Back to Journals » Drug Design, Development and Therapy » Volume 20
Artemisinin-Related Therapeutic Strategies for Autoimmune Thyroiditis: Chemokine–Receptor Networks, Spatial Thyroid Biology, and Molecular Mechanisms
Authors Sun Y, Yang X, Zhu Q, Yang R
Received 2 April 2026
Accepted for publication 9 June 2026
Published 18 June 2026 Volume 2026:20 614133
DOI https://doi.org/10.2147/DDDT.S614133
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Yuanyuan Sun,1 Xuelian Yang,2 Qimo Zhu,3 Rong Yang4
1Department of Endocrinology, The People’s Hospital of Liaoning Province, Shenyang, 110016, People’s Republic of China; 2Material Management Department, Shengjing Hospital of China Medical University, Shenyang, 110004, People’s Republic of China; 3The Second Clinical College, China Medical University, Shenyang, 110004, People’s Republic of China; 4Department of Endocrinology, Shengjing Hospital of China Medical University, Shenyang, 110004, People’s Republic of China
Correspondence: Rong Yang, Department of Endocrinology, Shengjing Hospital of China Medical University, No. 36, Sanhao Street, Shenyang, 110004, People’s Republic of China, Tel +86-24-24016709, Email [email protected]
Abstract: Autoimmune thyroiditis (AIT) is a chronic immune-mediated thyroid disorder characterized by lymphocytic infiltration, follicular epithelial injury, autoantibody production, and variable progression to thyroid dysfunction. Because current management is largely supportive or replacement based, there is growing interest in interventions that might modify inflammatory and immune processes before irreversible tissue damage develops. This narrative review critically evaluates the potential, but not yet established, role of Artemisia-derived and artemisinin-related compounds in AIT. The analysis focuses on chemokine–receptor networks, spatial organization of thyroid inflammation, and molecular mechanisms linking endoperoxide/redox chemistry, innate immune signaling, and immune-cell trafficking. Evidence was identified through targeted, non-systematic searches of PubMed and Google Scholar, supported by backward and forward citation tracking. Thyroid-specific evidence remains limited and predominantly preclinical. Dihydroartemisinin (DHA) has been reported to attenuate experimental autoimmune thyroiditis by reducing inflammatory infiltration, thyroid autoantibodies, Th1/Th17-associated responses, CXCL10/CXCR3-linked signaling, and oxidative stress. Evidence from non-thyroid autoimmune and inflammatory models suggests additional effects on NF-κB, MAPK, PI3K/Akt/mTOR, JAK/STAT, TLR/MyD88, NRF2/GPX4-related redox responses, inflammasome activity, regulatory T-cell balance, and possible epigenetic remodeling; however, these findings should be treated as mechanistic context rather than direct evidence for human AIT. Most available studies involve purified or semi-synthetic compounds rather than standardized botanical preparations, and robust clinical validation is absent. Artemisinin-related compounds are therefore framed as hypothesis-driven immunomodulatory candidates that require thyroid-specific replication, target validation, comparative pharmacology, safety assessment, formulation standardization, and carefully designed translational studies before clinical application can be considered. The infographic details artemisinin-related chemotypes in autoimmune thyroiditis (AIT). 1. Disease-Relevant Pathways Potentially Modulated: NF-kB and MAPK (Inflammation), PI3K/Akt/mTOR (Cell survival), JAK/STAT (Cytokine signaling), TLR/MyD88 (Innate immunity), NRF2/Redox (Oxidative balance). Modulation may vary. 2. Reported Mainly for DHA in EAT models: Inflammatory infiltration, Autoantibodies, Th1/Th17 responses, CXCL10/CXCR3-linked signaling, Oxidative stress. 3. Key Limitations: Standardization of botanical preparations, Compound authentication, Dose-response validation, Safety/toxicity, Lack of robust human AIT trials. 4. Candidate Chemotypes (Evidence Strength): D (Direct preclinical EAT evidence: Dihydroartemisinin), S (Supportive: Amino-artemisinins), H (Hypothesis-generating: Synthetic ozonides). 5. Cautious Translational Pathway: Target validation, Early translational studies, Clinical development if justified. All candidate compounds remain investigational.Artemisinin chemotypes in thyroiditis: pathways, limits and translation.
Keywords: autoimmune thyroiditis, artemisinin derivatives, chemokine–receptor networks, spatial thyroid biology, innate immune signaling
Introduction
Autoimmune thyroiditis (AIT), including chronic lymphocytic or Hashimoto thyroiditis, is an organ-specific autoimmune disease in which immune infiltration, follicular epithelial injury, autoantibody production, and progressive thyroid dysfunction may occur over time.1–3 Histologically, affected glands show lymphocytic infiltration, follicular disruption, oxyphilic change, fibrosis-prone remodeling, and sometimes tertiary lymphoid structures (TLSs).4 Chemokines help organize these infiltrates, and CXCR3 ligands (CXCL9, CXCL10, and CXCL11) contribute to Th1-skewed inflammation.5 Clinically, this inflammatory anatomy corresponds to measurable patient phenotypes, including TPOAb/TgAb positivity, hypoechoic thyroid ultrasound patterns, altered thyroid volume, and progression from euthyroidism or subclinical disease toward overt hypothyroidism.6,7 Recent cohorts also link higher thyroid autoantibodies with symptom burden, lower quality of life, and higher TNF-alpha/IFN-gamma, while diagnostic studies show that ultrasound and antibody testing provide useful but incomplete clinical anchors.8,9 Together, these data anchor immune mechanisms to the clinical variables by which AIT is diagnosed, monitored, and treated.
Thyroid follicular epithelial cells are active participants in this process. After cytokine exposure, they can produce CXCL10 and other chemokines that recruit CXCR3-positive Th1/Tc cells and reinforce local IFN-gamma/TNF-alpha-driven inflammation.10 Serum CXCL10/IP-10 is increased in chronic autoimmune thyroiditis, especially in hypothyroid or hypoechoic cases, but it is not sufficiently specific as a stand-alone biomarker.6,11–13 This clinical evidence is important because it links a mechanistic pathway to endocrine decline and sonographic remodeling, while also showing why CXCL10 cannot be interpreted in isolation. Therefore, CXCL10 is best interpreted alongside TSH/FT4, thyroid autoantibodies, ultrasound echogenicity or volume, symptoms, levothyroxine status, and inflammatory context.
Accordingly, CXCL10 is best treated as one node within a broader chemokine network. Thyrocytes, stromal cells, endothelial cells, and infiltrating leukocytes express multiple CC and CXC chemokines.14 These ligands signal through overlapping receptors, including CXCR3, CCR2, CCR5, CCR6, CCR7, CXCR5, CXCR1/2, CXCR4/ACKR3, CX3CR1, XCR1, and atypical chemokine receptors. Because these pathways are redundant and compensatory, blocking one ligand may be buffered by parallel trafficking circuits.14 Systems-level evidence is more consistent with coordinated dysregulation of several chemokine nodes than isolated CXCL10 perturbation.15–18 Clinical profiling supports this broader view: circulating CXCL9 and CXCL11 are also elevated in newly diagnosed autoimmune thyroiditis and increase with older age, hypoechogenicity, or hypothyroidism.19,20 Spatial and histological evidence further indicates that AIT is organized by tissue architecture rather than diffuse inflammation alone.21,22 Spatial transcriptomics has identified neighborhoods containing follicular cells, fibroblasts, endothelial cells, and immune subsets.23 TLS-like aggregates and perivascular inflammatory cuffs may carry distinct chemokine signatures, including CCL19, CCL21, and CXCL13 pathways that support antigen presentation, B-cell maturation, and T-cell priming.4,24–26 Recent human spatial data therefore strengthen the concept of intrathyroidal inflammatory niches, while preliminary serum data linking CCL21/CCR7 with thyroid autoantibodies suggest that TLS-related trafficking may have measurable clinical correlates.21,27 This spatial perspective also helps explain why circulating chemokines alone may not capture disease activity: the same serum value may reflect different follicular, perivascular, stromal, or TLS-like compartments.
Because thyroid-specific interventional data remain limited, this review applies an evidence-disciplined framework. AIT/EAT findings are prioritized, whereas non-thyroid autoimmune or inflammatory studies are used only as supportive mechanistic context. Artemisinin chemotypes, including semi-synthetic derivatives, amino-artemisinins, ozonides/trioxolanes, and dimeric or hybrid structures, show broad immunoregulatory activity outside malaria.28 Across experimental systems, these agents can influence NF-kappaB, MAPK, JAK-STAT, NRF2-linked redox responses, inflammasome activation, Th1/Th17-Treg balance, and macrophage polarization.28,29 Reports of reduced NF-kappaB-controlled chemokines and receptors create a rationale for studying chemokine-network modulation in AIT, but current care mainly restores thyroid hormone status and does not prove resolution of intrathyroidal autoimmunity. Thus, artemisinin-related mechanisms are framed as an untested immunomodulatory adjunct hypothesis, not established AIT efficacy.29–32 This boundary prevents mechanistic plausibility from being overstated as clinical efficacy.
At the molecular level, the review separates two overlapping mechanisms. First, peroxide-containing chemotypes and related natural products may alter reactive oxygen species through radical chemistry, redox buffering, or stress-response pathways. Second, they may reset inflammatory transcription by modulating redox-sensitive regulators such as NF-kappaB, AP-1/MAPK, STAT proteins, NRF2-linked antioxidant programs, and inflammasome-associated signaling. This distinction is clinically relevant because reduced oxidative injury does not automatically imply suppression of thyroid chemokine production. Recent human serum data connect this molecular layer to patients by showing persistent NF-kappaB-related inflammatory tone in Hashimoto thyroiditis despite thyroxine replacement.33 RCT meta-analysis evidence that selenium lowers TSH, TPOAb, and malondialdehyde also supports redox stress as a clinically measurable pathway, although not a substitute for thyroid-specific drug evidence.2
Direct thyroiditis evidence remains narrower than the broader artemisinin literature. AIT/EAT studies mainly link dihydroartemisinin (DHA) to chemokine-axis modulation in experimental disease.34 In EAT, DHA reduced thyroid inflammation, serum and tissue CXCL10, CXCL10-CXCR3-associated signaling, and NF-kappaB/p65 activation.28 Newer EAT evidence adds a thyroiditis-specific outcome layer: high-dose DHA reduced thyroglobulin antibody levels and thyroid inflammation while increasing Nrf2-linked antioxidant capacity, connecting antibody-linked disease activity with immune balance and oxidative-stress modulation.35 In non-thyroid autoimmune models, SM934 and SM905 have been associated with reduced inflammatory cytokines, oxidative stress, Th1/Th17 responses, nitric oxide production, and MAPK/NF-kappaB signaling.36,37 These data justify mechanistic exploration but do not establish class-wide efficacy in AIT.
The review is therefore organized to keep clinical context, mechanism, and evidence provenance aligned. First, it summarizes chemokine-receptor architecture in AIT and highlights where patient cohorts support phenotype-linked chemokine changes. Second, it maps these circuits onto intrathyroidal niches, including Th1 fields, myeloid-rich inflammatory belts, and TLS-like B-cell/T-cell zones. Third, it compares artemisinin chemotypes by upstream innate and redox pathways, while separating direct AIT/EAT evidence from supportive non-thyroid data (Table 1). Supplementary evidence tables are used to keep compound-level details, model provenance, candidate targets, and spatial predictions accessible without overloading the Introduction.
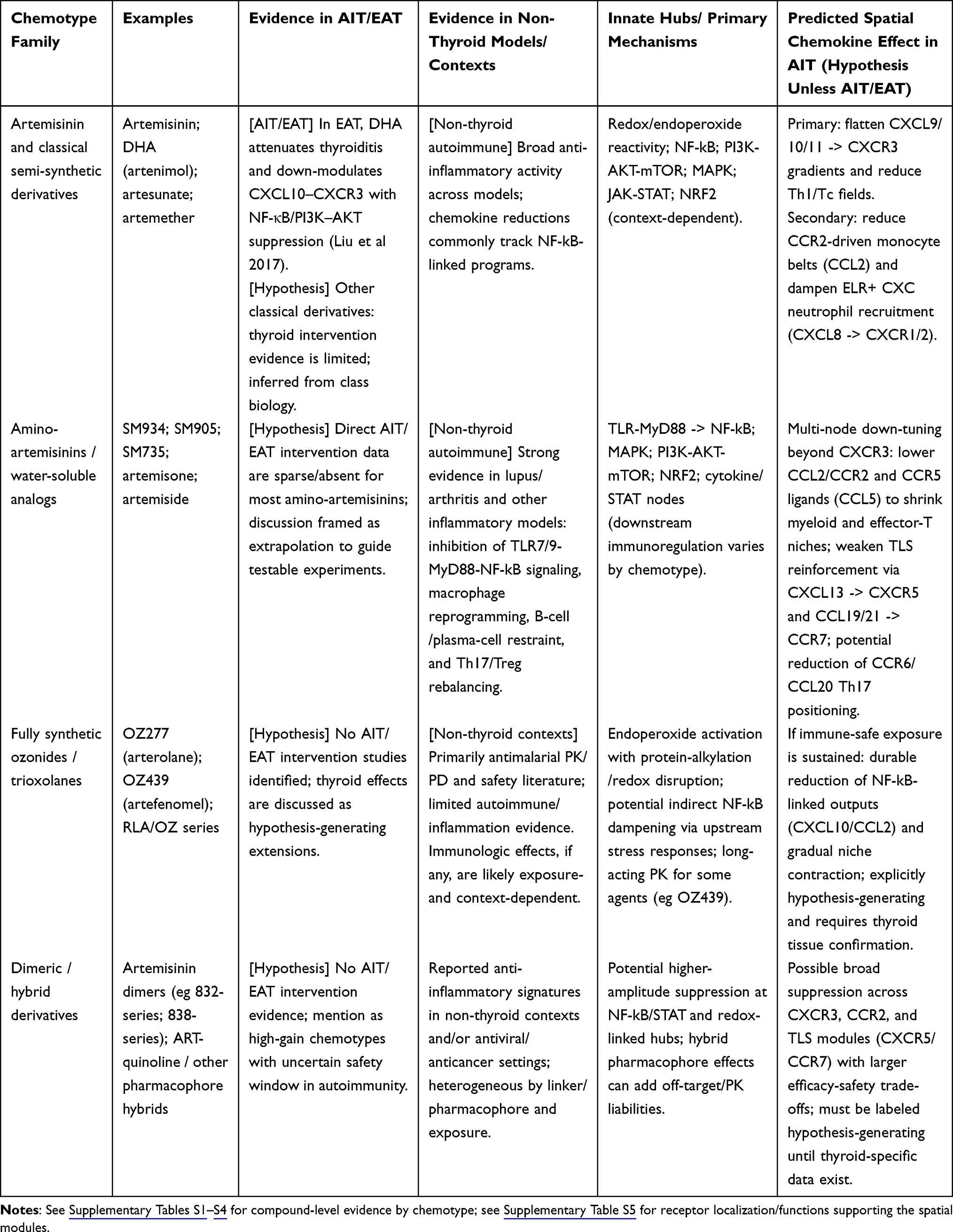 |
Table 1 Chemotype Snapshot and Evidentiary Boundary (Autoimmune Thyroiditis/Experimental Autoimmune Thyroiditis (AIT/EAT) vs Extrapolation) |
Four knowledge gaps motivate this review. First, thyroiditis studies still emphasize CXCL10-CXCR3 while giving less attention to TLS-relevant and myeloid-associated nodes such as CCL2/CCR2, CCL5/CCR5, CXCL8/CXCR1/2, CXCL13/CXCR5, and CCL19/21-CCR7.38 Second, spatial gradients across follicles, vessels, stromal regions, and TLS-like aggregates remain incompletely measured. Third, innate-pathway changes involving NF-kappaB, MAPK, PI3K-AKT-mTOR, NRF2/GPX4, and TLR-MyD88 are seldom quantified together with thyroid chemokine outputs.39–41 Fourth, chemokine readouts are rarely paired with patient-relevant endpoints such as TSH/FT4, TPOAb/TgAb, ultrasound echogenicity or volume, symptoms, levothyroxine need, and serum CXCL9/CXCL10/CXCL11/CCL2. These gaps matter because a treatment-relevant network model must explain not only pathway changes, but also whether those changes track endocrine function, antibody dynamics, imaging features, and tissue organization. This narrative review therefore asks whether artemisinin-related compounds may function as chemotype-defined modulators of innate signaling and spatial chemokine-network remodeling in AIT. The proposed triad--chemotype -> innate reprogramming -> spatial chemokine remodeling--is a testable framework, not an established pharmacodynamic account. Direct testing is needed to measure shifts in CXCR3+ Th1 fields, CCR2+/CX3CR1+ myeloid belts, and CXCR5+/CCR7+ TLS cores using spatially resolved methods.42–44
Overall, AIT is best viewed as a clinically measurable chemokine-network disease rather than a single-pathway disorder. This framework narrows the review question to a practical translational test: whether artemisinin-related compounds can shift thyroid-relevant immune niches in ways that correspond to antibody burden, thyroid function, ultrasound remodeling, and tissue chemokine architecture.
Scope and Evidence Framework
This article is a narrative review, not a systematic review or meta-analysis. Searches were targeted and non-systematic, using term combinations for AIT, Hashimoto thyroiditis, experimental AIT, chemokines, tertiary lymphoid structures, spatial transcriptomics, artemisinin, DHA, artesunate, amino-artemisinins, ozonides, and trioxolanes. Backward and forward citation tracking was used to identify additional mechanistic and disease-relevant publications. The absence of formal risk-of-bias assessment and quantitative synthesis is an important limitation and is considered when interpreting the evidence.
For scientific rigor, claims are interpreted at three evidence levels. “Direct AIT/EAT evidence” refers to findings in patients, thyroid tissue, primary thyroid systems, or experimental AIT models. “Supportive non-thyroid evidence” refers to data from other autoimmune or inflammatory systems that may inform mechanisms but do not establish thyroid relevance. “Hypothesis” refers to biologically plausible inferences that have not been tested in thyroiditis. These distinctions are essential because the field can otherwise overstate conclusions by merging mechanistic plausibility, animal data, and clinical applicability into a single therapeutic narrative (Table 2).
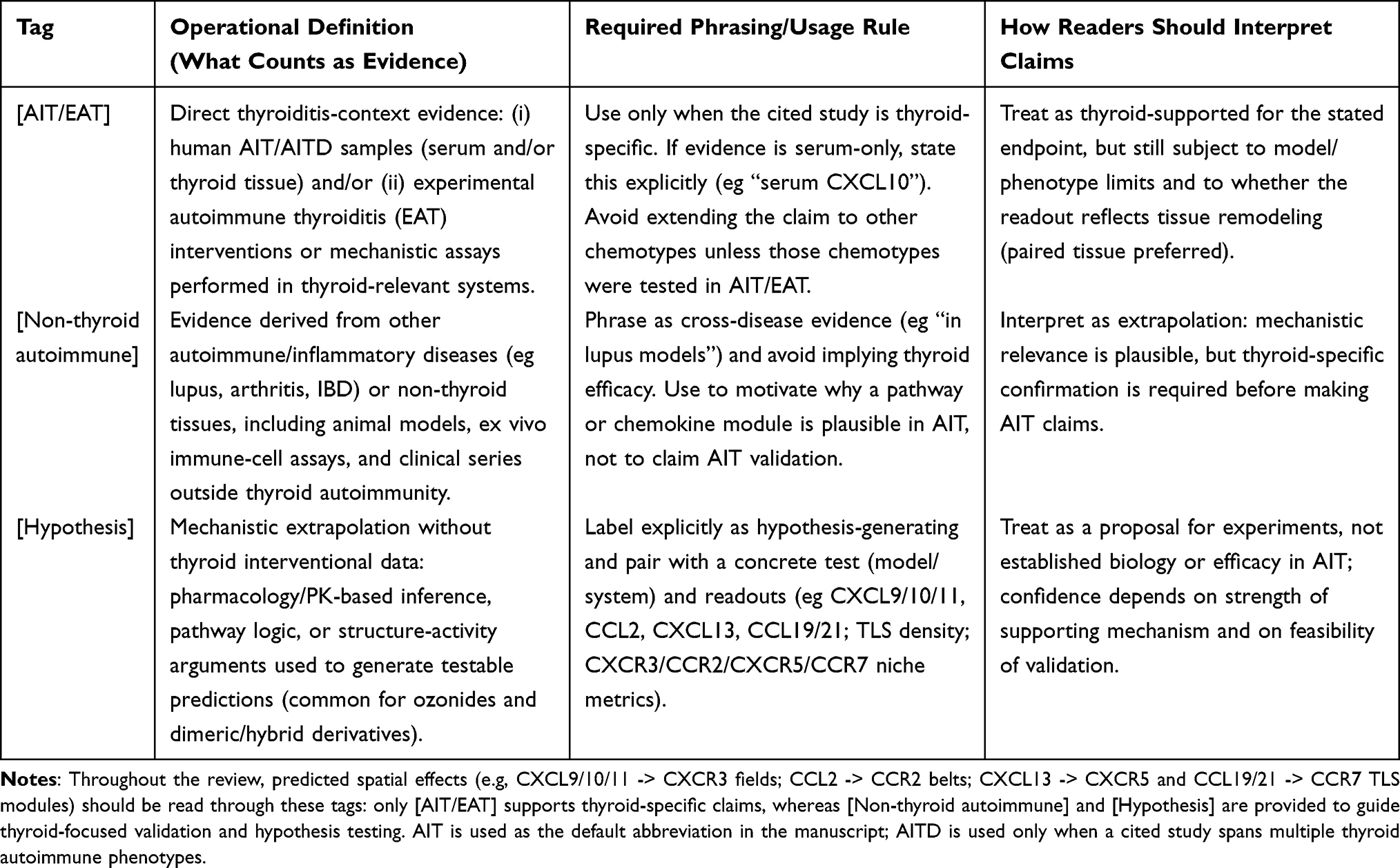 |
Table 2 Evidence-Tag Legend Used Throughout the Manuscript |
Mechanistic statements are also separated into cellular, biochemical, and epigenetic levels. Cellular statements refer to changes in T cells, B cells, macrophages, dendritic cells, thyrocytes, or stromal compartments. Biochemical statements refer to peroxide activation, ROS handling, kinase phosphorylation, receptor-proximal signaling, transcription-factor activity, and antioxidant pathway engagement. Epigenetic statements refer to proposed changes in chromatin accessibility, histone marks, DNA methylation, or trained-immunity-like memory; these are treated as hypotheses unless directly measured.
The goal is therefore not to claim clinical efficacy, but to map the current evidence, define mechanistic boundaries, and specify the experiments required before translational claims can be made. A second boundary concerns disease definition. Autoimmune thyroid disease (AITD) includes Graves disease and Hashimoto thyroiditis, whereas this manuscript centers on destructive AIT. Graves disease studies are cited only when they clarify shared chemokine biology across thyroid autoimmunity and are not interpreted as direct efficacy evidence for AIT.
Artemisinin Chemotypes: Evidence Boundaries and Candidate Molecular Mechanisms in AIT
Across the four chemotype families summarized in Supplementary Tables S1–S4, artemisinin-related compounds may influence upstream innate signaling hubs relevant to chemokine production.14,17,21,40,41,45 However, most evidence for these effects comes from non-thyroid systems. Selected compounds can be provisionally viewed as candidate network modulators rather than selective CXCL10 antagonists, but direct thyroiditis support is limited to a small subset.21,46 Accordingly, each interpretation in this section is labeled as direct AIT/EAT evidence, supportive non-thyroid autoimmune evidence, or mechanistic extrapolation (Table 2).
For mechanistic clarity, the chemotypes are best considered along a molecular continuum rather than as a single pharmacologic class. Classical semi-synthetic artemisinins provide relatively short peroxide-driven redox pulses; amino-artemisinins add side-chain-dependent solubility and immune-cell exposure; ozonides and trioxolanes extend peroxide pharmacokinetics; and dimers or hybrids may increase target engagement while also increasing off-target risk. The relevant question for AIT is not simply whether these compounds are anti-inflammatory, but whether they reproducibly alter thyroid-relevant chemokine transcription, receptor signaling, antigen-presentation programs, and oxidative-stress responses at achievable and safe concentrations.
Classical SSA: Thyroid-Specific Evidence Centered on DHA and CXCR3-Axis Modulation
Supplementary Table S1 summarizes classical or clinically used SSA, including artemisinin, DHA, artesunate, artemether, arteether, artelinic acid, and selected DHA ester prodrugs. The literature suggests class-level effects on NF-κB-regulated inflammatory mediators, including CXCL10, CCL2, and CCL20, but these effects vary by compound, model, and exposure context.37,45,47 Consequently, SSA are not described as uniform chemokine inhibitors. They are instead treated as a pharmacologic family with overlapping but incompletely characterized immunologic profiles.
A molecular interpretation of SSA activity remains compound-specific. DHA may reduce CXCL10-associated signaling partly by lowering PI3K/AKT/NF-κB activation, whereas other SSA members may differ in cellular uptake, reduction potential, active-metabolite formation, and exposure duration. Future thyroiditis studies need to report not only histology and autoantibodies but also kinase phosphorylation, nuclear translocation of NF-κB and STAT factors, NRF2 target-gene induction, and concordance between chemokine mRNA and protein changes.
DHA provides the strongest direct thyroiditis evidence among the compounds reviewed. In a C57BL/6 EAT model, DHA reduced thyroidal and serum CXCL10, CXCR3 expression, CXCR3-positive T-cell infiltration, and histologic thyroid injury, with effects linked to inhibition of the CXCR3/PI3K/AKT/NF-κB pathway.38,45 These findings support DHA as a thyroid-relevant proof-of-concept compound, not as evidence of class-wide clinical efficacy. Extrapolation to other SSA compounds or to human AIT requires direct thyroid validation and is presented as a testable hypothesis.35,37
The tissue location of these pathways is equally important. IFN-γ-inducible CXCL9/10/11 can arise from follicular epithelial, endothelial, and stromal compartments and recruit CXCR3-positive Th1/Tc cells, whereas CCL2-rich regions may preferentially support CCR2-positive monocyte and macrophage recruitment.21,48 Therefore, any SSA effect on CXCL10 or CCL2 requires evaluation as a tissue-structured event rather than inferred only from serum biomarkers.
Amino-Artemisinins and Water-Soluble Analogs: Non-Thyroid Evidence and Thyroiditis Hypotheses
Amino-artemisinins and water-soluble immunomodulatory analogs, including artemisone, artemiside, artemisox, 11-aza-artemisinin series, WHN-11, SM934, SM905, SM735, and related compounds, have been studied mainly in non-thyroid autoimmune models such as lupus and collagen-induced arthritis (Supplementary Table S2). Their pharmacologic profiles may involve TLR–MyD88–NF-κB and PI3K–AKT signaling.34,36 The SM series is useful for hypothesis generation because these agents can reduce inflammatory cytokines and chemokines, rebalance Th1/Th17 versus Treg responses, and attenuate tissue injury in non-thyroid systems.28,49 However, the absence of direct AIT/EAT data means that thyroid chemokine effects remain unproven.
Because thyroid-specific evidence for amino-artemisinins is sparse, this subsection is intentionally framed as mechanism-informed hypothesis generation. TLS formation in AIT may depend on CXCL13–CXCR5 and CCL19/21–CCR7 signaling that organizes B-cell follicles and T-cell zones.26,34 If compounds such as SM934 influence B cells, Tfh cells, or stromal–immune crosstalk in thyroid tissue, they could plausibly alter TLS-like niches. This prediction requires direct testing in thyroid explants, EAT models, or spatially resolved human tissue studies.
At the molecular level, amino-artemisinin side chains may influence ionization state, membrane permeability, lysosomal or endosomal accumulation, and proximity to TLR-rich intracellular compartments. These properties could affect MyD88-dependent signal initiation, B-cell receptor/TLR cooperation, and macrophage cytokine programs. In AIT, however, such predictions need to be tested in primary thyroid immune-cell mixtures, organoids, or explants rather than inferred from systemic lupus or arthritis models alone.
Fully Synthetic Ozonides/Trioxolanes: Pharmacokinetic Rationale and Untested Thyroid Predictions
There is currently no direct AIT/EAT evidence for fully synthetic ozonides or trioxolanes. This subsection therefore treats these compounds as pharmacologic comparators rather than established AIT therapies. Supplementary Table S3 covers peroxide ozonides/trioxolanes such as arterolane (OZ277) and artefenomel (OZ439), which share an artemisinin-like endoperoxide pharmacophore but differ in scaffold, half-life, and exposure profile.50–53
The key molecular distinction for ozonides is exposure geometry. A longer-lived peroxide may generate a lower but more sustained redox and signaling input, whereas DHA-like compounds may produce sharper and shorter perturbations. Whether chronic low-level pathway pressure would resolve inflammation, impair host defense, or induce compensatory chemokine escape in thyroid tissue is unknown and needs to be evaluated experimentally.
The main reason to discuss ozonides is pharmacokinetic rather than thyroid-specific. Long-acting agents such as OZ439 can sustain exposure for longer periods than classical SSA.52 In theory, this could support more durable modulation of NF-κB-regulated chemokines, but in AIT this remains untested. Claims that such agents would flatten chemokine gradients, destabilize TLS, or prevent re-establishment of CXCR3-positive and CCR2-positive infiltrates are presented only as experimental predictions. CXCL10 and CCL2 may be useful pharmacodynamic readouts if these compounds are tested in thyroiditis models.54,55
Dimeric and Hybrid Artemisinins: Experimental Chemotypes with Uncertain Thyroid Relevance
Dimeric and hybrid artemisinins currently have no direct AIT/EAT evidence. They are included only to define how future structure–activity studies might probe chemokine-network sensitivity. Supplementary Table S4 summarizes Posner-series dimers, ART–quinoline and ART–isoquinoline hybrids, organometallic hybrids, and photopharmacology-related structures. These compounds were largely optimized for antiviral, antimalarial, or anticancer applications.56,57 Their potency and prolonged target engagement may be immunologically informative, but thyroid relevance cannot be assumed.
From a molecular-biology perspective, dimeric and hybrid compounds may help determine whether stronger peroxide burden, altered subcellular localization, or hybrid target engagement changes chemokine transcription more profoundly than monomeric SSA. Their inclusion in this review does not imply clinical readiness. Rather, it identifies a structure–mechanism space that could be screened in thyroid-relevant inflammatory assays before animal or human translation is considered.
For dimeric and hybrid derivatives, claims of chemokine-network remodeling require particular caution. Supplementary Table S4 lists class-level reductions in pro-inflammatory mediators through NF-κB-related mechanisms, but translation to AIT would require direct evidence for effects on CXCL10, CCL2, CCL5, CCL20, CXCL13, CCL19/21, or CX3CL1 in thyroid tissue. These compounds may also raise greater safety, dose-selection, and off-target concerns than shorter-acting SSA. At present, they are best considered research tools or distant leads rather than AIT-ready therapeutic candidates.
A Hypothesis-Generating Innate–Spatial Network Model
The proposed model links innate hubs, chemokine circuits, and chemokine-defined spatial niches to organize available evidence and identify testable gaps. Supplementary Tables S1–S4 group compounds by chemotype family and compare their putative effects on NF-κB, PI3K–AKT–mTOR, JAK–STAT, TLR–MyD88, MAPK, and redox programs. The model assumes that these pathways shape the inflammatory set point of thyrocytes, macrophages, dendritic cells, and stromal compartments. This “set-point” terminology is conceptual and needs to be validated using pathway-activity assays rather than inferred from downstream cytokine changes alone.
Across autoimmune and inflammatory models, artemisinin derivatives have been reported to modulate NF-κB, JAK–STAT, PI3K–AKT–mTOR, and NRF2/GPX4-linked redox programs, with accompanying reductions in inflammatory mediators.58 In thyroiditis models, the clearest example is DHA-mediated reduction of CXCL10–CXCR3-associated PI3K/AKT/NF-κB signaling. This provides a direct AIT-linked example of innate-to-chemokine coupling, but it does not demonstrate that all chemotypes act through the same mechanism in thyroid tissue.
On the chemokine side, this review deliberately rejects a reductionist “CXCL10 disease” model. AIT and related thyroid autoimmunity involve multiple axes, including CXCL9/10/11–CXCR3, CCL2–CCR2, CCL5–CCR5, CCL20–CCR6, CXCL12–CXCR4/ACKR3, CXCL13–CXCR5, CCL19/CCL21–CCR7, and CX3CL1–CX3CR1. Clinical and experimental data support upregulation of IFN-γ-inducible CXCR3 ligands in thyroid tissue and circulation, with CXCL10 produced by thyrocytes and endothelial cells contributing to Th1-biased infiltration.59–61
These observations support a spatially structured, multi-node view of AIT. CXCL9/10/11 and CXCL12 may be produced by thyroid follicular, endothelial, and stromal cells; CCL2, CCL5, CCL20, and CX3CL1 may arise from epithelial, fibroblast, endothelial, and myeloid compartments. Receptors are probably distributed across partially overlapping niches: CXCR3 and CCR5 on Th1/Tc1 and NK cells; CCR2 and CX3CR1 on monocytes, macrophages, and myeloid dendritic cells; and CXCR4, CXCR5, and CCR7 on B cells, Tfh/Tfr cells, and central-memory or naïve T cells within lymphoid aggregates. This framework defines measurable tissue endpoints but are not read as a completed spatial atlas of AIT.
Within this framework, chemotype families can be compared along three cautious axes: depth of innate-pathway modulation, duration of exposure, and putative cell or niche selectivity. What is known comes mainly from pharmacology and non-thyroid immunology; what is predicted in AIT remains to be tested. A stronger or longer signal might, in principle, down-tune several chemokine nodes and reduce CXCR3-positive T-cell fields, CCR2/CX3CR1-positive myeloid belts, or CXCR5/CCR7-positive TLS cores. These are measurable endpoints for future work, not claims of demonstrated clinical benefit.
The model is useful only if it is falsifiable. In human AIT tissue, thyroid explants, or EAT models exposed to defined chemotypes, single-cell RNA-seq, spatial transcriptomics, and multiplex imaging are needed to test coordinated shifts in innate pathway signatures, chemokine gene and protein expression, and the geometry of CXCR3-positive, CCR2/CX3CR1-positive, and CXCR5/CCR7-positive niches. Biomarker strategies therefore include tissue and serum readouts rather than relying solely on circulating CXCL10.
Figure 1 presents a selected chemokine–receptor axis map in the AIT context, centered on CXCR3 rather than a global receptor-overlap chemokine network. The CXCL10–CXCR3 axis is highlighted as the strongest experimentally supported DHA-sensitive AIT axis, while CXCL9/CXCL11–CXCR3 and non-CXCR3 axes are shown as class-level, contextual, or validation-needed hypotheses.14 Overlaying artemisinin derivative classes onto this chemokine–chemokine network illustrates possible points of intersection rather than proven targets. SSA map to several AIT-associated hubs, whereas AAWS, PO/T, and D/HA expand the hypothesis space to additional inflammatory and lymphoid-organization nodes. At present, the strongest direct thyroiditis evidence remains DHA suppression of the CXCR3/PI3K/AKT/NF-κB pathway in EAT; evidence for SM905, SM934, and other chemotypes largely comes from non-thyroid systems.36,62 Accordingly, Figure 1 is read as a translational hypothesis map with explicit evidence gradients.
 |
Figure 1 Artemisinin and endoperoxide-class evidence mapped onto selected chemokine–receptor axes in autoimmune thyroiditis (AIT). Chemokine ligands are shown in boxed nodes and receptors in ovals; the CXCL10–CXCR3 axis is highlighted as the main experimentally supported AIT axis for DHA, while CXCL9/CXCL11–CXCR3 and non-CXCR3 axes are shown as class-level, contextual, indirect, or validation-needed evidence. Tags denote: SSA, semi-synthetic artemisinins; AAWS, water-soluble artemisinin analogs; PO/T, synthetic endoperoxides not standard artemisinin derivatives; and DHA, DHA-specific experimental AIT evidence. Navy lines indicate AIT-supported canonical axes, gray lines established ligand–receptor interactions, and dashed lines inferred or co-occurrence-based connections; symbols indicate experimental AIT evidence, indirect evidence, or need for validation. Colored tags summarize evidence context and do not prove direct regulation of every chemokine–receptor axis in human AIT. |
Chemokines and Receptor Networks
Figure 2 summarizes the AIT chemokine–receptor landscape as an interconnected ligand–receptor circuit. The schematic emphasizes key AIT-supported axes, particularly CXCL9/CXCL10/CXCL11–CXCR3 and CCL19/CCL21–CCR7, with additional inflammatory or lymphoid-organizing pathways involving CCL2–CCR2, CCL5–CCR5, and CXCL13.14 Thick dark-blue lines indicate AIT-supported axes, gray lines indicate established chemokine–receptor interactions, and dashed lines indicate indirect artemisinin-class modulation through upstream inflammatory pathways.36,63–66 Artemisinin-class tags indicate reported or inferred anti-inflammatory chemotype associations and do not prove direct modulation of every axis in human AIT.29,40
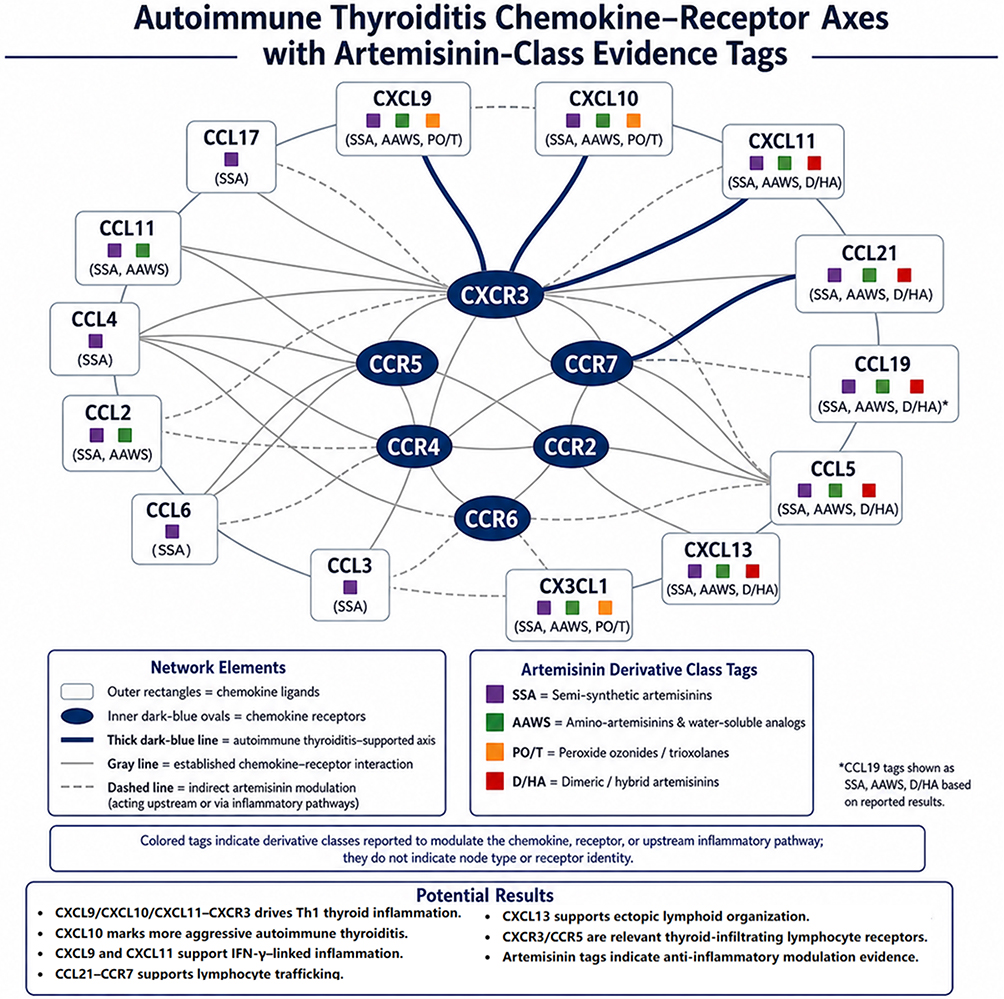 |
Figure 2 Autoimmune thyroiditis chemokine–receptor axes annotated with artemisinin-class evidence tags. Outer rectangles represent chemokine ligands, inner dark-blue ovals represent chemokine receptors, gray lines indicate established ligand–receptor interactions, thick dark-blue lines highlight autoimmune thyroiditis-supported axes, and dashed lines indicate indirect artemisinin-class modulation through upstream inflammatory pathways. The schematic emphasizes CXCL9/CXCL10/CXCL11–CXCR3 and CCL19/CCL21–CCR7 signaling, with additional inflammatory or lymphoid-organizing ligands including CCL2, CCL5, CXCL13, and CX3CL1. Colored tags denote artemisinin derivative classes with reported or inferred modulation evidence and do not imply proven direct regulation or established therapeutic efficacy in human autoimmune thyroiditis. Evidence supports the CXCR3 axis and CCL21/CCR7 involvement in thyroid autoimmunity, while artemisinin assignments should remain provisional pending thyroid-specific validation. |
Chemokine Receptor Functions in AIT
Spatial Distributions of Chemokine Receptors in Thyroid
Figure 3 projects the receptor network into thyroid space by depicting receptor distributions across perivascular, interfollicular, follicular, and TLS-like niches. The map includes CXCR3, CCR2, CCR5, CCR6, CCR7, CXCR5, CXCR1, CXCR2, CXCR4, ACKR3, CX3CR1, XCR1, ACKR1, ACKR2, and ACKR4. It emphasizes widespread CXCR3-rich inflammatory territories, stromal and interstitial CCR2/CCR5 regions, CCR7/CXCR5-enriched lymphoid domains, CXCR1/2-positive neutrophil-associated areas, central CXCR4/ACKR3 niches, CX3CR1-rich microvascular belts, XCR1-positive cDC1-like foci, and atypical receptor scavenger zones. Together with Supplementary Table S5, the schematic argues that receptor location may influence drug response. Direct evidence that artemisinin chemotypes remodel these niches in human AIT is not yet available.35
 |
Figure 3 Spatial mosaic of chemokine receptor distributions in autoimmune thyroiditis (AIT). The schematic thyroid template depicts receptor-enriched niches summarized in Supplementary Table S5: CXCR3-rich Th1-skewed infiltrates; CCR2-positive stromal and interfollicular monocyte/macrophage zones; CCR5-positive perivascular and interstitial effector T-cell regions; CCR6-positive Th17-associated areas; CCR7- and CXCR5-enriched lymphoid/TLS-like domains; CXCR1/2-positive neutrophil-associated areas; CXCR4/ACKR3 niches; CX3CR1-rich microvascular belts; XCR1-positive cDC1-like foci; and ACKR1/2/3/4 scavenger zones. The distribution is conceptual and should be validated by spatial transcriptomics, multiplex proteomics, or targeted histology in thyroid tissue. |
Associated Cell Types and Functions in AIT
Supplementary Table S5 organizes AIT chemokine receptors into a spatial and functional map. CXCR3, CCR2, and CCR5 dominate the classic Th1/monocyte axis. CXCR3-positive Th1/Tc and NK cells infiltrate interstitium and perivascular cuffs in response to thyrocyte-derived CXCL9/10/11, while CCR2-positive monocyte/macrophage precursors accumulate in stromal and interfollicular regions under CCL2. CCR5-positive effector T cells can cluster in perivascular and interstitial aggregates driven by CCL3/4/5.3,67,68 These axes plausibly sustain IFN-γ/TNF-α feedback, antigen presentation, and chronic inflammation.
Chemokine receptors also provide a molecular bridge between the network diagrams and cell behavior. Most canonical receptors are G protein-coupled receptors that activate Gαi-dependent signaling, phospholipase C, intracellular calcium flux, PI3K, small GTPases, integrin activation, and cytoskeletal remodeling. Thus, a chemokine axis does not merely label a cell type; it encodes receptor-proximal signals that influence adhesion, migration, retention, survival, and local cytokine production. Atypical chemokine receptors add another regulatory layer by shaping ligand availability and gradients without always producing classical migration signals.
A second receptor layer includes myeloid, endothelial, lymphoid-retention, and homeostatic circuits. CXCR1/2 ligands recruit neutrophil-associated cells; CXCR4 and ACKR3 respond to CXCL12 and may retain lymphocytes or plasma cells within follicles and TLS-like structures; and CX3CR1–CX3CL1 links cytotoxic T cells, NK cells, and inflammatory monocytes to activated endothelium.69 XCR1–XCL1/2 may position cross-presenting cDC1 in perivascular or interstitial niches. These pathways are important comparators when evaluating whether artemisinin effects extend beyond CXCL10.
Innate Reprogramming: Molecular Mechanisms
Peroxide Activation, Iron Biology, Redox Signaling, and Scavenging Across Chemotype Families
All artemisinin chemotypes contain an endoperoxide bridge that can be activated by ferrous iron or heme, producing short-lived radicals and reactive intermediates. In malaria parasites, this chemistry occurs largely in the digestive vacuole and damages multiple parasite proteins.70 In mammalian immune cells and thyrocytes, the same chemical principle is more likely to affect redox-sensitive signaling than to produce parasite-like toxicity. This distinction is essential when extrapolating antimalarial pharmacology to autoimmune disease.
In autoimmune settings, “antioxidant” is not used as a single mechanistic label. Direct ROS scavenging, reduction of lipid peroxidation, preservation of glutathione pools, activation of NRF2-dependent antioxidant genes, and inhibition of ROS-amplified NF-κB or inflammasome signaling are biologically distinct events. A thyroiditis study that measures only total ROS or a single antioxidant enzyme cannot determine which mechanism is dominant. Future work should therefore pair oxidative-stress readouts with pathway-specific assays, including NRF2 nuclear localization, Keap1/NRF2 target transcription, GPX4 activity, lipid-peroxidation markers, and phosphorylation states of redox-sensitive kinases.
Chemotype families differ along the peroxide–iron–signaling axis. Classical SSA such as ART, DHA, artesunate, artemether, and arteether are relatively short-lived and may generate transient redox signals. In thyroiditis models, DHA interrupts CXCL10–CXCR3-associated signaling and suppresses the CXCR3/PI3K/AKT/NF-κB cascade, linking peroxide chemistry to chemokine output in one thyroid-specific setting. Amino-artemisinins may improve solubility, cellular entry, and exposure; ozonides and dimers may extend exposure or alter tissue distribution. These differences can be tested as molecular pharmacology rather than presented as compound lists. Dose–response designs need to measure intracellular exposure, labile iron dependence, ROS kinetics, mitochondrial stress, glutathione balance, and reversibility after washout, because beneficial immunomodulation and nonspecific cytotoxicity can otherwise appear similar in bulk inflammatory readouts.
Innate Signaling Circuits and Epigenetic Interfaces: NF-κB, MAPK, PI3K–AKT–mTOR, JAK–STAT, NRF2/GPX4, and TLR Crosstalk
A recurring pattern across non-thyroid immune models is modulation of NF-κB, MAPK, PI3K–AKT–mTOR, and JAK–STAT signaling in macrophages or dendritic cells.71 In AIT-specific work, DHA reduces phosphorylation of PI3K, AKT, and NF-κB/p65 in thyroid tissue, paralleling lower CXCL10 and fewer CXCR3-positive infiltrating T cells.35 Other models suggest effects on NRF2–GPX4 antioxidant pathways, inflammasome activity, and Th1/Th17 versus Treg balance. These hubs sit upstream of several chemokines, including CXCL9/10/11, CCL2, CCL5, CCL20, CXCL8, CXCL13, and CCL19/21.72
A more molecular description of these pathways clarifies the testable links to chemokine output. NF-κB and AP-1 can directly drive transcription of inflammatory cytokines and chemokines; STAT1/STAT3 signaling can connect IFN-γ or IL-6-family cues to Th1/Th17-associated programs; PI3K–AKT–mTOR influences metabolic fitness, survival, and cytokine translation; and NRF2/GPX4-related programs can reduce lipid-peroxidation stress that otherwise amplifies inflammatory transcription. Therefore, the same compound could lower CXCL10 by reducing upstream IFN/NF-κB signaling, altering cellular redox tone, changing metabolic activation, or reducing viability. These alternatives require experimental separation.
TLR crosstalk is plausible but incompletely validated. Several artemisinin derivatives have been reported to dampen TLR4 or TLR7/9 signaling, thereby reducing MyD88-dependent NF-κB and MAPK activation in monocytes, macrophages, B cells, or dendritic cells.73 Whether this produces trained-immunity-like reprogramming in AIT is unknown. Current claims about persistent epigenetic effects in thyroiditis are therefore framed as hypotheses requiring ATAC-seq, histone-mark profiling, or longitudinal cell-state assays.74
Epigenetic modulation is important but underdeveloped. Natural compounds can influence chromatin indirectly by changing cellular redox state, acetyl-CoA and NAD+/sirtuin availability, DNA methylation pressure, histone acetylation or methylation, and the accessibility of NF-κB/IRF/STAT-bound enhancers. In AIT, this would matter if monocytes, dendritic cells, B cells, or thyrocytes retained a lower chemokine-producing state after drug removal. Such persistence is not assumed; this hypothesis requires testing with time-course washout experiments, ATAC-seq, histone-mark profiling, DNA-methylation analysis, and single-cell trajectory methods.
SM934, SM905, and SM735 as Non-Thyroid Exemplars of Innate Reprogramming
Among amino-artemisinins, SM934, SM905, and SM735 provide useful examples of immune reprogramming, but most evidence comes from non-thyroid models. SM934 has shown activity in systemic lupus erythematosus and related autoimmune systems by influencing TLR/MyD88/NF-κB signaling, B-cell activation, plasma-cell differentiation, autoantibody production, Th1/Th17 responses, and Foxp3-positive regulatory T cells. At the molecular level, this profile implies possible effects on B-cell transcriptional activation, Tfh/Treg differentiation cues, cytokine feedback loops, and chemokine programs that support ectopic lymphoid organization. In AIT, these mechanisms map conceptually onto CXCL13–CXCR5 and CCL19/21–CCR7 lymphoid niches, but direct evidence that SM934 modifies intrathyroidal TLS, germinal-center-like reactions, or autoantibody-generating structures is lacking.
SM905 illustrates macrophage-centered modulation. In RAW264.7 macrophages, SM905 inhibits nitric oxide and pro-inflammatory cytokines by suppressing p38/JNK/ERK phosphorylation and NF-κB activation. If similar effects occurred in thyroid tissue, they might reduce CCL2, CCL5, or CXCL8 output and alter CCR2-positive monocyte recruitment, CCR5-positive effector T-cell clustering, or CXCR1/2-dependent neutrophil influx. SM735 also suppresses lymphocyte proliferation and cytokine production in non-thyroid systems.28,45 These predictions require direct AIT testing.
Together, these amino-artemisinin examples highlight the need to integrate cellular and molecular endpoints. A reduction in macrophage cytokines remains mechanistically incomplete unless accompanied by evidence of MAPK/NF-κB target suppression, preserved cell viability, altered chromatin accessibility at inflammatory loci, and reduced chemokine production in a thyroid-relevant microenvironment. Similarly, Treg expansion gains mechanistic value when linked to transcriptional programs, cytokine milieu, and localization within thyroid tissue rather than reported only as a peripheral immune shift.
Spatial Transcriptomics and Proteomics of Thyroid Tissue: Reading Chemokine Niches
Spatial transcriptomics and proteomics can reduce over-extrapolation by testing whether proposed chemokine niches actually exist in thyroid tissue. In chronic inflammatory diseases, high-plex imaging and spatial-omics show that chemokines and receptors can be deployed in discrete neighborhoods containing epithelial, fibroblast, endothelial, and leukocyte populations.75 Comprehensive maps of AIT chemokine circuits remain limited, but the available thyroid and TLS literature supports spatial organization as a biologically plausible framework.
Future AIT studies should pair artemisinin exposure with spatial readouts rather than rely only on bulk cytokines. Spatial transcriptomics could quantify how DHA or candidate compounds alter CXCL9/10/11, CCL2, CCL5, CXCL13, and CCL19/21 at follicular borders, perivascular cuffs, and TLS-like aggregates.76 Multiplex proteomic imaging can add receptor expression, activation markers, cell proximity, and niche composition.77 These approaches would distinguish thyroid-specific effects from generalized anti-inflammatory activity.
Translational Considerations for AIT and Related Autoimmunity
Potential Use Cases and Competing Strategies
Among artemisinin-related compounds, short-acting SSA such as DHA currently have the most direct preclinical support in thyroiditis models, but the evidence does not define them as established therapies.37,78 In EAT, DHA affects CXCL10–CXCR3-associated signaling, autoantibodies, T-cell polarization, and histology. Non-thyroid data for SM934-like compounds suggest broader immune rebalancing. Any future AIT development should compare these candidates with existing supportive care, thyroid hormone replacement when indicated, risk-factor management, and other investigational anti-inflammatory or immune-modulating strategies. Without comparative data, artemisinin derivatives should not be prioritized over alternative approaches.
Mechanism-based comparison also includes non-artemisinin strategies. Approaches targeting cytokine signaling, JAK/STAT activation, B-cell/Tfh biology, oxidative stress, gut–immune interactions, or micronutrient insufficiency may influence overlapping inflammatory programs. Artemisinin derivatives would be most compelling if they demonstrate a distinct molecular signature, such as coordinated redox normalization and chemokine-network modulation, rather than a nonspecific decrease in inflammatory markers.
Dosing Horizons and Chemokine-Network Hysteresis
Pharmacokinetics may determine how chemotypes are tested, but dosing concepts remain speculative for AIT. Classical artemisinins and SSA often have short elimination half-lives and may produce brief exposure pulses, whereas artefenomel (OZ439) was engineered as a longer-acting peroxide.52 Short exposure might be useful for acute pharmacodynamic testing, while longer exposure could assess sustained network modulation. Both approaches require thyroid-specific dose–response, safety, and reversibility studies before clinical inference is justified.
Safety and Drug–Drug Interaction Considerations
Safety is central because AIT is usually chronic and often affects patients who require long-term follow-up rather than short-course anti-infective treatment. Pharmacogenomic resources indicate that artemisinin, artemether, and DHA can induce CYP3A activity and up-regulate CYP2B6 and CYP2C19.79 This raises the possibility of drug–drug interactions with CYP-metabolized immunomodulators, endocrine medications, or other chronic therapies. Conversely, Artemisia annua extracts and some formulations may inhibit CYP2B6 or CYP3A4, underscoring the need to distinguish purified chemotypes from botanical products.80,81
Reproductive safety is also important. Artemisinin-based combination therapies have established malaria-related use in later pregnancy, whereas first-trimester use has historically been approached cautiously. Autoimmune indications differ from malaria because the risk–benefit balance is usually less urgent. Until AIT-specific safety data exist, pregnancy, preconception, and lactation populations are excluded from early trials or studied only under highly controlled protocols with reproductive monitoring, contraception requirements, and independent safety review.
Translational Pipeline and Reporting Standards
A translational program begins with mechanism-rich preclinical and early-phase studies rather than large empirical efficacy trials. Candidate participants or samples are tightly phenotyped by thyroid function, antibody status, inflammatory activity, ultrasound or histologic features where available, and medication exposure. When tissue is obtainable through clinically indicated procedures, paired blood–tissue sampling connects systemic biomarkers to intrathyroidal mechanisms. Preclinical work includes primary thyrocytes, thyroid stromal cells, immune-cell co-cultures, organoids, explants, and EAT models so that molecular target engagement can be compared across levels of biological complexity.
Because these chemotypes are hypothesized to act upstream of chemokine networks, trials prespecify composite biomarker panels rather than rely on a single CXCL10 endpoint. Candidate panels include CXCL9/10/11, CCL2, CCL5, CCL20, CXCL13, CCL19, CCL21, thyroid autoantibodies, thyroid function, and innate-pathway activity scores. Molecular pharmacodynamic endpoints include NF-κB/STAT nuclear activity, PI3K–AKT–mTOR phosphorylation, NRF2/GPX4-related antioxidant markers, lipid-peroxidation indices, inflammasome markers, and, where feasible, chromatin accessibility or histone-mark readouts. Endpoints are layered across early target engagement, intermediate tissue or imaging readouts, and clinical outcomes such as thyroid function stability, symptom burden, or progression to replacement therapy. Enrichment for CXCR3-, CXCR5-, or CCR7-high/TLS-high phenotypes may improve mechanistic sensitivity but requires validation.
These translational steps must remain cautious because the current evidence base is predominantly preclinical and centered on DHA in EAT. Broad claims of class-wide efficacy in human AIT are not justified. Repurposing also addresses antimalarial stewardship, formulation quality, CYP-mediated interactions, reproductive safety, and the distinction between purified compounds and heterogeneous botanical preparations. Collaboration among pharmacologists, endocrinologists, immunologists, molecular biologists, spatial-omics specialists, and malaria experts will be needed before clinical protocols are mature.
Discussion: Synthesis of Cellular, Molecular, and Spatial Evidence
The revised interpretation is narrower and more clinically grounded than a general pro-artemisinin narrative. Direct interventional support in thyroid disease centers on DHA in EAT, including an earlier study linking DHA to reduced thyroiditis severity, autoantibodies, Th1/Th2 imbalance, and CXCL10/CXCR3-associated PI3K/AKT/NF-κB signaling, and a more recent study reporting reduced TgAb, inflammatory infiltration, Th1/Th17-related cytokines, and oxidative stress with high-dose DHA.21,35,62,82–84 These data strengthen the thyroid-specific rationale for DHA, but they do not validate all chemotypes, botanical preparations, or clinical use in patients.
Reconciling the CXCL10 Axis with Broader Chemokine Circuits
The CXCL10/IP-10–CXCR3–Th1 paradigm remains important in AIT. Thyrocytes and stromal or endothelial cells can produce CXCL9/10/11 after IFN-γ and TNF-α stimulation, recruiting CXCR3-positive Th1/Tc cells and amplifying glandular damage.5,14,15,85 Serum CXCL10 has been associated with thyroid autoimmunity and proposed as a disease-activity marker.11,15 However, biomarker associations vary across phenotypes, and CXCL10 alone cannot explain B-cell follicle organization, TLS formation, myeloid recruitment, or spatial heterogeneity. DHA/EAT studies show that interrupting this axis can reduce experimental disease, but they do not prove that CXCL10 is sufficient as a clinical target.
The innate–spatial network framework reconciles these findings by retaining CXCL10 as a hub while placing it within a broader thyroid map. Artemisinin chemotypes may act upstream through redox-sensitive and TLR-linked hubs such as NF-κB, MAPK, PI3K–AKT–mTOR, JAK–STAT, and NRF2/GPX4.86 Multiple chemotype classes could therefore converge on overlapping chemokine signatures, but the degree of convergence in thyroid tissue remains unknown and needs to be measured directly.
The CXCL10/CXCR3 axis is among the best-supported pathways in thyroid autoimmunity. Human thyrocytes from autoimmune glands produce IP-10/CXCL10 and Mig/CXCL9 after IFN-gamma stimulation, and stimulated thyrocyte supernatants attract CXCR3-positive T cells in a manner blocked by antibodies to IP-10, Mig, or CXCR3.67 Serum CXCL10 is higher in some chronic AIT cohorts and has been associated with hypothyroidism, hypoechogenicity, and TSH.6 However, not all circulating chemokine studies show consistent relationships with thyroid function or autoantibody titers across autoimmune thyroid disease phenotypes.87 Recent reviews therefore support CXCR3 ligands as relevant, but not exclusive, pathways in thyroid autoimmunity.14
Human thyroid tissue data support a broader chemokine architecture. Autoimmune glands may show increased lymphotoxin-alpha, lymphotoxin-beta, CCL21, CXCL12, CXCL13, and CCL22, with higher CXCL12, CXCL13, and CCL22 in glands containing ectopic lymphoid follicles.83 CXCL13/CXCR5 expression also tracks with lymphocytic infiltrates and germinal centers.88 Hashimoto thyroiditis has been linked to lymphoid tissue inducer-like ILC3, CXCR5-positive PD-1-high Tfh cells, TLSs, and germinal centers.89 Transcriptomic analyses further support active tertiary lymphoid organization in highly infiltrated glands.82 Spatial transcriptomics and single-cell studies have identified antigen-presenting thyrocytes, inflammatory fibroblasts, specialized vascular areas, and heterogeneous immune-cell subsets in Hashimoto tissue.21,84 Together, these findings make a network model more defensible than a single-axis model.
Therapeutic evidence remains much narrower than tissue biology. In the first thyroid-focused interventional study, DHA reduced EAT severity, anti-thyroglobulin and anti-thyroid peroxidase antibodies, Th1/Th2 imbalance, lymphocyte proliferation, and CXCL10/CXCR3-linked PI3K/AKT/NF-κB signaling.62 A later EAT study reported that high-dose DHA reduced TgAb and inflammatory infiltration, downregulated IFN-gamma, IL-17A, and IL-6, increased IL-4 and IL-10, and improved antioxidant readouts in thyroid tissue.35 These findings justify replication and mechanistic extension but do not establish that all artemisinin chemotypes remodel TLS-associated circuits or that serum markers reflect tissue architecture. Future studies should measure CXCL10 together with CXCL13, CCL21, histology, and spatial or single-cell readouts where possible.21,82,83,88
Strengths, Limitations, and Alternative Perspectives
A strength of this review is its explicit integration of interventional EAT data with human thyroid tissue observations while separating direct evidence from inference. It links the well-studied CXCL10/CXCR3 loop in thyrocytes and serum studies6,14,67 with independent evidence for CXCL13/CCL21-rich ectopic lymphoid organization and tertiary lymphoid structures.82,83,88,89 It also incorporates newer spatial and single-cell thyroid studies showing antigen-presenting thyrocytes, inflammatory stromal compartments, vascular niches, and heterogeneous immune subsets.21,84 This integrated map is the manuscript’s main contribution and distinguishes it from a generic review of artemisinin anti-inflammatory effects.
The limitations are equally important. Thyroid-specific efficacy data remain restricted to DHA in EAT, supported by two experimental studies but no robust human AIT trial.35,62 Human tissue studies defining chemokine circuits, TLS organization, and cellular heterogeneity are observational and do not test artemisinin exposure directly.21,67,82–84,88,89 Circulating chemokine associations vary by cohort and phenotype.6,87 Negative, null, or contradictory evidence is therefore not merely absent; it reflects an immature field in which many proposed effects have not yet been challenged in thyroid-specific systems. Alternative approaches, including conventional endocrine management, selenium or micronutrient studies where appropriate, cytokine/JAK/BTK-pathway modulation, and other anti-inflammatory strategies, are considered in parallel rather than treated as secondary to artemisinin development.
Next-Step Priorities: A Three-Tier Agenda
Three priorities follow from the evidence. First, independent thyroid pharmacology studies are needed to replicate or challenge DHA findings across doses, treatment windows, and comparator chemotypes, with prespecified histology, autoantibodies, T-cell polarization, oxidative-stress markers, CXCL10/CXCR3/PI3K/AKT/NF-κB, and Nrf2-related endpoints.67,83,88 Second, human mechanistic studies should use primary thyrocytes, stromal cells, thyroid organoids, or explants to test whether artemisinin derivatives suppress thyroid-relevant chemokine programs such as CXCL9/CXCL10, CXCL13, and CCL21. Third, translational studies should pair serum markers with thyroid histology, multiplex imaging, single-cell profiling, or spatial transcriptomics to determine whether circulating chemokines reflect intrathyroidal organization and disease activity.6,21,82,84,89 Early clinical work should therefore be mechanism-seeking and safety-focused, not efficacy-confirming.
Conclusions
Artemisia-derived and artemisinin-related interventions remain potential, not established, immunomodulatory candidates for AIT. The main contribution of this review is to connect three levels of evidence that are often considered separately: chemokine–receptor networks that guide immune trafficking, spatial thyroid biology that organizes inflammatory and lymphoid niches, and molecular mechanisms through which artemisinin chemotypes may influence redox tone, kinase signaling, transcription factors, and possibly epigenetic memory. The strongest thyroid-specific evidence still concerns DHA in experimental models, where reductions in inflammatory infiltration, autoantibodies, Th1/Th17-associated signaling, CXCL10/CXCR3-linked pathways, and oxidative stress have been reported. The evidence base nevertheless remains narrow, predominantly preclinical, and insufficient to support broad therapeutic claims for artemisinin derivatives as a class or for traditional botanical preparations. Key limitations include the absence of robust human AIT trials, limited thyroid-specific data for non-DHA chemotypes, inconsistent serum-chemokine associations across cohorts, and the absence of direct evidence that these compounds remodel intrathyroidal TLS or spatial chemokine niches. Future work should prioritize three linked tasks: replication and comparator testing in EAT; thyroid-specific molecular studies in primary cells, organoids, explants, or co-culture systems; and translational biomarker designs that pair serum markers with histology, multiplex imaging, single-cell profiling, spatial transcriptomics, and pathway-level pharmacodynamic readouts. Until these steps are completed, artemisinin-related compounds should be described as hypothesis-driven tools and early translational candidates rather than clinically validated therapies for AIT.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Xue H, Xu R. The lymphocyte levels of Hashimoto thyroiditis patients were significantly lower than that of healthy population. Front Endocrinol. 2025;16:1472856. doi:10.3389/fendo.2025.1472856
2. Huwiler VV, Maissen-Abgottspon S, Stanga Z, et al. Selenium supplementation in patients with Hashimoto thyroiditis: a systematic review and meta-analysis of randomized clinical trials. Thyroid. 2024;34(3):295–19. doi:10.1089/thy.2023.0556
3. Zhang Q-Y, Ye X-P, Zhou Z, et al. Lymphocyte infiltration and thyrocyte destruction are driven by stromal and immune cell components in Hashimoto’s thyroiditis. Nat Commun. 2022;13(1):775. doi:10.1038/s41467-022-28120-2
4. Zhao L, Jin S, Wang S, et al. Tertiary lymphoid structures in diseases: immune mechanisms and therapeutic advances. Signal Transduction Target Ther. 2024;9(1):225. doi:10.1038/s41392-024-01947-5
5. Rubinstein A, Kudryavtsev I, Arsentieva N, Korobova ZR, Isakov D, Totolian AA. CXCR3-expressing T cells in infections and autoimmunity. Front Biosci Landmark. 2024;29(8):301. doi:10.31083/j.fbl2908301
6. Antonelli A, Rotondi M, Fallahi P, et al. Increase of interferon-gamma inducible alpha chemokine CXCL10 but not beta chemokine CCL2 serum levels in chronic autoimmune thyroiditis. Eur J Endocrinol. 2005;152:171–177. doi:10.1530/eje.1.01847
7. Tywanek E, Michalak A, Swirska J, Zwolak A. Autoimmunity, new potential biomarkers and the thyroid gland-the perspective of Hashimoto’s thyroiditis and its treatment. Int J Mol Sci. 2024;25:4703. doi:10.3390/ijms25094703
8. Li J, Huang Q, Sun S, et al. Thyroid antibodies in Hashimoto’s thyroiditis patients are positively associated with inflammation and multiple symptoms. Sci Rep. 2024;14:27902. doi:10.1038/s41598-024-78938-7
9. Tan HL, Ge H, Qin ZE, Jiang YL, Chang S, Tang N. Evaluating the diagnostic efficiency of ultrasound and serum autoantibodies in Hashimoto’s thyroiditis: a cross-sectional study. Sci Rep. 2025;15:12674. doi:10.1038/s41598-025-97299-3
10. Wen Y, Yang Z, Rao M, Zhao L, Tang L. Histamine regulates the Th1/Th2 balance and modulates endometrial stromal autophagy through the CXCL9/CXCR3 pathway. FASEB J. 2025;39(14):e70885. doi:10.1096/fj.202403203R
11. Xue X, Wu D, Yao H, Wang K, Liu Z, Qu H. Mechanisms underlying the promotion of papillary thyroid carcinoma occurrence and progression by Hashimoto’s thyroiditis. Front Endocrinol. 2025;16:1551271. doi:10.3389/fendo.2025.1551271
12. Antonelli A, Fallahi P, Rotondi M, et al. Increased serum CXCL10 in Graves’ disease or autoimmune thyroiditis is not associated with hyper- or hypothyroidism per se, but is specifically sustained by the autoimmune, inflammatory process. Eur J Endocrinol. 2006;154:651–658. doi:10.1530/eje.1.02137
13. Antonelli A, Rotondi M, Fallahi P, et al. High levels of circulating CXC chemokine ligand 10 are associated with chronic autoimmune thyroiditis and hypothyroidism. J Clin Endocrinol Metab. 2004;89:5496–5499. doi:10.1210/jc.2004-0977
14. Ferrari SM, Paparo SR, Ragusa F, et al. Chemokines in thyroid autoimmunity. Best Pract Res Clin Endocrinol Metabol. 2023;37(2):101773. doi:10.1016/j.beem.2023.101773
15. Feng J, Xu X, Cai W, et al. Inhibiting Soluble Epoxide Hydrolase suppresses NF-κB p65 signaling and reduces CXCL10 expression as a potential therapeutic target in Hashimoto’s thyroiditis. J Clin Endocrinol Metabol. 2024;109(10):2579–2588. doi:10.1210/clinem/dgae163
16. Zheng H, Xu J, Chu Y, et al. A global regulatory network for dysregulated gene expression and abnormal metabolic signaling in immune cells in the microenvironment of graves’ disease and hashimoto’s thyroiditis. Front Immunol. 2022;13:879824. doi:10.3389/fimmu.2022.879824
17. Xu H, Lin S, Zhou Z, et al. New genetic and epigenetic insights into the chemokine system: the latest discoveries aiding progression toward precision medicine. Cell Mol Immunol. 2023;20(7):739–776.
18. Kryukova J, Vales S, Payne M, et al. Development of chemokine network inhibitors using combinatorial saturation mutagenesis. Commun Biol. 2025;8(1):549. doi:10.1038/s42003-025-07778-6
19. Antonelli A, Ferrari SM, Frascerra S, et al. Increase of circulating CXCL9 and CXCL11 associated with euthyroid or subclinically hypothyroid autoimmune thyroiditis. J Clin Endocrinol Metab. 2011;96:1859–1863. doi:10.1210/jc.2010-2905
20. Antonelli A, Ferri C, Ferrari SM, et al. High levels of circulating chemokine (C-X-C motif) ligand 11 are associated with euthyroid or subclinically hypothyroid autoimmune thyroiditis and with chemokine (C-X-C motif) ligand 10. J Interferon Cytokine Res. 2012;32:74–80. doi:10.1089/jir.2011.0051
21. Martínez-Hernández R, Sánchez de la Blanca N, Sacristán-Gómez P, et al. Unraveling the molecular architecture of autoimmune thyroid diseases at spatial resolution. Nat Commun. 2024;15(1):5895. doi:10.1038/s41467-024-50192-5
22. Liao T, Zeng Y, Xu W, et al. A spatially resolved transcriptome landscape during thyroid cancer progression. Cell Rep Med. 2025;6(4):102043. doi:10.1016/j.xcrm.2025.102043
23. Seok J, Choi H, Lee EK, et al. Tumor microenvironment remodeling across thyroid cancer differentiation states revealed by spatial transcriptomics. Cancer Immunol Immunother. 2025;74(12):357. doi:10.1007/s00262-025-04210-0
24. Yoshimitsu M, Nakamura M, Kano S, et al. CXCL13 and CCL21 induce tertiary lymphoid structures and enhance the efficacy of immunotherapy for melanoma. Cancer Sci. 2025;116:2075–2085. doi:10.1111/cas.70105
25. Zhao R, Gao D. Innate Immunity and Tertiary Lymphoid Structures. Immunolog Rev. 2025;332(1):e70052. doi:10.1111/imr.70052
26. Dong Y, Wang T, Wu H. Tertiary lymphoid structures in autoimmune diseases. Front Immunol. 2024;14:1322035. doi:10.3389/fimmu.2023.1322035
27. Gui X, Li Y, Weng Y, et al. Association of serum chemokine (C-C motif) 21 and receptor (C-C motif) 7 with Hashimoto’s thyroiditis: a preliminary clinical investigation. Front Endocrinol. 2025;16:1682553. doi:10.3389/fendo.2025.1682553
28. Gao X, Lin X, Wang Q, Chen J. Artemisinins: promising drug candidates for the treatment of autoimmune diseases. Med Res Rev. 2024;44(2):867–891. doi:10.1002/med.22001
29. Qiu F, Liu J, Mo X, Liu H, Chen Y, Dai Z. Immunoregulation by artemisinin and its derivatives: a new role for old antimalarial drugs. Front Immunol. 2021;12:751772. doi:10.3389/fimmu.2021.751772
30. Luo H, Li L, Han S, Liu T. The role of monocyte/macrophage chemokines in pathogenesis of osteoarthritis: a review. Int J Immunogenet. 2024;51(3):130–142. doi:10.1111/iji.12664
31. Yang Y, Hui C. Monoclonal antibodies to thyrotropin receptor with thyroid-stimulating activity activate the NF-κB pathway to induce chemokine expression. J Cell Mol Med. 2025;29(11):e70647. doi:10.1111/jcmm.70647
32. Ferrari SM, Ragusa F, Elia G, et al. Precision medicine in autoimmune thyroiditis and hypothyroidism. Front Pharmacol. 2021;12:750380. doi:10.3389/fphar.2021.750380
33. Yardim M, Deniz L, Saltabas MA, Celik N. Effect of thyroxine replacement therapy on serum Maresin 1 and NF-kB levels in patients with Hashimoto thyroiditis. Diagnostics. 2025;15:1248. doi:10.3390/diagnostics15101248
34. Efferth T, Oesch F. The immunosuppressive activity of artemisinin-type drugs towards inflammatory and autoimmune diseases. Med Res Rev. 2021;41(6):3023–3061. doi:10.1002/med.21842
35. Zhu S, Cui Y, Hu H, et al. Dihydroartemisinin inhibits the development of autoimmune thyroiditis by modulating oxidative stress and immune imbalance. Free Radic Biol Med. 2025;231:57–67. doi:10.1016/j.freeradbiomed.2025.02.028
36. Tong X, Chen L, He S-J, Zuo J-P. Artemisinin derivative SM934 in the treatment of autoimmune and inflammatory diseases: therapeutic effects and molecular mechanisms. Acta Pharmacolog Sinica. 2022;43(12):3055–3061. doi:10.1038/s41401-022-00978-4
37. Long Z, Xiang W, Xiao W, et al. Advances in the study of artemisinin and its derivatives for the treatment of rheumatic skeletal disorders, autoimmune inflammatory diseases, and autoimmune disorders: a comprehensive review. Front Immunol. 2024;15:1432625. doi:10.3389/fimmu.2024.1432625
38. Liu H, Tian Q, Ai X, et al. Dihydroartemisinin attenuates autoimmune thyroiditis by inhibiting the CXCR3/PI3K/AKT/NF-κB signaling pathway. Oncotarget. 2017;8(70):115028. doi:10.18632/oncotarget.22854
39. Chen L, Zhao X, Sheng R, Lazarovici P, Zheng W. Artemisinin alleviates astrocyte overactivation and neuroinflammation by modulating the IRE1/NF-κB signaling pathway in in vitro and in vivo Alzheimer’s disease models. Free Rad Biol Med. 2025;229:96–110. doi:10.1016/j.freeradbiomed.2025.01.027
40. Zhou F, Li G, Tan R, Wu G, Deng C. Artemisinins in autoimmune diseases: effects and mechanisms in systemic lupus erythematosus and rheumatoid arthritis. Brit J Pharmacol. 2025;182(15):3411–3427. doi:10.1111/bph.70086
41. Zhang L, Yu L, Deng D, et al. Artemisinin-hydroxychloroquine combination from traditional antimalarial medicine attenuates pulmonary fibrosis via PI3K/AKT/GSK3β-mediated inhibition of epithelial-mesenchymal transition. J Ethnopharmacol. 2025;355:120717. doi:10.1016/j.jep.2025.120717
42. Liu Y, Ye S-Y, He S, et al. Single-cell and spatial transcriptome analyses reveal tertiary lymphoid structures linked to tumour progression and immunotherapy response in nasopharyngeal carcinoma. Nat Commun. 2024;15(1):7713. doi:10.1038/s41467-024-52153-4
43. Inamo J, Fierkens R, Clay MR, et al. Spatial transcriptomics reveals immune-stromal crosstalk within the synovium of patients with juvenile idiopathic arthritis. JCI Insight. 2026;11(1). doi:10.1172/jci.insight.198074
44. Kondo A, McGrady M, Nallapothula D, et al. Spatial proteomics of human diabetic kidney disease, from health to class III. Diabetologia. 2024;67(9):1962–1979. doi:10.1007/s00125-024-06210-8
45. Xie K, Li Z, Zhang Y, Wu H, Zhang T, Wang W. Artemisinin and its derivatives as promising therapies for autoimmune diseases. Heliyon. 2024;10(7):1.
46. Sato Y, Silina K, van den Broek M, Hirahara K, Yanagita M. The roles of tertiary lymphoid structures in chronic diseases. Nat Rev Nephrol. 2023;19(8):525–537. doi:10.1038/s41581-023-00706-z
47. Zhu M, Wang Y, Han J, et al. Artesunate exerts organ-and tissue-protective effects by regulating oxidative stress, inflammation, autophagy, apoptosis, and fibrosis: a review of evidence and mechanisms. Antioxidants. 2024;13(6):686. doi:10.3390/antiox13060686
48. Wrońska K, Hałasa M, Szczuko M. The role of the immune system in the course of Hashimoto’s Thyroiditis: the current state of knowledge. Int J Mol Sci. 2024;25(13):6883. doi:10.3390/ijms25136883
49. Xu W-J, Wang J-S, Vijaykumar S, Arunachalam A. Immunosuppressive effect of Chinese medicine on T helper 17 cells. Chin J Integr Med. 2015;1–6. doi:10.1007/s11655-015-2073-x
50. Joshi A, Viswanathan C. Recent developments in antimalarial drug discovery. Anti Infect Agents Med Chem. 2006;5(1):105–122. doi:10.2174/187152106774755626
51. McCarthy JS, Baker M, O’Rourke P, et al. Efficacy of OZ439 (artefenomel) against early Plasmodium falciparum blood-stage malaria infection in healthy volunteers. J Antimicrob Chemother. 2016;71(9):2620–2627. doi:10.1093/jac/dkw174
52. Phyo AP, Jittamala P, Nosten FH, et al. Antimalarial activity of artefenomel (OZ439), a novel synthetic antimalarial endoperoxide, in patients with Plasmodium falciparum and Plasmodium vivax malaria: an open-label Phase 2 trial. Lancet Infect Dis. 2016;16(1):61–69. doi:10.1016/S1473-3099(15)00320-5
53. Kim HS, Hammill JT, Guy RK. Seeking the elusive long-acting ozonide: discovery of Artefenomel (OZ439). J Med Chem. 2017;60:2651–2653. doi:10.1021/acs.jmedchem.7b00299
54. Chen Y, Wang Y, Xu J, et al. Multiplex assessment of serum chemokines CCL2, CCL5, CXCL1, CXCL10, and CXCL13 following traumatic brain injury. Inflammation. 2023;46(1):244–255. doi:10.1007/s10753-022-01729-7
55. Bruschi F, Pinto B, Fallahi P, Ferrari SM, Antonelli A. Increased neutrophil derived chemokines (CXCL10 and CCL2) in human trichinellosis as possible serological markers of the polarization of the immune response against the parasite. Cytokine. 2023;166:156205. doi:10.1016/j.cyto.2023.156205
56. Marchesi E, Perrone D, Navacchia ML. Molecular hybridization as a strategy for developing artemisinin-derived anticancer candidates. Pharmaceutics. 2023;15(9):2185. doi:10.3390/pharmaceutics15092185
57. Çapcı A, Herrmann L, Sampath Kumar HM, Fröhlich T, Tsogoeva SB. Artemisinin-derived dimers from a chemical perspective. Med Res Rev. 2021;41(6):2927–2970. doi:10.1002/med.21814
58. Dai Y, Cui C, Jiao D, Zhu X. JAK/STAT signaling as a key regulator of ferroptosis: mechanisms and therapeutic potentials in cancer and diseases. Cancer Cell Int. 2025;25(1):83. doi:10.1186/s12935-025-03681-6
59. Nerviani A, Pitzalis C. Role of chemokines in ectopic lymphoid structures formation in autoimmunity and cancer. J Leukocyte Biol. 2018;104(2):333–341. doi:10.1002/JLB.3MR0218-062R
60. Marinkovic T, Garin A, Yokota Y, et al. Interaction of mature CD3+ CD4+ T cells with dendritic cells triggers the development of tertiary lymphoid structures in the thyroid. J Clin Investig. 2006;116(10):2622–2632. doi:10.1172/JCI28993
61. Muniz LR, Pacer ME, Lira SA, Furtado GC. A critical role for dendritic cells in the formation of lymphatic vessels within tertiary lymphoid structures. J Immunol. 2011;187(2):828–834. doi:10.4049/jimmunol.1004233
62. Liu H, Tian Q, Ai X, Qin Y, Cui Z, Li M. Dihydroartemisinin attenuates autoimmune thyroiditis by inhibiting the CXCR3/PI3K/AKT/NF-kappaB signaling pathway. Oncotarget. 2017;8(70):115028–115040. doi:10.18632/oncotarget.22854
63. Bachelerie F, Ben-Baruch A, Burkhardt AM, et al. Chemokine receptors (version 2020.5) in the IUPHAR/BPS guide to pharmacology database. IUPHAR/BPS Guide Pharmacol CITE. 2020;2020(5). doi:10.2218/gtopdb/F14/2020.5
64. Wang J, Qu C, Xiao P, Liu S, Sun J-P, Ping Y-Q. Progress in structure-based drug development targeting chemokine receptors. Front Pharmacol. 2025;16:1603950. doi:10.3389/fphar.2025.1603950
65. Rahman MF, Kurlovs AH, Vodnala M, et al. Immune disease dialogue of chemokine-based cell communications as revealed by single-cell RNA sequencing meta-analysis. Front Syst Biol. 2024;4:1466368. doi:10.3389/fsysb.2024.1466368
66. Li H, Wu M, Zhao X. Role of chemokine systems in cancer and inflammatory diseases. MedComm. 2022;3(2):e147. doi:10.1002/mco2.147
67. GarcÍa-López MA, Sancho D, Sánchez-Madrid F, Marazuela M. Thyrocytes from autoimmune thyroid disorders produce the chemokines IP-10 and Mig and attract CXCR3+ lymphocytes. J Clin Endocrinol Metabol. 2001;86(10):5008–5016. doi:10.1210/jcem.86.10.7953
68. Jiskra J, Antošová M, Krátký J, et al. CXCR3, CCR5, and CRTH2 chemokine receptor expression in lymphocytes infiltrating thyroid nodules with coincident hashimoto’s thyroiditis obtained by fine needle aspiration biopsy. J Immunol Res. 2016;2016(1):2743614. doi:10.1155/2016/2743614
69. Sun L, Su Y, Jiao A, Wang X, Zhang B. T cells in health and disease. Signal Transduction Targeted Ther. 2023;8(1):235. doi:10.1038/s41392-023-01471-y
70. Shi Q, Xia F, Wang Q, et al. Discovery and repurposing of artemisinin. Front Med. 2022;16(1):1–9. doi:10.1007/s11684-021-0898-6
71. Xu W, Zhu Q, Chen J, et al. Exploring the mechanisms of artemisinin and its derivatives in the treatment of atopic dermatitis based on network pharmacology and molecular docking: a review. Medicine. 2025;104(19):e42287. doi:10.1097/MD.0000000000042287
72. Yi M, Li T, Niu M, et al. Targeting cytokine and chemokine signaling pathways for cancer therapy. Signal Transduction Targeted Ther. 2024;9(1):176. doi:10.1038/s41392-024-01868-3
73. Xia Y, Shi C, Lu J, et al. Artemisinin and its derivatives from molecular mechanisms to clinical applications: new horizons beyond antimalarials. Int J Mol Sci. 2025;26(17):8409. doi:10.3390/ijms26178409
74. Chen J, Gao P, He X, et al. Single-cell profiling reveals immunoregulation of artemisinin on CD8+ GZMB+ T cells via JAK2-STAT3 in malaria-infected mice. Innovation. 2025;6(9):101080. doi:10.1016/j.xinn.2025.101080
75. Yang F, Yang J, Wu M, Chen C, Chu X. Tertiary lymphoid structures: new immunotherapy biomarker. Front Immunol. 2024;15:1394505. doi:10.3389/fimmu.2024.1394505
76. Rafiqi SI, Jaume JC. Thyroid cancer-tumor immune microenvironment (TIME) spatial transcriptomics reveals novel players. J Cancer Biol. 2025;6(2):76–81. doi:10.46439/cancerbiology.6.077
77. Tan C, Huang J, Gao N, et al. Dynamic remodeling of tertiary lymphoid structures in response to cancer therapy: a recent review. Cancer Immunol Immunother. 2025;74(10):313. doi:10.1007/s00262-025-04183-0
78. Huang Y, Yang Y, Liu G, Xu M. New clinical application prospects of artemisinin and its derivatives: a scoping review. Infect Dis Poverty. 2023;12(1):115. doi:10.1186/s40249-023-01152-6
79. Zhu F, Mao H, Du S, et al. CYP3A4-mediated metabolism of artemisinin to 10β-hydroxyartemisinin with comparable anti-malarial potency. Malaria J. 2024;23(1):328. doi:10.1186/s12936-024-05163-y
80. Nghilokwa ET, Petzer JP, Pheiffer W, Hassan IZ, Hayeshi RK. Phytochemical profiling and in silico prediction of interactions between Artemisia afra Jacq. ex-Wild, Catharanthus roseus (L.) G. Don and CYP2B6 enzyme. Discover Appl Sci. 2025;7(10):1134. doi:10.1007/s42452-025-07685-9
81. Kam R. Artemisia annua tea drug interactions: new method development. Worcester Polytechnic Institute.
82. Alvarez-Sierra D, Marin-Sanchez A, Gomez-Brey A, Bello I, Caubet E, Moreno-Llorente P. Lymphocytic thyroiditis transcriptomic profiles support the role of checkpoint pathways and B cells in pathogenesis. Thyroid. 2022;32(6):682–693. doi:10.1089/thy.2021.0694
83. Armengol MP, Cardoso-Schmidt CB, Fernandez M, Ferrer X, Pujol-Borrell R, Juan M. Chemokines determine local lymphoneogenesis and a reduction of circulating CXCR4+ T and CCR7+ B and T lymphocytes in thyroid autoimmune diseases. J Immunol. 2003;170(12):6320–6328. doi:10.4049/jimmunol.170.12.6320
84. Song K, Wang X, Yao W, Wang Y, Zhang Q, Tang Y. Construction of the single-cell landscape of Hashimoto’s thyroiditis tissue and peripheral blood by single-cell RNA sequencing. Immun Inflamm Dis. 2025;13(2):e70153. doi:10.1002/iid3.70153
85. Lu J, Song L, Luan J, et al. Identification of shared biomarkers and immune infiltration signatures between vitiligo and hashimoto’s thyroiditis. Clin Cosmetic Investig Dermatol. 2024;Volume 17:311–327. doi:10.2147/CCID.S451080
86. Long Z, Xiang W, Xiao W, et al. Natural Products and Their Derivatives in the Treatment of Inflammatory and Autoimmune Diseases. Comprehensive review. 2025.
87. Domberg J, Chao L, Papewalis C, Pfleger C, Xu K, Willenberg HS. Circulating chemokines in patients with autoimmune thyroid diseases. Horm Metab Res. 2008;40(6):416–421. doi:10.1055/s-2008-1073151
88. Aust G, Sittig D, Becherer L, et al. The role of CXCR5 and its ligand CXCL13 in the compartmentalization of lymphocytes in thyroids affected by autoimmune thyroid diseases. Eur J Endocrinol. 2004;150(2):225–234. doi:10.1530/eje.0.1500225
89. Mohr A, Tresallet C, Monot N, Bauvois A, Abiven D, Atif M. Tissue infiltrating LTi-like group 3 innate lymphoid cells and T follicular helper cells are associated with tertiary lymphoid structures in patients with Hashimoto’s thyroiditis. Front Immunol. 2020;11601278. doi:10.3389/fimmu.2020.601278
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.