Back to Journals » Nature and Science of Sleep » Volume 17

Beyond the Pillow: Linking Subjective and Objective Sleep Measures to Gut Microbiome Composition in Community-Dwelling Older Adults
Authors Cheng CH ![]() , Hung CC
, Hung CC ![]() , Wu CY
, Wu CY ![]() , Lin CM, Fang JT
, Lin CM, Fang JT
Received 15 April 2025
Accepted for publication 16 September 2025
Published 14 October 2025 Volume 2025:17 Pages 2657—2668
DOI https://doi.org/10.2147/NSS.S531730
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Valentina Alfonsi
Chia-Hsiung Cheng,1– 4 Chun-Che Hung,5 Ching-Yi Wu,1,2,6 Ciao-Ming Lin,7 Ji-Tseng Fang8,9 On behalf of the Integrated Systematic Geriatric Medicine Information and Solutions for Exploring Healthy Aging (ISDHA)
1Department of Occupational Therapy and Graduate Institute of Behavioral Sciences, Chang Gung University, Taoyuan, Taiwan; 2Healthy Aging Research Center, Chang Gung University, Taoyuan, Taiwan; 3Laboratory of Brain Imaging and Neural Dynamics (BIND Lab), Chang Gung University, Taoyuan, Taiwan; 4Department of Neurology, Chang Gung Memorial Hospital, Kaohsiung, Taiwan; 5Department of Psychology, University of Florida, Gainesville, FL, USA; 6Department of Physical Medicine and Rehabilitation, Chang Gung Memorial Hospital, Linkou, Taiwan; 7Medical Development Center, Formosa Plastics Group, Taipei, Taiwan; 8Department of Nephrology, Chang Gung Memorial Hospital, Linkou, Taiwan; 9School of Medicine, College of Medicine, Chang Gung University, Taoyuan, Taiwan
Correspondence: Chia-Hsiung Cheng, Department of Occupational Therapy and Graduate Institute of Behavioral Sciences, Chang Gung University, Taoyuan, Taiwan, Email [email protected]
Background: Sleep-related complaints are common among older adults, and recent research indicates that changes in sleep patterns may be associated with alterations in the composition of the gut microbiome (GM). However, investigations into the relationship between sleep measures and GM abundance among older adults have been limited thus far. This study represents the first large-scale effort to comprehensively explore the connection between GM composition and both subjective and objective sleep measures in older adults.
Methods: The study included 279 cognitively-normal older adults from the community who had not used sleep medication, antibiotics, or probiotics for at least one month before providing stool samples. Participants were categorized as good sleepers (GS) or poor sleepers (PS) based on the Pittsburgh Sleep Quality Index (PSQI) scores. GM diversity and relative abundance were compared between both groups, and their associations with PSQI scores and objective sleep measures were also examined.
Results: Alpha and beta diversity did not show significant differences between the GS and PS groups. However, significant differences in GM relative abundance across various taxonomic levels were found between the GS and PS groups. In the overall sample, higher PSQI scores were linked to lower abundance of the species Hungatella_hathewayi (p = 0.005, false discovery rate = 0.035). However, there were no significant associations between GM abundance and objective sleep measures after corrections for multiple comparisons.
Conclusion: These findings suggest that specific gut microbial taxa are associated with subjective sleep disturbances in older adults.
Keywords: sleep quality, gut microbiome, aging, actigraphy
Introduction
Sleep complaints are common among older adults and can significantly impact their overall quality of life. Prior research from both the United States and France indicated that approximately 30% of older adults aged 65 years and above have reported poor sleep quality.1,2 Findings from East Asia demonstrated that the sleep complaints among individuals aged 60 years and above were more prevalent, with a prevalence ranging from 30% to 40%.3,4 The presence of sleep disturbances has been implicated in deteriorating health outcomes,5,6 and an increased risk of dementia7,8 when compared to individuals without sleep-related complaints. The regulation of sleep behavior involves both homeostatic and circadian processes, with the latter being connected to the composition of gut microbiome (GM).9,10
Emerging evidence from animal models has reported a link between sleep disturbance and GM composition.11,12 To date, only a limited number of studies have explored the relationship between sleep quality and the composition of GM in humans, with highly inconsistent findings. For younger adults, a study involving 28 healthy young participants demonstrated that the self-reported sleep quality, as measured by the Pittsburgh Sleep Quality Index (PSQI), was linked to 10 bacterial taxa ranging from the phylum to genus level.13 Another study recruited 34 college students and divided them into two groups of good sleep quality and poor sleep quality based on the scores of PSQI; the results showed 24 taxa with significantly differential abundance, mainly focused on phyla Firmicutes and Proteobacteria.14 Among older adults, only a few studies have explored the relationship between subjective sleep quality and GM composition.15–17 Anderson et al examined 37 healthy older adults and found that better sleep quality, indicated by lower PSQI scores, was associated with higher proportions of the phyla Verrucomicrobia and Lentisphaerae.15 Conversely, Wijaya et al analyzed a sample of 42 healthy older adults and reported that lower PSQI scores were linked to reduced abundance of genus Collinsella and Holdemania.17 These contrasting findings highlight the complexity of the relationship between subjective sleep quality and GM composition, underscoring the need for further research with larger and more diverse samples to clarify these associations.
Beyond self-reported sleep quality, objective sleep measures have been increasingly employed to explore their associations with GM composition.16–21 For example, Smith et al found a positive correlation between sleep efficiency and the relative abundance of the phyla Bacteroidetes and Firmicutes in healthy young males.18 In a broader age group ranging from 18 to 94 years, lower alpha diversity was observed alongside reduced sleep efficiency and increased wake after sleep onset (WASO).19 Similarly, recent studies have demonstrated positive associations between objective sleep measures and Chao1 richness in healthy older adults17 and with the relative abundance of the genus Lachnoclostridium in older adults with insomnia.20 In contrast, no significant relationship was identified between objectively measured sleep and GM diversity in older men.16 While these findings offer valuable insights—particularly within aging populations—the generalizability of the existing literature remains limited. Notably, two studies primarily involved Western samples, with one focusing exclusively on males16 and the other on older adults with clinical sleep disorders.20 Given that GM composition and abundance are influenced by a range of factors, including geographic region, genetics, and dietary patterns,22 such sample characteristics may restrict the broader applicability of these results. Although one study included Taiwanese older adults,17 its generalizability was similarly constrained by the limited number of participants who provided fecal samples. To enhance the robustness and generalizability of future research, there is an urgent need to investigate how GM alterations impact sleep behaviors in a larger and more diverse sample of East Asian older adults. This is particularly crucial given the rapid growth of this population segment in East Asia, driven by increasing life expectancy.23
Notably, all previous studies in this area have relied on next-generation sequencing (NGS) to analyze GM, which, while valuable, offers only limited insights into genetic structures. To address this gap and advance the field, the present study adopts a cutting-edge approach by utilizing third-generation sequencing (TGS) technology, which provides more comprehensive and detailed information on GM composition. Furthermore, this study features a substantially larger sample size of older adults (n > 200) than prior research, enhancing the statistical power and generalizability of the findings. The present study aims to achieve three primary objectives: First, we seek to compare GM diversity and abundance between older adults with good sleep quality (PSQI ≤ 5) and those with poor sleep quality (PSQI > 5). Second, contingent on identifying differences in GM profiles between these groups, we will explore the specific relationship between subjective sleep quality (as measured via PSQI) and GM abundance. Finally, we aim to extend these insights by examining the associations between objective sleep indicators (as measured via actigraphy) and GM abundance.
Materials and Methods
This study was approved by the Institutional Review Board of Chang Gung Memorial Hospital, Linkou, Taiwan (IRB number: 201900702A3). All the research procedures were conducted in accordance with the Declaration of Helsinki. Written informed consent was obtained from all participants.
Participants
Participants were recruited through the “Integrating Systematic Data of Geriatric Medicine to Explore Solutions for Healthy Aging” study, an observational cohort designed to develop a comprehensive geriatric medicine database for Taiwan. We focused on older adults residing in Songshan District (Taipei City) and Chang Gung Health and Culture Village (Taoyuan City), who were undergoing routine health evaluations at Chang Gung Memorial Hospital. Eligibility criteria included being 60 years of age or older, having visited Chang Gung Memorial Hospital at least once in the past year, and residing in Taiwan for over 180 days within the previous year. Exclusion criteria comprised: evidence of significant organ system dysfunction, severe autoimmune disorders or recent cancer therapy, a Mini-Mental State Examination (license has been obtained from PAR) score of 26 or below, a 15-item Chinese-version Geriatric Depression Scale [GDS, used with permission from Liao et al] score of 5 or higher,24,25 cognitive problems requiring outpatient follow-up, physician-diagnosed dementia or major depressive disorder, substantial sensory or cognitive impairments, and an inability to participate in interviews or significant frailty affecting mobility. Participants who had received antibiotic treatments or used probiotics within one month before stool sample collection were also excluded.
The above inclusion and exclusion criteria were established to minimize confounding factors that could affect GM composition or sleep quality, such as severe physical illnesses, major psychiatric conditions, or cognitive impairment. Participants were drawn from communities with established access to healthcare, which facilitated consistent data collection and follow-up. However, this may have introduced a sampling bias favoring older adults who are relatively health-conscious or functionally independent.
Assessment of Sleep Quality
Sleep quality was assessed using the Chinese version of the Pittsburgh Sleep Quality Index (PSQI), which evaluates sleep over the preceding month through 19 items divided into seven components: subjective sleep quality, sleep latency, sleep duration, habitual sleep efficiency, sleep disturbances, use of sleep medication, and daytime dysfunction. The total PSQI score ranges from 0 to 21, with scores exceeding 5 indicative of poor sleep quality.26–29 Permission to employ the PSQI in this study was obtained in advance.
Quantification of Sleep Behaviors
Participants were instructed to wear an Actiwatch-2 (Philips Respironics Inc., Pittsburgh, Pennsylvania) for a duration of one week to gather objective sleep data. Nights where the device was removed were excluded from the analysis. Key objective measures analyzed included total sleep time (hours), sleep onset latency (minutes), sleep efficiency (percentage), wake after sleep onset (WASO, minutes), and number of awakenings, with data averaged across the nights.
Fecal Sample DNA Collection and Sequencing Methods
Fecal samples were collected and total genomic DNA was extracted from the samples using the iCatcher Stool DNA Kit (Cat. No. AD22025; CatchGene Co., Ltd., Taiwan). DNA concentration was determined with a Qubit 4.0 Fluorometer (Thermo Fisher Scientific, USA). The full-length 16S rRNA gene was amplified via polymerase chain reaction (PCR) with barcoded 16S-specific primers, and sequencing was performed using PacBio’s Single Molecule, Real-Time (SMRT) technology (PacBio, Menlo Park, CA, USA). The PCR reaction, employing 2 ng of genomic DNA and KAPA HiFi HotStart ReadyMix (Roche), followed these conditions: 95°C for 3 minutes, 20–30 cycles of 95°C for 30 seconds, 57°C for 30 seconds, and 72°C for 60 seconds, followed by a final extension at 72°C for 5 minutes and storage at 4°C. The PCR products were assessed on a 1% agarose gel, and those with a prominent band at approximately 1500 bp were purified with AMPure PB Beads and used to prepare the SMRTbell library.
The SMRTbell library was incubated with Sequel II primer 3.1 and sequel II Binding Kit 3.1 for the primer annealing and polymerase binding. At last, sequencing was performed in the circular consensus sequence (CCS) mode on a PacBio Sequel IIe instrument to generate the HiFi reads with Predicted Accuracy (Phred Scale) = 30 (detailed protocols of DNA collection and sequencing methods were provided in the Supplementary Materials).
GM Analysis and Quality Control
The average depth of each sample is greater than 10K reads. CCS reads were processed using PacBio’s SMRT Link software, applying a minimum predicted accuracy of 0.9 and requiring at least three sequencing passes. Only reads with a Q30 quality score or higher, designated as Q30 HiFi reads, were used for analysis. After demultiplexing, HiFi reads were processed with the DADA2 pipeline (version 1.20) to achieve single-nucleotide resolution of amplicon sequence variants (ASVs).30 Reads were trimmed and filtered with a maximum expected error rate of two per read (maxEE = 2). Taxonomic classification of representative sequences was performed using QIIME2 (v2022.11). ASV sequence similarity was further analyzed using multiple sequence alignment with the MAFFT tool in QIIME2.31 Detailed protocols of GM analysis and quality control were described in the Supplementary Materials.
Statistical Analysis
Data were presented as frequencies with percentages or means with standard deviations (SD), as appropriate. Normality of data distribution was assessed using the Shapiro–Wilk test, which revealed non-normal distributions for most variables. Consequently, differences in demographic and clinical characteristics between good sleepers (GS) and poor sleepers (PS) were analyzed using the Mann–Whitney U-test or chi-square test as applicable. Alpha-diversity measures (eg, Simpson index, Shannon index, Observed features) were compared between GS and PS groups using the Mann–Whitney U-test. Beta diversity was assessed through Principal Coordinates Analysis (Weighted UniFrac) and permutational multivariate analysis of variance (PERMANOVA). Differences in GM abundance at various taxonomic levels between GS and PS groups were evaluated using the Mann–Whitney U-test. The Linear Discriminant Analysis Effect Size (LEfSe) method was employed to identify taxa with significant differences between GS and PS groups, with a significance threshold set at alpha = 0.05 and an LDA score greater than 2.32,33 The relationships between GM abundance and both subjective (ie, PSQI) and objective (ie, actigraphic-recorded sleep indicators, including total sleep time, onset latency, sleep efficiency, WASO, and number of awakenings) sleep measures were explored using partial correlation coefficients with GDS as a covariate. A significant level of p < 0.05 was applied to all statistical tests. All of the correlations were adjusted for multiple comparisons using the Benjamini-Hochberg method with a false discovery rate (FDR) of 0.05. In addition, effect sizes were also reported in this study.
Results
From September 2019 to June 2023, a total of 468 participants were enrolled in the Integrating Systematic Data of Geriatric Medicine to Explore the Solution for Healthy Aging study. From this cohort, 21 individuals who did not provide stool samples and 168 individuals who reported probiotic use within the past month were excluded. As a result, the analysis of GM diversity and abundance was conducted on a subset of 279 participants.
Table 1 shows the demographic information in GS (n = 171) and PS (n = 108) groups. Compared to the GS group, the PS group demonstrated significantly higher scores of PSQI (p < 0.001). There were no significant differences in age, educational attainments, body mass index, smoking status, dietary pattern, bowel habit, and presence of diabetes or hypertension between GS and PS groups. The PS group exhibited slightly elevated levels of depressive symptoms, assessed by GDS, in comparison to the GS group.
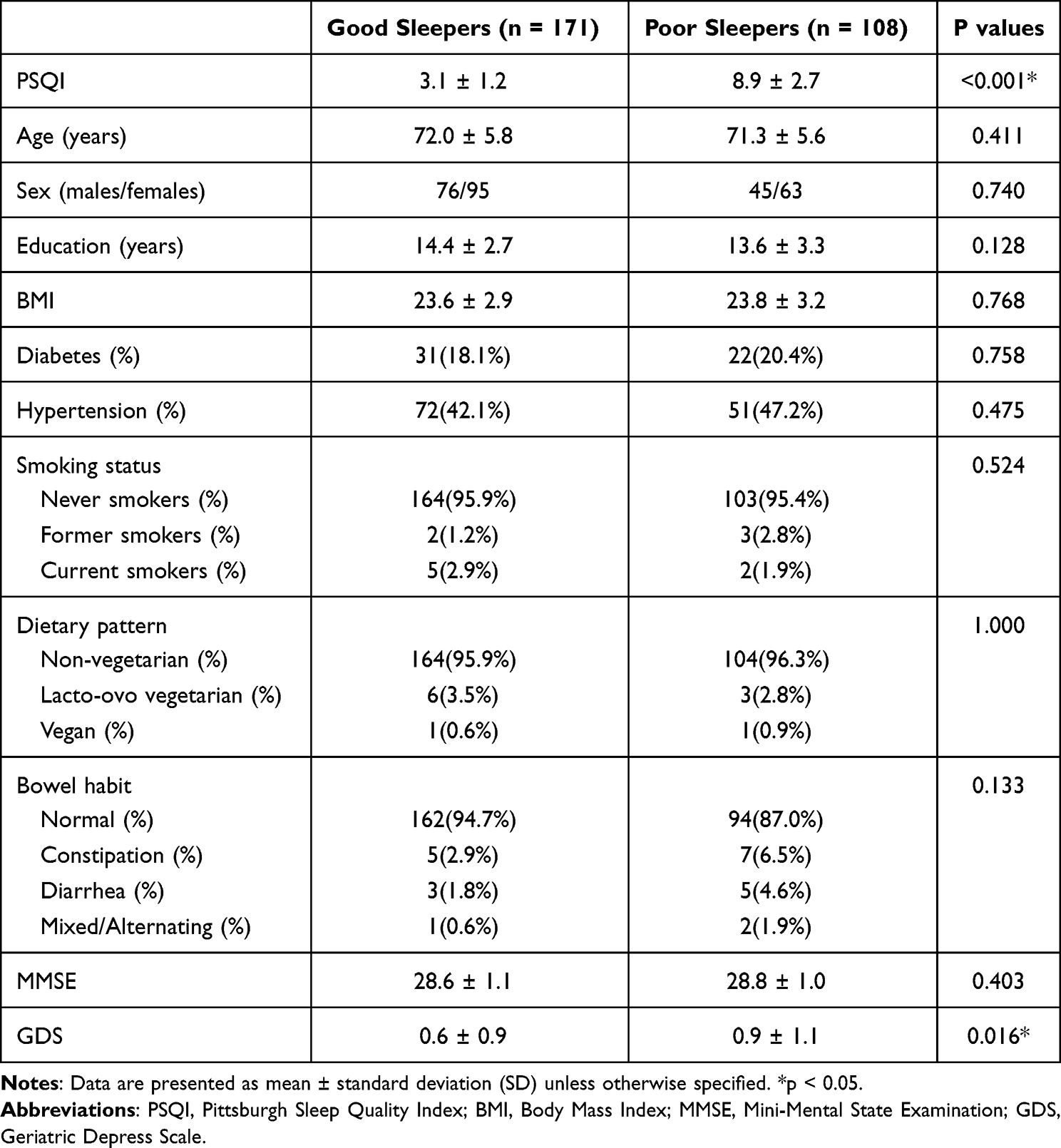 |
Table 1 Demographic Information in Good Sleepers (PSQI ≦ 5) and Poor Sleepers (PSQI > 5) |
Figure 1 displays the findings regarding GM diversity. There were no significant differences in alpha diversity between the GS and PS groups (Simpson index: p = 0.70, effect size = 0.023; Shannon index: p = 0.76, effect size = 0.018; Observed features: p = 0.67, effect size = 0.025). Additionally, beta-diversity analysis, conducted through Principal Coordinates Analysis with Ellipse (Weighted UniFrac), revealed no significant distinctions between the two groups (F = 0.311, R2 = 0.001, p = 0.97).
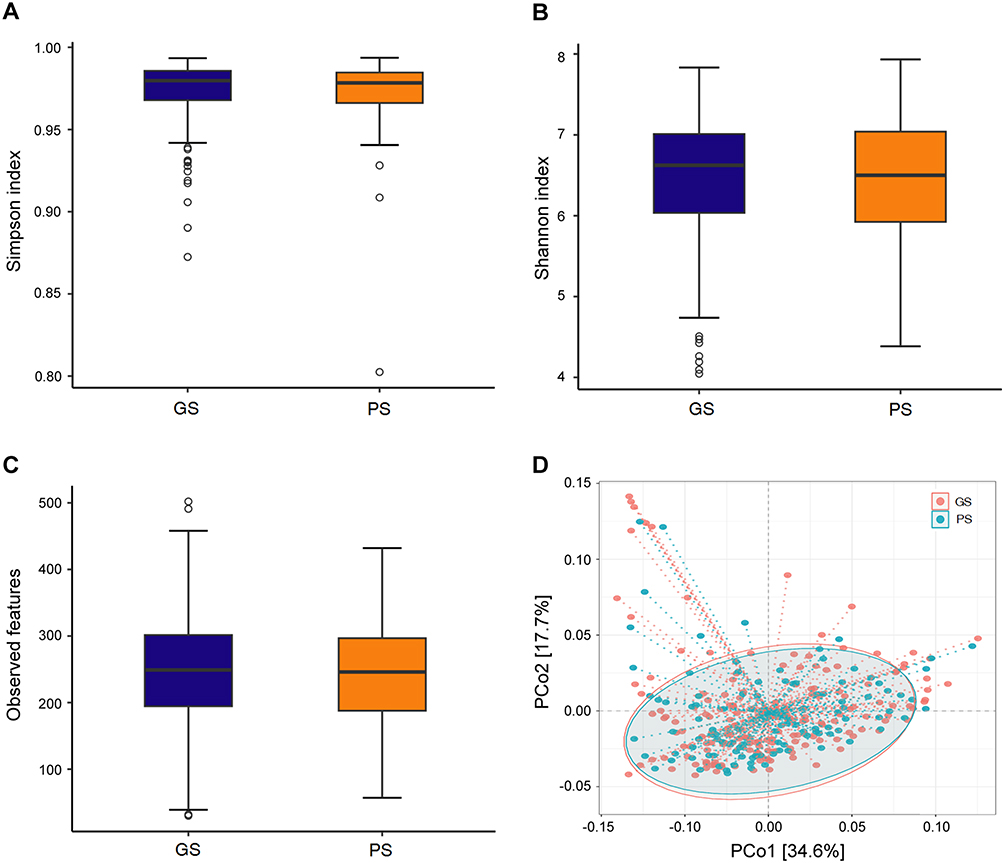 |
Figure 1 There were no significant differences in alpha diversity between the groups of good sleepers (GS) and poor sleepers (PS) as assessed by the Simpson index (A), Shannon index (B), and Observed features (C). Similarly, beta diversity, evaluated via weighted UniFrac by PCoA (D), showed no significant differences between the GS and PS groups. |
The results depicted in Figure 2 illustrate the variations in relative abundance of GM between the GS and PS groups. When considering the phylum level, no significant differences were shown in GM relative abundance between the two groups. However, at finer taxonomic levels, notable distinctions emerged. Specifically, at the class level, the PS group manifested a higher relative abundance of Erysipelotrichia (p = 0.038) compared to the GS group. Similarly, at the order level, the PS group displayed elevated relative abundances of both Erysipelotrichales (p = 0.038) and Moraxellales (p = 0.009) in contrast to the GS group. Moving further down the taxonomic hierarchy, at the family level, the PS group demonstrated more relative abundances of Dermacoccaceae (p = 0.029) and Moraxellaceae (p = 0.009) compared to the GS group. Examining at the genus level, the PS group exhibited increased relative abundances of Ructibacterium (p = 0.011), Intestinibacter (p = 0.028), and Dermacoccus (p = 0.029) compared to the GS group. Finally, at the species level, significant elevations in relative abundance were found in the PS group for Ructibacterium_gallinarum (p = 0.011), Bacteroides_fluxusYIT12057 (p = 0.001), Intestinibacter_bartlettii (p = 0.028), Dermacoccus_nishinomiyaensis (p = 0.029), and Sphingomonas_aquatilis (p = 0.025), whereas the relative abundances of Faecalibacterium_butyricigenerans (p = 0.045) and Hungatella_hathewayi (p = 0.030) were lower in the PS group. Please refer to the Supplementary Materials for the complete list of taxa and associated statistics.
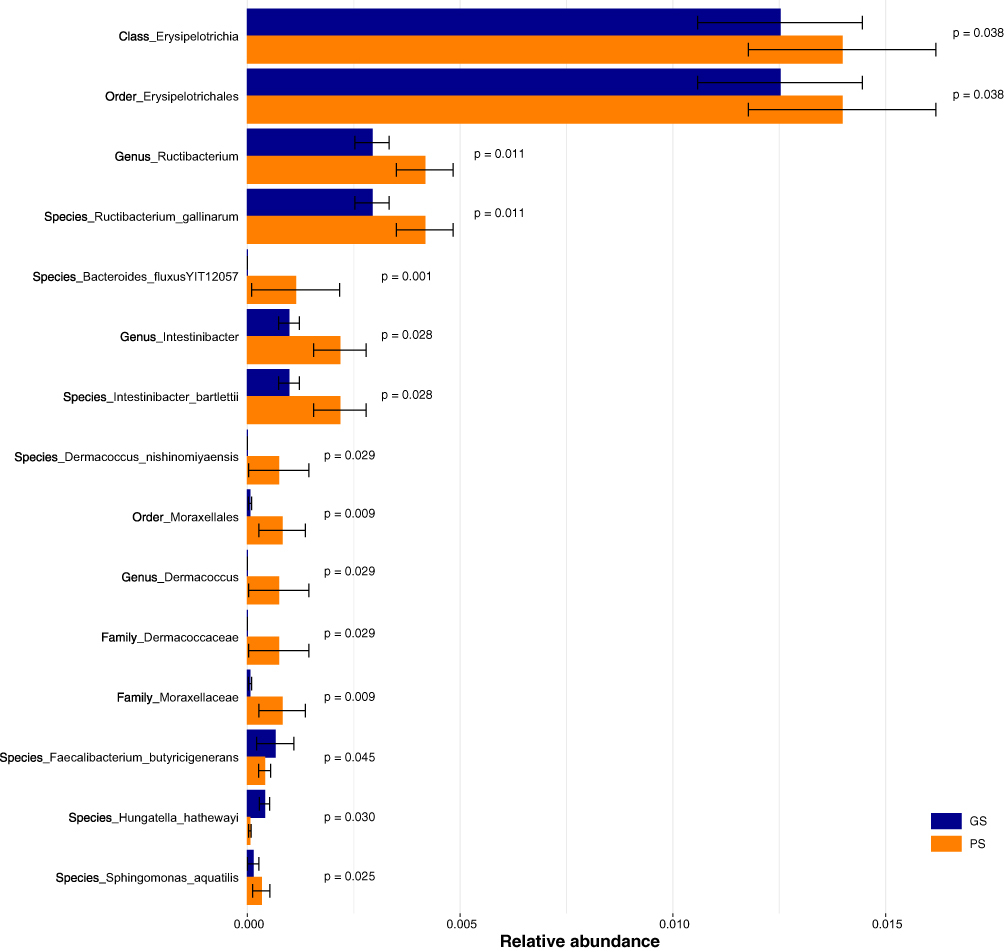 |
Figure 2 Comparisons of the relative abundance of gut microbiome between the good sleepers (GS) and poor sleepers (PS). The error bars represent standard deviation. |
Building upon the significant findings from the two-group comparisons mentioned above, we proceeded to investigate whether subjective perceptions of sleep quality correlated with GM abundance. As illustrated in Figure 3, we observed negative correlations between PSQI scores and the relative abundance of species Hungatella_hathewayi (partial r = −0.170, p = 0.005, FDR = 0.035, effect size = 0.353).
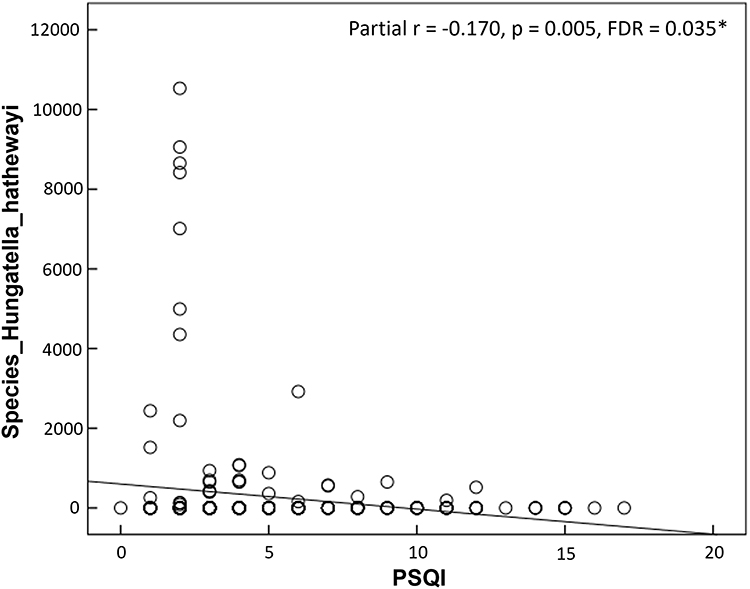 |
Figure 3 The scatter plots show the associations between self-reported sleep quality and relative abundance of gut microbiome (GM). Older adults with higher scores on Pittsburgh Sleep Quality Index (PSQI) were linked to reduced abundance of the species Hungatella_hathewayi. *p < 0.05. |
Beyond assessing self-reported sleep quality, we further examined whether objective measures of sleep behaviors (ie, total sleep time, onset latency, sleep efficiency, WASO, and number of awakenings) were linked to alterations in GM abundance. However, we did not detect any significant results after corrections for multiple comparisons. Table 2 also shows the comparisons of actigraphy-recorded data between the GS and PS groups.
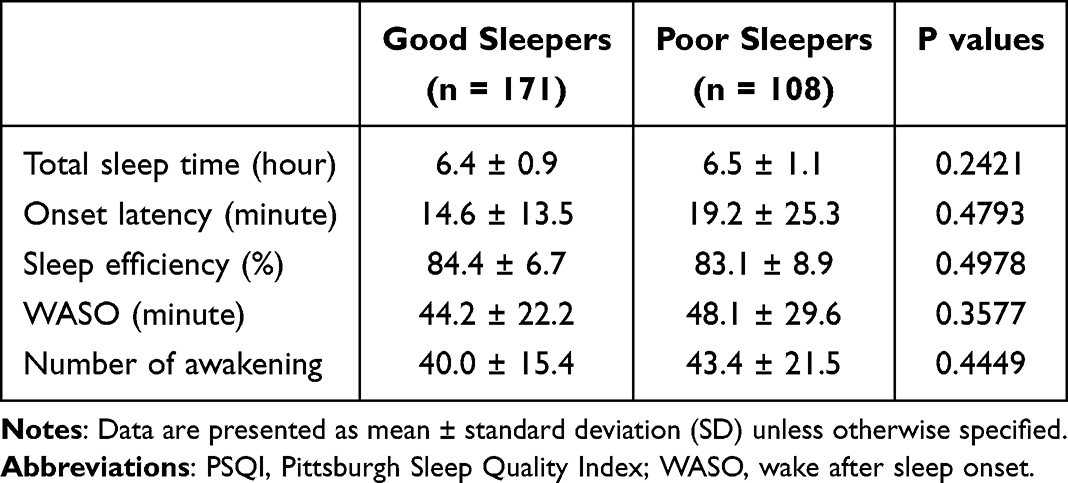 |
Table 2 Data Recorded by Actigraphy in Good Sleepers (PSQI ≦ 5) and Poor Sleepers (PSQI > 5) |
Discussion
To our knowledge, this study represents the first comprehensive investigation into the relationship of alterations in GM abundance with subjective and objective sleep measures among community-dwelling older adults aged 60 years and above, conducted on a large sample scale. Our research found significant differences of GM abundance were observed across various taxonomic levels between individuals classified as GS and PS. Furthermore, higher PSQI scores, indicative of poorer sleep quality, were associated with decreased relative abundance of species Hungatella_hathewayi. However, we did not detect any significant association between objective measures of sleep behaviors and alterations in GM abundance.
This study advances the current understanding of the relationship between sleep and GM composition by employing TGS technology (ie, PacBio) in a substantially larger sample of Taiwanese older adults (n = 279). Unlike previous research by Wijaya et al, which used NGS and included GM data from only 42 participants,17 our study offers enhanced statistical power and generalizability. Compared to NGS, the PacBio platform can achieve a higher estimate of richness and allow to determine organisms at a higher taxonomic and phylogenetic resolution.34,35 Accordingly, recent evidence has also suggested that PacBio is a more suitable technology than Illumina for human-associated microbiome studies.36,37 Furthermore, several factors are able to promote age-related degeneration and short-circuit possible compensatory mechanisms, such as individual genetic predisposition and health behaviors. In light of this, sleep is an important behavioral factor that has been closely tied to the inflammation and immune system. Specifically, sleep disturbances can weaken the immune system and increase the release of pro-inflammatory cytokines, including interleukin 6 and tumor necrosis factor-α.38,39 As comorbid sleep problems accompany normal aging, this health condition may set the stage for alterations in the microbiota-gut-brain axis and thus foster a brain vulnerable to pathology like the deposition of amyloid-β seen in Alzheimer’s disease.15,40 Hence, elucidating the association of sleep loss with GM composition is particular importance for older adults, which enables us to improve sleep problems through microbiota-targeted therapies,41,42 and eventually, prevent the initiation of dementia.
Of interest, GM has rhythmic activities, with fluctuations in both its composition and function over a 24-hour period.10 Sleep disruptions, however, can impair these rhythms and contribute to alterations in GM composition, driven by several interconnected physiological mechanisms.43 Specifically, disruptions in circadian rhythms can alter microbial diurnal patterns and reduce populations of beneficial bacteria,10 while activation of the hypothalamic–pituitary–adrenal (HPA) axis — a neuroendocrine system crucial for sleep regulation — and subsequent elevations in cortisol may impair gut barrier integrity and immune homeostasis, thereby disrupting communication along the microbiota–gut–brain axis.44 Similarly, dysregulation of autonomic nervous system, particularly reduced vagus nerve activity and increased sympathetic activity, has been shown to influence gut motility and microbial growth.45 Furthermore, depleted melatonin secretion in PS may decrease support for beneficial bacteria.46 Altogether, these mechanisms underline a bidirectional relationship between sleep and GM, where physiological changes from poor sleep contribute to GM alterations that can, in turn, further disrupt sleep. In our study, more abundance of species Hungatella_hathewayi is associated with better sleep quality in our study. It is noteworthy that Hungatella_hathewayi is a degrader of glycosaminoglycans, which plays a critical role in the cell signaling process (eg, wound repair and anticoagulation).47 Likewise, recent evidence has demonstrated that lower proportions of Hungatella_hathewayi are relevant to depleted concentrations of taurine, an anti-inflammatory metabolite known to prevent from traumatic brain injury or stroke.48 Moreover, Jiao and colleagues have reported that compared with non-nonagenarians, long-lived individuals manifest enrichment of agmatine as evident by increased levels of Hungatella_hathewayi, together with enhanced functional connectivity, suggesting its potential anti-aging effects on brain function.49 Taken together, the Hungatella_hathewayi is considered as an advantageous GM strain.
This study has several limitations. First, the composition of GM can be influenced by various factors, including geographic location and dietary habits. Additionally, the sampling approach—focused on specific residential areas and individuals actively engaged in healthcare services—may limit the generalizability of our findings. This recruitment strategy may introduce selection bias, as participants could differ systematically from the broader older adult population in terms of health status, lifestyle, or healthcare access. Although the sample size in our study is relatively large, caution should still be emphasized when extrapolating the results to other demographic and geographic groups.22 Second, the lack of cytokine data from participants limited our ability to explore potential links between sleep parameters, dysbiotic strains, and pro-inflammatory cytokines. Future research is needed to elucidate these relationships and to inform the development of targeted therapeutic strategies. Third, participants’ sleep behaviors were not assessed using a sleep diary. In both research and clinical contexts, combining actigraphy with a sleep diary is recommended, as they provide complementary information.50 A sleep diary can aid in the accurate interpretation of actigraphy data by helping to distinguish between sleep and periods of inactivity, as well as marking bedtime and wake time. Therefore, the absence of sleep diary data in our study may limit the precision of actigraphy-derived sleep parameters and should be considered when interpreting the findings. Finally, although actigraphy provides a practical and validated estimate of sleep-wake patterns in real-world settings, it lacks the precision of other objective measures such as polysomnography (PSG). This might be one of the reasons that we did not detect any significant association between GM abundance and objective sleep measures. Including PSG in future studies would allow for a more comprehensive evaluation of sleep architecture (eg, sleep stages, arousals, and apnea events) and may help further elucidate the relationship between GM and specific physiological components of sleep.
Although the current cross-sectional findings highlight significant associations between specific microbial taxa and subjective sleep parameters, they do not establish directionality or underlying mechanisms. To address this, longitudinal studies are needed to monitor changes in GM composition and sleep patterns over time, allowing for the identification of temporal relationships. Furthermore, interventional research—such as probiotic or dietary interventions designed to modify the gut microbiota—will be crucial to directly test whether altering GM can lead to measurable improvements in sleep quality. These research directions are essential for uncovering causal pathways and evaluating the therapeutic potential of microbiota-targeted interventions, particularly for individuals with poor sleep quality in aging populations.
Conclusion
To conclude, this population-based study indicated that specific GM taxa are associated with subjective sleep disturbances in older adults.
Data Sharing Statement
The datasets generated and analyzed during the current study are not publicly available due to privacy considerations and the protection of individual rights, but are available from the corresponding author on reasonable request.
Ethical Approval and Informed Consent
This study was approved by the Institutional Review Board of Chang Gung Memorial Hospital, Linkou, Taiwan (IRB number: 201900702A3). Written informed consent was obtained from all participants.
Acknowledgments
We thank the Genomic and Proteomic Core Laboratory of Chang Gung Memorial Hospital for the assistance of experiment conduction. We would like to acknowledge Ms. Wei-Li Chen, from the Formosa Plastics group, for helping us procure the Mini-Mental State Examination used in this study.
Author Contributions
Chia-Hsiung Cheng: Conceptualization, methodology, investigation, data curation, formal analysis, writing - the original draft.
Chun-Che Hung: Methodology, formal analysis, visualization, writing - the original draft.
Ching-Yi Wu: Methodology, writing – review & editing, supervision, funding acquisition.
Ciao-Ming Lin: Data curation, project administration, formal analysis, writing - review & editing.
Ji-Tseng Fang: Methodology, writing – review & editing, funding acquisition.
All authors have agreed on the target journal for submission and collectively take responsibility for the content of the article. Each author has reviewed and approved the final version to be published. During the preparation of this work, the authors used ChatGPT in order to improve language and readability. After using this tool, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication.
Funding
This study was supported by Chang Gung Memorial Hospital (CORPG3J0371, CMRPD1M0741, URRPD1P0131), Wang Chang Gung Charity Trust (XMRPG3L1021), National Science and Technology Council (NSTC-112-2410-H-182-030, NSTC-113-2314-B-182-064, NSTC-114-2314-B-182-016-MY3), Taiwan. The authors acknowledge the data collection, data management, and information technology services, based on the Structured Research and Medical Informatics Cloud (SRMIC) platform system, provided by the Clinical Trial Center, Chang Gung Memorial Hospital, Linkou, Taiwan, with support by the Ministry of Health and Welfare MOHW110-TDU-B-212-124005).
Disclosure
The authors have no conflict of interest to disclose.
References
1. Endeshaw YW, Yoo W. Association between social and physical activities and insomnia symptoms among community-dwelling older adults. J Aging Health. 2016;28(6):1073–1089. doi:10.1177/0898264315618921
2. Akbaraly TN, Jaussent I, Besset A, et al. Sleep complaints and metabolic syndrome in an elderly population: the Three-City Study. Am J Geriatr Psychiatry. 2015;23(8):818–828. doi:10.1016/j.jagp.2014.10.001
3. Wang P, Song L, Wang K, et al. Prevalence and associated factors of poor sleep quality among Chinese older adults living in a rural area: a population-based study. Aging Clin Exp Res. 2020;32(1):125–131. doi:10.1007/s40520-019-01171-0
4. Wang YM, Chen HG, Song M, et al. Prevalence of insomnia and its risk factors in older individuals: a community-based study in four cities of Hebei Province, China. Sleep Med. 2016;19:116–122. doi:10.1016/j.sleep.2015.10.018
5. Cross NE, Carrier J, Postuma RB, et al. Association between insomnia disorder and cognitive function in middle-aged and older adults: a cross-sectional analysis of the Canadian Longitudinal Study on Aging. Sleep. 2019;42(8). doi:10.1093/sleep/zsz114
6. Beydoun MA, Tate R, Georgescu MF, et al. Poor sleep quality, dementia status and their association with all-cause mortality among older US adults. Aging. 2024;16(17):12138–12167. doi:10.18632/aging.206102
7. Shi L, Chen SJ, Ma MY, et al. Sleep disturbances increase the risk of dementia: a systematic review and meta-analysis. Sleep Med Rev. 2018;40:4–16. doi:10.1016/j.smrv.2017.06.010
8. Ferini-Strambi L. Sleep disorders and increased risk of dementia. Eur J Neurol. 2022;29(12):3484–3485. doi:10.1111/ene.15562
9. Liang X, Bushman FD, FitzGerald GA. Rhythmicity of the intestinal microbiota is regulated by gender and the host circadian clock. Proc Natl Acad Sci U S A. 2015;112(33):10479–10484. doi:10.1073/pnas.1501305112
10. Thaiss CA, Zeevi D, Levy M, et al. Transkingdom control of microbiota diurnal oscillations promotes metabolic homeostasis. Cell. 2014;159(3):514–529. doi:10.1016/j.cell.2014.09.048
11. Badran M, Khalyfa A, Ericsson A, et al. Fecal microbiota transplantation from mice exposed to chronic intermittent hypoxia elicits sleep disturbances in naive mice. Exp Neurol. 2020;334:113439. doi:10.1016/j.expneurol.2020.113439
12. El Aidy S, Bolsius YG, Raven F, et al. A brief period of sleep deprivation leads to subtle changes in mouse gut microbiota. J Sleep Res. 2020;29(6):e12920. doi:10.1111/jsr.12920
13. Grosicki GJ, Riemann BL, Flatt AA, et al. Self-reported sleep quality is associated with gut microbiome composition in young, healthy individuals: a pilot study. Sleep Med. 2020;73:76–81. doi:10.1016/j.sleep.2020.04.013
14. Zhang J, Zhang X, Zhang K, et al. The component and functional pathways of gut microbiota are altered in populations with poor sleep quality - a preliminary report. Pol J Microbiol. 2022;71(2):241–250. doi:10.33073/pjm-2022-021
15. Anderson JR, Carroll I, Azcarate-Peril MA, et al. A preliminary examination of gut microbiota, sleep, and cognitive flexibility in healthy older adults. Sleep Med. 2017;38:104–107. doi:10.1016/j.sleep.2017.07.018
16. Estaki M, Langsetmo L, Shardell M, et al. Association of subjective and objective measures of sleep with gut microbiota composition and diversity in older men: the osteoporotic fractures in men study. J Gerontol a Biol Sci Med Sci. 2023;78(10):1925–1932. doi:10.1093/gerona/glad011
17. Wijaya MT, Fang JT, Liu GH, et al. Better objective sleep quality is associated with higher gut microbiota richness in older adults. Geroscience. 2025;47:4121–4137. doi:10.1007/s11357-025-01524-w
18. Smith RP, Easson C, Lyle SM, et al. Gut microbiome diversity is associated with sleep physiology in humans. PLoS One. 2019;14(10):e0222394. doi:10.1371/journal.pone.0222394
19. Holzhausen EA, Peppard PE, Sethi AK, et al. Associations of gut microbiome richness and diversity with objective and subjective sleep measures in a population sample. Sleep. 2024;47(3). doi:10.1093/sleep/zsad300
20. Haimov I, Magzal F, Tamir S, et al. Variation in gut microbiota composition is associated with sleep quality and cognitive performance in older adults with insomnia. Nat Sci Sleep. 2022;14:1753–1767. doi:10.2147/NSS.S377114
21. Carpena MX, Barros AJ, Comelli EM, et al. Accelerometer-based sleep metrics and gut microbiota during adolescence: association findings from a Brazilian population-based birth cohort. Sleep Med. 2024;114:203–209. doi:10.1016/j.sleep.2023.12.028
22. Hung CC, Chang CC, Huang CW, et al. Gut microbiota in patients with Alzheimer’s disease spectrum: a systematic review and meta-analysis. Aging. 2022;14(1):477–496. doi:10.18632/aging.203826
23. Abbasi BN, Sohail A. Ramification of healthcare expenditure on morbidity rates and life expectancy in the association of Southeast Asian nations countries: a dynamic panel threshold analysis. Int J Health Plann Manage. 2022;37(6):3218–3237. doi:10.1002/hpm.3551
24. Liao YC, Yeh TL, Ko HC, et al. Geriatric depression scale--validity and reliability of the Chinese-translated version: a preliminary study. Changhua J Med. 1995;1:11–17.
25. Sheikh JI, Yesavage JA. Geriatric Depression Scale (GDS): recent evidence and development of a shorter version. Clin Gerontologist. 1986;5:165–173. doi:10.1300/J018v05n01_09
26. Buysse DJ, Reynolds CF, Monk TH, et al. The Pittsburgh Sleep Quality Index: a new instrument for psychiatric practice and research. Psychiatry Res. 1989;28(2):193–213. doi:10.1016/0165-1781(89)90047-4
27. Tsai PS, Wang SY, Wang MY, et al. Psychometric evaluation of the Chinese version of the Pittsburgh Sleep Quality Index (CPSQI) in primary insomnia and control subjects. Qual Life Res. 2005;14(8):1943–1952. doi:10.1007/s11136-005-4346-x
28. Miller JK, Evans SL. Perceived stress, but not rumination, mediates the relationship between trait mindfulness and sleep quality in young adults. Nat Sci Sleep. 2024;16:1053–1065. doi:10.2147/NSS.S447469
29. Wei J, Song M, Mao HJ, et al. Analysis of the improvement sequence in insomnia symptoms and factors influencing the treatment outcomes of smartphone-delivered CBT in patients with insomnia disorder. Nat Sci Sleep. 2024;16:1365–1376. doi:10.2147/NSS.S486288
30. Callahan BJ, McMurdie PJ, Rosen MJ, et al. DADA2: high-resolution sample inference from Illumina amplicon data. Nat Methods. 2016;13(7):581–583. doi:10.1038/nmeth.3869
31. Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 2013;30(4):772–780. doi:10.1093/molbev/mst010
32. Qian Y, Yang X, Xu S, et al. Alteration of the fecal microbiota in Chinese patients with Parkinson’s disease. Brain Behav Immun. 2018;70:194–202. doi:10.1016/j.bbi.2018.02.016
33. Hung CC, Chao YP, Lee Y, et al. Cingulate white matter mediates the effects of fecal Ruminococcus on neuropsychiatric symptoms in patients with amyloid-positive amnestic mild cognitive impairment. BMC Geriatr. 2023;23(1):720. doi:10.1186/s12877-023-04417-9
34. Mosher JJ, Bowman B, Bernberg EL, et al. Improved performance of the PacBio SMRT technology for 16S rDNA sequencing. J Microbiol Methods. 2014;104:59–60. doi:10.1016/j.mimet.2014.06.012
35. Earl JP, Adappa ND, Krol J, et al. Species-level bacterial community profiling of the healthy sinonasal microbiome using Pacific Biosciences sequencing of full-length 16S rRNA genes. Microbiome. 2018;6(1):190. doi:10.1186/s40168-018-0569-2
36. Zhang J, Su L, Wang Y, et al. Improved high-throughput sequencing of the human oral microbiome: from Illumina to PacBio. Can J Infect Dis Med Microbiol. 2020;2020:6678872. doi:10.1155/2020/6678872
37. Buetas E, Jordan-Lopez M, Lopez-Roldan A, et al. Full-length 16S rRNA gene sequencing by PacBio improves taxonomic resolution in human microbiome samples. BMC Genomics. 2024;25(1):310. doi:10.1186/s12864-024-10213-5
38. Garbarino S, Lanteri P, Bragazzi NL, et al. Role of sleep deprivation in immune-related disease risk and outcomes. Commun Biol. 2021;4(1):1304. doi:10.1038/s42003-021-02825-4
39. Dzierzewski JM, Donovan EK, Kay DB, et al. Sleep inconsistency and markers of inflammation. Front Neurol. 2020;11:1042. doi:10.3389/fneur.2020.01042
40. Hung CC, Crowe-White KM, McDonough IM. A seed and soil model of gut dysbiosis in Alzheimer’s disease. Aging. 2023;15(12):5235–5237. doi:10.18632/aging.204840
41. Irwin C, McCartney D, Desbrow B, et al. Effects of probiotics and paraprobiotics on subjective and objective sleep metrics: a systematic review and meta-analysis. Eur J Clin Nutr. 2020;74(11):1536–1549. doi:10.1038/s41430-020-0656-x
42. Ito H, Tomura Y, Kitagawa Y, et al. Effects of probiotics on sleep parameters: a systematic review and meta-analysis. Clin Nutr ESPEN. 2024;63:623–630. doi:10.1016/j.clnesp.2024.07.006
43. Matenchuk BA, Mandhane PJ, Kozyrskyj AL. Sleep, circadian rhythm, and gut microbiota. Sleep Med Rev. 2020;53:101340. doi:10.1016/j.smrv.2020.101340
44. Buckley TM, Schatzberg AF. On the interactions of the hypothalamic-pituitary-adrenal (HPA) axis and sleep: normal HPA axis activity and circadian rhythm, exemplary sleep disorders. J Clin Endocrinol Metab. 2005;90(5):3106–3114. doi:10.1210/jc.2004-1056
45. Bonaz B, Bazin T, Pellissier S. The vagus nerve at the interface of the microbiota-gut-brain axis. Front Neurosci. 2018;12:49. doi:10.3389/fnins.2018.00049
46. Wang Z, Wang Z, Lu T, et al. The microbiota-gut-brain axis in sleep disorders. Sleep Med Rev. 2022;65:101691. doi:10.1016/j.smrv.2022.101691
47. Shang Q, Yin Y, Zhu L, et al. Degradation of chondroitin sulfate by the gut microbiota of Chinese individuals. Int J Biol Macromol. 2016;86:112–118. doi:10.1016/j.ijbiomac.2016.01.055
48. Li H, Xu H, Li Y, et al. Alterations of gut microbiota contribute to the progression of unruptured intracranial aneurysms. Nat Commun. 2020;11(1):3218. doi:10.1038/s41467-020-16990-3
49. Jiao B, Ouyang Z, Liu Q, et al. Integrated analysis of gut metabolome, microbiome, and brain function reveal the role of gut-brain axis in longevity. Gut Microbes. 2024;16(1):2331434. doi:10.1080/19490976.2024.2331434
50. Sadeh A. The role and validity of actigraphy in sleep medicine: an update. Sleep Med Rev. 2011;15(4):259–267. doi:10.1016/j.smrv.2010.10.001
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.