Back to Journals » International Journal of Nanomedicine » Volume 21
Apo-Transferrin Modified Natamycin-Loaded Liposomes Improve the Prognosis of Fungal Keratitis via Enhanced Antifungal and Anti-Inflammatory Activities
Authors Hu L ![]() , Luan J, Liu X, Zhang Z
, Luan J, Liu X, Zhang Z ![]() , Tong Z, Qu Y, Jiang Z
, Tong Z, Qu Y, Jiang Z ![]() , Lin J
, Lin J
Received 2 February 2026
Accepted for publication 29 April 2026
Published 28 May 2026 Volume 2026:21 595236
DOI https://doi.org/10.2147/IJN.S595236
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Xing Zhang
Luwen Hu,1,* Junjie Luan,1,* Xing Liu,1,2 Zhaoyang Zhang,1 Zhenglu Tong,1 Yutian Qu,1 Zhongtai Jiang,1 Jing Lin1
1Department of Ophthalmology, the Affiliated Hospital of Qingdao University, Qingdao, Shandong Province, 266003, People’s Republic of China; 2Department of Cataract, Eye Institute of Shandong First Medical University, Qingdao Eye Hospital of Shandong First Medical University, Qingdao, Shandong Province, 266003, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jing Lin, Department of Ophthalmology, the Affiliated Hospital of Qingdao University, Qingdao, Shandong Province, 266003, People’s Republic of China, Email [email protected]
Background: The proliferation and establishment of infections by pathogenic microorganisms require essential nutrients, such as host-derived iron. The sequestration of iron by the host represents an ancient defense mechanism against microbial invasion. Apo-Transferrin (Apo-Tf) is a promising and novel antimicrobial agent with intrinsic iron sequestration properties and therefore merits clinical investigation. Given the characteristics of natamycin (NAT) such as its low aqueous solubility and significant ocular irritation, we developed apo-Tf decorated liposomes encapsulating NAT for the treatment of fungal keratitis (FK).
Methods: NAT was encapsulated into liposomes using the lipid film hydration method. Natamycin-loaded transferrin-conjugated liposomes (NAT@TF-LP) were synthesized via the post-insertion technique and characterized through zeta-sizer analysis, morphology and Fourier transform infrared (FTIR) spectroscopy. The encapsulation efficiency, loading capacity, and in vitro release profile were determined by ultraviolet spectrophotometry. The cytotoxicity, ocular surface irritation, and systemic safety of liposomal formulations were evaluated in corneal epithelial cells and mice. In vitro studies investigated the antifungal properties of NAT@TF-LP, while in vivo experiments assessed eye surface retention and therapeutic efficacy in an FK mouse model.
Results: The prepared liposomes exhibited a well-defined nanostructure with uniform dimensions and apo-Tf was effectively conjugated to the surface, as verified by the emergence of characteristic amide bands in FTIR analysis. The loading efficiencies of NAT@LP and NAT@TF-LP were 5.6% and 3.53% respectively, with encapsulation efficiencies above 96%. Both formulations facilitated sustained NAT release and prolonged ocular surface retention. The in vitro release of NAT from NAT@TF-LPs followed a diffusion-controlled profile, exhibiting an excellent fit to both Higuchi and Korsmeyer–Peppas models. Additionally, they exhibited good biosafety profiles in both cytotoxicity and ocular irritation assays. In vivo, NAT@TF-LPs demonstrated superior therapeutic efficacy, resulting in a 15.4-fold reduction in corneal fungal burden (approximately 1.19-log reduction) compared to the free NAT group, alongside a marked decrease in the protein expression of pro-inflammatory cytokines, including IL-1β (65.0%), IL-6 (59.0%), and TNF-α (73.0%), with a 69.3% enhancement in anti-inflammatory IL-10 levels.
Conclusion: The apo-Tf functionalized liposomal system improves NAT ocular bioavailability and exhibits augmented antifungal and therapeutic efficacies. This enhancement is attributed to the combined benefits of the liposomal platform and the inherent antimicrobial properties of apo-Tf. This study highlights the promising potential of NAT@TF-LPs for the pharmacotherapy of FK. The diagram illustrates the preparation and application of functionalized liposomes for fungal studies. The top section shows the process of creating NAT@LP using film-ultrasonic dispersion with components including NATA, cholesterol, EPC, and DSPE-mPEG2000. Subsequently, the post-insertion of DSPE-mPEG2000-Tf (synthesized from Apo-Transferrin and DSPE-mPEG2000-MAL) into the preformed liposomes leads to the final Apo-Transferrin conjugated liposomes (NAT@TF-LP), followed by ultrafiltration. The bottom section depicts in vitro studies focusing on the impairment of conidial adherence and damage to fungal cell membranes and biofilms. The in vivo studies highlight fungal clearance and cytokine suppression.Diagram showing liposome preparation, in vitro and in vivo studies on fungal clearance and cytokine suppression.
Keywords: apo-transferrin, fungi, drug delivery, therapeutic effects, natamycin
Introduction
Fungal keratitis (FK), a severe sight-threatening corneal infection, is recognized as a major global cause of vision loss.1–3 Improper use of contact lenses and steroids are key contributors to its high incidence.4 The primary causative agents are the filamentous fungi Aspergillus spp. and Fusarium spp. and the yeast Candida spp.5 These fungal infections of the cornea often lead to poor clinical outcomes because of delayed microbiological identification, morphological variability in culture, and constrained drug efficacy and corneal penetration.6,7
The critical importance of iron for both microbial proliferation and virulence has been increasingly recognized in recent years. Exogenous iron is indispensable for the survival and proliferation of virtually all microbes, including pathogenic species that infect mammalian hosts.8–11 In several fungal species, including Aspergillus fumigatus, Cochliobolus heterostrophus and Gibberella zeae, the disruption of iron homeostasis impairs their sexual development and germination of conidia.12,13 Therefore, therapeutic approaches aimed at iron restriction have gained attention as a viable method for combating resistant infections. However, the development of small-molecule iron chelators into viable anti-infective therapies has proven challenging, partly due to their associated toxicity issues.14,15
In mammals, iron sequestration is primarily mediated by transferrin-family iron-binding proteins.16 Human transferrin (Tf), a 76-kDa glycoprotein, is primarily synthesized in the liver. Tf exists in different biochemical states, namely the iron-free (apo-Tf), monoferric, and iron-saturated (holo-Tf) forms.17 Apo-Tf has demonstrated potent cross-kingdom antimicrobial activity, inhibiting the growth of Gram-positive and Gram-negative bacteria, and the fungus Candida albicans.18–21 This iron-binding protein demonstrated synergy with rifampicin against S. aureus in vitro, reducing its MIC from 0.15 mg/L to 0.019 mg/L—an eight-fold reduction. Furthermore, apo-Tf (lacking iron) significantly suppressed the emergence of antibiotic resistance during the course of treatment.18 Some studies have explored the potential of administering exogenous Tf at supraphysiological levels as an iron-chelating therapeutic strategy for infections, which may overcome some limitations associated with small-molecule chelators.22 These findings provide evidence that microbial infections can be treated with apo-Tf by restricting metal availability. Given the broad-spectrum activity of apo-Tf, we sought to determine whether it could also restrict the growth of A. fumigatus in vitro and enhance therapeutic efficacy in FK.
In addition to its antimicrobial properties, apo-Tf can mitigate iron-induced oxidative stress and modulate the excessive inflammatory response, thereby conferring protective effects on organs and tissues.23–26 For instance, systemic iron levels in individuals with sickle cell disease (SCD) activate neutrophils to release neutrophil extracellular traps (NETs), directly promoting inflammation. The iron scavenger deferoxamine and apo-Tf significantly suppressed neutrophil extracellular trap (NET) formation in a majority of SCD serum samples.27 Furthermore, reducing levels of circulating iron by apo-Tf conferred protection against renal ischemia-reperfusion (I/R) injury by mitigating oxidative stress and inflammation, thereby preserving organ function.28 Apo-Tf could also alleviate hypoxia-ischemia-induced neuroinflammation through anti-apoptotic effects, preserve oligodendrocyte survival, and facilitate cerebral repair.24
The transferrin receptor (TfR) is widely distributed on the surfaces of most cells. Consequently, Tf has been explored as a ligand for functionalized drug delivery across biological barriers.29–31 The corneal epithelium constitutes a formidable obstacle to the anterior segment delivery of topical nanoparticles. Previous work has demonstrated TfR expression in human corneal epithelium.32,33 Nanoparticles functionalized with Tf have shown favorable uptake characteristics into various corneal layers, namely the epithelium, stroma and endothelium.32 Previous studies have also indicated that the transferrin-mediated liposomal system represents a promising platform to facilitate gene delivery to the corneal endothelium.34 Recently, transferrin-coated nanoparticles have shown potential in functionalized delivery to intestinal infection likely due to the overexpression of TfR during inflammation.35 Apo-Tf was selected for this functionalized system to leverage its intrinsic therapeutic adjuvant properties via iron sequestration, while exploring its role as a potential ligand to facilitate TfR-mediated corneal penetration.
Natamycin (NAT) is a natural tetraene macrolide antibiotic classified within the polyene antifungal family.36 Despite its global use, developing NAT formulations that offer enhanced bioavailability (BA) and reduced ocular irritation remains challenging. With a high molecular mass of 665.733 g/mol, NAT faces major delivery obstacles due to its low aqueous solubility and poor corneal permeability.37 Therefore, following topical application, NAT exhibits low ocular residence and minimal bioavailability (2%), thus requiring frequent dosing.38
Compared with free drugs or other delivery systems, liposomes (LPs) present the advantages of being food grade, biodegradable, biocompatible and non-toxic due to their natural phospholipid composition.39 Liposomal delivery systems have demonstrated efficacy in enhancing the therapeutic performance of antimicrobial agents, particularly antibiotics, by significantly improving their pharmacological activity, bioavailability, and stability.40–42 Many studies have reported the superior efficacy of liposomal systems against various pathogens, achieving enhanced bacterial killing even at antibiotic reduced doses.43,44 Both liposomal ciprofloxacin and amikacin have demonstrated superior efficacy over their non-encapsulated counterparts in treating lung infections, owing to their enhanced ability to infiltrate pre-formed biofilms and penetrate infected macrophages.45,46
No prior studies have described the preparation of apo-Tf decorated liposomes for the encapsulation of NAT. While various natamycin nanoformulations have been developed to enhance drug solubility and ocular bioavailability, most of these systems utilize pharmacologically inert materials. Although apo-Tf is known for its targeting capabilities, its intrinsic antimicrobial properties are less commonly exploited in nanocarrier design. In this study, our NAT@TF-LP platform represents a strategic shift toward bioactive functionalization by integrating the intrinsic antimicrobial properties of the carrier itself. The prepared liposomes were subjected to physicochemical characterization and stability evaluations. Furthermore, this investigation comprehensively evaluated the antifungal and anti-inflammatory potential of the liposomal system both in vitro and in vivo. Consequently, it presents a promising biomimetic nanotherapeutic platform for improving the clinical management of corneal fungal infections.
Materials and Methods
Materials
Cholesterol (ShFeng Biotechnology Co., Ltd).; Egg Phosphatidylcholine (EPC), DSPE-mPEG2000, and 2-Iminothiolane hydrochloride (Traut’s reagent) (Solarbio Co., Beijing, China); DSPE-mPEG2000-Maleimide (DSPE-mPEG2000-MAL) (Aladdin, Shanghai, China); Recombinant Human Apo-Transferrin Protein (Yeasen Biotechnology Co., Ltd., Shanghai, China); Natamycin (NAT) (Macklin Biochemical Co., Ltd., Shanghai, China).
Crystal violet (CV), Dimethyl sulfoxide (DMSO) from Solarbio Co. (Beijing, China); Dulbecco’s Modified Eagle Medium (DMEM), trypsin, penicillin–streptomycinfetal and bovine serum (FBS), from Gibco BRL (USA); Protein Concentration Determination Assay Kit from Solarbio; calcofluor white (CFW) stain (Sigma-Aldrich, Shanghai, China); Propidium Iodide (PI) (Leagene Biotechnology, Beijing, China); Cell Counting Kit-8 (CCK-8) (MCE, USA); ELISA kit for inflammatory factors (Elabscience, Wuhan, China); RNAiso Plus (TaKaRa, China); Prime Script RT Reagent Kit; and qPCR Master Mix from Vazyme (China) were utilized in this study.
Methods
Preparation of Natamycin-Loaded Liposomes
Natamycin Liposomes (NAT@LP) were prepared using the thin-film hydration method.47 Appropriate quantities of EPC, cholesterol, DSPE-mPEG2000, and NAT were weighed and dissolved in a mixed organic solvent at a mass ratio of 50:14:3:4. The mixture was transferred to a round-bottom flask after the addition of methanol/chloroform (1:2, v/v). Subsequent rotary evaporation under reduced pressure removed the organic solvent, affording a uniform thin lipid film on the flask’s inner wall. The flask was then removed, and the lipid film was hydrated with 15 mL of ultrapure water by vigorous vortex mixing. The mixture was sonicated for 5 min in an ice bath using a probe-type ultrasonicator at a power of 200 W with a pulse cycle of 3 s on’ and 2 s “off” to obtain the NAT@LP. The final concentration of NAT in the formulated suspension was 0.027% (w/w). Homogeneous liposomes were obtained with a 0.22 μm-filter (Millipore), used to sterilization.
Natamycin-Loaded Transferrin-Conjugated Liposomes (NAT@TF-LP) were prepared by the post-insertion method.48 A 10 mg/mL apo-Tf solution was mixed with 4 mg/mL Traut’s reagent at a molar ratio of 1:10 and incubated for 1 h with gentle shaking to allow thiolation. The thiolated apo-Tf was then purified via dialysis and subsequently reacted with DSPE-mPEG2000-MAL overnight at a 1:10 molar ratio to form the DSPE-mPEG2000-Tf conjugate.49 The preformed NAT@LP was incubated with DSPE-mPEG2000-Tf at 37 °C for 30 min to allow the hydrophobic lipid tails of the conjugate to spontaneously insert into the outer leaflet of the liposomal membrane, yielding NAT@TF-LP. Unbound apo-Tf was separated using a 100 kDa molecular weight cut-off ultrafiltration centrifuge tube (Merck Millipore, USA). The content of conjugated apo-Tf was then determined with a bicinchoninic acid (BCA) protein assay kit.50
The Physicochemical Characterization of the Liposomes
The mean zeta potential and particle size of LP, NAT@LP and NAT@TF-LP were measured at room temperature using a particle size/zeta analyzer (90Plus PALS, NanoBrook, American). The storage stability of NAT@LP and NAT@TF-LP was assessed through evaluation of key physicochemical parameters (particle size and zeta potential) on days 1, 3, 5, 7, and 14 under refrigerated conditions (4°C). The different preparations of liposomes were stained with phosphotungstic acid and visualized using transmission electron microscopy (TEM, JEOL-1200EX).
The chemical structures and successful surface functionalization of the liposomes were evaluated using FTIR spectroscopy. Briefly, the lyophilized samples (including free apo-Tf, NAT@LPs, and NAT@TF-LPs) were mixed with spectroscopic-grade potassium bromide (KBr) and pressed into thin pellets. The spectra were then recorded using a Nicolet iS5 FTIR spectrometer (Thermo Fisher Scientific, USA). All measurements were performed over a scanning range of 4000 to 500 cm−1 at room temperature.
NAT Standard Curve, Encapsulation Efficiency(EE), and Loading Efficiency(LE)
The absorbance of different concentrations of NAT was measured at 304 nm using a UV−vis spectrophotometer.51 The resulting absorbance values, plotted against the concentration of NAT, were utilized to generate a standard curve. The EEs of both NAT@LP and NAT@TF-LP were assessed via ultrafiltration centrifugation. In this process, the NAT@LP formulation was five-fold diluted with methanol and sonicated for 15 minutes to ensure complete drug release. The NAT concentration was determined based on the absorbance measured at 304 nm, with reference to a pre-established standard curve. The total amount of drug, defined as m(NAT), was analyzed after methanol demulsification. To isolate the free drug, a sample of NAT@LP was placed in a ultrafilter (10 kDa MWCO). Subsequent centrifugation at 9000 rpm for 10 min forced the unencapsulated NAT through the membrane into the aqueous filtrate below.49 The amount of the nonencapsulated insoluble drug was recorded as m(free NAT). Similarly, the total content of the prepared NAT@TF-LP and the content of free drugs can be determined. Additionally, the total mass of liposomes (m(LPs)) was obtained by freeze-drying an aliquot of the liposomal suspension using a freeze dryer (LGJ-10FD, SongYuan, Beijing, China).
The drug EE and LE were calculated as follows:
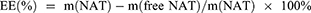
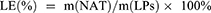
In vitro Drug Release Study
To assess the in vitro drug release profile, a dynamic dialysis model was employed.51,52 10 mL of free NAT or NAT@LP or NAT@TF-LP were filled into a 3500 kD MWCO dialysis bag (Solarbio, China), respectively. The dialysate bag was subsequently immersed in 250 mL of PBS on an oscillator. We collected 1 mL of the dialysate at scheduled times, immediately replenishing the volume with PBS. The released drug concentration in these samples was determined by measuring absorbance at 304 nm. The cumulative release percentage was calculated using the standard formula: (amount of drug released / total drug loaded)×100%.
To further investigate the drug release mechanism and kinetics of liposomes, the cumulative release data were fitted into several mathematical models, including the Zero-order 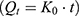 , First-order
, First-order 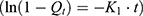 , Higuchi
, Higuchi 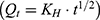 , and Korsmeyer–Peppas
, and Korsmeyer–Peppas 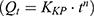 models. In these equations,
models. In these equations, 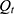 represents the cumulative percentage of NAT released at time t, while
represents the cumulative percentage of NAT released at time t, while 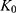 ,
, 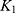 ,
, 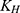 , and
, and 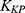 correspond to the respective release rate constants. For the Korsmeyer–Peppas model, the diffusion exponent (n) was utilized to characterize the transport mechanism. The model exhibiting the highest coefficient of determination (R2) was identified as the most suitable model for describing the NAT release kinetics.
correspond to the respective release rate constants. For the Korsmeyer–Peppas model, the diffusion exponent (n) was utilized to characterize the transport mechanism. The model exhibiting the highest coefficient of determination (R2) was identified as the most suitable model for describing the NAT release kinetics.
Evaluation of the Ocular Surface Retention Ability
The retention of the various formulations (NAT, NAT@LP, and NAT@TF-LP) on the ocular surface was assessed by a fluorescence-based method. Briefly, fluorescein sodium was used as a tracer and mixed with each formulation at a 1:5 mass ratio. A 5 μL dose of the drug mixture was applied via instillation into the mouse conjunctival sac. After gently closing the eye for 10 seconds, ocular surface clearance was monitored under cobalt blue light at 0, 1, 3, 5, 10, and 25 min by tracking the decrease in fluorescence intensity.
Preparation of A. fumigatus
Conidial and hyphal preparations of A. fumigatus (CGMCC 3.14869) were obtained following an established method.53 Conidial suspensions were obtained by washing Sabouraud’s agar cultures with PBS supplemented with 0.1% Tween 20. Conidia were resuspended following centrifugation and adjusted to target concentrations. To prepare hyphae, fungal cultures were harvested, ground into 20–40 μm fragments, and washed with PBS via centrifugation (4000×g, 40 min). The hyphal material was then inactivated using 75% ethanol, thoroughly rinsed with PBS, and finally resuspended in DMEM to the required concentration.
Detection of Antifungal Activity in vitro
To evaluate the antifungal efficacy, the minimum inhibitory concentration (MIC) was assessed. Liposomal formulations (NAT@LP or NAT@TF-LP) were introduced into wells containing A. fumigatus conidia at a density of 1×105 CFU/mL in Sabouraud medium. The final NAT concentrations, calculated from the drug loading data, followed a two-fold dilution series (0.25–16 μg/mL). After incubation for 48 hours at 37°C, the absorbance at 540 nm was recorded to quantify growth inhibition.
CFW was employed to stain the fungal cell wall, while PI was used to evaluate the extent of fungal cell membrane damage. After culturing at 37 ◦C for 32h, the hyphae treated with PBS, NAT, NAT@LP and NAT@TF-LP were subsequently stained with CFW or PI for 10 min. Then, the fungus exhibited a brilliant blue fluorescence (CFW) or a red fluorescence (PI), which was observed under a fluorescence microscope (Zeiss, Germany).
The biofilm formation was determined from the CV uptake of live A. fumigatus. The drug concentration and culture time of each group was the same as mentioned above. Following a 15-min incubation with 0.1% crystal violet, the samples were destained with 95% ethanol for 3 minutes, and the absorbance at 570 nm was recorded.
Conidial adherence to human corneal epithelial cells (HCECs, obtained from Xiamen Eye Center of Xia Men University) was assessed using hematoxylin and eosin (HE) staining. HCECs were maintained under culture conditions standardized in prior work.53 Following a 12-hour culture on four-chamber slides, HCECs were exposed to conidial suspensions (5 × 105/mL) in PBS, NAT, NAT@LP, or NAT@TF-LP for 3 h at identical concentrations. Each chamber was then rinsed with sterile PBS to remove non-adherent conidia, air-dried, and fixed using 4% paraformaldehyde (Elabscience, USA). Subsequently, the samples were stained with HE according to standard protocols. Then, the samples were imaged under a microscope (Zeiss, 400×). For quantification, four independent microscopic fields were randomly selected for each group. The conidia adhering to the HCECs were manually counted, and the adhesion level was calculated as the average number of adherent conidia per HCEC.
SEM and TEM were employed to examine the impact of NAT, NAT@LP, and NAT@TF-LP on fungal microstructure. Following collection, hyphal samples were processed for analysis by primary fixation with 2.5% glutaraldehyde (4°C, 2 h), PBS rinsing, and post-fixation in 1% osmium tetroxide (4°C, 1 h). These fixed samples were then dehydrated using ethanol, critically point-dried with CO2, and gold-sputtered prior to observation under a JEOL JSM-840 SEM. Imaging was conducted at 2000×and 5000×magnifications (scale bars = 20 or 10 μm).53
Drug Safety Assessment
Ocular tolerance was assessed in normal mice via the Draize test to ensure species consistency with the in vivo keratitis model, while adhering to the 3Rs principles of animal welfare.47,54 NAT, NAT@LP, and NAT@TF-LP were prepared as eye drops in sterile PBS at the same NAT concentration and applied to the conjunctival sac four times daily. On days 1, 3, and 5, corneal epithelial damage was evaluated by corneal fluorescein staining (CFS) under a cobalt blue slit-lamp microscope.55 Following the treatment period, tissues were collected on day 5. Eyeballs were encased in optimal cutting temperature (OCT) compound and snap-frozen for sectioning, while the livers, kidneys, hearts, and spleens were also collected. Tissue sections (8 μm thick) were then prepared, fixed in acetone, and dehydrated using ethanol. The staining procedure involved hematoxylin for 5 min, a tap water wash, eosin counterstaining for 30s, and a distilled water rinse. Finally, these sections were dehydrated, cleared, mounted, and imaged under a light microscope. HE staining of the retina, liver, kidney, heart and spleen were performed to evaluate the in vivo biosafety of liposomes. In addition, CCK- 8 assay in HCECs was carried out to assess the toxicity effect of liposomes with different drug concentrations. Following plating and culture to 80–90% confluence, HCECs were treated with formulations for 24 h. Cell viability was assessed using a CCK-8 assay, with absorbance measurement at 450 nm after a 2 h incubation.
Evaluation of Drug Efficacy in vivo
All experimental protocols involving animals were conducted in compliance with the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research. The mice were anesthetized with 8% chloral hydrate (0.4 mL/kg). Following anesthesia, the FK mouse model was established using our established method.56 Mouse cornea was infected with A. fumigatus conidia (1 × 107/mL). Corneas were treated with 3 μL of PBS, NAT (16 μg/mL) or NAT@LP or NAT@TF-LP four times per day and then observed using slit-lamp microscope on days 1, 3, and 5 post-infection. The severity of FK was graded based on established clinical scoring criteria.55 For euthanasia, mice were first rendered unconscious in a carbon dioxide chamber and then subjected to cervical dislocation. The corneas of mouse were separated with a surgical blade at 3 days post-infection Isolated corneas were homogenized in 1 mL of sterile PBS using a TissueLyser (QIAGEN, Germany). A 50 μL aliquot from each homogenate was subjected to culture on Sabouraud agar (28°C, 48 h) to observe fungal growth. The viable fungal burden on the corneas was determined by enumerating colony-forming units (CFU) using the plate count method (n = 3/group).
HE staining was conducted to facilitate a clear and intuitive observation of the integrity and changes in the cornea. On day 3 post-infection, mouse corneas from different treatment groups were harvested, embedded in OCT compound, frozen, and subsequently processed for HE staining as described above.
IL-1β, IL-6, TNF-α and IL-10 expression in murine corneal tissues was quantified by RT-PCR and ELISA (n = 3/group). Following centrifugation of homogenized lysates, the supernatant was treated with chloroform. Total RNA was then isolated from the aqueous phase by isopropyl alcohol precipitation. The resulting pellet was washed, air-dried, and resuspended in DEPC-treated water for concentration/purity measurement on a Nanodrop ND1000 (Thermo Fisher). Finally, cDNA was synthesized using a PrimeScript RT kit and RT-PCR was carried out as described previously,51 with primers listed in Table S1. The same method was used to collect the supernatant of corneal tissues. Concentrations of IL-1β, IL-6, TNF-α and IL-10 proteins were determined with specific ELISA kits (Biolegend, CA, USA) following the provided instructions.51
Statistical Analysis
Statistical analysis was performed using GraphPad Prism 8.0, with data reported as mean ± SD. Group differences were assessed by unpaired t‑test (two groups) or one-way ANOVA (≥ three groups). A P-value below 0.05 was considered significant (*P < 0.05, **P < 0.01, ***P < 0.001). All experiments included a minimum of three independent replicates.
Results and Discussion
Identification and Characterization of Properties of Liposomes
As shown in Figure 1A, the mean hydrodynamic diameters of the three liposomal formulations—plain liposomes (LPs), natamycin-loaded liposomes (NAT@LPs), and transferrin-conjugated natamycin-loaded liposomes (NAT@TF-LPs)—were 144.3 ± 2.88 nm, 174.7 ± 6.92 nm, and 205.5 ± 1.27 nm, respectively. Furthermore, the particle sizes of all formulations remained stable throughout the 14-day observation period (Figure 1B). The incorporation of apo-transferrin resulted in a moderate increase in liposome size, which can be attributed to the successful surface conjugation of the protein. Notably, the polydispersity index (PDI) showed no significant variation across all groups (Figure 1C), suggesting that the functionalization process did not compromise the stability of the liposomes. Zeta potential measurements further verified the surface modification. The values for LPs, NAT@LPs and NAT@TF-LPs were −16.23 ± 1.28 mV, −16.08 ± 3.12 mV, and −20.06 ± 3.19 mV, respectively (Figure 1D). Importantly, the zeta potential of NAT@TF-LPs was significantly more negative compared to NAT@LPs (P < 0.05). The more negative zeta potential of NAT@TF-LPs is consistent with the presence of negatively charged apo-Tf on the liposomal surface, aligning with previous reports on protein-decorated nanocarriers.57 After 14 days of continuous observation, all drug-loaded formulations exhibited a narrow particle size distribution with PDI values below 0.25 (Figure 1E and F). This finding suggests that storage at 4 °C effectively maintains the physical integrity of the liposomes, potentially minimizing premature drug leakage. Characterization confirmed that the lipid vesicles were uniform in size and composition, indicating their suitability for subsequent applications.58 Consequently, these results confirm the stability of the prepared liposomes.59,60 The morphology and structure of LPs, NAT@LPs and NAT@TF-LPs can be visualized by TEM (Figure 1G). The images indicated that all prepared formulations were well-dispersed and exhibited spherical nanostructures, consistent with those of previous studies.61 Importantly, TEM analysis could also confirm the presence of apo-Tf on the liposomal surface directly at the molecular level.50 Transferrin-conjugated liposomes clearly displayed a particulate surface coating that was absent in unconjugated preparations. FTIR spectroscopy was employed to verify the successful covalent conjugation of apo-Tf onto the liposomes (Figure 1H). The spectrum of free apo-Tf exhibited characteristic Amide I (1676 cm−1) and Amide II (1522 cm−1) bands,62 alongside a broad O-H/N-H stretching peak at 3538 cm−1. Plain NAT@LP displayed typical phospholipid markers,63 including lipid hydrocarbon C-H (2936 cm−1), ester C=O (1730 cm−1), and phosphate group vibrations (1239 cm−1 and 1073 cm−1), with no detectable absorption at the Amide II position. In the NAT@TF-LP spectrum, all fundamental lipid peaks were preserved, indicating that the liposomal structural integrity remained intact after functionalization. Crucially, prominent Amide I (1655 cm−1) and Amide II (1522 cm−1) signals emerged. The distinct appearance of the Amide II band provides direct evidence for the presence of apo-Tf on the liposomes, which aligns with established FTIR structural confirmations of apo-transferrin conjugated to lipidic platforms.64 Additionally, the narrowing and enhancement of the peak at 516 cm−1 is attributed to the C-S stretching vibration, which is consistent with the anticipated thioether bond formation via the maleimide-thiol reaction. Collectively, these spectral features confirm the successful functionalization of the NAT@TF-LP nanoplatform.
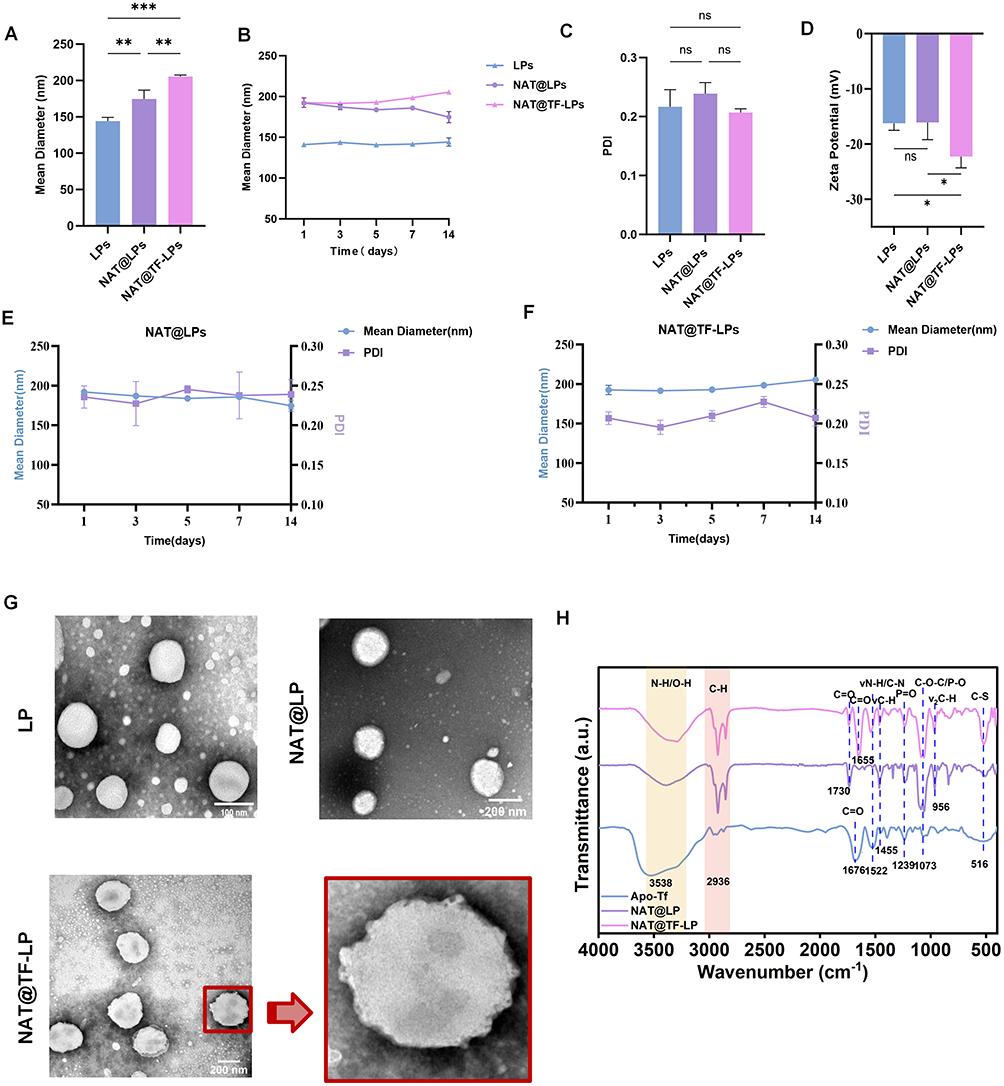 |
Figure 1 Physicochemical characterization of liposomal formulations. (A–C) Hydrodynamic diameter and PDI of plain LPs (blue), NAT@LPs (purple), and NAT@TF-LPs (pink) during 14-day storage at 4 ◦C. (D) Zeta potential of plain LPs, NAT@LPs, and NAT@TF-LPs. (E and F) Temporal changes in particle size and PDI of NAT@LPs and NAT@TF-LPs over 14 days, as measured by dynamic light scattering. (G) Representative TEM images illustrating the morphology and structural integrity of liposomes. (H) FTIR spectra of apo-Tf, NAT@LP, and NAT@TF-LP. ns represents no statistical significance; *P < 0.05, **P < 0.01,***P < 0.001. |
Drug Loading and Release from Liposomes
The linearity of NAT was established over a concentration range of 0–200 μg/mL, and the correlation coefficient R2 value was 0.999 (Figure 2A). Based on this standard curve, the encapsulation efficiency and loading efficiency were determined. The loading efficiencies of NAT@LP and NAT@TF-LP were 5.6% and 3.53% respectively, while the encapsulation efficiencies reached 96.85% and 96.3%. The stability of these parameters before and after functionalization indicates that no detectable NAT leakage occurred. Both NAT@LP and NAT@TF-LP exhibited high encapsulation efficiencies and comparable loading efficiency values. Such a disparity is an inherent trait of lipid-based nanocarriers engineered for hydrophobic or amphiphilic drugs. Since these molecules predominantly localize within the restricted space of the lipid bilayer, a substantial molar excess of structural components is mandated to provide adequate volume for NAT molecules and prevent precipitation.65,66 This structural requirement inherently establishes a physical upper limit on the overall loading efficiency. A parallel formulation strategy is evident in the clinically approved liposomal amphotericin B (AmBisome®), which maintains structural integrity by employing a distinctly low drug-to-lipid mass ratio of 0.143 mg/mg.67 Furthermore, the incorporation of cholesterol (optimally at 30–40 mol%) exerts an ordering effect on the phospholipid molecules.68,69 This ordering effect decreases bilayer permeability and prevents the uncontrolled leakage of the encapsulated natamycin.70 Surface modification with PEG-derivatives improves the overall pharmaceutical stability of the lipid-based nanocarriers.71 We evaluated the in vitro release of NAT from the liposomal formulations in PBS (Figure 2B). Compared to the rapid release observed with free NAT, both NAT@LPs and NAT@TF-LPs exhibited a more sustained release pattern. To further understand the release mechanism, the data were fitted to mathematical models. The release profile of NAT@TF-LPs best fit the Korsmeyer-Peppas model (R2 = 0.985) with a release exponent (n) of 0.54, closely followed by the Higuchi model (R2 = 0.962). These kinetic parameters demonstrate that the sustained release of NAT is primarily governed by a diffusion-controlled mechanism across the stable lipid bilayer, which aligns with recent kinetic evaluations of surface-functionalized ocular liposomes.72 Such controlled release behavior is highly advantageous for ocular drug delivery, as it prevents drug precipitation and extends the residence time of NAT on the ocular surface, potentially enhancing corneal penetration and improving overall bioavailability.73
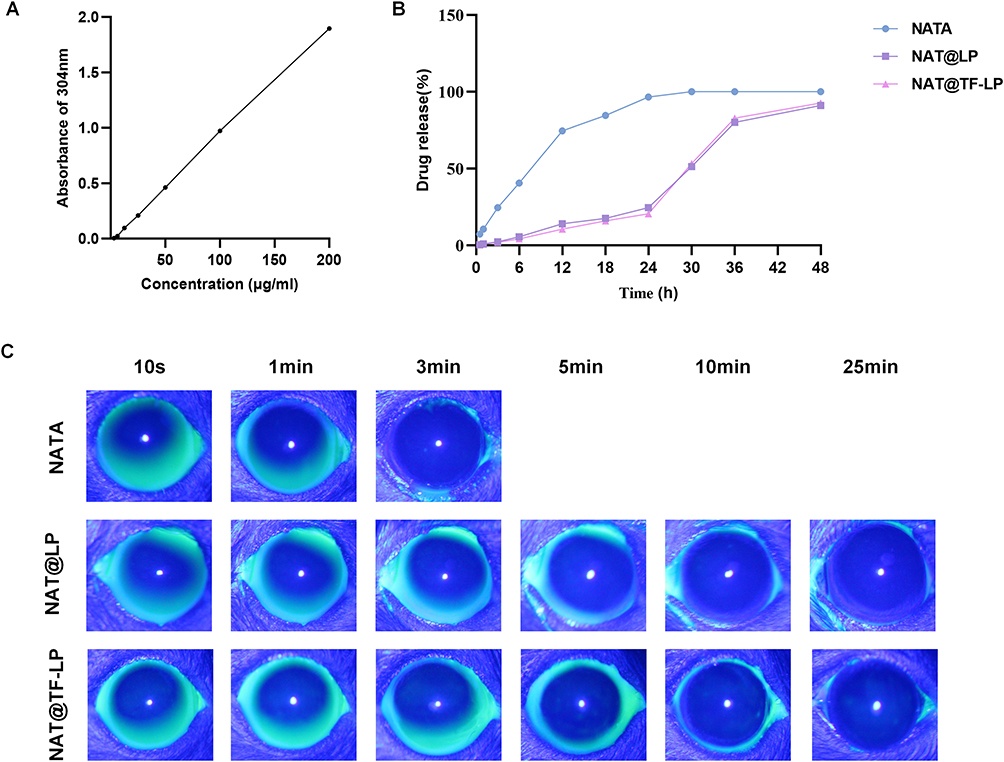 |
Figure 2 In vitro drug release and ocular retention assessment. (A) Standard curve of NAT based on UV–vis absorbance at 304 nm. (B) Cumulative release profiles of free NAT, NAT@LPs, and NAT@TF-LPs. (C) Ocular surface retention of fluorescein-labeled NAT, NAT@LPs, and NAT@TF-LPs in mice. |
Evaluation of Ocular Surface Retention
Physiological factors, such as the blinking reflex and tear secretion, lead to the rapid clearance of NAT from the ocular surface following administration. This phenomenon contributes to the short precorneal residence time of the drug, consequently reducing its topical bioavailability. Moreover, this rapid clearance poses challenges for dosage optimization, including the frequency of application and the risk of overdose.74 The therapeutic efficacy of commercially available antifungal suspensions is constrained by their short ocular residence and low bioavailability. Consequently, strategies aimed at prolonging ocular surface retention have emerged as a central approach to improving drug absorption.75 In this study, we evaluated the ocular surface retention ability of different formulations using anterior segment fluorescence imaging under cobalt blue illumination. Fluorescence of the NAT disappeared within 5 minutes after administration (Figure 2C). However, the fluorescence of NAT@LP and NAT@TF-LP completely disappeared after approximately 25 minutes. These results clearly demonstrate that liposomal formulations significantly prolong precorneal retention compared to the free drug solution. The extended residence time can be attributed to the exterior surfactant shell of liposomes, which facilitates sustained interactions with ocular tissues.76
Antifungal Effects of NAT@LPs and NAT@TF-LPs
The antifungal activity of the liposomal formulations was evaluated by determining the minimum inhibitory concentration (MIC) using absorbance readings at 540 nm. Owing to the sustained-release properties of liposomes, early-stage treatment of low-concentration liposomal NAT (0.25 µg/mL) did not exhibit significant antifungal effects compared to free NAT at equivalent concentrations (Figure 3A–F). Notably, apo-transferrin-functionalized liposomes (NAT@TF-LPs) achieved over 90% fungal growth inhibition at a concentration of 4 µg/mL (Figure 3G). At this concentration, the antifungal efficacy of NAT@TF-LPs was markedly higher than that of free NAT at the same concentration (4 µg/mL) and comparable to that of 8 µg/mL free NAT. Therefore, a concentration of 4 µg/mL was selected as the working concentration for subsequent in vitro experiments. Additionally, our study confirmed that apo-transferrin alone effectively inhibits the growth of A. fumigatus in vitro at relatively low concentrations (45 µg/mL) (Figure 3H). This observation is consistent with previous reports that the supplementary addition of apo-transferrin can effectively inhibit the growth of A. fumigatus by scavenging labile iron in the environment.77 Based on the BCA assay, the concentration of conjugated apo-Tf in the NAT@TF-LPs (at 4 µg/mL NAT) was 79.3 µg/mL. The antifungal efficacy of NAT@TF-LPs was significantly superior to that of NAT@LPs (non-functionalized liposomes containing the same 4 µg/mL of NAT but no apo-Tf). These results indicate that the combined presence of NAT and apo-Tf in the functionalized liposomes leads to a more potent antimicrobial activity at a lower NAT dosage. Moreover, the inhibitory effect of NAT@TF-LPs on fungal growth was sustained for up to 48 hours (Figure 3I).
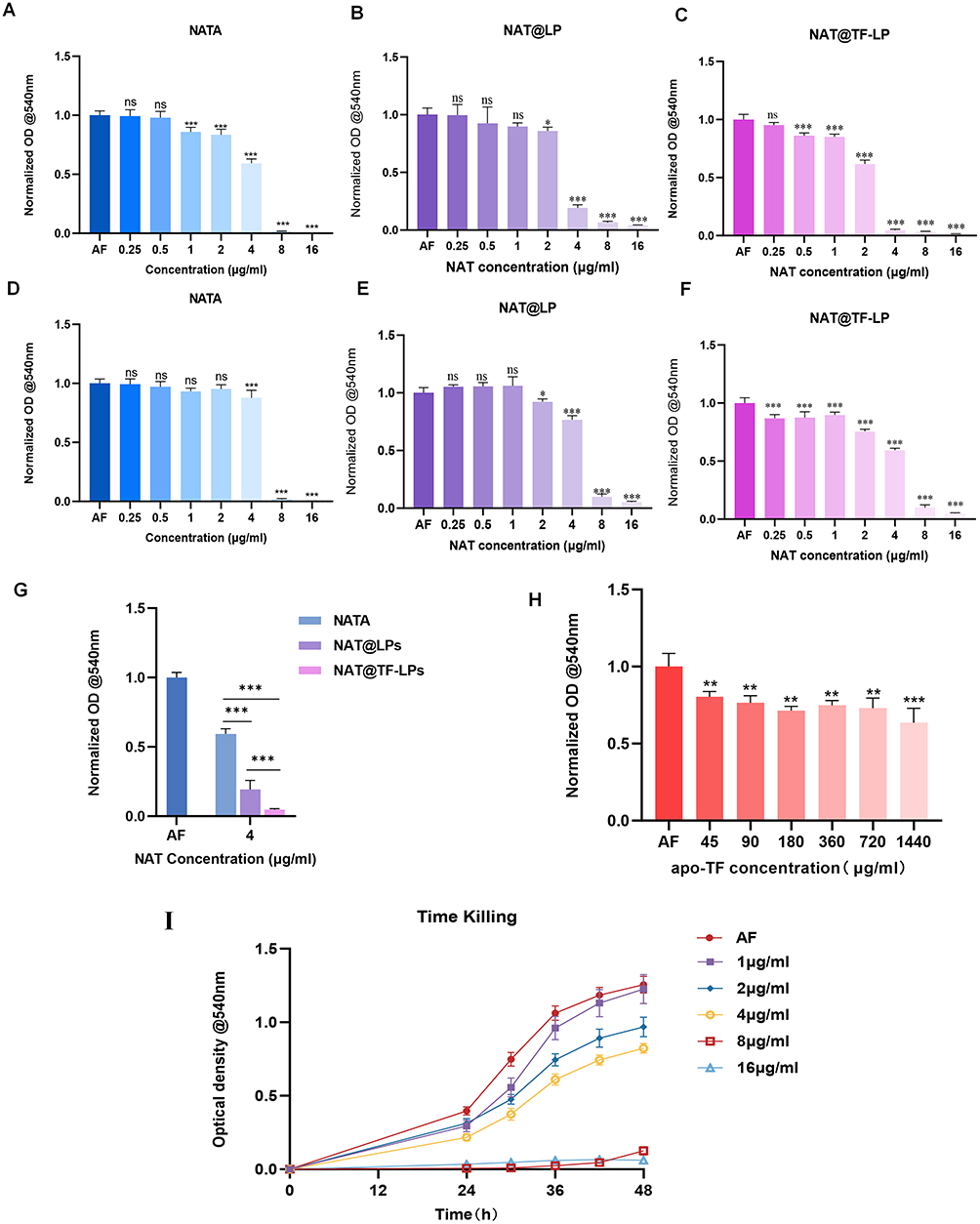 |
Figure 3 Antifungal activity of liposomal formulations against A. fumigatus conidia in vitro. MIC assays after 36 h (A–C) and 48 h (D–F) of treatment with NAT, NAT@LPs, or NAT@TF-LPs. (G) Comparison of antifungal efficacy between NAT, NAT@LPs, and NAT@TF-LPs at an equivalent NAT concentration of 4 μg/mL at 36 h. (H) MIC test of apo-Tf against A. fumigatus. (I) Time-kill kinetics of NAT@TF-LPs over 48h at varying concentrations. ns represents no statistical significance; *P < 0.05,**P < 0.01, ***P < 0.001. |
The integrity of fungal cell membranes is critical to cell viability.78 To evaluate membrane permeability, we performed PI staining, which revealed enhanced fluorescence intensity in fungal nuclei and chromosomal materials released from dead cells with compromised membranes. Notably, the NAT@TF-LPs group exhibited stronger fluorescence compared with the NAT and NAT@LPs groups (Figure 4A), indicating greater membrane disruption. Furthermore, CFW staining showed a marked reduction in hyphal growth of A. fumigatus after treatment with NAT@TF-LPs (Figure 4B), supporting the enhanced antifungal effect of the functionalized liposomes.
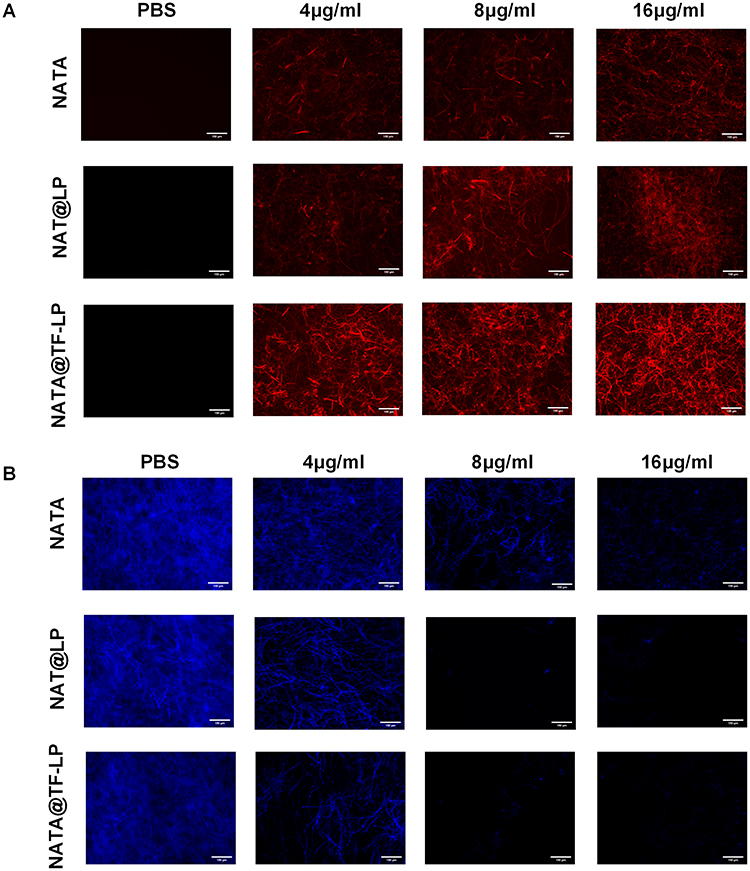 |
Figure 4 Evaluation of antifungal mechanisms in vitro by fluorescence and electron microscopy (scale bar =150 μm). (A) PI staining for membrane integrity. (B) CFW staining for cell wall morphology. |
The impact of the liposomal formulations on A. fumigatus was further examined by SEM and TEM to assess morphological alterations and damage to cell wall and membrane integrity (Figure 5A and B). SEM observations revealed that untreated hyphae maintained intact morphological structures with smooth surfaces. In contrast, hyphae treated with the liposomal formulations, particularly NAT@TF-LPs, exhibited obvious surface damage, including cracks and pores (indicated by black arrows). Furthermore, ultrastructural analysis by TEM corroborated these findings, revealing severe deformities in hyphae treated with NAT@TF-LPs. These included indistinct cell walls, membrane fragmentation, and disorganized cytoplasmic contents, which appeared more pronounced than the effects observed with NAT@LPs.
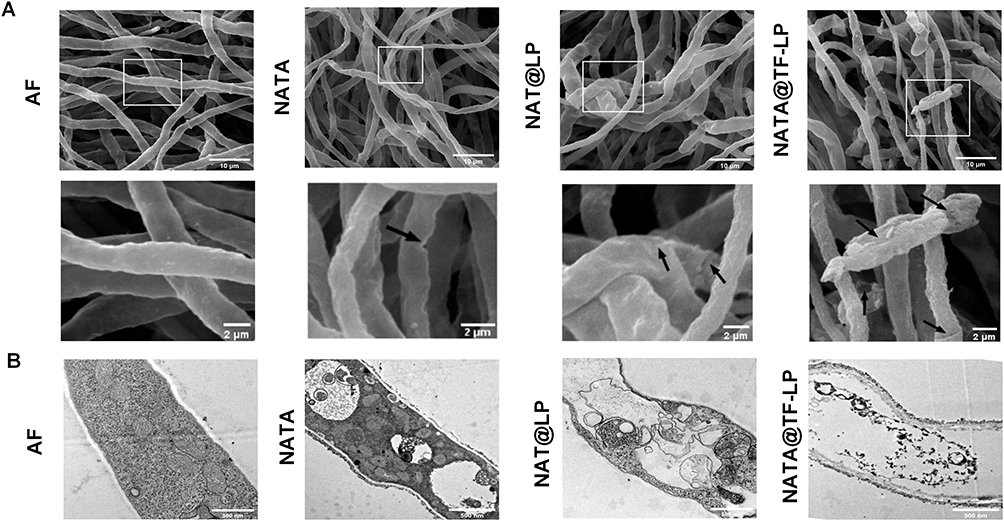 |
Figure 5 SEM (A; scale bars = 10 μm and 2 μm) and TEM (B; scale bar = 500 nm) images of A. fumigatus hyphae following treatment with PBS or liposomal formulations. |
The initial phase of FK involves the breach of the corneal epithelium, the primary defensive barrier of the cornea, which facilitates fungal colonization. Fungal adhesion to host epithelial cells represents a key step in infection establishment.79 Additionally, biofilm formation significantly enhances fungal tolerance to antifungal agents and promotes immune evasion.80 In this study, both NAT@LPs and NAT@TF-LPs decreased the adhesion of conidia to host cells (Figure 6A–F) and demonstrated a superior ability to inhibit biofilm formation (Figure 6G) compared to free NAT.
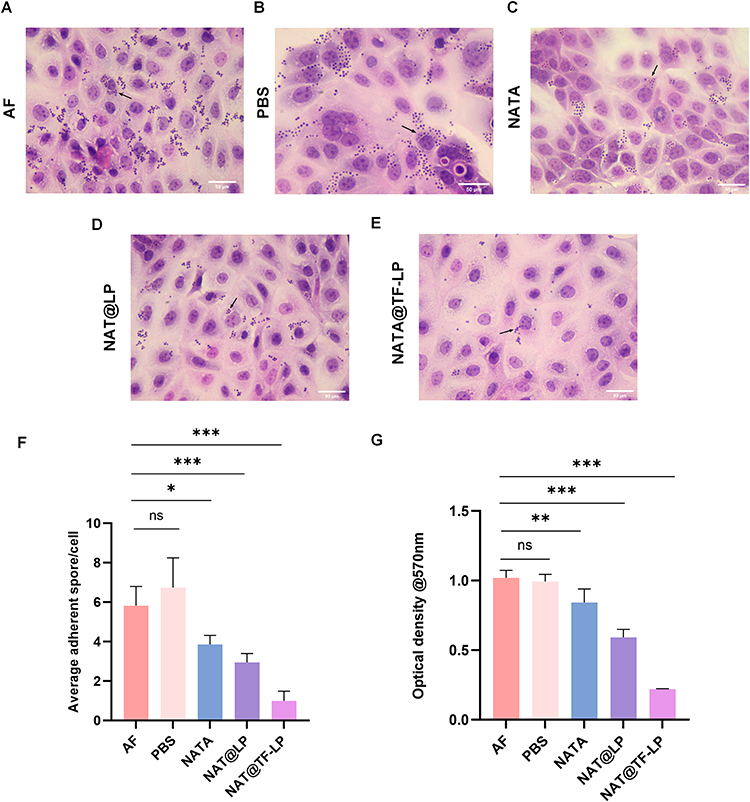 |
Figure 6 Fungal adhesion and biofilm formation on HCECs. HE staining of HCECs co-cultured with A. fumigatus conidia under different treatments: (A) conidia only, (B) PBS, (C) NAT, (D) NAT@LPs, (E) NAT@TF-LPs. (F) Quantitative analysis of adherent conidia per HCEC. (G) Biofilm formation assessed by crystal violet assay. ns represents no statistical significance; *P < 0.05,**P < 0.01, ***P < 0.001. |
Biocompatibility Evaluation of NAT@LPs and NAT@TF-LPs in vitro and in vivo
Currently marketed antifungal suspensions suffer from low bioavailability, high dosing frequency, and ocular irritation and discomfort during prolonged treatment of FK. Despite these limitations, NAT remains the drug of choice for treating FK.81 Liposomes functionalized with apo-transferrin provide a viable approach to address the limitations of conventional NAT formulations, potentially reducing side effects and ocular toxicity. In this study, equivalent concentrations of NAT were used as controls. CCK-8 assay results showed that neither NAT@LPs nor NAT@TF-LPs, at concentrations up to 32 µg/mL, significantly inhibited the viability of HCECs in comparison with the control. Conversely, a cytotoxic effect was observed with free NAT, which decreased cell viability to under 90% at 16 µg/mL and the cytotoxicity exhibited a clear concentration-dependent pattern (Figure 7A–C). In summary, both NAT@LPs and NAT@TF-LPs demonstrated excellent biocompatibility under in vitro conditions.
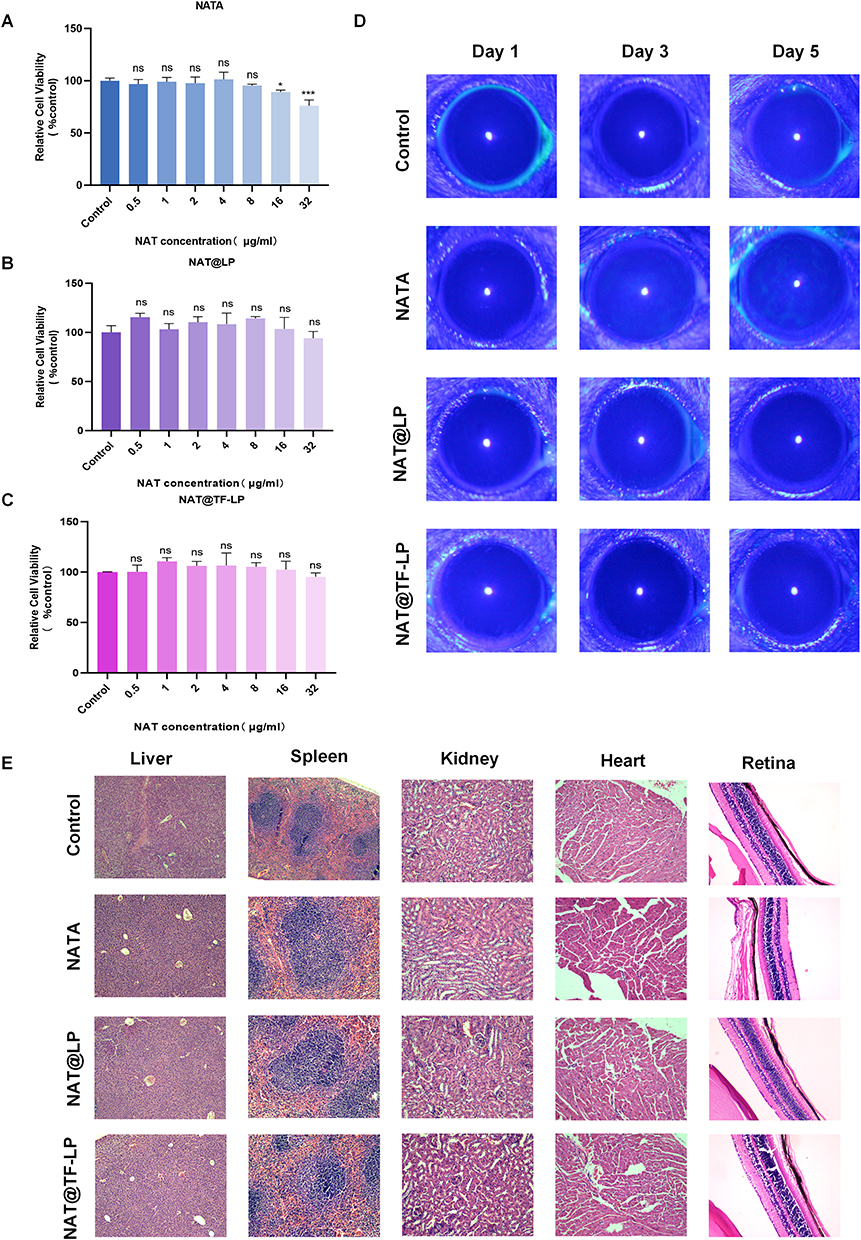 |
Figure 7 Biocompatibility assessment of liposomal formulations. (A–C) Viability of HCECs treated with NAT, NAT@LPs, or NAT@TF-LPs for 24h, measured by CCK-8 assay. (D) Representative corneal images from Draize eye tests in mice on days 1,3, and 5. (E) Histological sections of retina, heart, liver, spleen, and kidney tissues. ns represents no statistical significance; *P < 0.05,***P < 0.001. |
Draize eye tests were conducted to evaluate the ocular toxicity of NAT and liposomal formulations in mouse models. Healthy mice were treated with various drug formulations, and the ocular surface condition was monitored daily for five consecutive days using slit-lamp examination under cobalt blue light. Fluorescein sodium staining revealed no discernible fluorescence in any of the liposome-treated groups (Figure 7D). In contrast, mice treated with the same concentration of free NAT eye drops exhibited clear fluorescein staining in corneal tissue by the fifth day, indicating epithelial damage. These findings suggest that liposomal encapsulation significantly reduces the ocular toxicity associated with free NAT. Consistent with these observations, histological analysis of ocular sections (eg, retina) stained with HE showed an absence of pathological alterations in the eyes of treated mice compared to normal controls (Figure 7E). Furthermore, HE staining of major organs—including the liver, heart, spleen, and kidney—collected after the treatment period revealed no appreciable morphological differences between drug-treated and normal mice. No visible lesions were observed in any of the tissues examined. Collectively, these results demonstrate that apo-transferrin-modified liposomes exhibit high biosafety in vivo. Notably, unlike small-molecule iron chelators that often exhibit toxicity due to non-specific iron depletion, transferrin is an endogenous regulatory protein that maintains iron homeostasis without disrupting intracellular iron storage.22 Previous clinical studies have demonstrated that even high doses of exogenous apo-transferrin are well-tolerated in patients.82 Combined with our results, these findings suggest that NAT@TF-LPs do not adversely affect the iron homeostasis of the corneal microenvironment and exhibit high biosafety in vitro and vivo.
In vivo Efficacy of NAT@LPs and NAT@TF-LPs
The therapeutic efficacy of liposomal formulations was evaluated in a mouse model of FK. PBS and free NAT (16 μg/mL) served as controls alongside the liposomal formulations. Representative photographs (Figure 8A) showed that PBS-treated corneas developed severe opacity and swelling at 3 days post-infection. In comparison, corneas treated with free NAT exhibited moderate cloudiness, while those treated with NAT@LPs displayed noticeably reduced opacity and swelling. Notably, NAT@TF-LP treatment resulted in only minimal corneal opacity. Clinical scores, assessed using a standardized inflammation scoring system, further supported these observations (Figure 8B). In alignment with the improved clinical outcomes, NAT@TF-LPs achieved a substantial 15.4-fold reduction (approximately 1.19-log) in corneal fungal burden relative to the free NAT group (Figure 8C and D). These results highlight the potential of liposomes as effective drug delivery systems for ocular antifungal therapy. Composed of biodegradable and biocompatible phospholipid bilayers, liposomes are capable of encapsulating diverse therapeutic agents, which enhances both stability and bioavailability.39,73 In comparison with previously reported natamycin nanoformulations, such as conventional liposomes or PEGylated nanoparticles which primarily focus on enhancing drug solubility through passive carrier systems,71 our NAT@TF-LP platform represents a strategic functionalization. While existing inert delivery systems act solely as structural vehicles, the integration of apo-Tf into our liposomal surface introduces an adjunctive therapeutic dimension. Evidence from our in vitro studies confirms that the apo-Tf protein itself possesses inherent antimicrobial activity, which provides a complementary therapeutic contribution alongside the encapsulated NAT. This bioactive modification offers a distinct advantage over conventional formulations by potentially lowering the required therapeutic dose and improving the overall prognosis of FK, even when considering the biological barriers of the cornea for topical drug delivery. Additionally, it is recognized that TfR is expressed on the surface of most cell types,29–31 including the human corneal epithelium.32,33 Previous studies have shown that transferrin-functionalized nanoparticles are efficiently internalized across multiple corneal layers.32 Furthermore, TfR expression can be upregulated during inflammatory conditions,35 making apo-transferrin an especially promising ligand for functionalized drug delivery to infected corneas. While this may partly contribute to the observed efficacy, the precise molecular mechanisms require further specialized evaluation in future studies including the specific contribution of receptor-mediated pathways. From a clinical translation standpoint, the use of biodegradable phospholipids and a naturally occurring human protein (apo-Tf) ensures a favorable biocompatibility profile, potentially simplifying regulatory pathways. Future development could focus on optimizing large-scale manufacturing via microfluidic techniques and evaluating long-term storage stability, such as lyophilization, to meet clinical requirements for ocular anti-infective therapies. Furthermore, we acknowledge that the current in vivo evaluation serves as an exploratory proof-of-concept study. The relatively small sample size necessitates cautious interpretation, and future confirmatory studies with larger, prospectively powered cohorts are required to robustly validate these therapeutic outcomes. In conclusion, NAT@TF-LPs represent a promising strategy for the treatment of FK, achieving functionalization-enhanced ocular delivery of NAT and significantly improving its therapeutic efficacy.
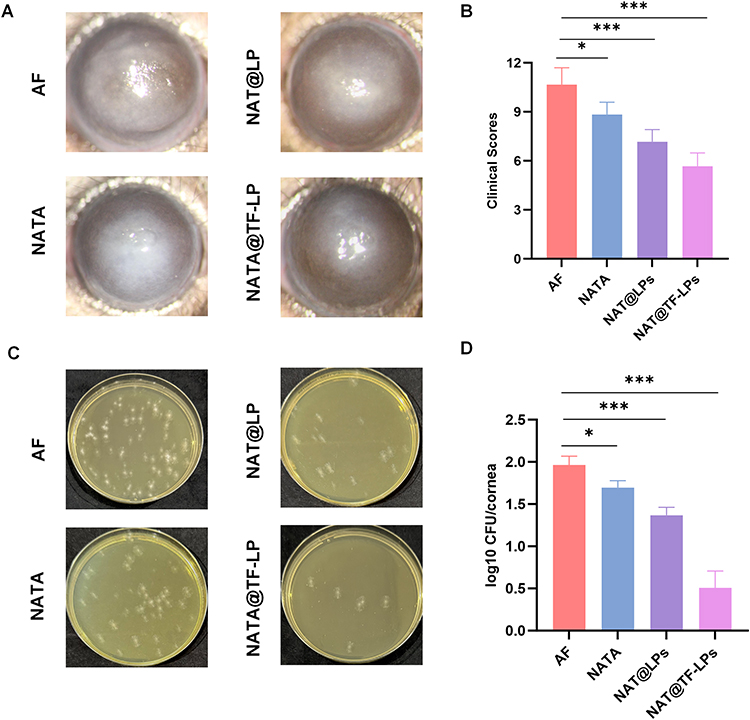 |
Figure 8 In vivo antifungal efficacy of liposomal formulations. (A) Slit-lamp photographs of infected corneas and (B) clinical scores of FK severity at 3 days post-infection after various treatments. (C and D) Representative images of corneal fungal burden assessed by plate counting. ns represents no statistical significance; *P < 0.05,***P < 0.001. |
Anti-Inflammatory Activity of Liposomes
Innate immunity represents the principal mechanism protecting the host from fungal infections. Nevertheless, an excessive inflammatory response can result in a poor FK prognosis by inducing corneal injury through elevated cytokine production. Therefore, it is crucial to eliminate the excessive production of proinflammatory cytokines in the cornea to alleviate FK.83 Notably, some studies have also demonstrated that iron overload could elevate inflammatory mediators, and this iron-mediated toxicity can be mitigated by iron-chelating agents such as transferrin.23,24 Building upon this rationale, several apo-transferrin-coated nanoparticles have been designed to provide a combined effect with therapeutic drugs.61,84,85 In our study, HE staining results illustrated that the NAT@TF-LPs group exhibited the most favorable prognosis at 3 days post-infection, characterized by less infiltration of inflammatory cells and tissue damage (Figure 9A). Treatment with NAT@TF-LPs significantly attenuated the overall inflammatory profile at both the mRNA and protein levels (Figure 9B–I). Specifically, compared to free NAT, the nanoplatform reduced the protein expression of key proinflammatory markers, including IL-1β (by 65.0%), IL-6 (by 59.0%), and TNF-α (by 73.0%), while concurrently upregulating the anti-inflammatory cytokine IL-10 by 69.3%. Consistent with previous studies,24,27,28 owing to the presence of the endogenous iron chelator apo-transferrin, NAT@TF-LPs effectively downregulated fungal-induced cytokine cascades, thereby alleviating inflammatory responses in FK. This integrated anti-inflammatory efficacy is likely derived from the combined contributions of each component: NAT indirectly limits inflammation by reducing the primary fungal stimulus, while apo-Tf modulates the inflammatory cascade as an endogenous iron chelator. The liposomal platform further sustains this therapeutic synergy by prolonging ocular retention. While NAT@TF-LPs demonstrate significant potential, the precise molecular contributions of TfR-mediated endocytosis versus iron sequestration remain to be quantified. Future studies employing receptor-blocking assays will be essential to further decouple these mechanisms.
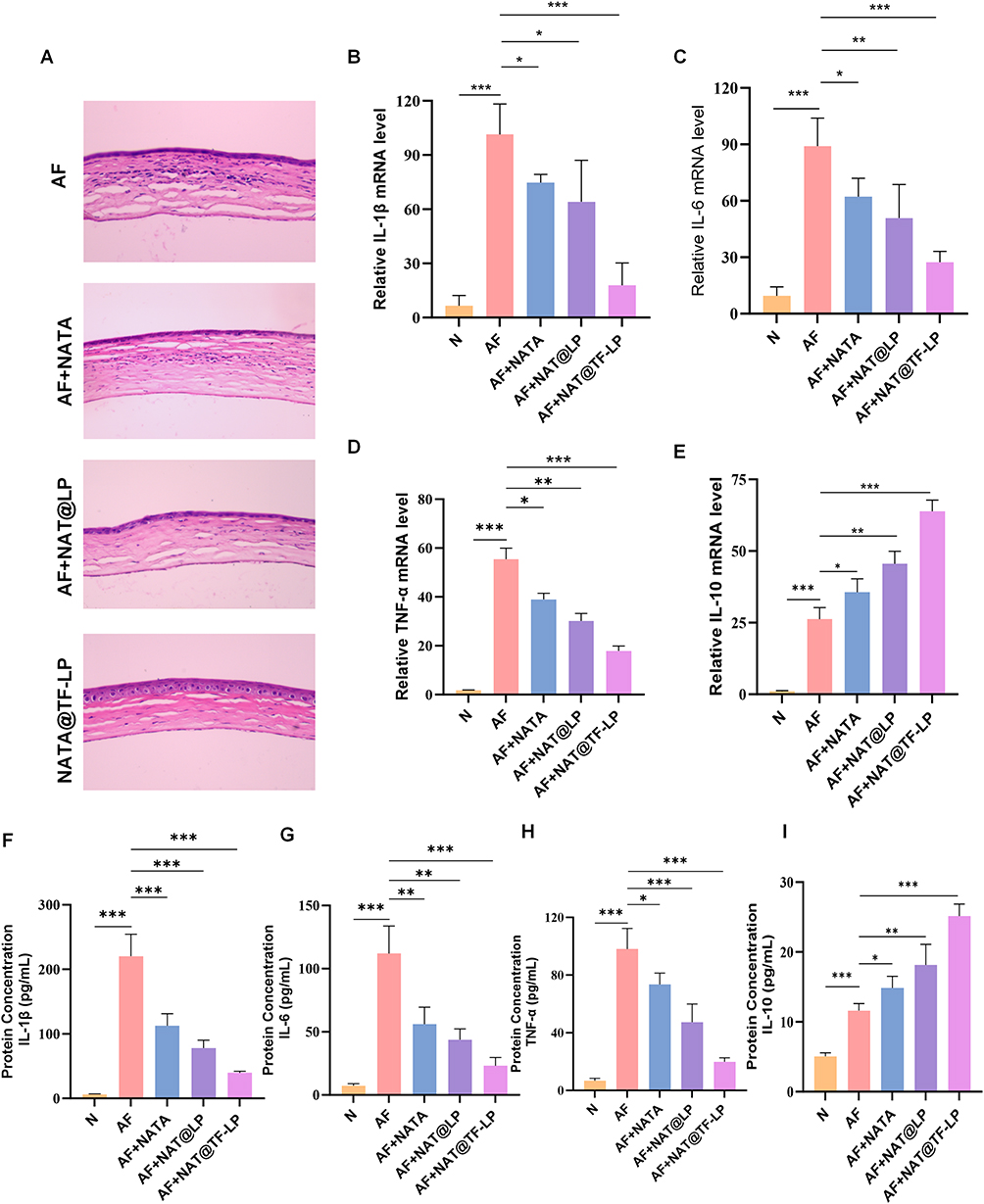 |
Figure 9 Anti-inflammatory effects of liposomal treatments in vivo. (A) HE-stained corneal sections from each treatment group at 3 days post-infection. (B–E) mRNA expression levels of IL-1β, IL-6, TNF-α, IL-10 in corneal tissues. (F–I) Protein levels of IL-1β, IL-6, TNF-α, IL-10 measured by ELISA. ns represents no statistical significance; *P < 0.05, **P < 0.01, ***P < 0.001. |
Conclusion
In this study, we designed a novel and highly efficient liposomal formulation for the treatment of FK by encapsulating NAT into apo-transferrin-functionalized liposomes. Apo-Tf itself exhibited strong antifungal activity at low concentrations. Meanwhile, liposomal encapsulation conferred sustained-release characteristics on NAT, thereby enhancing its bioavailability. Consequently, the NAT@TF-LP formulation demonstrated remarkable antifungal efficacy in vitro by inhibiting fungal growth, suppressing biofilm formation, and reducing conidial adhesion. In vivo experiments revealed that NAT@TF-LPs exhibited excellent biocompatibility. Treatment with NAT@TF-LPs significantly reduced corneal fungal burden, exhibited anti-inflammatory activity, and alleviated FK severity. In conclusion, apo-transferrin-conjugated liposomes for the delivery of NAT represent a promising biomimetic nanoplatform with great potential to enhance the clinical management and therapeutic efficacy of FK.
Ethics
All animal experiments were performed in full adherence to the principles outlined in the Basel Declaration and the ethical standards established by the International Council for Laboratory Animal Science and were approved by the Ethics Review Committee of the Affiliated Hospital of Qingdao University (QYFYWZLL 30540), Qingdao, Shandong, China.
Acknowledgment
This work was supported by the Natural Science Foundation of Shandong Province (No. ZR2023MH068). Graphical Abstract was drawn by the authors.
Funding
Supported by the Natural Science Foundation of Shandong Province (No. ZR2023MH068).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bisen AC, Sanap SN, Agrawal S, et al. Etiopathology, epidemiology, diagnosis, and treatment of fungal keratitis. ACS Infect Dis. 2024;10(7):2356–21. doi:10.1021/acsinfecdis.4c00203
2. Brown L, Leck AK, Gichangi M, Burton MJ, Denning DW. The global incidence and diagnosis of fungal keratitis. Lancet Infect Dis. 2021;21(3):e49–e57. doi:10.1016/S1473-3099(20)30448-5
3. Whitcher JP, Srinivasan M, Upadhyay MP. Corneal blindness: a global perspective. Bull World Health Organ. 2001;79(3):214–221.
4. Lalitha P, Prajna NV, Kabra A, Mahadevan K, Srinivasan M. Risk factors for treatment outcome in fungal keratitis. Ophthalmology. 2006;113(4):526–530. doi:10.1016/j.ophtha.2005.10.063
5. Niu L, Liu X, Ma Z, et al. Fungal keratitis: pathogenesis, diagnosis and prevention. Microb Pathog. 2020;138:103802. doi:10.1016/j.micpath.2019.103802
6. Pintye A, Bacsó R, Kovács GM. Trans-kingdom fungal pathogens infecting both plants and humans, and the problem of azole fungicide resistance. Front Microbiol. 2024;15:1354757. doi:10.3389/fmicb.2024.1354757
7. Ghannoum MA, Rice LB. Antifungal agents: mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin Microbiol Rev. 1999;12(4):501–517. doi:10.1128/CMR.12.4.501
8. Schaible UE, Kaufmann SHE. Iron and microbial infection. Nat Rev Microbiol. 2004;2(12):946–953. doi:10.1038/nrmicro1046
9. Schrettl M, Haas H. Iron homeostasis--Achilles’ heel of Aspergillus fumigatus? Curr Opin Microbiol. 2011;14(4):400–405. doi:10.1016/j.mib.2011.06.002
10. Holbein BE, Ang MTC, Allan DS, Chen W, Lehmann C. Iron-withdrawing anti-infectives for new host-directed therapies based on iron dependence, the Achilles’ heel of antibiotic-resistant microbes. Environ Chem Lett. 2021;19(4):2789–2808. doi:10.1007/s10311-021-01242-7
11. Sánchez-Jiménez A, Marcos-Torres FJ, Llamas MA. Mechanisms of iron homeostasis in Pseudomonas aeruginosa and emerging therapeutics directed to disrupt this vital process. Microb Biotechnol. 2023;16(7):1475–1491. doi:10.1111/1751-7915.14241
12. Misslinger M, Hortschansky P, Brakhage AA, Haas H. Fungal iron homeostasis with a focus on Aspergillus fumigatus. Biochim Biophys Acta Mol Cell Res. 2021;1868(1):118885. doi:10.1016/j.bbamcr.2020.118885
13. Wallner A, Blatzer M, Schrettl M, Sarg B, Lindner H, Haas H. Ferricrocin, a siderophore involved in intra- and transcellular iron distribution in Aspergillus fumigatus. Appl Environ Microbiol. 2009;75(12):4194–4196. doi:10.1128/AEM.00479-09
14. Vichinsky E. Clinical application of deferasirox: practical patient management. Am J Hematol. 2008;83(5):398–402. doi:10.1002/ajh.21119
15. Spellberg B, Ibrahim AS, Chin-Hong PV, et al. The deferasirox-AmBisome therapy for mucormycosis (DEFEAT Mucor) study: a randomized, double-blinded, placebo-controlled trial. J Antimicrob Chemother. 2012;67(3):715–722. doi:10.1093/jac/dkr375
16. Hood MI, Skaar EP. Nutritional immunity: transition metals at the pathogen-host interface. Nat Rev Microbiol. 2012;10(8):525–537. doi:10.1038/nrmicro2836
17. Kemp SF, Creech RH, Horn TR. Glycosylated albumin and transferrin: short-term markers of blood glucose control. J Pediatr. 1984;105(3):394–398. doi:10.1016/s0022-3476(84)80011-6
18. Lin L, Pantapalangkoor P, Tan B, et al. Transferrin iron starvation therapy for lethal bacterial and fungal infections. J Infect Dis. 2014;210(2):254–264. doi:10.1093/infdis/jiu049
19. Han Y. Apotransferrin has a second mechanism for anticandidal activity through binding of Candida albicans. Arch Pharm Res. 2014;37(2):270–275. doi:10.1007/s12272-013-0263-7
20. Han Y. The identification of surface interaction of apotransferrin with Candida albicans. Arch Pharm Res. 2014;37(10):1301–1307. doi:10.1007/s12272-013-0301-5
21. Luna BM, Ershova K, Yan J, et al. Adjunctive transferrin to reduce the emergence of antibiotic resistance in Gram-negative bacteria. J Antimicrob Chemother. 2019;74(9):2631–2639. doi:10.1093/jac/dkz225
22. Bruhn KW, Spellberg B. Transferrin-mediated iron sequestration as a novel therapy for bacterial and fungal infections. Curr Opin Microbiol. 2015;27:57–61. doi:10.1016/j.mib.2015.07.005
23. Vinchi F, Porto G, Simmelbauer A, et al. Atherosclerosis is aggravated by iron overload and ameliorated by dietary and pharmacological iron restriction. Eur Heart J. 2020;41(28):2681–2695. doi:10.1093/eurheartj/ehz112
24. Guardia Clausi M, Paez PM, Pasquini LA, Pasquini JM. Inhalation of growth factors and apo-transferrin to protect and repair the hypoxic-ischemic brain. Pharmacol Res. 2016;109:81–85. doi:10.1016/j.phrs.2016.01.010
25. Youale J, Bigot K, Kodati B, et al. Neuroprotective effects of transferrin in experimental glaucoma models. Int J Mol Sci. 2022;23(21):12753. doi:10.3390/ijms232112753
26. Bigot K, Gondouin P, Bénard R, et al. Transferrin non-viral gene therapy for treatment of retinal degeneration. Pharmaceutics. 2020;12(9):836. doi:10.3390/pharmaceutics12090836
27. Van Avondt K, Schimmel M, Bulder I, et al. Circulating iron in patients with sickle cell disease mediates the release of neutrophil extracellular traps. Transfus Med Hemother. 2023;50(4):321–329. doi:10.1159/000526760
28. de Vries B, Walter SJ, von Bonsdorff L, et al. Reduction of circulating redox-active iron by apotransferrin protects against renal ischemia-reperfusion injury. Transplantation. 2004;77(5):669–675. doi:10.1097/01.tp.0000115002.28575.e7
29. Guo Q, Qian C, Wang X, Qian ZM. Transferrin receptors. Exp Mol Med. 2025;57(4):724–732. doi:10.1038/s12276-025-01436-x
30. Li H, Qian ZM. Transferrin/transferrin receptor-mediated drug delivery. Med Res Rev. 2002;22(3):225–250. doi:10.1002/med.10008
31. Gandapu U, Chaitanya RK, Kishore G, Reddy RC, Kondapi AK. Curcumin-loaded apotransferrin nanoparticles provide efficient cellular uptake and effectively inhibit HIV-1 replication in vitro. PLoS One. 2011;6(8):e23388. doi:10.1371/journal.pone.0023388
32. Kompella UB, Sundaram S, Raghava S, Escobar ER. Luteinizing hormone-releasing hormone agonist and transferrin functionalizations enhance nanoparticle delivery in a novel bovine ex vivo eye model. Mol Vis. 2006;12:1185–1198.
33. Lauweryns B, van den Oord JJ, Missotten L. The transitional zone between limbus and peripheral cornea. An immunohistochemical study. Invest Ophthalmol Vis Sci. 1993;34(6):1991–1999.
34. Tan PH, King WJ, Chen D, et al. Transferrin receptor-mediated gene transfer to the corneal endothelium. Transplantation. 2001;71(4):552–560. doi:10.1097/00007890-200102270-00011
35. Reddy GA, Handa M, Garabadu D, Kumar R, Kushawaha PK, Shukla R. Transferrin decorated PLGA encumbered moxifloxacin nanoparticles and in vitro cellular studies. Drug Dev Ind Pharm. 2023;49(1):129–138. doi:10.1080/03639045.2023.2185463
36. Qiu S, Zhao GQ, Lin J, et al. Natamycin in the treatment of fungal keratitis: a systematic review and meta-analysis. Int J Ophthalmol. 2015;8(3):597–602. doi:10.3980/j.issn.2222-3959.2015.03.29
37. Brik H. New high-molecular decomposition products of natamycin (pimaricin) with intact lactone-ring. J Antibiot. 1976;29(6):632–637. doi:10.7164/antibiotics.29.632
38. Patil A, Majumdar S. Echinocandins in Ocular Therapeutics. J Ocul Pharmacol Ther. 2017;33(5):340–352. doi:10.1089/jop.2016.0186
39. Obuobi S, Julin K, Fredheim EGA, Johannessen M, Škalko-Basnet N. Liposomal delivery of antibiotic loaded nucleic acid nanogels with enhanced drug loading and synergistic anti-inflammatory activity against S. aureus intracellular infections. J Control Release. 2020;324:620–632. doi:10.1016/j.jconrel.2020.06.002
40. Ferreira M, Ogren M, Dias JNR, et al. Liposomes as antibiotic delivery systems: a promising nanotechnological strategy against antimicrobial resistance. Molecules. 2021;26(7):2047. doi:10.3390/molecules26072047
41. Drulis-Kawa Z, Dorotkiewicz-Jach A. Liposomes as delivery systems for antibiotics. Int J Pharm. 2010;387(1–2):187–198. doi:10.1016/j.ijpharm.2009.11.033
42. Gonzalez Gomez A, Hosseinidoust Z. Liposomes for antibiotic encapsulation and delivery. ACS Infect Dis. 2020;6(5):896–908. doi:10.1021/acsinfecdis.9b00357
43. Bassetti M, Vena A, Russo A, Peghin M. Inhaled liposomal antimicrobial delivery in lung infections. Drugs. 2020;80(13):1309–1318. doi:10.1007/s40265-020-01359-z
44. Nwabuife JC, Pant AM, Govender T. Liposomal delivery systems and their applications against Staphylococcus aureus and Methicillin-resistant Staphylococcus aureus. Adv Drug Deliv Rev. 2021;178:113861. doi:10.1016/j.addr.2021.113861
45. Meers P, Neville M, Malinin V, et al. Biofilm penetration, triggered release and in vivo activity of inhaled liposomal amikacin in chronic Pseudomonas aeruginosa lung infections. J Antimicrob Chemother. 2008;61(4):859–868. doi:10.1093/jac/dkn059
46. Blanchard JD, Elias V, Cipolla D, Gonda I, Bermudez LE. Effective treatment of mycobacterium avium subsp. hominissuis and mycobacterium abscessus species infections in macrophages, biofilm, and mice by using liposomal ciprofloxacin. Antimicrob Agents Chemother. 2018;62(10):e00440–18. doi:10.1128/AAC.00440-18
47. Wang K, Jiang L, Zhong Y, et al. Ferrostatin-1-loaded liposome for treatment of corneal alkali burn via targeting ferroptosis. Bioeng Transl Med. 2022;7(2):e10276. doi:10.1002/btm2.10276
48. Wang X, Zhao Y, Dong S, et al. Cell-penetrating peptide and transferrin co-modified liposomes for targeted therapy of glioma. Molecules. 2019;24(19):3540. doi:10.3390/molecules24193540
49. Liu X, Dong S, Dong M, et al. Transferrin-conjugated liposomes loaded with carnosic acid inhibit liver cancer growth by inducing mitochondria-mediated apoptosis. Int J Pharm. 2021;607:121034. doi:10.1016/j.ijpharm.2021.121034
50. Anabousi S, Laue M, Lehr CM, Bakowsky U, Ehrhardt C. Assessing transferrin modification of liposomes by atomic force microscopy and transmission electron microscopy. Eur J Pharm Biopharm. 2005;60(2):295–303. doi:10.1016/j.ejpb.2004.12.009
51. Liu X, Zhang Y, Peng F, et al. Macrophage membrane-coated nanoparticles for the delivery of natamycin exhibit increased antifungal and anti-inflammatory activities in fungal keratitis. ACS Appl Mater Interfaces. 2024;16(44):59777–59788. doi:10.1021/acsami.4c11366
52. Zhang Y, Liu X, Hou Y, et al. Platelet membrane-camouflaged PLGA loaded natamycin improve the prognosis of fungal keratitis. Biomater Adv. 2025;172:214239. doi:10.1016/j.bioadv.2025.214239
53. Liu W, Qi Y, Diao W, et al. Chelerythrine ameliorates Aspergillus fumigatus keratitis through suppressing the LOX-1/p38 MAPK signaling pathway. Cytokine. 2024;182:156717. doi:10.1016/j.cyto.2024.156717
54. Fu X, Tian X, Lin J, et al. Zeolitic imidazolate framework-8 offers an anti-inflammatory and antifungal method in the treatment of aspergillus fungal keratitis in vitro and in vivo. Int J Nanomed. 2024;19:11163–11179. doi:10.2147/IJN.S480800
55. Wu TG, Wilhelmus KR, Mitchell BM. Experimental keratomycosis in a mouse model. Invest Ophthalmol Vis Sci. 2003;44(1):210–216. doi:10.1167/iovs.02-0446
56. Chi M, Gu L, Zhang L, et al. Pentoxifylline treats Aspergillus fumigatus keratitis by reducing fungal burden and suppressing corneal inflammation. Eur J Pharmacol. 2023;945:175607. doi:10.1016/j.ejphar.2023.175607
57. Cheung TH, Xue C, Kurtz DA, Shoichet MS. Protein release by controlled desorption from transiently cationic nanoparticles. ACS Appl Mater Interfaces. 2023;15(44):50560–50573. doi:10.1021/acsami.2c19877
58. Danaei M, Dehghankhold M, Ataei S, et al. Impact of particle size and polydispersity index on the clinical applications of lipidic nanocarrier systems. Pharmaceutics. 2018;10(2):57. doi:10.3390/pharmaceutics10020057
59. Loureiro JA, Andrade S, Duarte A, et al. Resveratrol and grape extract-loaded solid lipid nanoparticles for the treatment of alzheimer’s disease. Molecules. 2017;22(2):277. doi:10.3390/molecules22020277
60. Andrade S, Loureiro JA, Pereira MC. Transferrin-functionalized liposomes for the delivery of gallic acid: a therapeutic approach for alzheimer’s disease. Pharmaceutics. 2022;14(10):2163. doi:10.3390/pharmaceutics14102163
61. Mao K, Zhang W, Yu L, Yu Y, Liu H, Zhang X. Transferrin-decorated protein-lipid hybrid nanoparticle efficiently delivers cisplatin and docetaxel for targeted lung cancer treatment. Drug Des Devel Ther. 2021;15:3475–3486. doi:10.2147/DDDT.S296253
62. Hadden JM, Haris PI, Srai KS, Chapman D. Conformational studies on human transferrin. Biochem Soc Trans. 1992;20(2):200S. doi:10.1042/bst020200s
63. De Leo V, Maurelli AM, Ingrosso C, Lupone F, Catucci L. Easy preparation of Liposome@PDA microspheres for fast and highly efficient removal of methylene blue from water. Int J Mol Sci. 2021;22(21):11916. doi:10.3390/ijms222111916
64. Amin HH, Meghani NM, Park C, et al. Fattigation-platform nanoparticles using apo-transferrin stearic acid as a core for receptor-oriented cancer targeting. Colloids Surf B Biointerfaces. 2017;159:571–579. doi:10.1016/j.colsurfb.2017.08.014
65. Torchilin VP. Recent advances with liposomes as pharmaceutical carriers. Nat Rev Drug Discov. 2005;4(2):145–160. doi:10.1038/nrd1632
66. Guimarães D, Cavaco-Paulo A, Nogueira E. Design of liposomes as drug delivery system for therapeutic applications. Int J Pharm. 2021;601:120571. doi:10.1016/j.ijpharm.2021.120571
67. Faustino C, Pinheiro L. Lipid systems for the delivery of amphotericin b in antifungal therapy. Pharmaceutics. 2020;12(1):29. doi:10.3390/pharmaceutics12010029
68. Róg T, Pasenkiewicz-Gierula M, Vattulainen I, Karttunen M. Ordering effects of cholesterol and its analogues. Biochim Biophys Acta. 2009;1788(1):97–121. doi:10.1016/j.bbamem.2008.08.022
69. Briuglia ML, Rotella C, McFarlane A, Lamprou DA. Influence of cholesterol on liposome stability and on in vitro drug release. Drug Deliv Transl Res. 2015;5(3):231–242. doi:10.1007/s13346-015-0220-8
70. Bouaoud C, Lebouille JGJL, Mendes E, De Braal HEA, Meesters GMH. Formulation and antifungal performance of natamycin-loaded liposomal suspensions: the benefits of sterol-enrichment. J Liposome Res. 2015;1–10. doi:10.3109/08982104.2015.1046079
71. Patil A, Lakhani P, Taskar P, et al. Formulation development, optimization, and in vitro–in vivo characterization of natamycin-loaded pegylated nano-lipid carriers for ocular applications. J Pharmaceut Sci. 2018;107(8):2160–2171. doi:10.1016/j.xphs.2018.04.014
72. Mirkani A, Nabid MR, Pakian S. Mucoadhesive cationic liposome nanoparticles coated with methacrylated hyaluronic acid for ocular drug delivery. ACS Appl Nano Mater. 2025;8(49):23370–23387. doi:10.1021/acsanm.5c03934
73. Gbian DL, Omri A. Lipid-based drug delivery systems for diseases managements. Biomedicines. 2022;10(9):2137. doi:10.3390/biomedicines10092137
74. Gu L, Li C, Lin J, et al. Drug-loaded mesoporous carbon with sustained drug release capacity and enhanced antifungal activity to treat fungal keratitis. Biomater Adv. 2022;136:212771. doi:10.1016/j.bioadv.2022.212771
75. Li S, Chen L, Fu Y. Nanotechnology-based ocular drug delivery systems: recent advances and future prospects. J Nanobiotechnology. 2023;21(1):232. doi:10.1186/s12951-023-01992-2
76. López-Cano JJ, González-Cela-Casamayor MA, Andrés-Guerrero V, Herrero-Vanrell R, Molina-Martínez IT. Liposomes as vehicles for topical ophthalmic drug delivery and ocular surface protection. Expert Opin Drug Deliv. 2021;18(7):819–847. doi:10.1080/17425247.2021.1872542
77. Petzer V, Wermke M, Tymoszuk P, et al. Enhanced labile plasma iron in hematopoietic stem cell transplanted patients promotes Aspergillus outgrowth. Blood Adv. 2019;3(11):1695–1700. doi:10.1182/bloodadvances.2019000043
78. Huang X, Dong Q, Zhou Q, et al. Genomics insights of candidiasis: mechanisms of pathogenicity and drug resistance. Front Microbiol. 2025;16:1531543. doi:10.3389/fmicb.2025.1531543
79. Moyes DL, Richardson JP, Naglik JR. Candida albicans-epithelial interactions and pathogenicity mechanisms: scratching the surface. Virulence. 2015;6(4):338–346. doi:10.1080/21505594.2015.1012981
80. Donovan C, Arenas E, Ayyala RS, Margo CE, Espana EM. Fungal keratitis: mechanisms of infection and management strategies. Surv Ophthalmol. 2022;67(3):758–769. doi:10.1016/j.survophthal.2021.08.002
81. Mascarenhas M, Chaudhari P, Lewis SA. Natamycin ocular delivery: challenges and advancements in ocular therapeutics. Adv Ther. 2023;40(8):3332–3359. doi:10.1007/s12325-023-02541-x
82. Sahlstedt L, von Bonsdorff L, Ebeling F, Ruutu T, Parkkinen J. Effective binding of free iron by a single intravenous dose of human apotransferrin in haematological stem cell transplant patients. Br J Haematol. 2002;119(2):547–553. doi:10.1046/j.1365-2141.2002.03836.x
83. Yuan K, Zhao G, Che C, et al. Dectin-1 is essential for IL-1β production through JNK activation and apoptosis in Aspergillus fumigatus keratitis. Int Immunopharmacol. 2017;52:168–175. doi:10.1016/j.intimp.2017.09.008
84. Zhu L, Yu T, Wang W, et al. Responsively degradable nanoarmor-assisted super resistance and stable colonization of probiotics for enhanced inflammation-targeted delivery. Adv Mater. 2024;36(18):e2308728. doi:10.1002/adma.202308728
85. Yang D, Feng Y, Yuan Y, et al. Protein coronas derived from mucus act as both spear and shield to regulate transferrin functionalized nanoparticle transcellular transport in enterocytes. ACS Nano. 2024;18(10):7455–7472. doi:10.1021/acsnano.3c11315
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.