genes involved in DNA repair would confer susceptibility to the tumor and would be associated to disease aggressiveness through the alteration of DNA repair pathways.4 Among them, BER pathway is responsible for repairing small lesions such as oxidative damage, alkylation, or methylation.5 This pathway is a multistep process that requires the activity of several proteins.6 Apurinic/apyrimidinic endonuclease 1 (APE1, also known as APE, APEX, HAP1, and REF-1) is a multifunctional protein that plays a central role in the BER pathway through hydrolyzing the phosphodiester backbone immediately 5′ to the AP site.5,7 APE1 can act as a 3′-phosphodiesterase to initiate the repair of 3′-blocking damage at DNA single-strand breaks, which are produced either directly by reactive oxygen species or indirectly through the enzymatic removal of damaged bases.8,9 It also acts as a transcriptional coactivator for numerous transcription factors (AP-1, p53, Pax-5, and TTF-1) involved in cancer development.10
The human APE1 gene (2.6 kb in size) is localized to chromosome14 q11.2-12 and consists of four introns and five exons.11,12 A total of 18 polymorphisms in APE1 have been identified.13 But the most extensively studied polymorphism is a T to G transversion, Asp148Glu (rs1130409 and T1349G). It was reported that the Glu was associated with an increased mitotic delay after exposure to ionizing radiation.13,14 Functional studies on APE1 Asp148Glu suggested that the Glu may alter endonuclease and DNA-binding activity, reduce the ability to communicate with other BER proteins, and decrease the capacity to repair DNA oxidative damage.15 In the study on X-ray exposure to lymphocytes and polymorphisms of DNA repair genes on chromosome aberrations, samples from individuals with the Asp/Glu or Glu/Glu genotype showed higher levels of damage with regard to all the studied measures, including aberrant cells, chromatid breaks, chromatid exchanges, deletions, and dicentrics.16 Although numerous epidemiological studies have been conducted to explore the association between APE1 Asp148Glu and the risk of urinary cancers,17–37 the results are to some extent inconsistent, which may be due to the limitations in individual studies. In this study, we combined all the published case–control studies regarding the association between APE1 Asp148Glu and urinary cancers to better explore this genetic variation on the risk of developing urinary cancers.
Materials and methods
Study identification and selection
Case–control studies regarding the APE1 Asp148Glu and the risk of urinary cancers published before September 12, 2015 were included through searches of PubMed and EMBASE by using the following terms and key words: “apurinic endonuclease” or “apyrimidinic endonuclease” or APE1 or APEX or APEX1 or HAP1 or REF-1; polymorphism or variant or variation or mutation; and kidney or renal or urothelial or “transitional cell carcinoma” or bladder or prostatic or prostate. The search was limited to human studies. The criteria used for the study selection were as follows: 1) the articles were concerned about the association between APE1 Asp148Glu and urinary cancers, including prostate cancer, kidney cancer, and bladder cancer; 2) the studies were designed as case–control studies; 3) detailed genotyping data were available; and 4) there were no overlapping data.
Data extraction
Information was carefully extracted from all the eligible studies independently by three investigators according to the selection criteria listed earlier. The following data were collected: first author’s name, publication year, country, ethnicity (categorized as Asians, Caucasians, or the African–Americans), source of controls, genotyping method, numbers of cases and controls, genotype frequency of cases and controls, and the result of Hardy–Weinberg equilibrium (HWE) test. We did not require a minimum number of patients to be included in our meta-analysis.
Quality score assessment
The study quality was assessed by using a quality assessment score developed for genetic association studies by Thakkinstian et al.38 Total scores range from 0 (worst) to 12 (best). The criteria for quality assessment of genetic associations between the APE1 Asp148Glu and urinary cancers are described in Table S1.
Statistical analysis
Data in the control group were used to estimate a pooled allelic prevalence. HWE was tested by chi-squared test (P<0.05 was considered representative of statistical significance). The minor allele frequency was also calculated for the controls. The strength of association between APE1 Asp148Glu and the risk of urinary cancers was measured by odd ratios (ORs) with 95% confidence intervals (CIs). The pooled ORs were calculated for recessive model (Glu/Glu vs Asp/Glu+Asp/Asp), dominant model (Glu/Glu+Asp/Glu vs Asp/Asp), homozygote comparison (Glu/Glu vs Asp/Asp), heterozygote comparison (Asp/Glu vs Asp/Asp) and additive model (Glu vs Asp). Heterogeneity assumption was checked by a chi-square-based Q test, and I2 statistics was calculated to quantify the proportion of the total variation across studies due to heterogeneity.39 A P-value of >0.05 for the Q test indicated a lack of heterogeneity among studies, so that the pooled OR estimate of each study was calculated by the fixed-effects model (the Mantel–Haenszel method).40 Otherwise, the random-effects model (the DerSimonian and Laird method) was used.41 Subgroup analyses were conducted according to ethnicity (ie, Asians, African–Americans, Caucasians), cancer type (ie, all urinary cancers, kidney cancer, prostate cancer, and bladder cancer), genotyping methods (ie, polymerase chain reaction-restriction fragment length polymorphism [PCR-RFLP] and TaqMan), source of control (ie, population based and hospital based), and quality score (score <7 or score ≥7). Sensitivity analyses were performed to test the reliability of results by sequential omission of individual studies.42 Publication bias is the tendency on the parts of investigators, reviewers, and editors to submit or accept manuscripts for publication based on the direction or strength of the study findings.43 An estimate of potential publication bias was carried out by the funnel plot, in which the standard error of log (OR) of each study was plotted against its log (OR). Funnel plot asymmetry was further assessed by the method of Egger’s linear regression test (P<0.05 was considered a significant publication bias).44 Statistical analyses were performed by using STATA statistical software (version 11.0).
Results
Extraction process and study characteristics
The selection process of the studies is presented in Figure 1. In total, 419 potentially relevant articles were identified after the initial search, and 20 of them concentrated on the APE1 Asp148Glu. Also, four additional articles were identified from retrieved articles. Thus, following implementation of our search criteria, we found 24 publications on the association between APE1 Asp148Glu and the risk of urinary cancers.10,17,19–28,30–37,45–48 Among them, eight studies were excluded because of duplicated data or lack of usable data.10,24,26,31,35,45,46,48 Because the two studies by Andrew et al18,25 contain overlapping data and the source of control in one of them25 is mixed, the study by Andrew et al18 was excluded when calculating the pooled OR, while the study by Andrew et al25 was excluded when conducting the subgroup analysis according to the source of controls. Hence, 16 publications including 18 studies (5,539 cases and 7,348 controls) were selected in the meta-analysis. Table 1 lists the studies identified and their main characteristics. Of them, there were ten bladder cancer studies, seven prostate cancer studies, and one kidney cancer study. There were six studies32,33,36,37,47 involving Asians, ten studies with Caucasians,17,19–23,25,27,28,34 and two studies with African–Americans.19,30 The quality of all 18 studies ranged from 3 to 12, with a mean value of 6.9 (standard deviation: 2.18). The distribution of genotypes in the controls of each study was consistent with HWE except for two studies,28,34 which were not included in further pooling (Table 2). The pooled prevalence of the Glu were 0.424 (95% CI=0.393–0.456) in control group, while the pooled prevalence of the Glu among Asians were 0.383 (95% CI=0.311–0.456), Caucasians 0.467 (95% CI=0.454–0.480), and African–Americans 0.354 (95% CI=0.354–0.378).
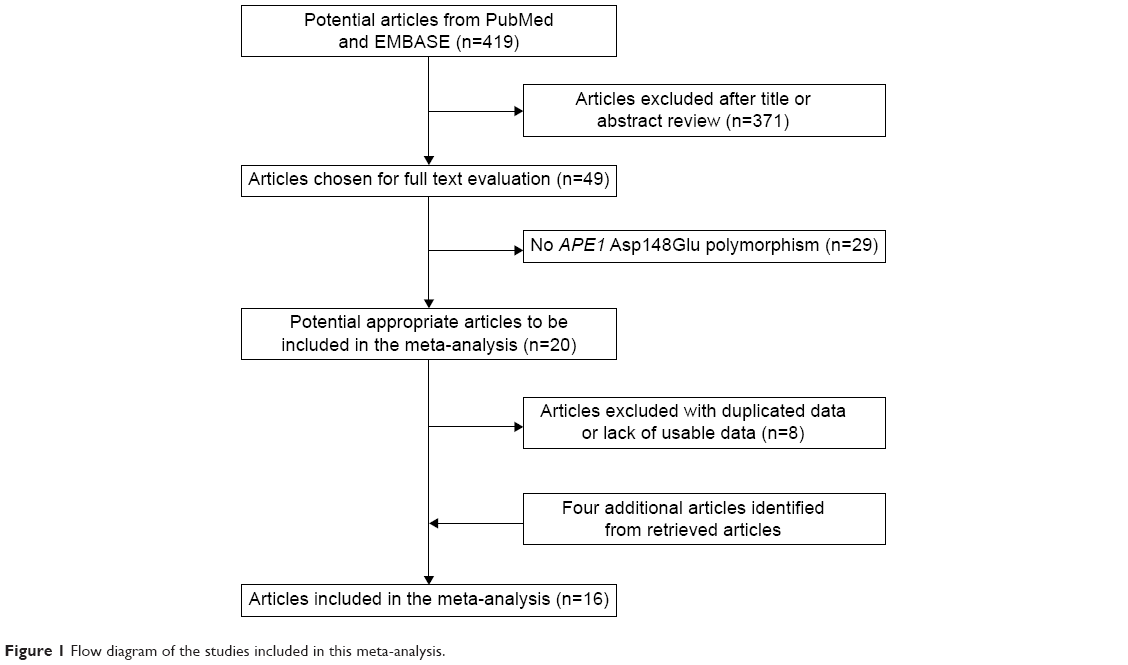 | Figure 1 Flow diagram of the studies included in this meta-analysis. |
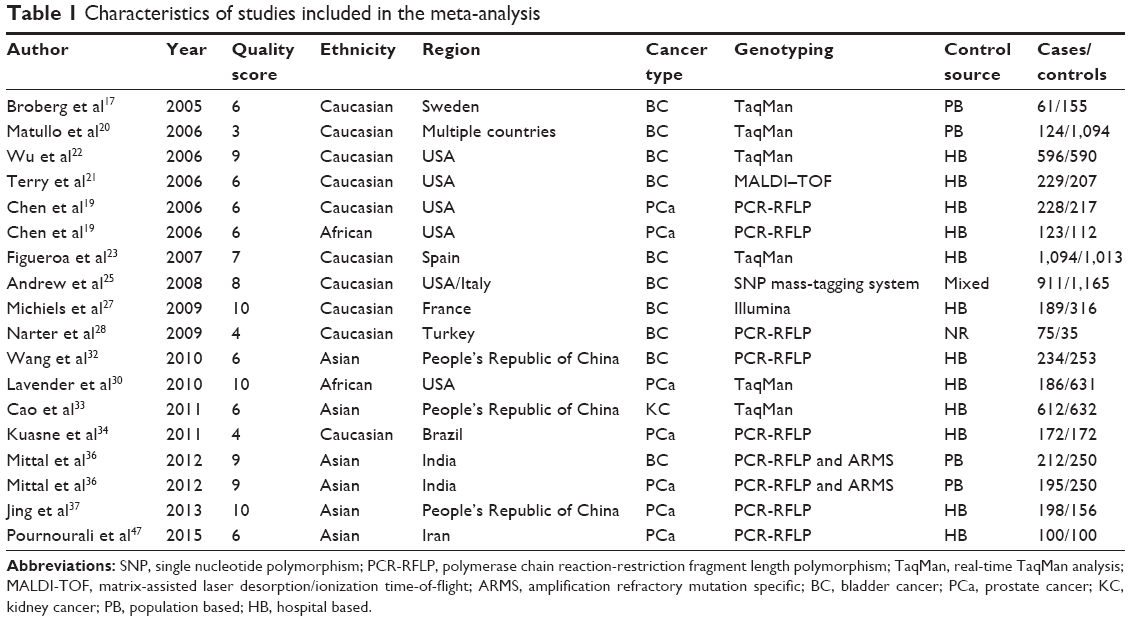 | Table 1 Characteristics of studies included in the meta-analysis |
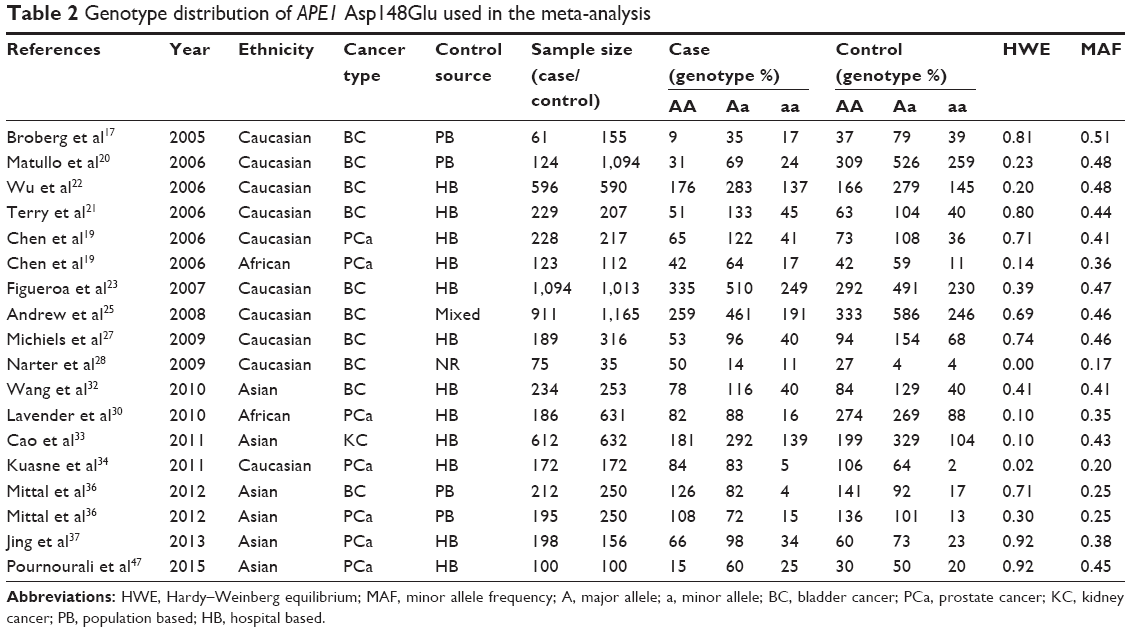 | Table 2 Genotype distribution of APE1 Asp148Glu used in the meta-analysis |
Meta-analysis results
Table 3 lists the main results of the meta-analysis for APE1 Asp148Glu. Overall, no significant association was found between Asp148Glu and the risk of urinary cancers (dominant model: OR=1.06, 95% CI=0.98–1.15; Figure 2A). In the stratified analysis by ethnicity, we did not find significant associations among Caucasians (dominant model: OR=1.06, 95% CI=0.96–1.17; Table 3). Similarly, no significant associations were observed among the African–Americans (dominant model: OR=1.02, 95% CI=0.77–1.35; Table 3). However, among Asians, the individuals who carried the Glu/Glu genotype had an increased risk of urinary cancers (recessive model: OR=1.26, 95% CI=1.03–1.54, P=0.022; Table 3, Figure 2B). When stratified by the source of controls, no significant associations were found in both population-based studies and hospital-based studies (Table 3). When stratified by the quality score, no significant association was found in high-quality studies (score ≥7), but a significant increased risk of urinary cancers was observed in lower quality studies (score <7) (dominant model: OR=1.27, 95% CI=1.11–1.45). Subgroup analyses based on cancer type also showed that there was no significant association of bladder cancer. However, significant association was found between Asp148Glu and the risk of prostate cancer (dominant model: OR=1.21, 95% CI=1.03–1.42, Figure 3A; heterozygote comparison: OR=1.12, 95% CI=1.03–1.44, Figure 3B; additive model: OR=1.12, 95% CI=1.00–1.26). Meanwhile, a significant increased risk of prostate cancer was observed among Caucasians (dominant model: OR=1.45, 95% CI=1.08–1.94; heterozygote comparison: OR=1.44, 95% CI=1.06–1.94; additive model: OR=1.26, 95% CI=1.02–1.56) and Asians (homozygote comparison: OR=1.61, 95% CI=1.05–2.46) but not African–Americans.
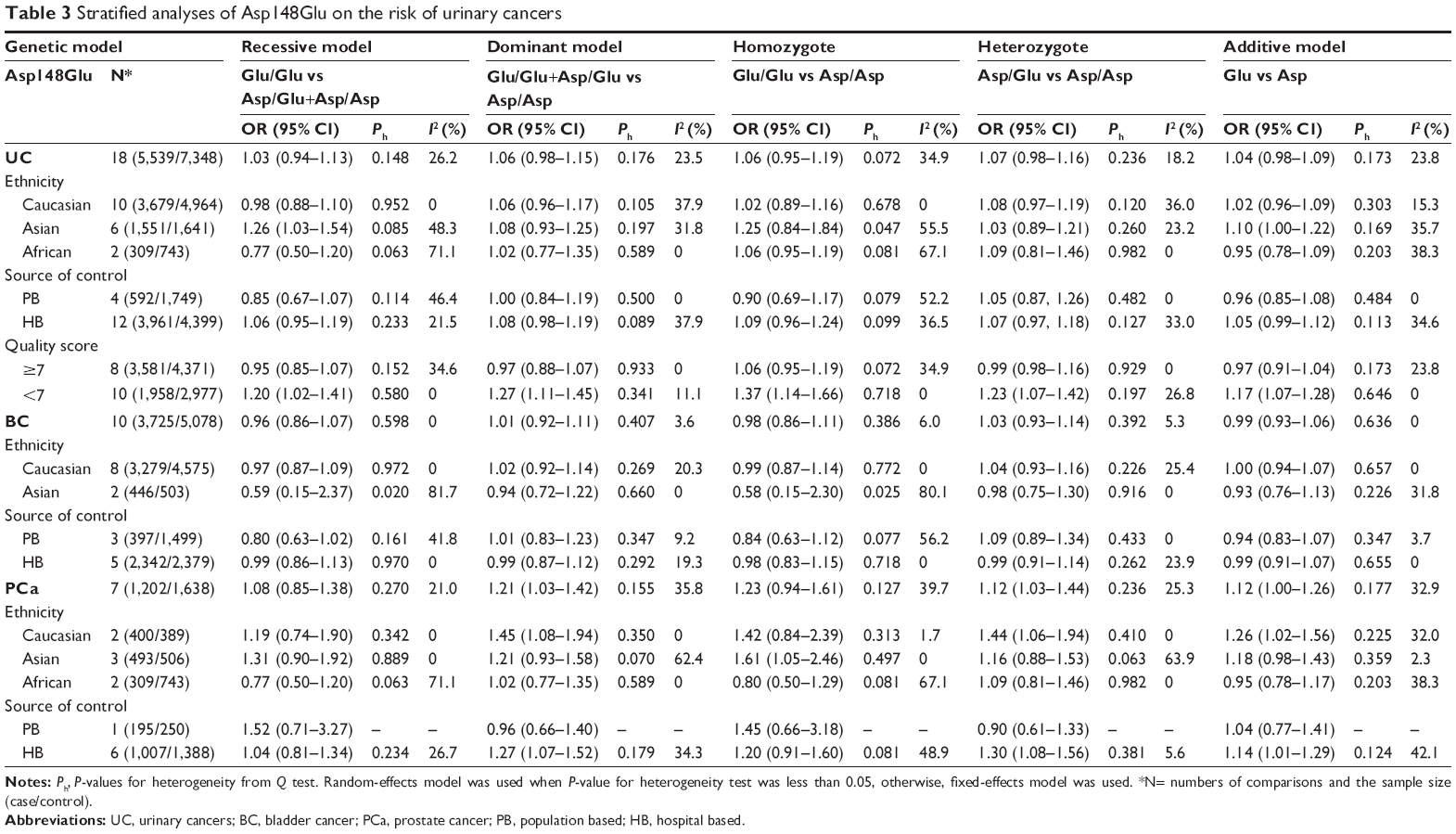 | Table 3 Stratified analyses of Asp148Glu on the risk of urinary cancers |
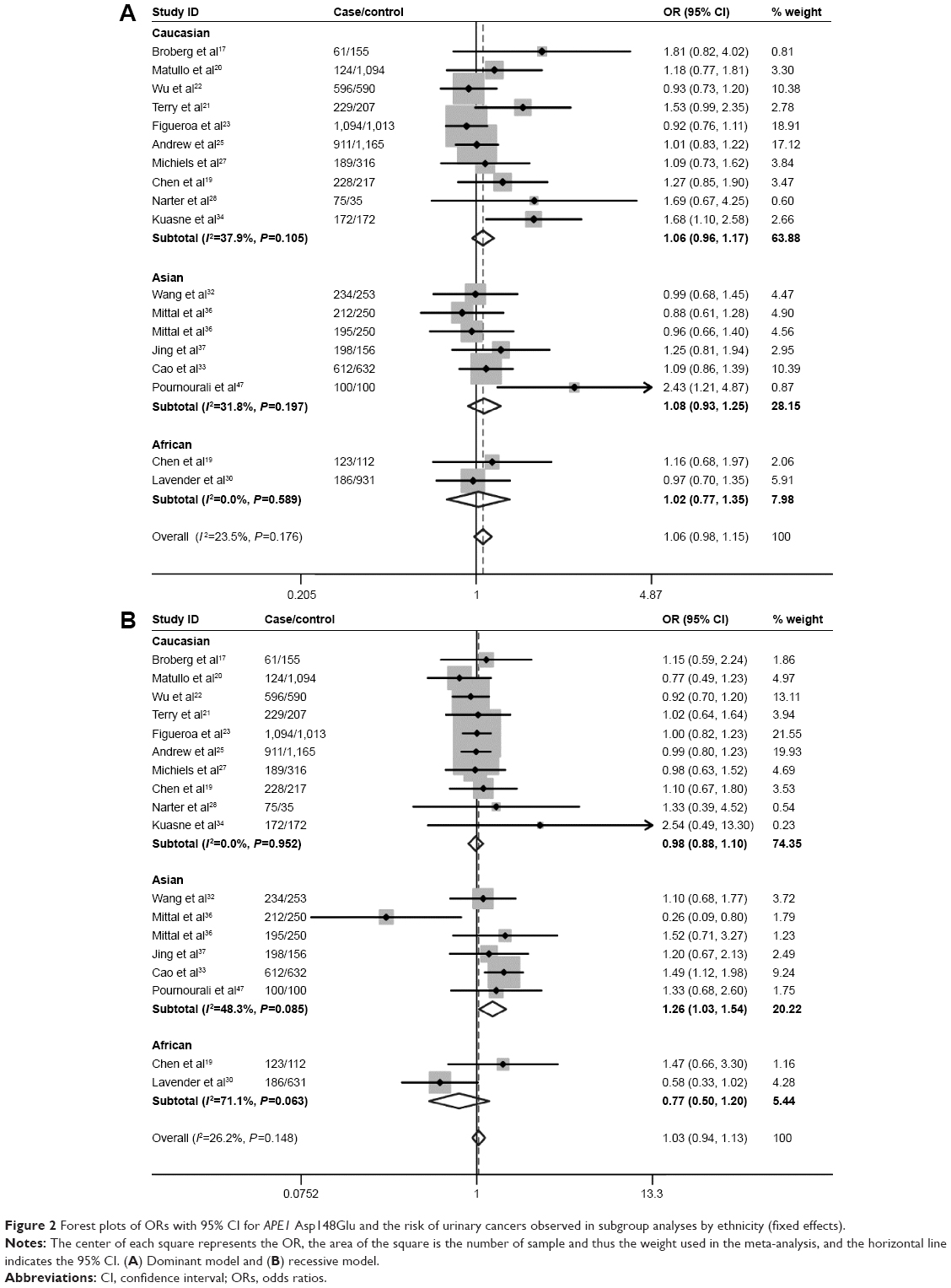 | Figure 2 Forest plots of ORs with 95% CI for APE1 Asp148Glu and the risk of urinary cancers observed in subgroup analyses by ethnicity (fixed effects). |
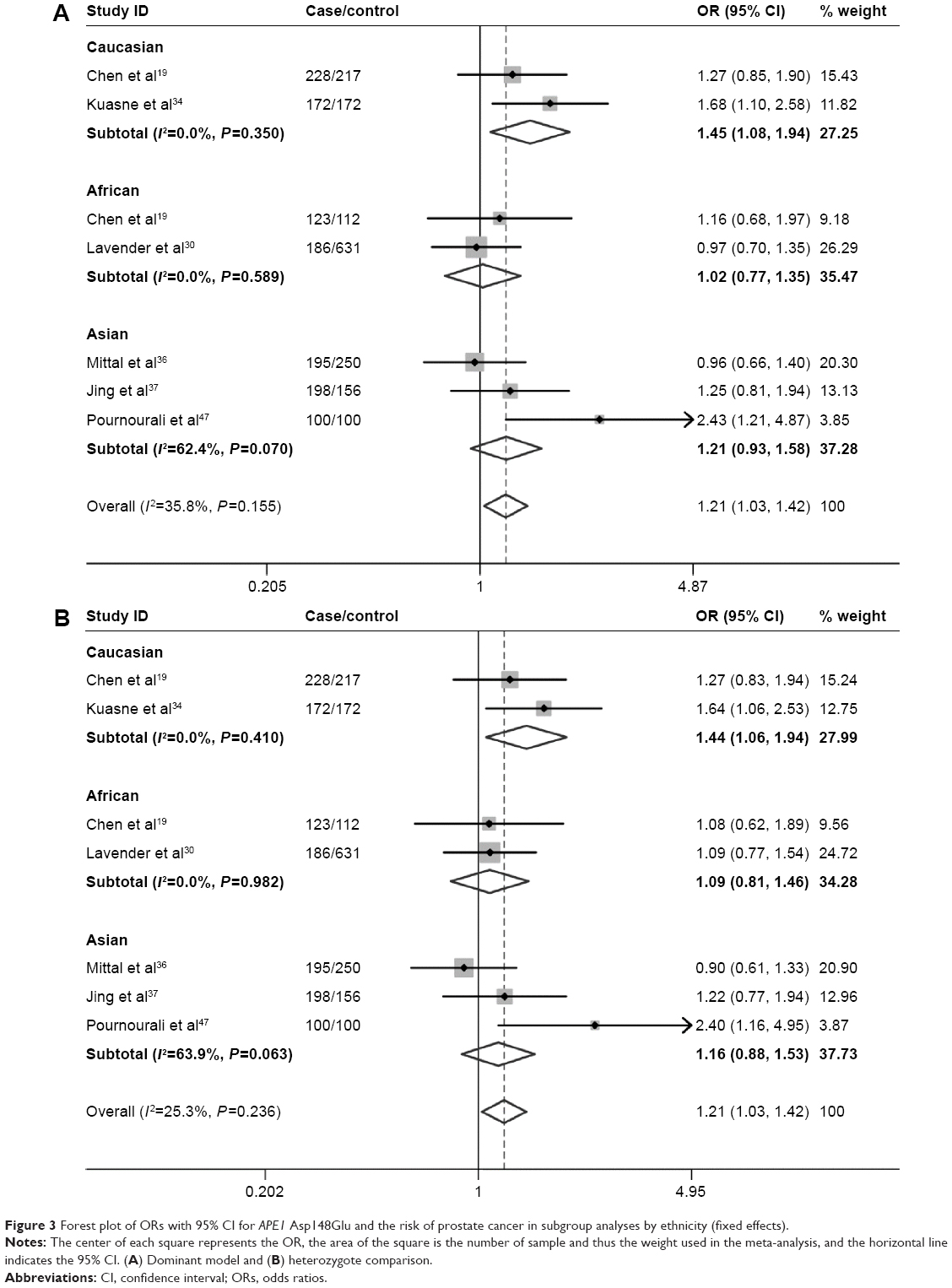 | Figure 3 Forest plot of ORs with 95% CI for APE1 Asp148Glu and the risk of prostate cancer in subgroup analyses by ethnicity (fixed effects). |
Test of heterogeneity and sensitivity analyses
The heterogeneity test showed that there was no significant heterogeneity in overall comparisons (Table 3). Although the genotype distributions in two studies28,34 did not follow the HWE, the corresponding pooled ORs were not materially altered by including or excluding the studies. Additionally, we also assessed the influence of each individual study on the pooled ORs by sequential omission of individual studies. The results showed that the pooled ORs of this polymorphism were altered by omission of the study by Figueroa et al23 (Figure 4).
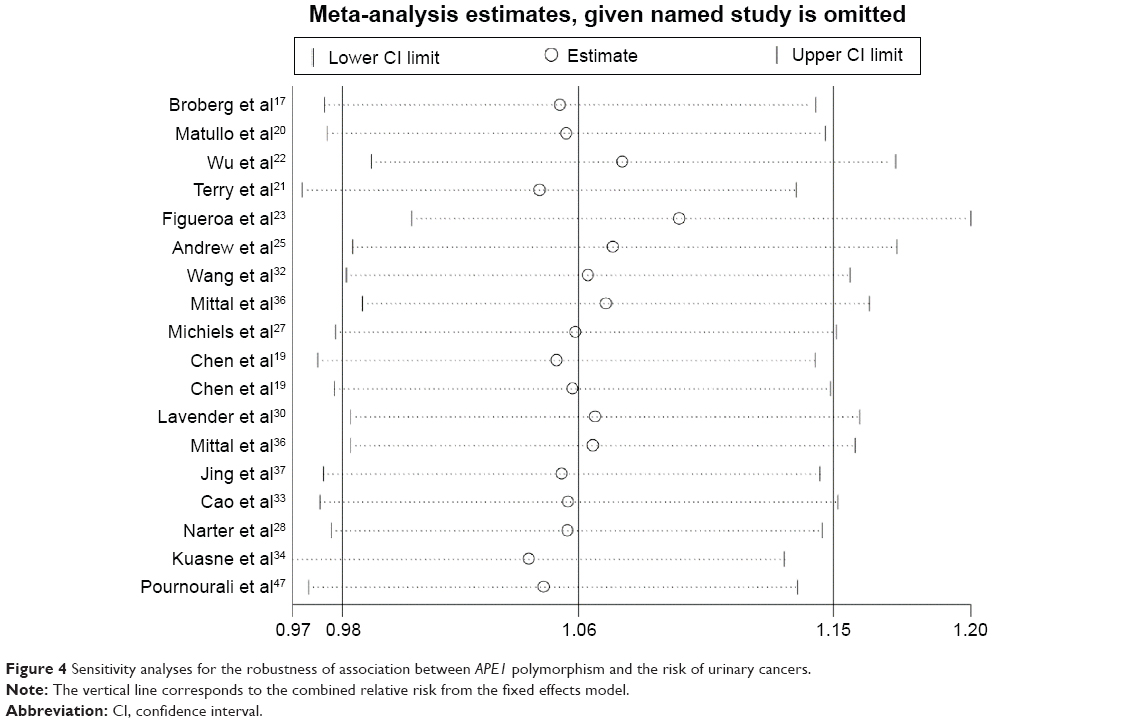 | Figure 4 Sensitivity analyses for the robustness of association between APE1 polymorphism and the risk of urinary cancers. |
Publication bias
Begg’s funnel plot and Egger’s test were conducted to estimate the publication bias of studies. It was showed that Egger’s test was significant for publication bias in heterozygote comparison (P<0.001), additive model (P=0.04), and dominant model (P=0.001). However, the Egger’s test was not statistically significant for publication bias in recessive model (P=0.893) and homozygote comparison (P=0.237). It suggested that a possibility of publication bias could have existed in the studies.
Discussion
Numerous epidemiological studies have been conducted to explore the association between APE1 Asp148Glu and the risk of urinary cancers.17,19–23,25,27,28,30–34,36,37 However, the results are to some extent inconsistent, which may be due to the limitations in individual studies. Meta-analysis has been widely used in epidemiological research, especially for evaluating genetic polymorphisms in cancer susceptibility. It can improve statistical power, subsequently drawing a more reliable conclusion.49 Therefore, we performed a meta-analysis to explore the association between APE1 Asp148Glu and the risk of urinary cancers. Our results indicated that the individuals who carried the Glu/Glu genotype have an increased risk of urinary cancers among Asians. Meanwhile, significant association was found between Asp148Glu and the risk of prostate cancer.
Several genome-wide association studies have identified susceptibility variants,50–52 providing evidence in support of the role of genetic susceptibility in developing urinary cancers. As for prostate cancer, the prostate carcinogenesis is a result of multiple environmental and hereditary risk factors, and genetic factors play important roles in the development of prostate cancer.53 Functional studies on APE1 Asp148Glu suggested that the Glu may alter endonuclease and DNA-binding activity, reduce the ability to communicate with other BER proteins, and decrease the capacity to repair DNA oxidative damage.15 In the study on X-ray exposure to lymphocytes and polymorphisms of DNA repair genes on chromosome aberrations, samples from individuals with the Asp/Glu or Glu/Glu genotype showed higher levels of damage with regard to all the studied measures, including aberrant cells, chromatid breaks, chromatid exchanges, deletions, and dicentrics.16
The combined results based on all the studies showed that no significant association was found between Asp148Glu and the risk of urinary cancers. Meanwhile, no significant associations were observed among Caucasians and African–Americans. However, a significant association was found among Asians. The discrepancy suggested a possible role of ethnic difference in genetic background and the environment. The same polymorphisms play different roles in cancer susceptibility among different ethnic populations, because cancer is a complicated multi-genetic disease, and different genetic background may contribute to the discrepancy.54 Meanwhile, when stratified by the quality score, no significant association was found in high-quality studies (score ≥7) but a significant increased risk of urinary cancers risk was observed in lower quality studies (score <7). The combined results based on all the high-quality studies further supported the previous conclusion that no significant association was found between Asp148Glu and the risk of urinary cancers.
One recent meta-analysis by Liu et al46 estimated the association between Asp148Glu and the risk of bladder cancer, which was basically in accordance with our opinion that Asp148Glu may not contribute to the susceptibility to bladder cancer. However, another published meta-analysis by Zhou et al48 showed that no significant association was found between Asp148Glu and the risk of prostate cancer, which is contrast with our results. One possible explanation of the contrast may be that different studies were included in the meta-analysis. As shown in the selection process of the studies, we mentioned that the studies by Mandal et al35 and Mittal et al36 were reported from the same organization. Actually, these two studies may contain partial overlapping data when carefully reading the full texts, and only the larger study36 should be selected for the analysis. However, the meta-analysis by Zhou et al48 included the studies by Mandal et al35 and Mittal et al,36 which might be biased by not taking into account the effects of overlapping data. Meanwhile, the study by Zhou et al48 was not included in the previous meta-analysis. Thus, the results of our meta-analysis are more accurate and reliable. However, in our meta-analysis, only two or three studies on prostate cancer were available for each specific ethnicity, and they had limited sample size; hence, larger studies are needed to explore the association between Asp148Glu and prostate cancer risk among Africans, Asians, and Caucasians.
Some limitations of our meta-analysis should be considered. First, the number of published studies included in our meta-analysis was not large enough for subgroup analyses by ethnicity and cancer type. Second, our results were unadjusted estimates because of lack of detailed data, such as age, sex, and environmental factors in the studies included. Third, some inevitable publication bias might exist in the results because only published studies were available to be included.
Conclusion
Our meta-analysis suggests that Asp148Glu was not associated with the risk of urinary cancers but might increase the risk of urinary cancers among Asians. Stratification by cancer type identified a significant association of Asp148Glu with prostate cancer. Additional larger studies, stratified by gene–gene and gene–environmental interactions, are needed to further explore the association between Asp148Glu and the susceptibility to urinary cancers.
Acknowledgment
This work was supported by the Program of the Pearl River Young Talents of Science and Technology in Guangzhou, People’s Republic of China (2013J2200042), National Natural Science Foundation of China (81201565, 81101536), and Natural Science Foundation of Guangdong Province, People’s Republic of China (S2012010009404).
Disclosure
The authors report no conflicts of interest in this work.
References
Jemal A, Bray F, Center MM, et al. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. | ||
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. 2015;65:5–29. | ||
Yu Z, Chen J, Ford BN, Brackley ME, Glickman BW. Human DNA repair systems: an overview. Environ Mol Mutagen. 1999;33:3–20. | ||
Zhu Y, Spitz MR, Amos CI, et al. An evolutionary perspective on single-nucleotide polymorphism screening in molecular cancer epidemiology. Cancer Res. 2004;64:2251–2257. | ||
Barzilay G, Hickson ID. Structure and function of apurinic/apyrimidinic endonucleases. Bioessays. 1995;17:713–719. | ||
Friedberg EC, McDaniel LD, Schultz RA. The role of endogenous and exogenous DNA damage and mutagenesis. Curr Opin Genet Dev. 2004;14:5–10. | ||
Tell G, Quadrifoglio F, Tiribelli C, Kelley MR. The many functions of APE1/Ref-1: not only a DNA repair enzyme. Antioxid Redox Signal. 2009;11:601–620. | ||
Izumi T, Hazra TK, Boldogh I, et al. Requirement for human AP endonuclease 1 for repair of 3′-blocking damage at DNA single-strand breaks induced by reactive oxygen species. Carcinogenesis. 2000;21:1329–1334. | ||
Fortini P, Pascucci B, Parlanti E, et al. The base excision repair: mechanisms and its relevance for cancer susceptibility. Biochimie. 2003;85:1053–1071. | ||
Bhakat KK, Mantha AK, Mitra S. Transcriptional regulatory functions of mammalian AP-endonuclease (APE1/Ref-1), an essential multifunctional protein. Antioxid Redox Signal. 2009;11:621–638. | ||
Robson CN, Hochhauser D, Craig R, et al. Structure of the human DNA repair gene HAP1 and its localisation to chromosome 14q 11.2-12. Nucleic Acids Res. 1992;20:4417–4421. | ||
Evans AR, Limp-Foster M, Kelley MR. Going APE over ref-1. Mutat Res. 2000;461:83–108. | ||
Xi T, Jones IM, Mohrenweiser HW. Many amino acid substitution variants identified in DNA repair genes during human population screenings are predicted to impact protein function. Genomics. 2004;83: 970–979. | ||
Hu JJ, Smith TR, Miller MS, et al. Amino acid substitution variants of APE1 and XRCC1 genes associated with ionizing radiation sensitivity. Carcinogenesis. 2001;22:917–922. | ||
Hadi MZ, Coleman MA, Fidelis K, Mohrenweiser HW, Wilson DM 3rd. Functional characterization of Ape1 variants identified in the human population. Nucleic Acids Res. 2000;28:3871–3879. | ||
Au WW, Salama SA, Sierra-Torres CH. Functional characterization of polymorphisms in DNA repair genes using cytogenetic challenge assays. Environ Health Perspect. 2003;111:1843–1850. | ||
Broberg K, Bjork J, Paulsson K, Hoglund M, Albin M. Constitutional short telomeres are strong genetic susceptibility markers for bladder cancer. Carcinogenesis. 2005;26:1263–1271. | ||
Andrew AS, Nelson HH, Kelsey KT, et al. Concordance of multiple analytical approaches demonstrates a complex relationship between DNA repair gene SNPs, smoking and bladder cancer susceptibility. Carcinogenesis. 2006;27:1030–1037. | ||
Chen L, Ambrosone CB, Lee J, et al. Association between polymorphisms in the DNA repair genes XRCC1 and APE1, and the risk of prostate cancer in white and black Americans. J Urol. 2006;175:108–112; discussion 112. | ||
Matullo G, Dunning AM, Guarrera S, et al. DNA repair polymorphisms and cancer risk in non-smokers in a cohort study. Carcinogenesis. 2006;27:997–1007. | ||
Terry PD, Umbach DM, Taylor JA. APE1 genotype and risk of bladder cancer: evidence for effect modification by smoking. Int J Cancer. 2006;118:3170–3173. | ||
Wu X, Gu J, Grossman HB, et al. Bladder cancer predisposition: a multigenic approach to DNA-repair and cell-cycle-control genes. Am J Hum Genet. 2006;78:464–479. | ||
Figueroa JD, Malats N, Real FX, et al. Genetic variation in the base excision repair pathway and bladder cancer risk. Hum Genet. 2007;121:233–242. | ||
Huang M, Dinney CP, Lin X, et al. High-order interactions among genetic variants in DNA base excision repair pathway genes and smoking in bladder cancer susceptibility. Cancer Epidemiol Biomarkers Prev. 2007;16:84–91. | ||
Andrew AS, Karagas MR, Nelson HH, et al. DNA repair polymorphisms modify bladder cancer risk: a multi-factor analytic strategy. Hum Hered. 2008;65:105–118. | ||
Gangwar R, Ahirwar D, Mandhani A, Mittal RD. Influence of XPD and APE1 DNA repair gene polymorphism on bladder cancer susceptibility in north India. Urology. 2009;73:675–680. | ||
Michiels S, Laplanche A, Boulet T, et al. Genetic polymorphisms in 85 DNA repair genes and bladder cancer risk. Carcinogenesis. 2009;30:763–768. | ||
Narter KF, Ergen A, Agachan B, et al. Bladder cancer and polymorphisms of DNA repair genes (XRCC1, XRCC3, XPD, XPG, APE1, hOGG1). Anticancer Res. 2009;29:1389–1393. | ||
Park JY, Huang Y, Sellers TA. Single nucleotide polymorphisms in DNA repair genes and prostate cancer risk. Methods Mol Biol. 2009;471:361–385. | ||
Lavender NA, Komolafe OO, Benford M, et al. No association between variant DNA repair genes and prostate cancer risk among men of African descent. Prostate. 2010;70:113–119. | ||
Ricceri F, Guarrera S, Sacerdote C, et al. ERCC1 haplotypes modify bladder cancer risk: a case-control study. DNA Repair (Amst). 2010;9:191–200. | ||
Wang M, Qin C, Zhu J, et al. Genetic variants of XRCC1, APE1, and ADPRT genes and risk of bladder cancer. DNA Cell Biol. 2010;29:303–311. | ||
Cao Q, Qin C, Meng X, et al. Genetic polymorphisms in APE1 are associated with renal cell carcinoma risk in a Chinese population. Mol Carcinog. 2011;50:863–870. | ||
Kuasne H, Rodrigues IS, Losi-Guembarovski R, et al. Base excision repair genes XRCC1 and APEX1 and the risk for prostate cancer. Mol Biol Rep. 2011;38:1585–1591. | ||
Mandal RK, Gangwar R, Kapoor R, Mittal RD. Polymorphisms in base-excision & nucleotide-excision repair genes & prostate cancer risk in north Indian population. Indian J Med Res. 2012;135:64–71. | ||
Mittal RD, Mandal RK, Gangwar R. Base excision repair pathway genes polymorphism in prostate and bladder cancer risk in North Indian population. Mech Ageing Dev. 2012;133:127–132. | ||
Jing B, Wang J, Chang WL, et al. Association of the polymorphism of APE1 gene with the risk of prostate cancer in Chinese Han population. Clin Lab. 2013;59:163–168. | ||
Thakkinstian A, McEvoy M, Minelli C, et al. Systematic review and meta-analysis of the association between b2-adrenoceptor polymorphisms and asthma: a HuGE review. Am J Epidemiol. 2005;162:201–211. | ||
Higgins JP, Thompson SG, Deeks JJ, Altman DG. Measuring inconsistency in meta-analyses. BMJ. 2003;327:557–560. | ||
Mantel N, Haenszel W. Statistical aspects of the analysis of data from retrospective studies of disease. J Natl Cancer Inst. 1959;22:719–748. | ||
DerSimonian R, Laird N. Meta-analysis in clinical trials. Control Clin Trials. 1986;7:177–188. | ||
Tobias A. Assessing the influence of a single study in the meta-analysis estimate. Stata Tech Bull. 1999;8:15–17. | ||
Dickersin K. The existence of publication bias and risk factors for its occurrence. JAMA. 1990;263:1385–1389. | ||
Egger M, Davey Smith G, Schneider M, Minder C. Bias in meta-analysis detected by a simple, graphical test. BMJ. 1997;315:629–634. | ||
Li X, Zhang G, Huai YJ, Cao ZQ. Association between APE1 T1349G polymorphism and prostate cancer risk: evidence from a meta-analysis. Tumour Biol. 2014;35(10):10111–10119. | ||
Liu C, Yin Q, Li L, et al. APE1 Asp148Glu gene polymorphism and bladder cancer risk: a meta-analysis. Mol Biol Rep. 2013;40:171–176. | ||
Pournourali M, Tarang AR, Yousefi M. The association between 1349T>G polymorphism of ApE1 gene and the risk of prostate cancer in northern Iran. Cell Mol Biol (Noisy-le-grand). 2015;61:21–24. | ||
Zhou X, Wei L, Jiao G, et al. The association between the APE1 Asp148Glu polymorphism and prostate cancer susceptibility: a meta-analysis based on case-control studies. Mol Genet Genomics. 2015;290(1):281–288. | ||
Qin Q, Zhang C, Zhu H, et al. Association between survivin-31G>C polymorphism and cancer risk: meta-analysis of 29 studies. J Cancer Res Clin Oncol. 2014;140:179–188. | ||
Hindorff LA, Gillanders EM, Manolio TA. Genetic architecture of cancer and other complex diseases: lessons learned and future directions. Carcinogenesis. 2011;32:945–954. | ||
Wiklund F. Prostate cancer genomics: can we distinguish between indolent and fatal disease using genetic markers? Genome Med. 2010;2:45. | ||
Varghese JS, Easton DF. Genome-wide association studies in common cancers – what have we learnt? Curr Opin Genet Dev. 2010;20:201–209. | ||
Amankwah EK, Sellers TA, Park JY. Gene variants in the angiogenesis pathway and prostate cancer. Carcinogenesis. 2012;33:1259–1269. | ||
Hirschhorn JN, Lohmueller K, Byrne E, Hirschhorn K. A comprehensive review of genetic association studies. Genet Med. 2002;4:45–61. |
Supplementary material
 | Table S1 Criteria for quality assessment of genetic associations of the APE1 Asp148Glu polymorphism with the risk of urinary cancers |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.