Back to Journals » International Journal of Nanomedicine » Volume 21
Application and Future Perspectives of Extracellular Vesicle-Loaded Scaffold in Spinal Cord Injury
Authors Zhang W, Chai X, Xu N, Ma W, Zhou Y, Lou X, Yang H
Received 16 April 2026
Accepted for publication 13 June 2026
Published 23 June 2026 Volume 2026:21 617539
DOI https://doi.org/10.2147/IJN.S617539
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Eng San Thian
Weihu Zhang,1,2,* Xiaoyin Chai,1,2,* Nanjian Xu,2,3 Weihu Ma,2,3 Yun Zhou,1,2 Xiaohan Lou,1,2 Hailin Yang1,2
1Department of Nursing, Ningbo No.6 Hospital, Ningbo, Zhejiang, People’s Republic of China; 2Ningbo Clinical Research Center for Orthopedics, Sports Medicine & Rehabilitation, Ningbo, Zhejiang, People’s Republic of China; 3Spine Surgery Center, Ningbo No.6 Hospital, Ningbo, Zhejiang, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Hailin Yang, Department of Nursing, Ningbo No.6 Hospital, 1059# Zhongshan East Road, Ningbo, 315040, People’s Republic of China, Email [email protected]
Abstract: Following the initial trauma of spinal cord injury (SCI), the secondary injury phase—characterized by inflammation, oxidative stress, neuronal death, and axonal demyelination—establishes an adverse microenvironment that hinders functional recovery. Despite the availability of existing clinical therapies, they often yield suboptimal functional recovery. In this review, the applications and future prospects of extracellular vesicle (EV)-loaded scaffolds for SCI repair are summarized. This review discusses the biological properties and therapeutic action of EVs, as well as limitations of using them as a single agent, such as rapid clearance and poor targeting. Other scaffold loading strategies and types that change in response to the microenvironment are also reviewed. This review highlights the advancement of EV-loaded scaffolds in alleviating secondary injury, controlling inflammation, enhancing neural regeneration and remyelination, and promoting angiogenesis. Microvesicles, a high-capacity and large subtype of EVs with rapid release and functional surface proteins, in particular, have been shown to have multi-target repair capabilities in various disease models. Their integration with scaffolds for SCI is suggested as a promising translational direction. Current challenges encompass subtype standardization, safety evaluation, as well as large-scale production. The review concludes that EV-loaded scaffolds are not just delivery systems, but a regenerative platform that enables spatiotemporal structure-signal synergy. Subtype-specific selection, pathology-stage-directed release, and industry-compliant quality control should be the focus in future studies to develop this strategy beyond basic research into clinical translation.
Keywords: spinal cord injury, extracellular vesicles, microvesicles, scaffold, neural regeneration, microenvironment regulation, tissue engineering
Introduction: Therapeutic and Nursing Dilemmas in Spinal Cord Injury
Spinal Cord Injury (SCI) is a devastating disorder that grossly interferes with the homeostasis of the Central Nervous System (CNS). Considering its high disability rate and the higher risk of premature mortality in young people, SCI has emerged as a major cause of the loss of healthy life expectancy and a major public health issue of concern in the world.1,2 From a pathophysiological standpoint, normal spinal cord function can be dependent upon complex interactions among neurons, astrocytes, microglia, and oligodendrocytes. SCI results in physical disruption and dysfunction of this multicellular network, thereby impeding neural repair.3 The majority of human SCIs stem from mechanical traction, penetrating injuries, or acute or chronic compression caused by displaced bone fragments or herniated intervertebral discs.4,5 The process generally starts with a primary injury following sudden spinal trauma, manifested as vertebral dislocation, fracture, and ligament tear.6,7 Under the effect of the primary injury, substantial amounts of intracellular calcium ions accumulate, reactive oxygen species (ROS) increase, and glutamate is released excessively. As a result, severe neuronal excitotoxicity is triggered. This is accompanied by pro-apoptotic signaling, ischemia, and inflammatory cell infiltration. Continuous cell necrosis releases ATP, DNA, and free radicals. On that basis, a destructive cytotoxic microenvironment is created, which damages the molecular structure of nucleic acids, proteins, and phospholipids. Consequently, permanent neurological dysfunction is triggered.8,9
In response to spinal canal mechanical compression caused by secondary edema and hemorrhage post-SCI, which results in severe inflammation and nerve damage, early surgical decompression has become the standard intervention. The aim of early surgical decompression is at blocking the progression of secondary damage through physical decompression.10 Concerning drug therapy, methylprednisolone (MPSS), a classic corticosteroid, can improve local blood flow and preserve neuronal excitability by inhibiting lipid peroxidation, safeguarding spinal cord neurofilaments, and boosting robust Na+-K+-ATPase activity.11,12 Moreover, anti-inflammatory drugs, sodium channel blockers, hypothermia therapy, and systematic rehabilitation training display some neuroprotective potential in clinical practice.9,13–15 Nonetheless, existing treatment strategies are ineffective in fully reversing the pathological damage caused by SCI. Existing research indicates that the clinical prognosis of surgical decompression is largely determined by the timing of the intervention and exhibits considerable individual variability.16 Notably, the application of high-dose MPSS remains subject to ongoing debate, uncertainty surrounding its long-term benefits and potential dose-dependent risks.17 Beyond acute interventions, long-term SCI care confronts profound systemic and bedside dilemmas. Rehabilitation demands are resource-intensive yet often inaccessible, especially in under-resourced regions, while fragmented care pathways undermine continuity. Nursing faces persistent challenges in pressure ulcer prevention, neurogenic bladder/bowel management, recurrent infections, and spasticity control, compounded by patient non-adherence and inadequate family/caregiver training. Psychosocially, depression, anxiety, and diminished quality of life are pervasive, straining support networks. Economically, lifelong direct/indirect costs impose catastrophic financial burdens on families and healthcare systems alike. Thus, despite incremental refinements in surgery and pharmacotherapy, SCI remains a condition where restoration lags far behind stabilization—and where daily clinical reality involves trade-offs between suboptimal options amid unmet needs.
Thus, the development of safe and effective new regenerative medicine strategies is urgently needed. To be specific, EV-borne biological scaffolds, showing exceptional potential for neural microenvironment remodeling and repair, offer new avenues for structural reconstruction and functional recovery after SCI.
EVs: Biology and Therapeutic Potential
Biogenesis, Classification, and Composition
EVs refer to a class of membrane-bound nanoparticles released by cells that serve as crucial carriers of intercellular communication. Their biological functions are largely determined by their contents, and they are extensively involved in a variety of physiological and pathological processes. Existing research confirms that these vesicles can efficiently transport bioactive substances (eg., proteins, lipids, and nucleic acids), thereby regulating the function and phenotype of target cells. As endogenous natural carriers, EVs are capable of crossing biological barriers, delivering their contents stably, and mediating complex intercellular signal transduction within the local microenvironment. Consequently, they hold critical significance in multiple biological processes (eg., immune regulation, tissue repair, and neural modulation).18 Virtually all living cells can release EVs, which are generally classified into three categories based on their origin and formation method: exosomes, microvesicles, and apoptotic bodies.19,20 Their classification primarily follows their origin and biogenesis, such that their molecular composition and functional characteristics can be determined. Thus, each EV subtype plays a unique role in intercellular communication while holding different significance for physiological regulation, disease progression, and emerging therapeutic applications.21,22 Microvesicles (MVs), which are membrane-encapsulated, express endosome-related markers, and can cross the blood-brain barrier, efficiently deliver neurotrophic factors and exhibit low immunogenicity, hold significant application prospects in neurobiological research.23–25
The Therapeutic Mechanism of EVs in Neural Repair
EVs, as endogenous lipid bilayer structures, exhibit favorable biocompatibility profiles. These vesicles function as natural delivery carriers for bioactive substances (eg., nucleic acids and proteins). EVs are recognized as a promising delivery platform for genome editing drugs.26 In terms of delivery advantages, certain subpopulations of EVs can penetrate tissue and cell barriers, thereby crossing the blood-brain barrier under specific pathophysiological conditions. In this regard, a necessary precondition is given to targeted therapy in central nervous system diseases.27,28 Specific subpopulations of EVs derived from particular cell types have inherent targeting characteristics in certain microenvironments, and they are attracted to specific cells or tissues, which enables the selective delivery of drugs and minimizes off-target effects.29 Autologous or low-immunogenic EVs generally exhibit favorable biocompatibility profiles, often demonstrating reduced immunogenicity and systemic toxicity compared to certain artificial carriers (eg., LNPs and polymer micelles). However, the safety and efficacy margins are highly dependent on the specific EV source, engineering strategy, and route of administration.30 Furthermore, EVs have the potential to effectively deliver large molecules, including proteins, with a wider payload capacity.31 It is noteworthy that specific subtypes of EVs can circumvent endocytosis and lysosomal degradation, thereby directly delivering payloads (eg., siRNA) to the cytoplasm in certain experimental models. Thus, molecular transfection and intervention efficacy can be enhanced.32
Despite EVs’ multiple delivery advantages, their application in vivo as a single treatment still faces significant limitations. As a result, their clinical translation and effectiveness are greatly restricted. Firstly, the heterogeneity in EV particle size and surface components directly impacts their biodistribution in vivo, reducing their efficiency in penetrating tissues, biological barriers, and targeting cell membranes, and resulting in insufficient effective enrichment at lesion sites.33 Secondly, research aimed at enhancing the delivery efficiency of EV payloads to intracellular action sites is still relatively limited; a substantial proportion of EVs taken up by cells enter degradation pathways, which compromises their therapeutic impact.33 Additionally, free EVs are rapidly cleared from the body. Notably, despite the availability of various isolation methods (eg., ultracentrifugation, microfluidics), standardized, high-purity, and scalable EV separation and purification technologies remain lacking. This results in low production yields and inconsistent product quality that do not meet clinical demands. Large-scale production and quality control are also expensive, thus posing significant challenges to the industrialization of EVs.34,35 When coupled with therapeutic loads, EVs may also trigger non-specific adverse reactions, such as complement activation-related pseudoallergy, thrombogenicity due to surface protein expression, or unintended pro-inflammatory responses induced by specific RNA or protein cargos, further affecting therapeutic stability and safety.36,37 Therefore, the use of free EVs alone is not sufficient to produce stable, long-term, and specific neural repair outcomes. Engineering approaches should be used to load and enhance EVs. In this regard, the critical issues of fast in vivo clearance, low targeting, low local effective concentration, and low efficiency of intracellular delivery can be resolved. While bio-scaffolds are generally designed to be biocompatible and minimize immune recognition, they are not entirely inert; certain scaffold materials may also elicit foreign body reactions, fibrosis, or chronic inflammation, potentially exacerbating the overall risk profile rather than eliminating it38 (Figure 1).
 |
Figure 1 Exosomal biogenesis and pathways to biological roles.24 |
Strategies for Loading EVs into Scaffolds
Loading Strategies
Carrier embedding has been widely used in conjunction with strategies like physical adsorption, chemical coupling, and biofunctionalization in the compositional integration of EVs and scaffold materials. The retention of EV bioactivity, release kinetics, and functional outcomes are significantly affected by different loading methods.
Physical adsorption primarily incorporates EVs onto the scaffold surface via electrostatic interactions, hydrophobic interactions, or hydrogen bonding. This method is conducted under mild conditions and optimizes the preservation of the EV membrane structure and bioactivity.39 For instance, Zhang et al functionalized EVs by anchoring cholesterol-coupled aptamers to the EV membrane surface through hydrophobic interactions. Subsequently, aptamer-modified EVs can be loaded onto a Polyetheretherketone-polydopamine scaffold through electrostatic adsorption. This approach has demonstrated efficient antibacterial activity and improved targeting of EVs in orthopedic implants.40 Implanting EVs into scaffolds through physical adsorption can address the spatial delivery control issue. Notably, its loading efficiency is heavily dependent on the surface properties of the material, lacking other bonding methods’ specificity and strength. Moreover, EVs loaded onto the surface are susceptible to “burst release” under physiological conditions, which results in excessively high initial release rates.41 In comparison, physical embedding involves directly mixing EVs into the scaffold precursor solution and subsequently encapsulating them within the scaffold through gelation or solidification. The release behavior is primarily governed by diffusion and scaffold degradation rates, enabling sustained release over several days to weeks. In some existing studies, 3D molding technology was used for directly load EVs into polymeric composite scaffolds. Specifically, embedding EVs in a PLA-gelatin composite scaffold can promote cell proliferation, migration, and collagen synthesis related to tympanic membrane repair.42 Moreover, the use of low-temperature 3D printing to create composites of IFN-γ-preconditioned exosomes with collagen/chitosan scaffolds not only preserves the bioactivity of EVs maximally but also ensures long-term retention and sustained release of EVs at the injury site, thereby guaranteeing prolonged therapeutic effects.43 However, it is crucial to acknowledge that shear forces, organic solvents, or UV crosslinking during the preparation process may compromise the integrity of EVs.41,44
Chemical coupling anchors EVs to scaffold surfaces or frameworks through covalent bonds, which significantly arrest the diffusion of EVs and support the sustained local maintenance of their concentrations.45 However, covalent bonding may lead to the loss of some biological activity by occupying crucial functional proteins such as CD63 and targeting peptides found on the EV surface. Recently, the development of breakable linkages has emerged as a promising direction for improvement, enabling on-demand release of active EVs after reaching the target site.46 Therefore, the literature emphasizes the need for protecting EVs’ integrity and functionality during the binding process with materials, suggesting the introduction of orthogonal functional groups through metabolic engineering and other strategies under mild physiological conditions to minimize interference with active molecules on the EV surface.47,48 Biospecific binding employs affinity interactions, such as streptavidin-biotin, antigen-antibody, or membrane fusion proteins, for achieving efficient and directional fixation of EVs, balancing high loading rates and spatial orientation control under mild conditions. The above-mentioned strategies mimic the receptor-ligand interactions between EVs and matrix components in the natural extracellular matrix. They are expected to achieve precise anchoring while preserving EV activity.49–51 As indicated by existing research, a combined strategy of chemical conjugation and physical embedding can achieve stable loading and functional preservation of EVs. For instance, the surface chemical modification of PLGA-based porous scaffolds with PEI enables an efficient immobilization of melatonin-regulated EVs via electrostatic interactions. Combined with a physical embedding process using ice particle leaching, the constructed PLGA/Mg (OH)2-RA/ECM/ZnO-ALA scaffold with melatonin-modulated extracellular vesicles scaffold accomplishes both an initial burst release and a prolonged sustained release of EVs. Importantly, after loading, the EVs maintain intact surface marker expression and endogenous miRNA regulatory functionality, continuing to elicit pro-angiogenic and anti-inflammatory repair effects at the injury site.52
In general, physical strategies offer significant advantages in activity preservation, whereas the release regulation capabilities are limited. Chemical strategies provide precise control over release but require balancing activity loss. Biospecific binding represents the direction for precise loading. The selection among these strategies requires a comprehensive trade-off based on the scaffold material properties, the desired release profile, and the repair demands of the injury microenvironment41,53 (Figure 2).
 |
Figure 2 EV biogenesis, composition, and uptake.54 |
Scaffold Types: How to Adapt to the Microenvironment of SCI
To overcome the pathological features of a severe inflammatory response and a severe neural regeneration microenvironment after SCI, different types of scaffolds, such as hydrogels, 3D-printed scaffolds, and electrospun fiber scaffolds, are essential. These scaffolds are able to provide specific adaptation to the injury microenvironment by regulating core characteristics including material stiffness, porosity, and degradability, thereby facilitating EV delivery and neural repair.
Hydrogel scaffolds, whose physicochemical characteristics are similar to the natural extracellular matrix (ECM), have emerged as important vectors in SCI repair. Gelatin methacrylamide (GelMA) hydrogels possess adjustable mechanical properties and demonstrate favorable biocompatibility, facilitating the adhesion, survival, and neuronal differentiation of neural progenitor cells (NPCs). Their composite systems with tetramethylpyrazine (TMP) and neural stem cell-derived EVs (NSC-EVs) can reduce cystic cavity formation and promote axonal regeneration by mimicking the neural niche microenvironment.55 Collagen-based hydrogels (CbH), owing to their good biodegradability, synergistically construct a controlled-release system with placental mesenchymal stem cell-derived EVs (PMSCs-Exo) microspheres. During the slow degradation process at the injury site, these EVs are continuously released, significantly reducing the levels of TNF-α and IL-1β, and increasing antioxidant enzyme activity, effectively inhibiting the spread of damage.56 Furthermore, peptide-based hierarchical microgel scaffolds constructed from aptamer conjugates possess both antioxidant capacity and nuclease sensitivity. They can anchor MSC-Exo cells through immunoaffinity mimicry to reduce lesion loss and achieve precise release of EVs through degradation properties. These actions guide neural stem cells to differentiate into neurons rather than astrocytes, thus creating conditions for microenvironmental immune regulation and neural regeneration.57 Cryo-assembled silk fibroin (SF) sponge scaffolds, formed through low-temperature phase separation technology, display unique silk I structures characterized by excellent extensibility and enzyme-responsive degradation. Their porous structure efficiently loads EVs and facilitates a degradation-driven sustained release, maintaining EV retention and activity for up to two months in vivo. This performance is significantly superior to that of clinically used fibrin glue and promotes angiogenesis and cell infiltration through continuous EV release.58 3D printing technology further enhances the repair effects of SCI by precisely controlling the scaffold’s structure and properties. For instance, 3D bioprinted gelatin methacryloyl/tetramethylpyrazine composite scaffolds create porous structures that not only provide stable loading spaces for EVs and NPCs but also promote nutrient exchange and cell infiltration. In a rat model of whole-segment SCI, these scaffolds have been shown to inhibit neuroinflammation and protect the integrity of the parenchyma.55 Additionally, low-temperature 3D-printed collagen/chitosan scaffolds43 preserve EV bioactivity while ensuring mechanical stability through a low-temperature molding process. Their controllable degradation rate and continuous release characteristics match the repair demands of different stages post-SCI, achieving long-term therapeutic effects. Another study embedded Angiopep2-modified brown adipose tissue-derived extracellular matrix (BAT-EVs) in a targeted delivery system constructed from GelMA hydrogel,59 demonstrating the synergistic effects of scaffold material properties and functional modifications. The slow degradation of the hydrogel, combined with the targeted delivery of EVs, enables precise accumulation of bioactive cargoes such as miR-692 at the injury site. This strategy effectively regulates the anti-inflammatory polarization of microglia, providing an optimized approach that integrates structural support with functional targeting to adapt scaffold material properties to the SCI microenvironment.
Although there are currently no studies on direct EV loading for SCI repair using electrospun fiber scaffolds, their fundamental properties and interdisciplinary achievements provide important references for their application. Electrospun scaffolds based on polycaprolactone (PCL) and polyvinylpyrrolidone (PVP), with their ECM-like nanofiber structure, high porosity of 80%-90%, and adaptive stiffness of 10–50 kPa, can promote nutrient exchange and axon directional growth.39 Embedding EVs into PVP-based electrospun fibers can create a solid-state storage system, maintaining the fiber structure and EV activity even after 12 weeks of storage at 4°C or room temperature. Furthermore, the water-soluble matrix can achieve rapid dissolution and release of EVs, addressing their storage and transportation issues.60 After modification with collagen, polydopamine (PDA), etc., the stability of EV loading can be enhanced through specific binding. The core mechanism of this strategy could be referenced in SCI repair, providing insights for structure-guided and EV-loaded delivery-assisted therapies, and offering a feasible solution.39
Different scaffold types, through distinct material properties, are designed to meet the pathological needs of SCI from dimensions such as structural support, microenvironment regulation, and stable delivery of EVs. Among these, the hydrophilicity and sustained-release properties of hydrogels, the structural customization of 3D-printed scaffolds, and the axonal guidance and storage advantages of electrospun scaffolds provide multi-dimensional solutions for SCI repair. Furthermore, the combination of EVs and scaffolds achieves a synergistic improvement in both structure and function.
Applications of EV-Loaded Scaffolds in SCI Repair
Mitigating Secondary Damage and Regulating the Microenvironment
The pathological cascade reactions and adverse repair microenvironment following SCI, such as the secondary inflammatory storm, oxidative stress, and mitochondrial dysfunction, represent significant barriers to the recovery of neurological function.61 Single scaffolds often fail to provide multi-dimensional intervention. In contrast, EV-loaded scaffolds offer an efficient solution for regulating the microenvironment and inhibiting secondary injury through the synergistic effects of their components. The 3D bioprinted gelatin methacrylamide (GelMA) multifunctional scaffold developed by Liu et al55 innovatively incorporates neural progenitor cells (NPCs), neural stem cell-derived extracellular vesicles (NSC-EVs), and tetramethylpyrazine (TMP). This configuration achieves a triple effect of “structural support, cellular regulation, and biochemical intervention.” NSC-EVs deliver biochemical signals that enhance NPC activity by regulating inflammatory responses, inhibiting oxidative stress and apoptosis, promoting neurogenesis, and preserving mitochondrial function.62–64 TMP, derived from Ligusticum chuanxiong, possesses antioxidant, anti-inflammatory, and mitochondrial protective properties.65,66 Integrating these two elements with the scaffold allows for local continuous delivery, effectively promoting neuroprotection, axonal regeneration, myelin repair, angiogenesis, and immune regulation. This combinatorial approach reprograms the unfavorable microenvironment post-injury into one conducive to structural reconstruction and functional recovery. Using stem cell-derived exosomes combined with biomimetic electromagnetic hydrogels can synergistically improve the early inflammatory response of spinal cord injury and enhance the regeneration of new neurons and axons.67 In a rat model of SCI, this approach markedly enhanced the expression levels of antioxidant enzymes such as GSH, SOD, and CAT, and the anti-inflammatory factor IL-10. Simultaneously, it reduced levels of pro-inflammatory cytokines such as TNF-α and IL-1β, as well as the lipid peroxidation product MDA. This intervention effectively curtailed the extension of secondary injury and fostered a stable microenvironment for nerve repair.
Promoting Neural Regeneration and Remyelination
Impairment of neural regeneration and myelin damage are central pathological features that limit functional recovery after SCI. EV-loaded scaffolds provide vital support for axonal regeneration, neuronal differentiation, and myelin repair by precisely delivering bioactive signals and creating a biomimetic microenvironment. Inspired by antibody-antigen binding mechanisms, Zheng et al developed a hierarchical microgel scaffold based on peptide-aptamer (Peptide-AptCD63) conjugates.57 This scaffold ensures precise anchoring and controllable release of EVs at the injury site through immunoaffinity mimicry targeting CD63-labeled MSC-Exo cells. Moreover, its outstanding antioxidant properties help alleviate local oxidative stress. The MSC-Exo cells that are released off the scaffold greatly increase the proliferation and migration of the neural stem cells and selectively cause them to develop into neurons as opposed to astrocytes. This synergistic action, comprising of glial scar inhibition and neural regeneration, is effective in improving motor performance in SCI mice.
Conductive hydrogels are biomimetic scaffolds that combine mechanical support with electrical signal conduction to form a distinct synergistic microenvironment to regenerate nerves.68 Their tissue-like softness and biomimetic electrical fields mimic the natural nervous system environment, which greatly enhances the neuronal differentiation efficiency of neural stem cells and directs the directional growth of neurites.69,70 The neurogenic conductive hydrogel developed by Fan et al,71 which is loaded with bone marrow stem cell-derived exosomes, effectively alleviates local inflammation and clears obstacles for nerve regeneration in a mouse model of SCI through the dual effects of electrical signal regulation and EV bioactive delivery. It also directly fosters axonal regeneration and local neurogenesis, leading to significant functional recovery in the early stages of injury. This approach provides a multidimensional synergistic treatment strategy for nerve regeneration and myelin repair post-SCI.
Supporting Angiogenesis and Tissue Remodeling
Impairment in local blood supply and an imbalance in tissue repair capabilities following SCI severely hinder the recovery of neurological functions. EV-loaded scaffolds play a vital role in supporting blood supply reconstruction and structural remodeling in the injured area by modulating angiogenesis-related pathways and tissue repair processes.
Core-shell nanofiber scaffolds, loaded with bone marrow mesenchymal stem cell-derived EVs, significantly improve the tubular structure formation ability of endothelial cells and effectively regulate the repair microenvironment in the injured region. By activating downstream pro-angiogenesis signaling pathways, these scaffolds expedite the formation of new blood vessel networks and promote orderly tissue remodeling.72 Long-acting EV delivery systems, based on cryogel technology, preserve vesicle structure and bioactivity over extended periods in complex physiological environments. By alleviating local oxidative stress, regulating the phenotypic transformation of inflammatory cells, and promoting orderly collagen fiber deposition, these systems concurrently advance angiogenesis and functional tissue repair.73 Additionally, adipose-derived endothelial cells effectively increase the efficiency and maturity of vascular network formation through their enriched pro-angiogenic miRNA profiles. The pro-repair effects of these cells exhibit significant dependence on scaffold properties. Optimizing the scaffold material, structure, and degradation rate can further regulate the topological morphology, connectivity, and structural stability of newly formed blood vessels, ultimately facilitating the synergistic reconstruction of the vascular system and matrix structure after tissue injury, and providing comprehensive microenvironmental support for tissue regeneration74 (Figure 3).
 |
Figure 3 Applications of EV-Loaded Scaffolds in SCI Repair. |
Harnessing the Specific Potential of MVs: Lessons from Other Diseases
Unique Advantages of MVs
EVs are primarily classified into three subtypes based on their biological development pathways: exosomes, MVs, and apoptotic bodies. Among these, exosomes and MVs are currently recognized as important carriers for local and long-distance intercellular communication.75 Exosomes originate from the endocytic pathway, with early budding from the endosome membrane to form intraluminal vesicles (ILVs). As this process progresses, the endosomes gradually mature into multivesicular bodies (MVBs) and eventually fuse with the plasma membrane, releasing exosomes with a diameter of about 10–150 nm into the extracellular space.76,77 The endosomal sorting transport complex (ESCRT) mechanism is the main regulator of this process, although it may also take place via non-ESCRT-dependent pathways.77–79
In contrast, MVs bud and fission directly outward from the plasma membrane, with diameters ranging from 200 nm to several micrometers, and as small as 30 nm.76,80 Their biogenesis is a concerted effort of several molecular processes: the directional movement of certain membrane proteins to the budding location on the plasma membrane is mediated by the activation of the small GTPase ARF6. RhoA and its downstream effector molecules, ROCK and ERK, regulate actin-myosin-mediated vesicle neck contraction, ultimately leading to fission and release. Furthermore, the eversion of phosphatidylserine (PS) and changes in lipid composition mediated by phospholipase D2 also play key roles in membrane curvature regulation and vesicle formation.81 Since MVs originate directly from the plasma membrane, their contents are primarily enriched with cytoplasmic and plasma membrane-associated proteins, especially tetraspan proteins and other molecules known to aggregate on the plasma membrane surface.82,83 Differences in membrane components bound by different tetraspan proteins determine the functional diversity of MVs in processes such as promoting or inhibiting translocation.83
Functionally, the larger diameter of MVs allows them to carry larger fragments of proteins, nucleic acids, and lipids. Furthermore, MVs have been reported in specific experimental settings to encapsulate complex cargo, such as organelle fragments or macromolecular complexes, which may contribute to their potent delivery capabilities.84,85 The cytoskeletal proteins, heat shock proteins, integrins, and post-translationally modified proteins, such as glycosylated and phosphorylated proteins commonly found on their surface, may be the key molecular basis for their targeting of other cells and mediating interactions.86–88 It is important to note that the fundamental differences in the biological mechanisms of MVs and exosomes result in significant differences in their content composition and biological functions, thereby providing a theoretical basis for targeting different EV subtypes for specific therapeutic strategies.81
Apoptotic bodies, as a third subtype, are larger than 1 micrometer in diameter and are released exclusively by cells undergoing programmed cell death. Their formation is related to cytoskeletal dissociation and membrane vesicle formation78 and will not be discussed further in this review.
The Multifunctional Role of MV-Loaded Scaffold: Insights from Other Disease Domains
MVs, as novel messengers for intercellular communication, play a central role in the construction and remodeling of the tumor microenvironment.20 Tumor cell-derived MVs selectively load miRNAs, mRNAs, and proteins, stably delivering them to both nearby and distant non-malignant cells, thereby facilitating the horizontal transfer of genetic information.20,89,90 In the tumor microenvironment, MV-mediated communication exhibits bidirectional characteristics: on the one hand, tumor cells “educate” endothelial cells, immune cells, and fibroblasts through MVs, transforming them into phenotypes that support tumor growth; on the other hand, stromal cells in the microenvironment can transmit pro-proliferation and pro-drug resistance signals to tumor cells through MVs, forming a vicious cycle.20,89–91 The regulatory roles of MVs in the tumor microenvironment are primarily reflected in the following aspects: First, they promote angiogenesis—monocyte-derived MVs significantly enhance the migration ability of endothelial cells by delivering miR-150,92 while leukemia cell-derived MVs promote endothelial cell lumen formation through mechanisms such as miR-17-92 clusters;93 second, they facilitate immune escape—tumor MVs carry immunosuppressive molecules like TGF-β1, inhibiting functions of natural killer cells. This includes 1) inhibiting Th1 cell-mediated anti-tumor immune responses by delivering miRNAs such as let-7d via MVs derived from regulatory T cells94,95 and 2) enhancing invasion and metastasis—MVs derived from macrophages boost the invasive capability of breast cancer cells by delivering miRNAs such as miR-223, while MVs derived from fibroblasts promote tumor cell movement and metastasis through the Wnt signaling pathway.96,97 These studies underscore that MVs, with their stable membrane structures, selective loading mechanisms, and broad targeting capabilities, have evolved into mediators of intercellular communication within the tumor microenvironment.20,89,98,99
MVs, which are major mediators of paracrine effects, have shown great promise in the treatment of cardiovascular diseases, especially myocardial infarction (MI) and lower limb ischemia. Mesenchymal stromal cell (MVs) are rich in bioactive factors such as pro-angiogenic (VEGF and HIF-1α) and anti-inflammatory (cytokines) and other regulatory miRNAs. They respond to target cells through horizontal transfer, which enhances angiogenesis, immune homeostasis, and local microenvironment. In vitro studies have emphasized that these MVs can be effectively internalized by human umbilical vein endothelial cells (HUVECs) and have a profound effect on endothelial cell migration and lumen formation by increasing the expression of pro-angiogenic genes such as VEGF and HIF-1α; and anti-inflammatory effects by suppressing the expression of pro-inflammatory factors like IL-6 and TNF-α, exerting anti-inflammatory effects. In animal models of lower limb ischemia, local injection of MVs in animal models of lower limb ischemia has been shown to significantly restore blood perfusion in the ischemic limbs, enhance limb preservation rates, and effectively decrease inflammatory infiltration, fibrosis, and apoptosis in muscle tissue.100
Meanwhile, MVs derived from endothelial progenitor cells (EPCs) (EPC-MVs) have demonstrated substantial cardioprotective effects in the treatment of myocardial infarction.101 EPC-MVs exert their therapeutic effects through several synergistic mechanisms. Firstly, in terms of anti-inflammation, EPC-MVs markedly inhibit the activation of NLRP3 inflammasome, which is induced by myocardial infarction. They suppress the expression of NLRP3, Caspase-1, and ASC, thus inhibiting the release of pro-inflammatory cytokines like IL-6 and preventing the inflammatory cascade reaction. Secondly, in terms of anti-apoptosis, EPC-MVs have a significant effect on cardiomyocyte apoptosis in the infarct border zone, increasing the expression of the anti-apoptotic protein Bcl-2 and decreasing the expression of the pro-apoptotic proteins Bax and cleaved caspase-3. Thirdly, concerning the maintenance of myocardial structural integrity, EPC-MVs can restore the downregulated level of CSRP3 in myocardial infarction and reduce the expression of the stress protein NPM1, suggesting that they help alleviate cellular stress and maintain myocardial architecture. Proteomics analysis has further revealed that EPC-MVs regulate key signaling networks such as the HIF-1 signaling pathway, the cGMP-PKG pathway, and the vascular smooth muscle contraction pathway, which collaboratively participate in cellular stress response, metabolic reprogramming, and myocardial repair processes. Histological evaluation confirmed that treatment with EPC-MVs significantly reduced myocardial fibrosis and effectively protected the myocardial structure.
Furthermore, MVs exhibit multiple regulatory functions in bone and cartilage tissue repair, involving various processes such as osteogenic, chondrogenic, anti-inflammatory, and intercellular communication. Regarding gene delivery and osteogenic processes, researchers have developed a non-viral gene delivery vector (MVs-PEI/phBMP2) by loading the bone morphogenetic protein 2 (BMP2) plasmid onto the surface of mesenchymal stem cell-derived MVs using a layer-by-layer self-assembly technique. This vector was then loaded onto a demineralized bone matrix (DBM) scaffold. This gene-activated scaffold enhanced the osteogenic differentiation of mesenchymal stem cells in vitro, promoted the expression of collagen fibers, osteocalcin, osteopontin, and CD34 in an in vivo subcutaneous implantation model, and promoted bone regeneration in a rabbit femoral condyle defect model, while also exhibiting pro-angiogenic effects.102
Secondly, in the context of cartilage protection and anti-inflammation, osteoclast-derived MVs display a unique triple activity, promoting cartilage and bone formation while simultaneously exhibiting anti-inflammatory properties. This triple activity is exclusive to osteoclast-derived MVs and is not observed in osteoclast apoptotic bodies, exosomes, or MVs derived from other cell types.103 In a mouse model of tibial plateau injury-induced osteoarthritis, intra-articular injection of osteoclast-derived MVs significantly reduced the loss of articular cartilage area and thickness, decreased cartilage surface irregularity, inhibited osteophyte formation, alleviated synovial inflammation, and effectively reduced the OARSI osteoarthritis score. These results indicate that osteoclast-derived MVs have the potential to simultaneously treat various components of osteoarthritis and slow down or even reverse the disease.
Moreover, with regard to intercellular communication and bone homeostasis, osteoblast-derived MVs have the ability to deliver RANKL proteins to osteoclast precursor cells. This transfer stimulates osteoclast formation via the RANKL-RANK signaling pathway, thus contributing to the regulation of bone remodeling.104 This finding reveals a new pathway through which MVs can be used as a communicative linkage between osteoblasts and osteoclasts, which can be a new target of interventions in bone metabolic diseases.104
MVs have a multifaceted neurorepair capability in the management of neurodegenerative diseases. The underlying mechanisms are multidimensional, encompassing the control of neurogenesis, systemic inflammation intervention, and repair at the organelle level.
On the neurogenesis and differentiation regulation level, MVs of human umbilical cord mesenchymal stem cells (hUC-MSC MVs) can significantly promote the proliferation and survival of adult neural stem cells (NSCs). This improvement is demonstrated by the fact that the number of neurospheres and derived cells increases, and the expression of the proliferation marker Ki67 is upregulated. In terms of differentiation regulation, hUC-MSC MVs dose-dependently induce NSCs to differentiate into neurons (β-tubulin+) while promoting differentiation into oligodendrocyte precursors (Olig2+) and mature oligodendrocytes (MBP+). It is important to note that the dose effect on mature oligodendrocytes is negative, meaning that lower doses are more effective. These MVs do not have any significant effect on NSCs differentiation into astrocytes (GFAP +). Mechanistic analyses have revealed that MVs increase the expression of Sox10, a key transcription factor in the oligodendrocyte lineage, and thus increase the ability of NSCs to differentiate into oligodendrocytes.105
At the systemic regulatory level, research utilizing Parkinson’s disease (PD) models has disclosed a novel mechanism by which MVs exert neuroprotective effects through the “gut microbiota-brain axis.” Mesenchymal stem cell-derived MVs (MSC-MVs) reverse 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced reductions in dopamine transporter (DAT) and tyrosine hydroxylase (TH) expression in both the striatum and substantia nigra, and decrease the phosphorylated α-synuclein (p-α-Syn)/α-Syn ratio. Concurrently, MSC-MVs ameliorate MPTP-induced disturbances in gut microbiota composition, with the relative abundance of Dubosiella positively associated with the p-α-Syn/α-Syn ratio in both the brain and colon. This association suggests that gut microbiota participates in the neuroprotective effects of MVs. Furthermore, MSC-MVs counteract MPTP-induced declines in specific blood metabolites; the levels of these metabolites are inversely correlated with the p-α-Syn/α-Syn ratio in the brain and colon, thereby confirming the systemic regulatory role of MVs through the “gut microbiota-metabolite-brain” axis106 (Table 1).
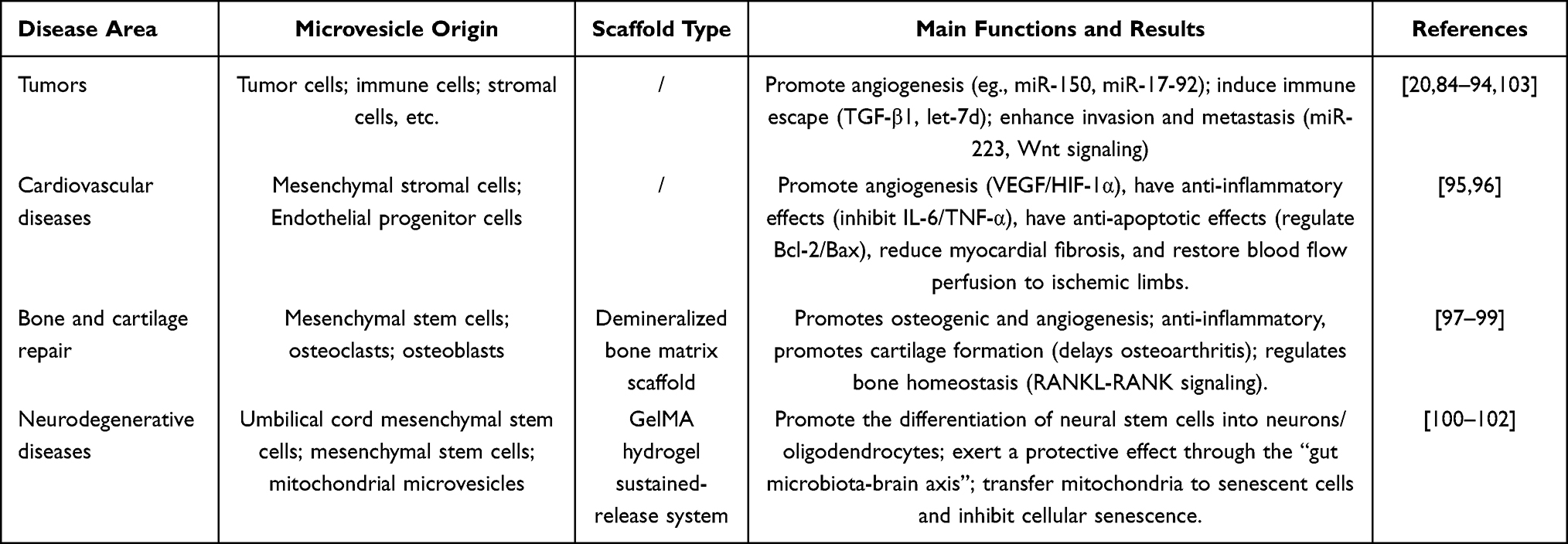 |
Table 1 Harnessing the Specific Potential of MVs: Lessons from Other Diseases |
At the organelle level of repair, studies employing an intervertebral disc degeneration (IVDD) model have demonstrated that mitochondrial MVs (mitoMVs) derived from mesenchymal stem cells can transfer functional mitochondria to senescent nucleus pulposus cells (NPCs). This transfer rescues mitochondrial dysfunction and inhibits cellular senescence. Further investigations have revealed that intervention with a static magnetic field (SMF) significantly enhances the secretion and proportion of mitoMVs. This enhancement is achieved by augmenting kinesin Kif5b-mediated intracellular mitochondrial transport and promoting the interaction between Kif5b and Rab22a, thereby regulating the recruitment of MVs at the plasma membrane budding site. To address the rapid clearance of MVs following local injection, researchers have developed a methacrylamide gelatin (GelMA) hydrogel sustained-release system, which allows for the continuous release of mitoMVs for approximately two weeks. In a rat model of caudal intervertebral disc degeneration, this system has significantly improved the intervertebral disc height index, reduced Pfirrmann grade, inhibited the expression of nucleus pulposus aging markers (P16, P21), and promoted extracellular matrix retention.107
Drawing on the experience of MVs regulating immune cell polarization in oncology, we hypothesize that in SCI, MVs can induce a shift in microglia/macrophages from a pro-inflammatory (M1) phenotype to an anti-inflammatory/repair (M2) phenotype by transporting specific miRNAs or immunomodulatory proteins. The use of engineered MVs or MV-scaffold composite systems may allow for precise remodeling of the inflammatory microenvironment at the injury site, thereby reducing secondary damage and enhancing neurological function recovery. Research in the cardiovascular domain has illustrated that MVs synergistically reverse ischemic tissue damage through mechanisms that include angiogenesis, anti-inflammation, and anti-apoptosis. Inspired by these findings, engineered MVs show promise for precisely targeting key pathological processes following SCI, such as ischemia-hypoxia, inflammatory cascades, and apoptosis. Moreover, studies in bone and cartilage repair have confirmed that MVs possess multi-target regulatory capabilities, which include osteogenic, chondrogenic, anti-inflammatory, and angiogenesis-promoting effects, enabling synergistic intervention in complex pathological processes. Introducing this strategy into SCI repair suggests potential for achieving multifaceted, synergistic repair through inflammatory microenvironment remodeling, angiogenesis, and neuroprotection. Research into neurodegenerative diseases has demonstrated that MVs can promote the directed differentiation of neural stem cells, regulate the gut microbiota-brain axis, and deliver functional mitochondria. Given that SCI also presents core challenges such as neuronal death, inflammatory disorders, and energy metabolism disturbances, engineered MVs are anticipated to facilitate comprehensive, multifaceted repair of damaged spinal cords by synergistically activating endogenous neurogenesis, remodeling the immune microenvironment, and restoring mitochondrial function.
Translational Vision: Potential Research Directions for MV-Scaffold in SCI
By integrating the biophysical and biofunctional properties of MVs, the pathological microenvironment requirements of SCI, and the broad applications across various disease areas, the MV-scaffold composite system can be explored through research focusing on three principal directions: functional targeting, scaffold synergy, and multi-level delivery. This approach is poised to deliver more precise and efficient novel strategies for SCI repair. By harnessing the homing characteristics of tumor-derived MVs, mesenchymal stem cell-derived MVs can be engineered and modified. The coupling with ligands such as Angiopep2 and neural targeting peptides, or the introduction of inflammatory/ROS-responsive elements, facilitates site-specific enrichment, thus addressing the fundamental issues of poor targeting and rapid clearance of free MVs.59,108 Concurrently, employing a mild biomimetic modification strategy may optimize the preservation of functional proteins and bioactivity on the MV membrane surface, ensuring safe application in vivo. Capitalizing on the high surface area and membrane protein richness of MVs, it is possible to design novel scaffolds that exhibit high compatibility with them. Techniques such as grafting CD63 antibodies and peptide-aptamers onto hydrogels like GelMA and collagen/chitosan could enhance loading stability through biospecific binding. Alternatively, the fabrication of large-pore adaptable scaffolds using low-temperature 3D printing (to preserve MV integrity) might reduce damage to MV activity during production. Long-term sustained release can be achieved through the simultaneous regulation of degradation-release, overcoming the limitations associated with physical adsorption’s burst release and chemical coupling’s damaging effects on activity.43,57,109 Additionally, the development of a three-tiered delivery system of “scaffold-MV-internal drug” could be envisioned. In this system, MVs serve as multifunctional delivery carriers to encapsulate therapeutic agents such as neurotrophic factors, miRNAs, and anti-inflammatory small molecules, while preserving the immunomodulatory, regenerative, and anti-apoptotic effects of the membrane proteins themselves, achieving dual treatment of membrane function and internal load.71,107 This system could operate synergistically in sequence: the scaffold providing structural support and inhibiting glial scar formation; MVs continuously regulating the inflammatory microenvironment and promoting angiogenesis in the damaged area; and internal drugs targeting and enhancing neurogenesis, axonal extension, and remyelination. This comprehensive intervention could cover aspects such as the inhibition of secondary injury, nerve regeneration, vascular remodeling, and myelin repair.55,56
Challenges and Considerations
Despite the promising application prospects of MV-based combination therapies in SCI repair, the clinical translation of these therapies still confronts core challenges including insufficient standardization, questionable safety, and difficulties in large-scale production.
In terms of separation and purification, the accurate differentiation and standardized identification of EV subtypes remain primary challenges. Currently, the academic community has not reached a consensus on specific markers for subtypes such as exosomes, MVs, and apoptotic bodies. Traditional detection techniques are not sufficient to clearly differentiate between endosome-derived exosomes and MVs generated by plasma membrane budding; only by monitoring the biogenetic process by in vivo imaging110 can the classification be done accurately. The purity and quality of EV preparation must be of clinical grade. Current techniques, including ultracentrifugation, density gradient centrifugation, ultrafiltration, and immunoaffinity capture, have their own limitations. The most common technique, differential ultracentrifugation, is controversial because it tends to lead to EV aggregation and no single optimal solution has been determined.111–113 In addition, exosome quantification and purity determination does not have a gold standard, and total protein, particle count, and total lipids are prone to changes due to storage and extraction conditions.114 Flow cytometry, pulse resistance sensing, surface plasmon resonance, and atomic force microscopy are all techniques that can be used to characterize exosome particles, but the results are prone to variation because of equipment algorithms and data processing approaches, which require standardized reference materials and standardized methodologies.115,116
In terms of safety, the heterogeneity of exosome sources and the possible pathogenic risks of exosomes cannot be neglected. On the one hand, a growing body of evidence has found that there is a strong connection between viruses and exosomes which have a dual and subtle role in viral infection. Viruses can proliferate, release, and evade the immune system via the biogenetic mechanisms of endothelial cells (EVs). For instance, retroviruses bud using the vesicle pathway, while hepatitis viruses utilize EVs for intercellular transmission, posing potential biosafety risks to the clinical application of EVs.117,118 On the other hand, EVs from pathological sources, such as tumor cells, carry oncogenic nucleic acids and abnormal proteins, potentially transmitting pathological signals. Their potential pathogenic effects have not been fully assessed.119
In terms of large-scale production and clinical translation, the batch preparation and quality uniformity of EVs are the core bottlenecks in industrialization. Current technologies struggle to reliably produce sufficient quantities of homogeneous therapeutic-grade EVs, exhibiting low separation and purification efficiency and high costs for large-scale production, which impedes meeting the needs of clinical research and translational applications.34,35 Future research should focus on validating the safety and efficacy of EVs in large animal models and developing an integrated storage and transportation system to facilitate the advance of MV-scaffold combination therapy from basic research to clinical application.34,35,120
Overall, the future of EV-scaffold combination therapy must not be limited to the use of one exosome subtype, but must capitalize on the biological diversity of the EV family to explore and capitalize on the distinct functions of the various subtypes, including MVs. MVs, which are EV subtypes that are directly generated by budding of plasma membranes, have a number of benefits, such as a high loading capacity, fast release, and unique biological functions, which makes them a very promising candidate in translation. Future studies must focus on mechanism studies, separation method standardization, systematic safety evaluation, and development of large-scale preparation systems. EV-scaffold therapy can be developed as a basic research to clinical application through rigorous preclinical studies and standardized clinical trials.120
Summary
EV-loaded scaffolds are not merely an innovative delivery system, but a regenerative platform that tackles the complex pathology of SCI. They surmount the shortcomings of single drugs or scaffolds by organically combining the anti-inflammatory, regenerative, and angiogenic bioactive cues of EVs with the structural support and sustained-release capabilities of the scaffolds, thus realizing multidimensional, spatiotemporal synergistic control of the injury environment. This integrated “structure-signal” strategy, by modulating the injury microenvironment, enables the platform to concurrently tackle multiple challenges in repair, such as secondary inflammation, axonal damage, myelin loss, and impaired blood supply.
However, existing research generally considers EVs as homogenized therapeutic agents, with a significant knowledge gap evident in the absence of a systematic comparison of the differentiated roles of various EV subtypes, such as exosomes and MVs, at different stages of SCI. MVs, which are large, have high loading capacity, release quickly and have functional protein-carrying membrane surfaces, have a special therapeutic potential. However, their best release mechanisms and signs following incorporation with scaffolds are yet to be elucidated. Thus, the most important step towards clinical implementation is to develop a GMP-compliant, large-scale fabrication and scaffold loading procedure that can differentiate EV subtypes, and to develop potency assays using specific active components. EV-loaded scaffolds will only be able to make the transition between the laboratory and the bedside of SCI patients in the future by advancing towards an engineered platform that is subtype-specific, pathology-stage-guided, and industrially accessible to quality control.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
1. National Key Research and Development Program of China(2023YFC3604401). 2. Ningbo Clinical Research Center for Orthopedics, Sports Medicine & Rehabilitation (2024L004). 3.Ningbo Municipal Health Technology/Science and Technology Program Project(2025Y49).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Ackery A, Tator C, Krassioukov A. A global perspective on spinal cord injury epidemiology. J Neurotrauma. 2004;21(10):1355–18. doi:10.1089/neu.2004.21.1355
2. Global. Regional, and national burden of spinal cord injury, 1990-2019: a systematic analysis for the global burden of disease study 2019. Lancet Neurol. 2023;22(11):1026–1047. doi:10.1016/s1474-4422(23)00287-9
3. O’Shea TM, Burda JE, Sofroniew MV. Cell biology of spinal cord injury and repair. J Clin Invest. 2017;127(9):3259–3270. doi:10.1172/jci90608
4. McDonald JW, Sadowsky C. Spinal-cord injury. Lancet. 2002;359(9304):417–425. doi:10.1016/s0140-6736(02)07603-1
5. Norenberg MD, Smith J, Marcillo A. The Pathology of human spinal cord injury: defining the problems. J Neurotrauma. 2004;21(4):429–440. doi:10.1089/089771504323004575
6. Katoh H, Yokota K, Fehlings MG. Regeneration of spinal cord connectivity through stem cell transplantation and biomaterial scaffolds. Front Cell Neurosci. 2019;13:248. doi:10.3389/fncel.2019.00248
7. Ohnmar H, Das S, Naicker AS. An interesting case of autonomic dysreflexia. Clin Ter. 2009;160(5):371–373.
8. Dimitrijevic MR, Danner SM, Mayr W. Neurocontrol of movement in humans with spinal cord injury. Artif Organs. 2015;39(10):823–833. doi:10.1111/aor.12614
9. Ahuja CS, Fehlings M. Concise review: bridging the gap: novel neuroregenerative and neuroprotective strategies in spinal cord injury. Stem Cells Transl Med. 2016;5(7):914–924. doi:10.5966/sctm.2015-0381
10. Fehlings MG, Vaccaro A, Wilson JR, et al. Early versus delayed decompression for traumatic cervical spinal cord injury: results of the surgical timing in acute spinal cord injury study (STASCIS). PLoS One. 2012;7(2):e32037. doi:10.1371/journal.pone.0032037
11. Hall ED, Braughler JM. Glucocorticoid mechanisms in acute spinal cord injury: a review and therapeutic rationale. Surg Neurol. 1982;18(5):320–327. doi:10.1016/0090-3019(82)90140-9
12. Braughler JM, Hall ED. Effects of multi-dose methylprednisolone sodium succinate administration on injured cat spinal cord neurofilament degradation and energy metabolism. J Neurosurg. 1984;61(2):290–295. doi:10.3171/jns.1984.61.2.0290
13. Wells JE, Hurlbert RJ, Fehlings MG, Yong VW. Neuroprotection by minocycline facilitates significant recovery from spinal cord injury in mice. Brain. 2003;126(Pt 7):1628–1637. doi:10.1093/brain/awg178
14. Fehlings MG, Wilson JR, Frankowski RF, et al. Riluzole for the treatment of acute traumatic spinal cord injury: rationale for and design of the nactn phase i clinical trial. J Neurosurg Spine. 2012;17(1 Suppl):151–156. doi:10.3171/2012.4.Aospine1259
15. Levi AD, Green BA, Wang MY, et al. Clinical application of modest hypothermia after spinal cord injury. J Neurotrauma. 2009;26(3):407–415. doi:10.1089/neu.2008.0745
16. Fehlings MG, Perrin RG. The timing of surgical intervention in the treatment of spinal cord injury: a systematic review of recent clinical evidence. Spine. 2006;31(11):S28–35. doi:10.1097/01.brs.0000217973.11402.7f
17. Hurlbert RJ, Hadley MN, Walters BC, et al. Pharmacological Therapy for Acute Spinal Cord Injury. Neurosurgery. 2013;72:93–105. doi:10.1227/NEU.0b013e31827765c6
18. Lintsen D, Broux B. Effects and mechanisms of adipose tissue-derived extracellular vesicles in vascular inflammation and dysfunction. Neural Regen Res. 2026;21(5):2005–2006. doi:10.4103/nrr.Nrr-d-24-01619
19. Choi DS, Kim DK, Kim YK, Gho YS. Proteomics of Extracellular Vesicles: exosomes and Ectosomes. Mass Spectrom Rev, 2015;34(4):474–490. doi:10.1002/mas.21420
20. Akers JC, Gonda D, Kim R, Carter BS, Chen CC. Biogenesis of Extracellular Vesicles (Ev): exosomes, Microvesicles, Retrovirus-Like Vesicles, and Apoptotic Bodies. J Neurooncol. 2013;113(1):1–11. doi:10.1007/s11060-013-1084-8
21. Kumar MA, Baba SK, Sadida HQ, et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct Target Ther. 2024;9(1):27. doi:10.1038/s41392-024-01735-1
22. Takahashi Y, Takakura Y. Extracellular vesicle-based therapeutics: extracellular vesicles as therapeutic targets and agents. Pharmacol Ther. 2023;242:108352. doi:10.1016/j.pharmthera.2023.108352
23. Wang Y, Xu H, Wang J, Yi H, Song Y. Extracellular vesicles in the pathogenesis, treatment, and diagnosis of spinal cord injury: a mini-review. Curr Stem Cell Res Ther. 2022;17(4):317–327. doi:10.2174/1574888x17666220330005937
24. Li J, Song J, Jia L, et al. Exosomes in central nervous system diseases: a comprehensive review of emerging research and clinical frontiers. Biomolecules. 2024;14(12). doi:10.3390/biom14121519
25. Chen S, Bao Q, Xu W, Zhai X. extracellular particles: emerging insights into central nervous system diseases. J Nanobiotechnol. 2025;23(1):263. doi:10.1186/s12951-025-03354-6
26. Leandro K, Rufino-Ramos D, Breyne K, et al. Exploring the potential of cell-derived vesicles for transient delivery of gene editing payloads. Adv Drug Deliv Rev. 2024;211:115346. doi:10.1016/j.addr.2024.115346
27. Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJ. Delivery of sirna to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol. 2011;29(4):341–345. doi:10.1038/nbt.1807
28. Kooijmans SA, Vader P, van Dommelen SM, van Solinge WW, Schiffelers RM. Exosome mimetics: a novel class of drug delivery systems. Int J Nanomed. 2012;7:1525–1541. doi:10.2147/ijn.S29661
29. Kooijmans SAA, Schiffelers RM, Zarovni N, Vago R. Modulation of tissue tropism and biological activity of exosomes and other extracellular vesicles: new nanotools for cancer treatment. Pharmacol Res. 2016;111:487–500. doi:10.1016/j.phrs.2016.07.006
30. Ha D, Yang N, Nadithe V. Exosomes as therapeutic drug carriers and delivery vehicles across biological membranes: current perspectives and future challenges. Acta Pharm Sin B. 2016;6(4):287–296. doi:10.1016/j.apsb.2016.02.001
31. Haney MJ, Klyachko NL, Zhao Y, et al. Exosomes as drug delivery vehicles for parkinson’s disease therapy. J Control Release. 2015;207:18–30. doi:10.1016/j.jconrel.2015.03.033
32. Mehrotra N, Tripathi RM. Short interfering rna therapeutics: nanocarriers, prospects and limitations. IET Nanobiotechnol. 2015;9(6):386–395. doi:10.1049/iet-nbt.2015.0018
33. Murphy DE, de Jong OG, Brouwer M, et al. Extracellular vesicle-based therapeutics: natural versus engineered targeting and trafficking. Exp Mol Med. 2019;51(3):1–12. doi:10.1038/s12276-019-0223-5
34. Petersen KE, Manangon E, Hood JL, et al. A review of exosome separation techniques and characterization of b16-f10 mouse melanoma exosomes with af4-uv-mals-dls-tem. Anal Bioanal Chem. 2014;406(30):7855–7866. doi:10.1007/s00216-014-8040-0
35. Taylor DD, Shah S. Methods of isolating extracellular vesicles impact down-stream analyses of their cargoes. Methods. 2015;87:3–10. doi:10.1016/j.ymeth.2015.02.019
36. Cui X, Lai W, Zhao Y, Chen C. The exosome-mediated cascade reactions for the transfer and inflammatory responses of fine atmospheric particulate matter in macrophages. Environ Sci Technol. 2023;57(21):7891–7901. doi:10.1021/acs.est.3c01436
37. Park KY. Adverse reactions following intradermal injection of exosome-based formulations: a case series. J Cosmet Dermatol. 2025;24(10):e70520. doi:10.1111/jocd.70520
38. Nochalabadi A, Khazaei M, Zamani S, Rezakhani L. The effect of decellularization processes on the immune response and compatibility of tissues. Tissue Cell. 2026;98:103135. doi:10.1016/j.tice.2025.103135
39. Stiliyanov-Atanasov K, Bagur-Cardona S, Chiera S, Capella-Monsonís H, Gomez-Florit M. Interfacing extracellular vesicles with bioengineered materials for regenerative medicine. Acta Biomater. 2025;208:20–37. doi:10.1016/j.actbio.2025.10.022
40. Zhang C, You J, Lin R, et al. Engineering self-assembled peek scaffolds with marine-derived exosomes and bacteria-targeting aptamers for enhanced antibacterial functions. J Funct Biomater. 2025;17(1). doi:10.3390/jfb17010023
41. Ju Y, Hu Y, Yang P, Xie X, Fang B. Extracellular vesicle-loaded hydrogels for tissue repair and regeneration. Mater Today Bio. 2023;18:100522. doi:10.1016/j.mtbio.2022.100522
42. Hu H, Yang J, Chen X, et al. Electrospun pla-gelatin composite scaffolds loaded with 3d-derived extracellular vesicles for repairing tympanic membrane perforation in rats. Int J Biol Macromol. 2025;323(Pt 2):147267. doi:10.1016/j.ijbiomac.2025.147267
43. Chen C, Chang ZH, Yao B, et al. 3d printing of interferon γ-preconditioned nsc-derived exosomes/collagen/chitosan biological scaffolds for neurological recovery after tbi. Bioact Mater. 2024;39:375–391. doi:10.1016/j.bioactmat.2024.05.026
44. Drury JL, Mooney DJ. Hydrogels for tissue engineering: scaffold design variables and applications. Biomaterials. 2003;24(24):4337–4351. doi:10.1016/s0142-9612(03)00340-5
45. Muir VG, Burdick JA. Chemically modified biopolymers for the formation of biomedical hydrogels. Chem Rev. 2021;121(18):10908–10949. doi:10.1021/acs.chemrev.0c00923
46. Henriques-Antunes H, Cardoso RMS, Zonari A, et al. The kinetics of small extracellular vesicle delivery impacts skin tissue regeneration. ACS Nano. 2019;13(8):8694–8707. doi:10.1021/acsnano.9b00376
47. Smyth T, Petrova K, Payton NM, et al. Surface functionalization of exosomes using click chemistry. Bioconjug Chem. 2014;25(10):1777–1784. doi:10.1021/bc500291r
48. Xing Y, Yerneni SS, Wang W, Taylor RE, Campbell PG, Ren X. Engineering pro-angiogenic biomaterials via chemoselective extracellular vesicle immobilization. Biomaterials. 2022;281:121357. doi:10.1016/j.biomaterials.2021.121357
49. Capella-Monsonís H, Crum RJ, D’Angelo W, Hussey GS, Badylak SF. Matrix-bound nanovesicles promote prohealing immunomodulation without immunosuppression. Tissue Eng Part A. 2025;31(17–18):1121–1131. doi:10.1089/ten.tea.2024.0238
50. Mu W, Rana S, Zöller M. Host matrix modulation by tumor exosomes promotes motility and invasiveness. Neoplasia. 2013;15(8):875–887. doi:10.1593/neo.13786
51. Sung BH, Ketova T, Hoshino D, Zijlstra A, Weaver AM. Directional cell movement through tissues is controlled by exosome secretion. Nat Commun. 2015;6:7164. doi:10.1038/ncomms8164
52. Rhim WK, Woo J, Kim JY, et al. Multiplexed PLGA scaffolds with nitric oxide-releasing zinc oxide and melatonin-modulated extracellular vesicles for severe chronic kidney disease. J Adv Res. 2025;69:75–89. doi:10.1016/j.jare.2024.03.018
53. Chabria Y, Duffy GP, Lowery AJ, Dwyer RM. Hydrogels: 3d drug delivery systems for nanoparticles and extracellular vesicles. Biomedicines. 2021;9(11). doi:10.3390/biomedicines9111694
54. Zhang C, Wu Y, Wang Y, et al. Bioengineering of extracellular vesicles with scaffold proteins for drug delivery. J Nanobiotechnol. 2026;24(1):160. doi:10.1186/s12951-026-04142-6
55. Liu Y, Kim G, Kim JY, et al. 3d bioprinted multifunctional gelma/tmp scaffold integrated with neural stem cell-derived extracellular vesicles and neural progenitor cells for spinal cord injury repair. J Tissue Eng. 2026;17:20417314261425659. doi:10.1177/20417314261425659
56. Zou J, Razali MH, Sun Z, Gao Y. Collagen-based hydrogel loaded with microspheres encapsulated with placental mesenchymal stem cells-derived exosomes synergistically promote recovery after spinal cord injury in rats. J Biomed Mater Res B Appl Biomater. 2026;114(1):e70014. doi:10.1002/jbmb.70014
57. Zheng D, Bai Y, Liu Z, et al. Immunoaffinity-mimetic assembly of peptide-aptamer conjugates and stem cell-derived exosomes into hierarchical microgels for spinal cord injury repair. Adv Sci. 2026;13(16):e19701. doi:10.1002/advs.202519701
58. Sun M, Li Q, Yu H, et al. Cryo-self-assembled silk fibroin sponge as a biodegradable platform for enzyme-responsive delivery of exosomes. Bioact Mater. 2022;8:505–514. doi:10.1016/j.bioactmat.2021.06.017
59. Xiang S, Qin T, Qin Y, et al. Angiopep-2-engineered extracellular vesicles derived from spinal cord injury-responsive brown adipose tissue for targeted neuroinflammation modulation. J Nanobiotechnol. 2026. doi:10.1186/s12951-026-04163-1
60. Németh K, Kazsoki A, Visnovitz T, et al. Nanofiber formation as a promising technology for preservation and easy storage of extracellular vesicles. Sci Rep. 2022;12(1):22012. doi:10.1038/s41598-022-25916-6
61. Zhang N, Yin Y, Xu SJ, Wu YP, Chen WS. Inflammation & apoptosis in spinal cord injury. Indian J Med Res. 2012;135(3):287–296.
62. Cheng J, Chen Z, Liu C, et al. Bone mesenchymal stem cell-derived exosome-loaded injectable hydrogel for minimally invasive treatment of spinal cord injury. Nanomedicine. 2021;16(18):1567–1579. doi:10.2217/nnm-2021-0025
63. Lee HH, Lien SE, Alpizar Vargas V, et al. Controlled spatial and temporal release of neurotrophic growth factor from genetically modified tissue engineering living scaffolds. J Tissue Eng. 2025;16:20417314251389333. doi:10.1177/20417314251389333
64. Larrea Murillo L, Chen Z, Song J, et al. Blood Vessels bioengineered from induced pluripotent stem cell derived mesenchymal stem cells and porous silk fibroin coated functional scaffolds. J Tissue Eng. 2025;16:20417314251355723. doi:10.1177/20417314251355723
65. Hu J, Lang Y, Cao Y, Zhang T, Lu H. The neuroprotective effect of tetramethylpyrazine against contusive spinal cord injury by activating pgc-1α in rats. Neurochem Res. 2015;40(7):1393–1401. doi:10.1007/s11064-015-1606-1
66. Fan X, Zang C, Lao K, Mu XH, Dai S. Neuroprotective effects of tetramethylpyrazine on spinal cord injury-related neuroinflammation mediated by p2x7r/nlrp3 interaction. Eur J Pharmacol. 2024;964:176267. doi:10.1016/j.ejphar.2023.176267
67. Liu W, Liu Q, Li Z, et al. Multifunctional magneto-electric and exosome-loaded hydrogel enhances neuronal differentiation and immunoregulation through remote non-invasive electrical stimulation for neurological recovery after spinal cord injury. Bioact Mater. 2025;48:510–528. doi:10.1016/j.bioactmat.2025.02.034
68. Wang LS, Chung JE, Chan PP, Kurisawa M. Injectable biodegradable hydrogels with tunable mechanical properties for the stimulation of neurogenesic differentiation of human mesenchymal stem cells in 3d culture. Biomaterials. 2010;31(6):1148–1157. doi:10.1016/j.biomaterials.2009.10.042
69. Wu Y, Chen YX, Yan J, et al. Fabrication of conductive gelatin methacrylate-polyaniline hydrogels. Acta Biomater. 2016;33:122–130. doi:10.1016/j.actbio.2016.01.036
70. Tseng TC, Tao L, Hsieh FY, Wei Y, Chiu IM, Hsu SH. An Injectable, self-healing hydrogel to repair the central nervous system. Adv Mater. 2015;27(23):3518–3524. doi:10.1002/adma.201500762.
71. Fan L, Liu C, Chen X, et al. Exosomes-Loaded electroconductive hydrogel synergistically promotes tissue repair after spinal cord injury via immunoregulation and enhancement of myelinated axon growth. Adv Sci. 2022;9(13):e2105586. doi:10.1002/advs.202105586
72. Xue Y, Wang A, Liu F, et al. Extracellular vesicle-integrated core-shell nanofibers synergize multi-target healing mechanisms for accelerated repair of large-volume skin defects. Mater Today Bio. 2026;37:102889. doi:10.1016/j.mtbio.2026.102889.
73. Shiekh PA, Tariq U, Athmuri DN, et al. An off-the-shelf bioactive ev delivering cryogel orchestrating immunomodulatory reprogramming and angiogenesis for diabetic ulcer healing. Biomaterials. 2026;330:124053. doi:10.1016/j.biomaterials.2026.124053
74. Getova VE, Orozco-García E, Palmers S, Krenning G, Narvaez-Sanchez R, Harmsen MC. Extracellular vesicles from adipose tissue-derived stromal cells stimulate angiogenesis in a scaffold-dependent fashion. Tissue Eng Regen Med. 2024;21(6):881–895. doi:10.1007/s13770-024-00650-4
75. Harding CV, Heuser JE, Stahl PD. Exosomes: looking back three decades and into the future. J Cell Biol. 2013;200(4):367–371. doi:10.1083/jcb.201212113
76. Zaborowski MP, Balaj L, Breakefield XO, Lai CP. Extracellular vesicles: composition, biological relevance, and methods of study. Bioscience. 2015;65(8):783–797. doi:10.1093/biosci/biv084
77. Doyle LM, Wang MZ. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells. 2019;8(7). doi:10.3390/cells8070727
78. Gurung S, Perocheau D, Touramanidou L, Baruteau J. The exosome journey: from biogenesis to uptake and intracellular signalling. Cell Commun Signal. 2021;19(1):47. doi:10.1186/s12964-021-00730-1
79. Zhang Y, Liu Y, Liu H, Tang WH. Exosomes: biogenesis, biologic function and clinical potential. Cell Biosci. 2019;9:19. doi:10.1186/s13578-019-0282-2
80. Crescitelli R, Lässer C, Szabó TG, et al. Distinct rna profiles in subpopulations of extracellular vesicles: apoptotic bodies, microvesicles and exosomes. J Extracell Vesicles. 2013:2. doi:10.3402/jev.v2i0.20677.
81. D’Souza-Schorey C, Schorey JS. Regulation and mechanisms of extracellular vesicle biogenesis and secretion. Essays Biochem. 2018;62(2):125–133. doi:10.1042/ebc20170078
82. Escola JM, Kleijmeer MJ, Stoorvogel W, Griffith JM, Yoshie O, Geuze HJ. Selective enrichment of tetraspan proteins on the internal vesicles of multivesicular endosomes and on exosomes secreted by human b-lymphocytes. J Biol Chem. 1998;273(32):20121–20127. doi:10.1074/jbc.273.32.20121
83. Zöller M. Tetraspanins: push and pull in suppressing and promoting metastasis. Nat Rev Cancer. 2009;9(1):40–55. doi:10.1038/nrc2543
84. van Niel G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev Mol Cell Biol. 2018;19(4):213–228. doi:10.1038/nrm.2017.125
85. Menck K, Sivaloganathan S, Bleckmann A, Binder C. Microvesicles in cancer: small size, large potential. Int J Mol Sci. 2020;21(15). doi:10.3390/ijms21155373
86. Heijnen HF, Schiel AE, Fijnheer R, Geuze HJ, Sixma JJ. Activated platelets release two types of membrane vesicles: microvesicles by surface shedding and exosomes derived from exocytosis of multivesicular bodies and alpha-granules. Blood. 1999;94(11):3791–3799.
87. Di Vizio D, Morello M, Dudley AC, et al. Large oncosomes in human prostate cancer tissues and in the circulation of mice with metastatic disease. Am J Pathol. 2012;181(5):1573–1584. doi:10.1016/j.ajpath.2012.07.030
88. Morello M, Minciacchi VR, de Candia P, et al. Large oncosomes mediate intercellular transfer of functional microrna. Cell Cycle. 2013;12(22):3526–3536. doi:10.4161/cc.26539
89. Ghosh AK, Secreto CR, Knox TR, Ding W, Mukhopadhyay D, Kay NE. Circulating microvesicles in b-cell chronic lymphocytic leukemia can stimulate marrow stromal cells: implications for disease progression. Blood. 2010;115(9):1755–1764. doi:10.1182/blood-2009-09-242719
90. Balaj L, Lessard R, Dai L, et al. Tumour microvesicles contain retrotransposon elements and amplified oncogene sequences. Nat Commun. 2011;2:180. doi:10.1038/ncomms1180
91. Streubel B, Chott A, Huber D, et al. Lymphoma-specific genetic aberrations in microvascular endothelial cells in b-cell lymphomas. N Engl J Med. 2004;351(3):250–259. doi:10.1056/NEJMoa033153
92. Zhang Y, Liu D, Chen X, et al. Secreted monocytic mir-150 enhances targeted endothelial cell migration. Mol Cell. 2010;39(1):133–144. doi:10.1016/j.molcel.2010.06.010
93. Umezu T, Ohyashiki K, Kuroda M, Ohyashiki JH. Leukemia cell to endothelial cell communication via exosomal mirnas. Oncogene. 2013;32(22):2747–2755. doi:10.1038/onc.2012.295
94. Szczepanski MJ, Szajnik M, Welsh A, Whiteside TL, Boyiadzis M. Blast-derived microvesicles in sera from patients with acute myeloid leukemia suppress natural killer cell function via membrane-associated transforming growth factor-beta1. Haematologica. 2011;96(9):1302–1309. doi:10.3324/haematol.2010.039743
95. Okoye IS, Coomes SM, Pelly VS, et al. Microrna-containing t-regulatory-cell-derived exosomes suppress pathogenic t helper 1 cells. Immunity. 2014;41(1):89–103. doi:10.1016/j.immuni.2014.05.019
96. Luga V, Wrana JL. Tumor-stroma interaction: revealing fibroblast-secreted exosomes as potent regulators of wnt-planar cell polarity signaling in cancer metastasis. Cancer Res. 2013;73(23):6843–6847. doi:10.1158/0008-5472.Can-13-1791
97. Yang M, Chen J, Su F, et al. Microvesicles Secreted by macrophages shuttle invasion-potentiating micrornas into breast cancer cells. Mol Cancer. 2011;10:117. doi:10.1186/1476-4598-10-117
98. Théry C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses. Nat Rev Immunol. 2009;9(8):581–593. doi:10.1038/nri2567
99. Cocucci E, Racchetti G, Meldolesi J. Shedding microvesicles: artefacts no more. Trends Cell Biol. 2009;19(2):43–51. doi:10.1016/j.tcb.2008.11.003
100. Liu C, Xie W, Li X, Dong Z, Fu X. Intravesicular gelation of mesenchymal stromal cell-derived microvesicles for enhanced therapeutic angiogenesis in lower limb ischemia. J Mater Chem B. 2026;14(1):273–288. doi:10.1039/d5tb01411e
101. Song Y, Wang S, Mai H, et al. Endothelial progenitor cell-derived microvesicles therapy relieves myocardial infarction symptoms by altering left ventricular protein expression. Am J Transl Res. 2025;17(7):5332–5343. doi:10.62347/zjox1177
102. Liang Z, Luo Y, Lv Y. Mesenchymal stem cell-derived microvesicles mediate BMP2 gene delivery and enhance bone regeneration. J Mater Chem B. 2020;8(30):6378–6389. doi:10.1039/d0tb00422g
103. Sheng MH-C, Rundle CH, Lau K-HW. Microvesicles released by osteoclastic cells exhibited chondrogenic, osteogenic, and anti-inflammatory activities: an evaluation of the feasibility of their use for treatment of osteoarthritis in a mouse model. Cells. 2025;14(3):193. doi:10.3390/cells14030193
104. Deng L, Wang Y, Peng Y, et al. Osteoblast-derived microvesicles: a novel mechanism for communication between osteoblasts and osteoclasts. Bone. 2015;79:37–42. doi:10.1016/j.bone.2015.05.022
105. Ghanbari A, Rad F, Shahraki MH, Hosseini E, Barmak MJ, Zibara K. Human mesenchymal stem cells-derived microvesicles increase oligodendrogenesis and neurogenesis of cultured adult neural stem cells. Neurosci Lett. 2024;841:137951. doi:10.1016/j.neulet.2024.137951
106. Pu Y, Wu Q, Zhang Q, et al. Mesenchymal stem-cell-derived microvesicles ameliorate mptp-induced neurotoxicity in mice: a role of the gut-microbiota-brain axis. Psychopharmacology. 2023;240(5):1103–1118. doi:10.1007/s00213-023-06348-0
107. Shi P, Gao H, Cheng Z, et al. Static magnetic field-modulated mesenchymal stem cell-derived mitochondria-containing microvesicles for enhanced intervertebral disc degeneration therapy. J Nanobiotechnol. 2024;22(1):457. doi:10.1186/s12951-024-02728-6
108. Zhang L, Valencia CA, Dong B, Chen M, Guan PJ, Pan L. Transfer of micrornas by extracellular membrane microvesicles: a nascent crosstalk model in tumor pathogenesis, especially tumor cell-microenvironment interactions. J Hematol Oncol. 2015;8:14. doi:10.1186/s13045-015-0111-y
109. Guo S, Redenski I, Levenberg S. Spinal cord repair: from cells and tissue engineering to extracellular vesicles. Cells. 2021;10(8). doi:10.3390/cells10081872
110. Cocucci E, Meldolesi J. Ectosomes and exosomes: shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015;25(6):364–372. doi:10.1016/j.tcb.2015.01.004
111. Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med. 1996;183(3):1161–1172. doi:10.1084/jem.183.3.1161
112. Théry C, Amigorena S, Raposo G, Clayton A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol. 2006;3:
113. Chen C, Skog J, Hsu CH, et al. Microfluidic isolation and transcriptome analysis of serum microvesicles. Lab Chip. 2010;10(4):505–511. doi:10.1039/b916199f
114. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (misev2018): a position statement of the international society for extracellular vesicles and update of the misev2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750. doi:10.1080/20013078.2018.1535750
115. Cointe S, Judicone C, Robert S, et al. Standardization of microparticle enumeration across different flow cytometry platforms: results of a multicenter collaborative workshop. J Thromb Haemost. 2017;15(1):187–193. doi:10.1111/jth.13514
116. Valkonen S, van der Pol E, Böing A, et al. Biological reference materials for extracellular vesicle studies. Eur J Pharm Sci. 2017;98:4–16. doi:10.1016/j.ejps.2016.09.008
117. Feng Z, Hensley L, McKnight KL, et al. A pathogenic picornavirus acquires an envelope by hijacking cellular membranes. Nature. 2013;496(7445):367–371. doi:10.1038/nature12029
118. Yang Y, Han Q, Hou Z, Zhang C, Tian Z, Zhang J. Exosomes mediate hepatitis b virus (hbv) transmission and nk-cell dysfunction. Cell Mol Immunol. 2017;14(5):465–475. doi:10.1038/cmi.2016.24
119. Thakur A, Rai D. Global requirements for manufacturing and validation of clinical grade extracellular vesicles. J Liq Biopsy. 2024;6:100278. doi:10.1016/j.jlb.2024.100278
120. Primorac D, Brlek P, Bulić L, et al. Therapeutic potential of extracellular vesicles: from biogenesis, isolation and molecular characterization to addressing translational gaps and regulatory barriers. Int J Mol Sci. 2026;27(4). doi:10.3390/ijms27041676
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.