Back to Journals » Drug Design, Development and Therapy » Volume 20
Antimicrobial Peptides as Novel Ecological Approaches to Caries Prevention
Authors Jiang W, Liu J, Chen A, Zhao T, Xu Z, Xu L, Du Y, Guan X, Ding J, Liu Y, Sun D ![]()
Received 30 December 2025
Accepted for publication 20 March 2026
Published 26 March 2026 Volume 2026:20 592732
DOI https://doi.org/10.2147/DDDT.S592732
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Anastasios Lymperopoulos
Wentao Jiang,1,2 Jintao Liu,1 Aijia Chen,2 Tongxin Zhao,2 Zhenkun Xu,3 Laijun Xu,4 Yijie Du,3 Xin Guan,3 Jingzhe Ding,3 Yuming Liu,3 Dongyuan Sun1
1School of Stomatology, Shandong Second Medical University, Affiliated Hospital of Shandong Second Medical University, Weifang, People’s Republic of China; 2Guanghua School of Stomatology, Sun Yat-Sen University, Guangzhou, People’s Republic of China; 3School of Stomatology, the National University of Malaysia, Kuala Lumpur, Malaysia; 4School of Stomatology, Changsha Medical University, Changsha, People’s Republic of China
Correspondence: Dongyuan Sun, School of Stomatology, Shandong Second Medical University, Affiliated Hospital of Shandong Second Medical University, Weifang, People’s Republic of China, Tel +0086-18328581142, Email [email protected]
Abstract: Antimicrobial strategies have consistently been a central theme in caries prevention research. Due to their potent antibacterial properties and the potential for multifunctionality through sequence modification, antimicrobial peptides (AMPs) are emerging as promising candidates for anticaries therapeutics. Traditional investigations primarily focused on the broad-spectrum bactericidal efficacy of AMPs to eradicate cariogenic bacteria; however, this approach may inadvertently harm the beneficial oral microbiome. With advancing knowledge of oral microecology, the focus has progressively shifted towards ecological approaches. These strategies aim to selectively modulate cariogenic populations or the disease-associated microbiota while preserving microbial diversity and maintaining ecological balance, as demonstrated in multispecies biofilm models and sequencing-based microbiome analyses. Current innovations include targeting specific pathogens, regulating virulence factors, utilizing bio-responsive mechanisms within the cariogenic microenvironment, and disrupting dental plaque biofilms. This article reviews the current progress and limitations of antimicrobial peptide-based ecological approaches to caries control and offers valuable insights for the future design and optimization of antimicrobial peptides with microecology regulating effects or other promising functions aiding caries prevention.
Keywords: antimicrobial peptides, dental caries, anticaries agents, ecological prevention of dental caries, multifunctional peptides
Background
Dental caries is among the most prevalent chronic diseases worldwide, leading to complications such as pain, tooth loss, and masticatory dysfunction, all of which significantly affect quality of life and impose considerable economic burdens.1 Under physiological conditions, cariogenic and commensal bacteria maintain a dynamic homeostasis, wherein transient acidogenic activity is effectively neutralized by the salivary buffering system and innate antimicrobial components.2 However, this ecological equilibrium can be disrupted by the convergence of multiple determinants, including frequent carbohydrate consumption, diminished salivary flow or buffering capacity, altered host immune responses, and innate variations in enamel composition and structure.3 Excessive carbohydrate intake and inadequate oral hygiene drive the accumulation of exopolysaccharides (EPS) and prolonged acid production. When coupled with insufficient salivary clearance and neutralization, this promotes the progressive acidification of the dental plaque biofilm. This process, heavily modulated by host susceptibility and microbial succession patterns, selects for highly aciduric and acidogenic microbiota, ultimately establishing a dysbiotic microenvironment. Consequently, when the localized drop in pH disrupts the delicate balance between demineralization and remineralization—a threshold further dictated by enamel solubility and salivary mineral content—the progressive destruction of dental hard tissues ensues (Figure 1). 2–7
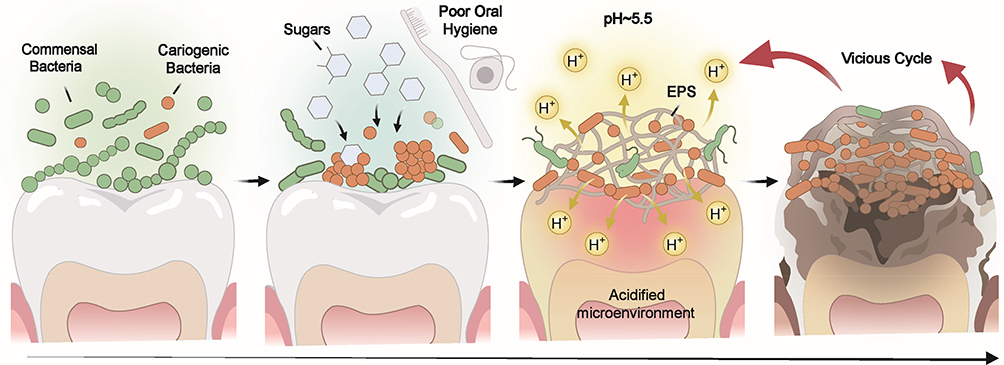 |
Figure 1 Ecological plaque hypothesis of dental caries. The development of dental caries is caused by the dysbiosis of oral microecology, the acidification of the plaque microenvironment, and the disequilibrium of enamel mineral homeostasis.4–6 |
Antimicrobial peptides (AMPs) can combat bacteria, viruses, and fungi.8 They generally disrupt bacterial cell membranes, leading to cell lysis, and pose a minimal risk of inducing drug resistance.9 Additionally, they can undergo conformational transitions to form nanonets or nanofibers, thereby preventing biofilm formation, and can inhibit cellular metabolism and translation through intracellular mechanisms.10–12 Previous research has primarily focused on their bactericidal efficacy against cariogenic bacteria and dental biofilms. These compounds have demonstrated antimicrobial activity against a wide array of oral bacteria.13 Their potential anti-caries effects are attributed to their broad-spectrum antimicrobial properties, which, however, result in a lack of specificity and may lead to microbial dysbiosis. A significant challenge associated with the use of AMPs is that their broad-spectrum nature may indiscriminately target both pathogenic and commensal bacteria, potentially causing unpredictable disruptions to the oral microbiota and adverse effects on host health.14,15 Consequently, researchers are advancing the development of novel AMPs to establish an effective and ecological strategy for the prevention and management of dental caries.13,16,17 These novel ecological antimicrobial strategies aim to achieve caries control while strictly preserving microbial diversity. This preservation is typically substantiated by complex experimental frameworks, such as saliva or dental plaque-derived biofilm models and in vivo studies, where microbiome sequencing analyses confirm that overall microbial diversity remains intact despite the suppression of cariogenic phenotypes and caries progression. Furthermore, advanced approaches actively restore a balanced microecology by selectively reducing the abundance of caries-associated pathogens without disrupting commensal populations. At a minimum, in vitro multispecies models must demonstrate the suppression of cariogenic bacteria or their virulence, without altering the overall proportion of commensal species.18–20 Beyond their antimicrobial efficacy and concerns about side effects on the oral microbiome, the applicability of AMPs is also constrained by their potential cytotoxic effects on host cells and their capacity to affect local immune responses. Uncontrolled or non-selective activity could lead to inflammation or soft tissue impairment.21–24 Therefore, the design of anticaries peptides must also focus on biocompatibility to preserve oral tissue health. Novel strategies, such as pH-triggered activation, where antimicrobial activity is enhanced specifically within the acidic cariogenic niche, offer a promising approach to minimize off-target effects on healthy oral tissues under neutral physiological conditions, ensuring both safety and ecological balance.25 In addition to biocompatibility issues, the clinical translation of AMPs is significantly hindered by their proteolytic instability in the complex oral environment. Notably, the oral cavity is replete with diverse proteases originating from saliva, host cells, and bacteria, which can rapidly degrade linear peptides. This rapid enzymatic cleavage leads to severely shortened half-lives and reduced therapeutic efficacy.26,27 Therefore, addressing this structural vulnerability is essential for the successful formulation of AMP-based therapeutics for caries prevention.
This literature review presents a comprehensive overview of some novel typical antimicrobial peptides with ecological properties and discusses future development strategies, aiming to inspire more researches in antimicrobial peptide-based caries prevention strategies.
Specifically Targeted Antimicrobial Peptides (STAMP): Eliminating Cariogenic Bacteria or Virulence Factors Precisely
AMPs exhibit significant structural diversity and exert their antimicrobial effects on a broad spectrum of extracellular and intracellular targets through various mechanisms.15 This diversity of mechanisms presents substantial opportunities for the design of AMPs that can selectively interact with specific microorganisms, leading to the development of STAMPs targeting cariogenic bacteria.
Usually, STAMPs incorporate a species-targeting peptide domain conjoined with an antimicrobial killing peptide domain, a longstanding yet effective design concept that persists today.28,29 The targeting domains increase the concentration of antimicrobial peptides on the surface of target bacteria, thereby enhancing the killing efficacy and dynamic properties of the antimicrobial peptides. Several studies have demonstrated that STAMPs can be engineered to selectively target and eliminate Streptococcus mutans without affecting other non-cariogenic oral streptococci (Figure 2). 30–33 Furthermore, the incorporation of a targeting domain enables the antimicrobial peptide to specifically act on cariogenic bacteria while sparing host oral cells and tissues, thereby enhancing its potential safety profile during application.
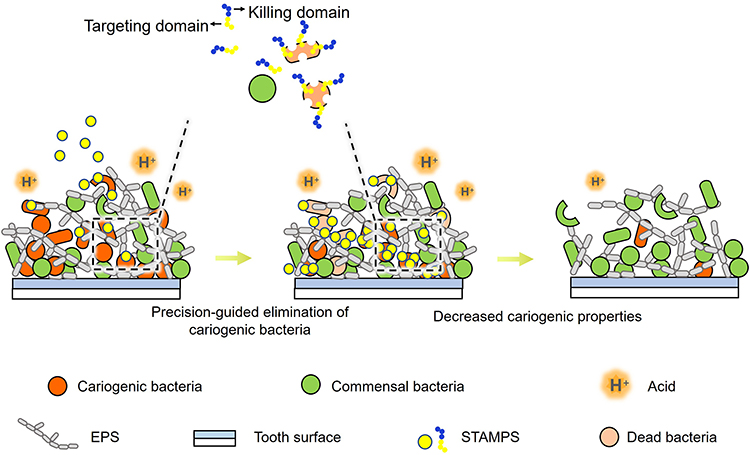 |
Figure 2 As illustrated, STAMPS consist of two functional components, a targeting domain and a killing domain. The targeting domain guides STAMPS to specifically recognize and bind to cariogenic bacteria (eg., S. mutans), enabling the killing domain to destroy them. This precision-targeted approach specifically eliminates cariogenic bacteria while leaving commensal bacteria unharmed. Ultimately, this decreases the dominance of cariogenic pathogens and significantly inhibits the cariogenic properties of the dental plaque biofilm, such as EPS accumulation and acid production.30–33 |
C16G2 and C16G2-Like STAMPs
An early and notable instance of the STAMP is C16G2, which consists of three interconnected yet distinct domains.33 The targeting domain of the STAMP, CSP-C16, is a truncated variant of the competence stimulating peptide (CSP) pheromone from S. mutans. This domain is fused with G2, a fragment derived from the broad-spectrum antimicrobial peptide novispirin G10. The two regions are linked by a flexible tri-glycine linker. Within a multispecies community, C16G2 selectively inhibits S. mutans and suppresses some harmful bacteria interacting with S. mutans, while concurrently allowing for the relative enrichment of beneficial symbiotic bacteria such as S. mitis and S. cristatus.31,33 This selective modulation facilitates the development of a more benign oral microbial community, indicating that STAMPs possess the capability to regulate microbial ecology.15,31 Furthermore, C16G2 exhibits rapid action, affecting bacteria within a minute, and is water-soluble, rendering it suitable for delivery via mouth rinse.34,35 R. Sullivan et al reported that C16G2 rinse usage was associated with reductions in plaque and salivary S. mutans, lactic acid production, and enamel demineralization.35 C16G2 has completed a Phase II clinical trial (ClinicalTrials.gov Identifier: NCT03196219), which evaluated various varnish and strip formulations in both male and female dental subjects using a single-blind, open-label design.36,37 The primary outcomes assessed included safety and tolerability, as well as reductions in S. mutans levels. However, the open-label nature of the trial and the absence of detailed microbiome-profiling outcomes limit the assessment of its true ecological impact. Additionally, robust sample size justification and long-term efficacy data were not reported in the available trial summaries, representing further limitations of the current clinical evidence. Consequently, antimicrobial peptides hold significant potential for practical applications. However, further clinical translation has been hindered by the high costs of chemical synthesis, which pose limitations for the use of antimicrobial peptides in caries prevention. Some studies have suggested biosynthesis strategies for antimicrobial peptide production to enhance efficiency and reduce costs.38,39 Montesinos et al utilized rice oleosin 18 as a carrier protein and applied oleosin fusion technology to efficiently synthesize cecropin A in rice seeds, indicating that the use of plants as biofactories for AMPs may represent a cost-effective and safe alternative.40 In a related study, Amir Pandi et al developed a cell-free protein synthesis platform for the rapid and economical production of antimicrobial peptides directly from DNA templates.41
Building upon the design strategy of STAMPs C16G2, subsequent research has integrated antimicrobial peptide domains with targeting domains such as C16 to develop targeted anticaries peptides. For instance, Che et al engineered a human-specific targeted antimicrobial peptide, C16-LL-37, aimed at S. mutans by combining the human antimicrobial peptide domain LL-37 with the targeting domain C16.42 Similarly, Huo et al designed and chemically synthesized a series of STAMPs exhibiting selective antibacterial activity against S. mutans in both planktonic and biofilm states, while demonstrating minimal effects on oral Streptococci and multi-species biofilms. They also investigated the potential of various fragments of CSP to function as the STAMP-targeting domain (C8: TFFRLFNR; C11: TFFRLFNRSFT; C12: LSTFFRLFN; C13: STFFRLFNRSFTQ; C14: LSTFFRLFNRSFTQ).32 This approach has facilitated the exploration of additional STAMPs. Employing this methodology, a greater number of STAMPs were investigated. He et al employed synthetic targeting and antimicrobial peptide libraries as foundational elements to create several novel STAMPs exhibiting selective activity against S. mutans. The STAMPs developed through these methodologies were subsequently optimized by modifying the linker and the incorporated antimicrobial peptides. The refined STAMPs exhibited efficacy against S. mutans and biofilms, thereby illustrating an enumeration method for screening STAMPs.43 This enumeration method also serves as an inspiration for the development and screening of antimicrobial peptides with enhanced efficacy through sequence assembly (Figure 3).
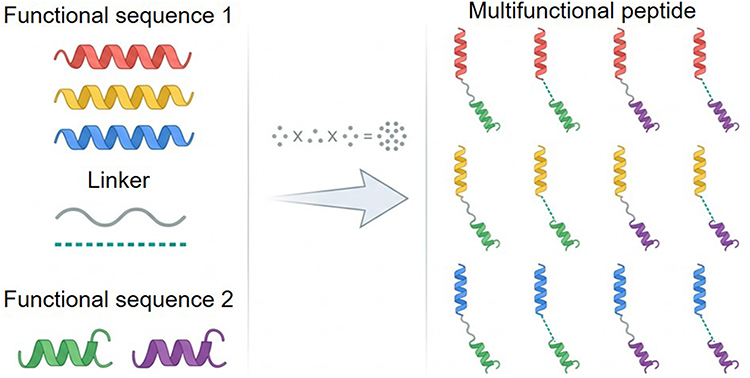 |
Figure 3 Schematic of the combinatorial enumeration method for generating multifunctional peptides. Inspired by the design of STAMPS, this schematic illustrates a modular approach. As illustrated, different “Functional sequence 1” modules (such as antimicrobial domains) are linked to “Functional sequence 2” modules (such as targeting domains) using distinct “Linkers”. Through combinatorial assembly, these components generate a comprehensive library of theoretically multifunctional peptides for subsequent screening. |
In summary, C16G2 could serve as a reference model for similar studies, encompassing its design strategy and anticaries functional validation.
Screening of Phage Peptide Libraries for Antimicrobial Peptides
Phage display is a robust and high-throughput screening methodology wherein exogenous genes are incorporated into the DNA of bacteriophages, facilitating the expression of a library of peptide or protein variants on the exterior of the phage virion.44 This technique has enabled researchers to identify peptide sequences with specific binding affinities for Candida albicans,45 Staphylococcus aureus,46 and Acinetobacter baumannii.47 While some investigations have focused on human antibody fragments capable of recognizing Streptococcus mutans with high specificity through human antibody variable gene phage-display libraries, there is a paucity of research dedicated to designing specific antimicrobial peptides targeting Streptococcus mutans using this technology.48,49 Lee et al employed a target non-immobilization approach for phage selection, successfully screening three effective sequences: SmP (SLSTATPLSLIR), SoP (SSSLSYPTTSHL) and LcP2 (GHNRAQPMTTPF), for S. mutans, Streptococcus oralis and lactobacillus casei respectively.50 SmP showed high bind affinity for both S. mutans and S. oralis and decreased the biofilm thickness of the two species, but SmP only inhibited biofilm formation of S. mutans, suggesting the selectivity to some extent. Therefore, this approach could represent a new direction for research of targeted antimicrobial peptides against cariogenic bacteria.
Other STAMPs
Several studies have identified that certain anticaries peptides, which were not initially developed for targeted action, demonstrate selective inhibitory effects. Nonetheless, the precise mechanisms underlying their targeting capabilities remain unclear.
Proteomic analyses have shown significantly elevated levels of lactotransferrin in the acquired enamel pellicle of individuals susceptible to dental caries, indicating its crucial role as a caries-associated protein.51 Based on this research, Luo et al designed and synthesised novel lactotransferrin-derived antimicrobial peptides, designated as LF-1.52 In follow-up studies and subsequent analyses, LF-1 displayed selective antibacterial activity against S. mutans rather than common oral streptococci strains.53 While the design of LF-1 deviates from traditional targeting strategies, it exhibits a strong interaction with the membrane of S. mutans, causing significant membrane disruption compared to the commensal bacteria S. sanguinis and S. gordonii. This observation suggests that LF-1 may engage with specific binding sites on the S. mutans membrane.53 This study advocates for the use of proteomic techniques to analyze protein profiles in acquired enamel pellicle or saliva, aiming to identify proteins that are differentially expressed between individuals susceptible to dental caries and those who are caries-free. These differentially expressed proteins could serve as templates for designing biomimetic anticaries peptides, offering a promising strategy for dental health. Numerous salivary proteins, such as lysozyme, play a crucial role in defending against cariogenic bacteria and maintaining oral microbiota balance.2 However, due to the limited exploration of underlying mechanisms and the restricted inclusion of diverse bacterial strains, further assays are necessary to validate the selectivity.
Peptides Targeting Cariogenic Virulence Factors
Targeting cariogenic virulence factors is another promising precision-guided strategy for caries prevention. Precisely inhibiting cariogenic properties without excessively impacting the growth of oral bacteria may minimize harm to the oral microbiota. Furthermore, the reduction in virulence could diminish the ecological advantages of cariogenic bacteria, thereby aiding in the reestablishment of homeostasis. For instance, common targets include antigen I/II, which mediates bacterial adhesion to the tooth surface and co-aggregation; glucosyltransferases, which play essential roles in EPS production and biofilm establishment; and sortase A, a membrane-associated enzyme responsible for anchoring surface virulence proteins.54,55 Multiple small molecules have been synthesized using computer-aided drug design to perturb the adhesion and biofilm accumulation of S. mutans via the aforementioned targets.54,55 Beyond biofilm formation, efflux pumps and proton pumps represent other promising virulence targets that confer bacterial pathogenicity and colonization. For instance, reserpine—an inhibitor of multidrug resistance pumps—significantly impairs the efflux capacity of S. mutans without affecting its growth, thereby weakening its acid tolerance, mutacin production, and transformation efficiency.56 Similarly, the F-type ATPase is a crucial proton pump responsible for bacterial survival in the acidified cariogenic microenvironment. Small molecules such as bedaquiline, piceatannol, curcumin, and demethoxycurcumin have been shown to act as F-type ATPase inhibitors, effectively reducing enzymatic activity and bacterial growth under acidic conditions.57 Although the inhibitors discussed above are small molecules rather than peptides, they are explicitly included here as conceptual analogues. They successfully illustrate the feasibility of targeting specific virulence factors and highlight the utility of computational drug discovery—approaches that remain significantly underexplored in peptide design. Ultimately, while these proof-of-concept studies demonstrate the viability of virulence-targeted interventions, such strategies have yet to be widely adopted for anticaries peptides. Therefore, these small-molecule paradigms provide a highly valuable framework to inspire and guide future peptide-based therapeutics.
In most studies of anticaries peptides, their anti-virulence properties are derived from their antibacterial effects, serving primarily as a complement to antibacterial mechanisms. This limitation can be attributed not only to the complex structures of antimicrobial peptides, which pose challenges for binding to specific targets, but also to several additional physicochemical and delivery-related constraints. First, the proteolytic instability of peptides in the oral environment leads to rapid degradation before they can reach and interact with intracellular or membrane-associated virulence targets. Second, poor tissue penetration limits the ability of peptides to access deeper layers of dental plaque biofilms or infiltrate the complex extracellular polymeric matrix. Third, many key virulence targets, such as sortase A or components of two-component signal transduction systems, are located intracellularly or on the inner surface of the membrane, posing challenges for intracellular accessibility that most cationic linear peptides cannot readily overcome. These limitations highlight the need for innovative delivery strategies and structural modifications, such as conjugation with cell-penetrating peptides, encapsulation in nanoparticles, or incorporation of biofilm-penetrating domains, to enhance the translational potential of virulence-targeting peptides. For instance, Duprez et al found that a nine-residue ligand (PSPFATCDF) from the Escherichia coli oxidative folding machinery protein DsbB could bind the partner enzyme DsbA.58 The subsequent structure-activity relationship study provided additional modified peptides and confirmed their selective binding effects, thereby offering a novel approach for the design of anti-virulence peptides in caries prevention.
Do the so-Called Targeting Peptides Really Selectively Inhibit Cariogenic Bacteria?
Although numerous antimicrobial peptides reported in recent years have been proposed to selectively target cariogenic bacteria, many of them did not fully achieve such precision in practice. In this section, a table (Table 1) of antimicrobial peptides with claimed targeting and selective activity on S. mutans in the studies of 2020–2025 is summarized using a simple retrieval on PUBMED, ((target) OR (selective)) AND (peptide) AND (Streptococcus mutans), among which 64 articles were searched and 10 original research articles (6 peptides) were selected because only the peptides, not combination or biomaterials, were used as antimicrobial strategies for caries prevention in these studies.25,50,52,53,59–64 All six peptides were thought to possess selective inhibitory effects on cariogenic bacteria. Specifically, SmP and 8DSS-C8-P-113 were tested against only three bacterial species, whereas GHb and its derivatives were evaluated with a single oral commensal strain. In contrast, only LF-1, LH12, and GH12 were subjected to a more comprehensive analysis involving a broader array of cariogenic and commensal bacteria. Although SmP, GHb and its derivatives, and GH12 demonstrated potent antibacterial effects against S. mutans and other cariogenic species, they also showed similar inhibitory activity against beneficial commensals such as S. oralis, S. sanguinis, and S. salivarius. The restricted scope of testing and the unintended suppression of commensal bacteria undermine the evidence supporting their purported specificity. Among these peptides, only LF-1 and GH12 were evaluated in vivo. However, LF-1 was not subjected to in vivo microbial profiling, rendering its impact on oral microbiota composition and S. mutans colonization ambiguous. While GH12 effectively decelerated caries progression, sequencing data did not reveal a significant reduction in oral S. mutans levels. While LH12 has yet to undergo in vivo testing, its efficacy in multi-species biofilm models has demonstrated enhanced selectivity, effectively inhibiting cariogenic pathogens while promoting commensal populations within an S. mutans–S. gordonii model. Among the six peptides analyzed, only LF-1 and LH12 exhibited notably selective inhibitory effects across various caries-associated species, although their underlying mechanisms remain largely unexplored. These findings underscore the necessity for future research to encompass a broader spectrum of bacterial species, employ more complex multi-species models, conduct rigorous in vivo studies, and pursue mechanistic investigations to accurately assess the selective antibacterial efficacy of targeted antimicrobial peptides.
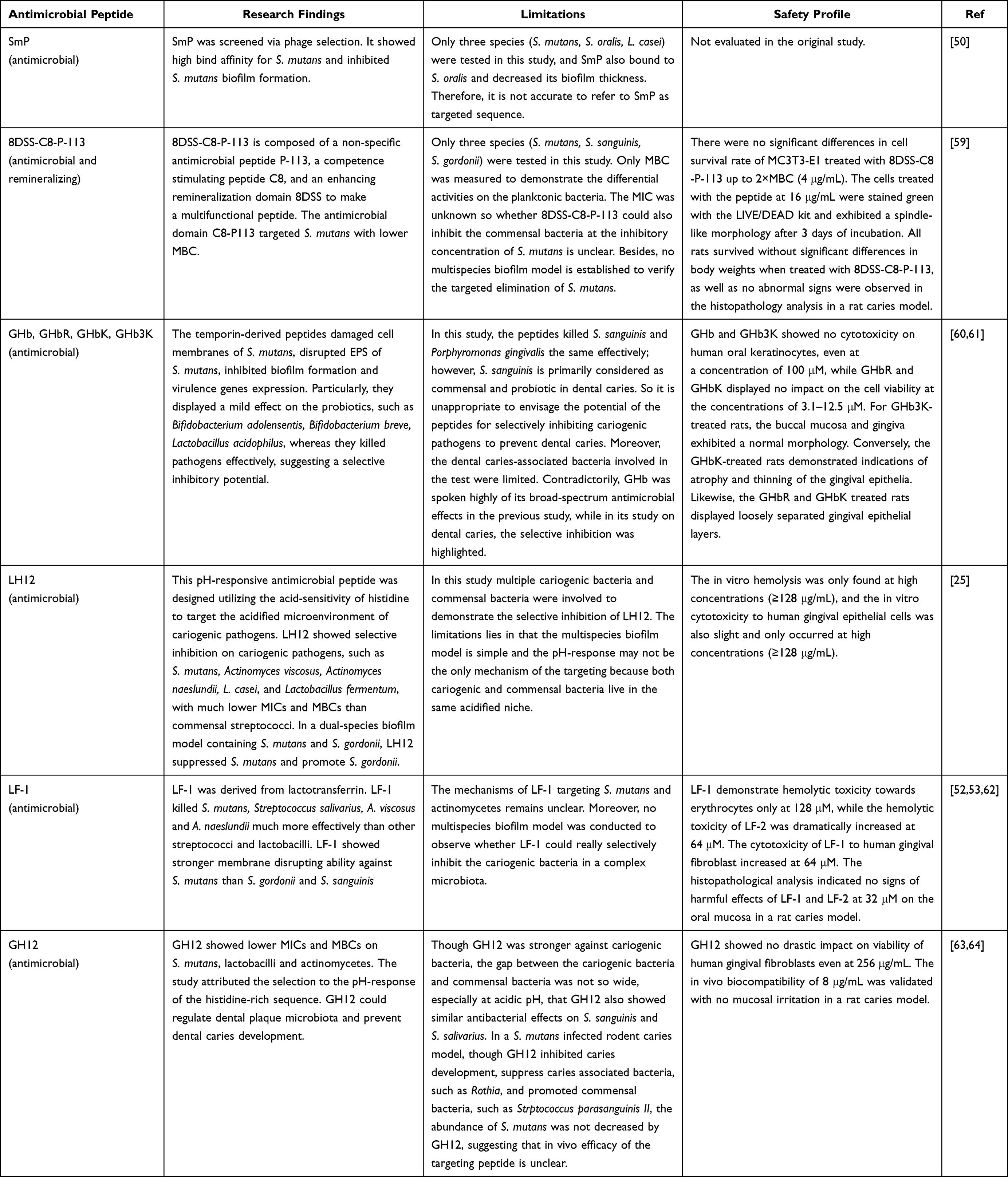 |
Table 1 The so-Called Targeting Peptides for Dental Caries from 2020 to 2025 |
Bioresponsive Antimicrobial Peptides: Establishing a Balanced Oral Microbial Community in the Pathogenic Acidified Microenvironment
Targeted antimicrobial peptides undoubtedly offer a promising ecological strategy for caries prevention. However, microbial analysis of saliva samples and plaque samples has revealed that the abundance of S. mutans is not directly correlated with the occurrence and development of dental caries.65,66 Some studies have reported that S. mutans is not consistently detected in all cases of early childhood caries and may also be present in caries-free children, indicating a complex relationship between specific bacterial species and caries development.66,67 It is found that S. mutans is not the primary cariogenic bacterium in all patients, and there are other non-Streptococcus mutans cariogenic bacteria associated with high caries, such as S. sanguinis, S. mitis, S. sinensis, S. rubneri, S. vestibularis, S. cristatus and S. massiliensis.68 These findings suggest that an exclusive focus on S. mutans for caries prevention may not yield optimal outcomes and could potentially disrupt microbial balance, allowing other cariogenic bacteria with similar biological traits to occupy larger ecological niches. Cariogenic bacteria may exist as part of normal oral flora in healthy environments and contribute to caries only in highly cariogenic oral environments. Therefore, antimicrobial caries prevention strategies should encompass not only the inhibition of cariogenic bacteria but also the management of high-risk microenvironments such as the acidification-induced microecological imbalances caused by various acid-producing and acid-tolerant bacteria.69 Therefore, bioresponsive strategies are proposed, which targets the dysbiotic microenvironment associated with dental caries and helps recover microbial homeostasis, is more consistent with ecological principles (Figure 4).
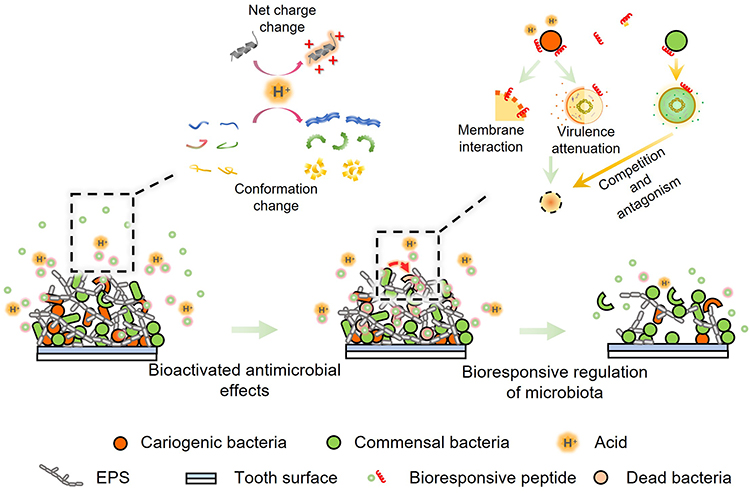 |
Figure 4 Bioresponsive antimicrobial peptides for dental caries. The antimicrobial effects of the peptides can be activated by the pathogenic conditions via net charge change and conformation change. The bioresponsive antimicrobial peptides may be more sensitive to cariogenic bacteria because of the cariogenic microenvironment created by the pathogens. Apart from the direct bactericidal effects and virulence inhibiting effects on cariogenic bacteria, the peptides may also stimulate the production of antimicrobial substances from the commensal bacteria and promote their ecological advantages, and subsequently regulate the composition of the microbiota. |
pH-Responsive Antimicrobial Peptides
Given that acidification of the microenvironment results in dysbiosis of dental plaque and enamel demineralization, the development of pH-responsive antimicrobial peptides, which exploit the pH differential between normal oral buffering conditions and acidic biofilms, represents a promising ecological strategy for caries prevention.
Wang et al designed and synthesized a cationic amphipathic α-helical AMP GH12 (GLLWHLLHHLLH-NH2) which showed rapid and strong antimicrobial activity against oral streptococci in vitro and inhibited various virulence factors of S. mutans effectively.70–72 In subsequent investigations, Jiang et al demonstrated that GH12 to some degree selectively inhibits S. mutans within a controlled multispecies biofilm model containing both pathogenic bacteria (S. mutans) and commensal bacteria (S. gordonii and S. sanguinis).73 Moreover, the potential of this antimicrobial peptide for ecological caries prevention has been substantiated. It exhibited minimal impact on the microbiota of human dental plaque-derived multispecies biofilms from healthy volunteers but could regulate the microbiota of dental plaque in the rat caries model in vivo, reducing the abundance of some cariogenic bacteria.64 Subsequent mechanistic studies have suggested that this modulation effect is attributable to the histidine-rich sequence present in GH12, which enhances the cationic and amphipathic properties of the peptide.63 Under acidic pH conditions, the histidine residue in GH12 becomes protonated, thereby increasing the net positive charge of the molecule.74 This enhanced positive charge promotes the electrostatic attraction of GH12 to anionic microbial membranes. In addition to the pH-responsive charge change, conformation change in the secondary structure of peptides, such as transitioning from a random coil at neutral pH to an α-helix at acidic pH, is also utilized to design the pH-responsive AMPs. For example F5 (FLEHLIPHVIHGLVHAIHH-NH2) was designed with an increased helical structure at acidic pH to enhance selective killing activity for bacterial abscess therapy.75
While GH12 exhibits ecological caries-preventive properties, it nonetheless impacts some symbiotic bacterial populations. Jiang et al further utilized the pH-sensitivity of histidine to design a pH-responsive antimicrobial peptide, LH12 (GLLHLLHHLLHH-NH2).25 LH12 exhibited significantly lower minimum inhibitory concentrations against the tested cariogenic bacteria compared to commensal bacteria. LH12 underwent protonation within the acidified cariogenic microenvironment, resulting in increased cationicity and stronger interactions with bacterial membranes. This acid-activated mechanism enabled LH12 to selectively target cariogenic pathogens and acidified niches without disrupting commensal bacteria. Furthermore, the acid-activated mechanism endows the peptide with an enhanced biosafety profile. Because healthy oral tissues are maintained in a buffered, neutral physiological environment, the peptide’s positive charge and membrane-interacting capabilities are substantially reduced, thereby minimizing unintended effects on host tissues.
LH12 demonstrated significant inhibitory effects on various virulence factors of S. mutans, including biofilm formation, acid tolerance, acid production, and polysaccharide synthesis. The expression of virulence-associated genes, such as gtfBCD, atpD, and ldh, was notably downregulated. Simultaneously, the transcriptional levels of genes involved in two-component signal transduction systems related to stress tolerance, cell viability, and the regulation of cariogenic virulence, including vicR, liaR, and comDE, were also diminished. Beyond its suppressive effects on S. mutans, LH12 enhanced the production of hydrogen peroxide and the generation of reactive oxygen species by S. gordonii, thereby increasing its competitive advantage. These findings underscore the dual ecological regulatory role of LH12 in modulating the composition of biofilm-associated bacteria. LH12 effectively inhibits S. mutans activity while promoting the ecological benefits of S. gordonii, contributing to a healthier oral microbiome and reducing the risk of dental caries. Although in vivo model results are currently insufficient, preliminary findings indicate that the ecological modulation mechanism of antimicrobial peptides not only targets cariogenic bacteria but also promotes the growth of symbiotic bacteria. This dual-functional mechanism of LH12 offers a novel bioresponsive ecological approach for caries prevention.
Dual-Sensitive Antimicrobial Peptides
Zhang et al designed pHly-1 (KIWFHAMHIIAHFIAHQHLHQHL) by primarily replacing lysine with histidine residues.76 The AMP exhibited pH-sensitive activity under acidic conditions and demonstrated lipid-binding capabilities, suggesting its potential application in the treatment of dental caries. Under acidic conditions, it adopted random coil conformation and formed nanoparticles with an average diameter of approximately 40 nm. In contrast, at neutral pH, the peptide underwent a conformational shift to β-sheet structures, leading to the assembly of nanofibers. Within the acidic microenvironment of cariogenic biofilms, interaction with the bacterial membrane induced a coil-helix transition in the peptide pHly-1, facilitating its penetration into the lipid bilayer and subsequent disruption of the S. mutans biofilm. Conversely, at neutral pH, pHly-1 adopted a β-sheet configuration, forming nanofibers that remain in an aggregated state due to stabilization by hydrogen bonding and hydrophobic interactions. This aggregation impeded the penetration of pHly-1 into bacterial membranes, thereby exhibiting minimal toxicity to oral microbiota and host tissues. Subsequent in vitro and in vivo experiments have corroborated that this antimicrobial peptide demonstrates significant efficacy against cariogenic bacteria and biofilms, without adversely impacting oral microbial communities or host tissues. This design strategy thereby augments both the safety and ecological effectiveness of the antimicrobial peptide in combating dental caries.
In conclusion, bioresponsive strategies constitute a promising ecological approach for the application of antimicrobial peptides in the prevention and treatment of dental caries. Nevertheless, these studies are not without their limitations. Numerous researchers have expressed optimism regarding pH-responsive strategies, expecting these approaches to exert ecological regulatory effects that inhibit cariogenic bacteria and restore the balance between acid-producing and acid-tolerant bacterial populations. Despite this optimism, limitations in model construction, experimental design, and sequencing accuracy hinder the ability of these studies to conclusively demonstrate whether pH-responsive antimicrobial peptides can effectively regulate the dysbiosis in the microecology associated with dental caries as anticipated. This highlights the necessity for the advancement of both in vitro and in vivo models, as well as the exploration of microbiome analysis, in future research on pH-responsive antimicrobial peptides. Such improvements are essential to more effectively demonstrate the theoretical benefits of pH-responsive strategies.
Antibiofilm Peptides: Preventing the Accumulation of Cariogenic Initiator
Dental plaque biofilm has been considered as the initiator of dental caries. Biofilms serve as a physical barrier, safeguarding embedded bacteria from antimicrobial agents and the host defense system, while also providing a conducive environment for bacterial interaction and the accumulation of cariogenic virulence factors.77 Consequently, the antibiofilm properties of various agents are of significant interest for their potential application in plaque control and caries prevention. Many antimicrobial peptides exhibit antibiofilm properties to varying degrees, as they can inhibit bacterial growth or directly kill bacteria, similar to other antimicrobial peptides discussed in this manuscript. Notably, certain peptides demonstrate antibiofilm efficacy without possessing bactericidal activity, which is believed to result in minimal adverse effects on the oral microbiota. This aspect will be further explored in this section.
Chen et al utilized computational methods to simulate short amyloid-forming peptides (ASNIVI, TSFVLV, GIDLKI) derived from the amyloid-forming truncated protein C123 of Streptococcus mutans. These amyloid-forming hexapeptides (AFhPs) displayed broad-spectrum antibiofilm activity against S. mutans as well as other Gram-positive bacteria, Gram-negative bacteria, and fungus, by aggregating into rigid amyloid fibrils that entangle microbe cells without killing bacteria.78 In addition, one of the AFhPs, GIDLKI, demonstrated efficacy in inhibiting multi-species biofilm formation and mitigating the progression of caries in both human tooth blocks and rat teeth.79 Kokilakanit et al found a series of pomegranate-derived peptides (LLKLFFPFLETGE, GAVGSVV, LGTY, FPSFLVGR) could inhibit biofilm formation of S. mutans without exhibiting strong bactericidal effects.80 The sequence LLKLFFPFLETGE was found to reduce glucosyltransferase activity, thereby impeding bacterial adhesion. In addition, taking the peptide sequence for other gram-positive pathogenic biofilm is useful to extend the scope of cariogenic biofilm control. A cationic synthetic peptide, called P1 (PARKARAATAATAATAATAATAAT), derived from the Ixodes tick antifreeze glycoprotein with potent antibiofilm activity against S. aureus, also strongly inhibited biofilm formation of S. mutans. This inhibitory action is likely attributable to the interaction between P1 and S. mutans, which disrupts surface attachment and microcolony aggregation.81 The anti-adhesion and anti-aggregation characteristics of these antibiofilm peptides offer promising avenues for the inhibition of biofilm formation and dispersion, potentially facilitating the removal of persistent dental plaque (Figure 5).
 |
Figure 5 Antibiofilm peptides for dental caries. The antibiofilms peptides entangle bacteria to prevent the microbial aggregation, inhibit the activity of glucosyltransferase to reduce matrix production and bacteria adhesion, or serve as an obstacle to tooth surface attachment and bacteria co-aggregation, which destroy the formation and accumulation of the dental plaque microenvironment without killing bacteria. |
Bifunctional Anticaries Peptides: Providing Not Only Bacteriostasis but a Bonus of Tooth Surface Protection
Microenvironmental acidification and microbial dysbiosis lead to a localized reduction in pH, thereby disrupting the balance between mineral loss caused by oral microbial biofilms and physiological mineral uptake from the oral environment.82 This disruption is a fundamental cause of dental caries. It can be concluded that the initiation of bacterial activity and the subsequent demineralization of enamel ultimately result in the formation of dental caries. Traditional antimicrobial peptides primarily inhibit bacterial proliferation to prevent caries but lack the capability to repair existing enamel damage. However, due to the variability in amino acid sequences of antimicrobial peptides, these molecules can exhibit a range of additional activities.83 Several studies have focused on the design of bifunctional antimicrobial peptides that target both antibacterial effects and the protection of tooth surfaces to prevent caries (Figure 6).
 |
Figure 6 Bifunctional peptides for dental caries. Certain peptides can bind tooth surface and act as protective layer to inhibit bacterial colonization and biofilm formation. Other bifunctional peptides can kill cariogenic bacteria to eliminate the initiator of dental caries and promote the formation of hydroxyapatite and remineralize the damaged tooth surface. |
Dual-Action Peptide with Antimicrobial and Remineralizing Properties
It has been demonstrated that the design of peptides, through the assembly of two or more peptides, can yield distinct functionalities derived from their individual components and facilitate their formulation into pharmaceutical products.84,85 The initial design strategy involved the fusion of an antibacterial peptide with a short peptide possessing established remineralizing capabilities. Wang et al developed and synthesized a bifunctional anticaries peptide, TVH19 (TKRQQVVGLLWHLLHHLLH-NH2), which exhibits both antibacterial and remineralizing properties.86 Its N-terminal mineralizing sequence, derived from the hydrophilic C-terminal tail of amelogenin TD7 (TKREEVD), is TKRQQVV, while its C-terminal antimicrobial sequence is antimicrobial peptide GH12 (GLLWHLLHHLLH-NH2). Research has shown that TVH19 exhibited the antimicrobial activities and could inhibit biofilm formation. Furthermore, researchers assessed the remineralizing activity of TVH19 in vitro, demonstrating its efficacy. It could promote remineralization of demineralized enamel, reduce mineral loss on the tooth surface and restore microhardness of demineralized enamel. Furthermore, Li et al designed bifunctional peptides, 8DSS-C8-P113, by fusing the non-specific antimicrobial peptide P-113 (AKRHHGYKRKFH) with a competence-stimulating peptide C8 (TFFRLFNR) to create a targeted antimicrobial peptide against S. mutans. C8-P113 was fused with enhancing remineralization domain 8DSS (DSSDSSDSSDSSDSSDSSDSSDSS) for addition remineralising activity. 8DSS-C8-P113 showed both anti-S. mutans effects and remineralising effects on etched enamel.59 An alternative design strategy for dual-action peptides involves the modification of antibacterial peptide sequences with mineralizing groups. In a study by Niu et al, the N-terminus of the LL-37-derived antimicrobial peptide KR12 (KRIVQRIKDFLR) was modified with gallic acid (GA), resulting in the novel peptide GA-KR12. This peptide comprises KR12 as the antimicrobial domain and gallic acid as the mineralizing domain.87 Studies have shown that the novel antimicrobial peptide GA-KR12 inhibited the growth of S. mutans biofilm and enhanced the remineralization of artificial dentine caries.88,89
Although TVH19 demonstrated both antibacterial and remineralizing properties, its antimicrobial efficacy was inferior to that of the template antimicrobial peptide GH12. The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of TVH19 against S. mutans were 64 μM (149.0 μg/mL) and 128 μM (249.9 μg/mL), respectively, which were much higher than the MIC and MBC of GH12 against S. mutans (8.0 μg/mL, around 6 μM).70,86 The fusion drastically lowered the antimicrobial activity of 8DSS-C8-P113. The MBC of C8-P113 against S. mutans was 30 μM (79.0 μg/mL), compared to 2000 μM (9922.0 μg/mL) for 8DSS-C8-P113. Notably, in remineralization assays, 8DSS-C8-P113 required a concentration of 16000 μM to exhibit significant effects, whereas the control group C8-P113 was effective at only 250 μM. When both peptides were administered at their respective MBC, they displayed similarly limited remineralizing capabilities. Consistent with this pattern, GA-KR12 exhibited a MIC of 160 μM and an MBC of 320 μM against S. mutans, values significantly higher than those of the parent peptide KR-12.87 Furthermore, GA-KR12 required concentrations up to 640 μM to function effectively in an in vitro biofilm demineralization model.88 These findings suggest that the fusion of functional domains may compromise the intrinsic activity of each component, resulting in functional interference. Consequently, achieving the intended bifunctionality necessitates elevated dosages, which invariably increases application costs and hinders clinical translation. This reduction in antimicrobial efficacy likely stems from a biophysical conflict between the domains: antimicrobial peptides rely on specific cationic and hydrophobic properties to penetrate biological membranes,90 which may be disrupted by the introduction of mineralizing sequences. Conversely, the process of mineralization typically demands hydrophilic and negatively charged domains to facilitate calcium ion binding and subsequent hydroxyapatite nucleation. This creates a structural paradox, as these requirements often oppose the physicochemical traits necessary for antimicrobial action. Therefore, a critical objective for future research is to delineate the optimal equilibrium between antimicrobial potency and mineralizing capacity. Achieving this balance is essential to engineering dual-action peptides that maintain high efficacy at reduced concentrations, thereby improving their therapeutic viability. It is also worth noting that remineralizing domains, owing to their interactions with calcium ions or hydroxyapatite, may inherently possess an affinity for the tooth surface, thereby contributing to peptide retention. For instance, as will be discussed in Dual-Action Peptide with Antibacterial Efficacy and Improved Retention on Tooth Surface, the phosphoserine (Sp) domain of Sp-H5 not only facilitates enamel adsorption but also actively drives the remineralization process.
Dual-Action Peptide with Antibacterial Efficacy and Improved Retention on Tooth Surface
Furthermore, the clinical utility of AMPs in the oral environment is frequently hindered by rapid salivary clearance and susceptibility to proteolytic degradation. To circumvent these stability issues, an alternative strategic approach focuses on enhancing the retention of AMPs directly onto tooth surfaces. By establishing a contact-active antimicrobial barrier on the enamel, this method effectively inhibits initial bacterial adhesion and subsequent biofilm maturation, thereby providing a potent mechanism for in situ caries prevention. Crucially, this site-specific anchoring ensures that antimicrobial activity is targeted strictly toward cariogenic plaque, minimizing disruption to the broader commensal oral microbiome. Additionally, this strong adsorption to the tooth surface prevents the peptide from accumulating on surrounding soft tissues, further mitigating any unintended impact on the oral mucosa.
To implement this strategy, Huang et al engineered a hydroxyapatite-binding antimicrobial peptide (HBAMP, NNHYLPR-GGG-KKVVFWVKFK) by fusing a broad-spectrum antimicrobial peptide (KSLW, KKVVFWVKFK) with a specific hydroxyapatite-binding heptapeptide (NNHYLPR) identified via phage display bio-panning.91 While the fusion resulted in a modest reduction in antimicrobial potency—HBAMP exhibited a MIC of 125 μg/mL and an MBC of 250 μg/mL against S. mutans, compared to 62.5 μg/mL and 125 μg/mL for the parent KSLW—its surface-functionalization properties were superior. In vitro assays demonstrated that HBAMP rapidly binds to hydroxyapatite, creating a contact-active antibacterial surface capable of eliminating adhering bacteria and effectively inhibiting biofilm formation on tooth substrates.92
Beyond simple surface retention, recent efforts have aimed to engineer molecules that possess simultaneous antibiofouling and mineralizing capabilities. In this vein, Zhou et al synthesized a bioactive conjugate, phosphoserine-histatin 5 (Sp-H5, phosphoserine-DSHAKRHHGYKRKFHEKHHSHRGY), by attaching a phosphoserine (Sp) moiety to the N-terminus of Histatin 5 (H5).93 As a naturally occurring salivary peptide, native H5 is recognized for its broad-spectrum antimicrobial activity against oral pathogens, including S. mutans, as well as its inherent ability to adsorb onto dental surfaces. The strategic addition of Sp was designed to augment this functionality by recruiting calcium ions to the tooth surface. Regarding its antimicrobial profile, Sp-H5 exhibited an MIC of 2 mM and an MBC of 4 mM against planktonic S. mutans. Although these concentrations are significantly lower than those required to eradicate established biofilms, they indicate a moderate efficacy against free-floating bacteria. Importantly, in vitro assays confirmed that Sp-H5 exhibits superior hydroxyapatite binding affinity compared to the native peptide, alongside an enhanced capacity to inhibit demineralization and promote remineralization. Consequently, conjugating the Sp domain to H5 not only anchors the peptide to the tooth surface to resist biofilm accumulation and exert sustained in situ antimicrobial activity, but also endows it with the capacity to repair damaged enamel through active remineralization. This highlights a highly promising two-pronged strategy that simultaneously tackles the two most critical targets in caries management: bacterial pathogenesis and structural mineral loss. Mechanistically, this binding is driven by strong electrostatic attractions between the cationic amino acid residues (Lys12, Arg13, Lys14, Lys18, and Arg23) of the peptide and the anionic phosphate groups (PO43-) of the enamel lattice. Concurrently, the negatively charged phosphoserine moiety acts as a nucleation site by sequestering calcium ions, thereby facilitating hydroxyapatite crystal growth.
Building upon this conceptual framework, further optimization studies have focused on refining the functional domains of H5.94 Researchers selected P-113 (AKRHHGYKRKFH), the minimal active fragment of H5, as the core antimicrobial motif. To engineer dual-bioactive conjugates, specific functional moieties were appended to the C-terminus of P-113, yielding three distinct variants: P-113-DPS (incorporating a bisphosphoserine motif, SpSp), P-113-DA (conjugated with D-3,4-dihydroxyphenethylamine, or dopamine), and P-113-SK (fused with the hydroxyapatite-binding peptide SKHKGGKHKGGKHKG). Comparative analysis revealed that P-113-DPS exhibited the most comprehensive therapeutic profile, demonstrating superior surface adsorption, antibiofouling efficacy, and inhibition of demineralization, alongside enhanced remineralization capacity, stability, and cytocompatibility. It is hypothesized that the interaction between the bisphosphoserine residues and divalent metal ions facilitates the accumulation of P-113-DPS on bacterial membranes, thereby increasing membrane permeability. Furthermore, in the P-113-DPS-hydroxyapatite complex, the exposed anionic DPS moieties serve as potent calcium attractors, significantly amplifying the peptide’s remineralizing potential. Collectively, this design strategy, which integrates antibiofouling and mineralizing functionalities into a single construct, offers a promising paradigm for caries prevention and in situ enamel regeneration.
Potential Novel Strategy for Designing Multifunctional Peptides
Consistent with the examples discussed above, bifunctional peptides engineered via conventional sequence assembly strategies frequently suffer from compromised biological activity. These assembly protocols generally fall into three categories: (1) conjugation, where two active fragments are joined via a linker; (2) fusion, involving the direct coupling of functional domains; and (3) merging (or hybridization), where shared structural motifs between two fragments are utilized for integration (Figure 7). In such constructs, functional domains with divergent physicochemical properties—such as charge, hydrophobicity, and conformation—often exhibit mutual interference. This structural incompatibility increases molecular complexity while attenuating specific biological functions, creating significant hurdles for multifunctional peptide design. Consequently, there is a pressing need to optimize design strategies beyond simple physical assembly. While computational methodologies have been extensively applied to develop antimicrobial agents for caries management, their application specifically for bifunctional anticaries peptides remains largely unexplored. A relevant precedent is provided by Hsueh et al, who employed a super-learning-based algorithm to engineer multifunctional peptides capable of efficient cell entry and melanin binding.95 They successfully screened a lead candidate, HR97, for effective cell penetration and sustained drug release. Although distinct from the oral environment, this study underscores the immense potential of bioinformatics approaches in accelerating the discovery of novel anticaries agents. A refined iteration of the assembly approach involves the hybridization of chemically compatible functional segments. For instance, Christian et al successfully integrated two independent domains to construct a multifunctional peptide possessing both cell-penetrating and antimicrobial activities.96 Similarly, Wang et al engineered a peptide targeting autoimmune and oxidative stress-related disorders by combining immunomodulatory and antioxidant sequences.97 While fundamentally remaining a form of sequence assembly, this strategy distinguishes itself by selecting functions and sequences that are intrinsically compatible, thereby circumventing the activity loss typically caused by structural conflicts. However, the success of this method relies heavily on the extensive screening of compatible candidates. In the context of caries prevention, this highlights the critical need to deeply explore the structural interplay between antibacterial and mineralizing fragments to identify combinations that permit harmonious functional integration.
 |
Figure 7 The design concept of a group of mutlifunctional peptides.98 A novel design strategy named sequence transformation beyond sequence assembly was proposed in this study. Amino acid mutations at specific sites were conducted within the first functional sequence, based on its inherent conformational propensities, to progressively align it with the secondary functional sequence and thus facilitate multiple functions. |
Beyond physical assembly, a more innovative strategy is “sequence transformation.” Xie et al demonstrated this by modifying osteogenic growth peptide (OGP) into a multifunctional agent with integrated anti-infective, immunomodulatory, and osteogenic activities (Figure 7). 98 Initially, they employed a conventional fusion strategy to create GK16 by combining sequences from OGP and lactoferrin. However, they subsequently developed two superior peptides, WL14 and GL14, using a novel transformation approach. Recognizing that OGP inherently possesses a hydrophobic-hydrophilic alternation pattern similar to that of antimicrobial peptides, they preserved its basic skeletal framework while substituting specific amino acids to optimize amphiphilicity and net charge. Through these modifications, the osteogenic peptide underwent a structural transformation, acquiring additional immunomodulatory and antibacterial capabilities. Essentially, this transformation entails introducing site-specific mutations within a primary functional sequence—guided by its conformational propensities—to align it with the structural requirements of a secondary function. This similarity-driven shift enables distinct functional motifs to operate concertedly within a single-sequence framework. This concept offers a compelling alternative for harmonizing antimicrobial and remineralizing fragments, paving the way for next-generation bifunctional anticaries peptides that simultaneously regulate the oral microecology and protect tooth surfaces.
Notably, beyond incorporating functional domains for enhanced tooth surface retention or remineralization, structurally modifying the peptide sequences themselves is a promising strategy to improve resistance against proteolytic degradation. The oral cavity contains a diverse array of proteases from saliva, host tissues, and bacteria that can rapidly cleave linear peptides and reduce their therapeutic efficacy.26,27,99,100 Although the complex oral environment poses a high risk of degrading AMPs and lowering their efficacy, most anticaries AMP studies do not routinely test for peptide stability. Instead, this focus is largely seen in pure peptide engineering research.101,102 This difference is likely due to disciplinary gaps, as dental research focuses more heavily on anticaries properties. This highlights a limitation in current oral research and suggests that stability testing must be included to broaden the characterization of anticaries AMPs.86 To overcome this challenge, various chemical and structural approaches have been utilized. These include cyclization to constrain peptide conformation and limit protease access; incorporating D-amino acids unrecognized by natural proteases; terminal modifications (N-terminal acetylation or C-terminal amidation) to prevent exopeptidase cleavage; and substituting labile residues with resistant analogs.26,103–105 These modifications can be seamlessly added without losing antimicrobial activity, thereby extending the peptide’s half-life and clinical viability. Future development of anticaries peptides should therefore consider not only the optimization of dual functionalities but also the engineering of proteolytic stability as an integral design criterion.
Conclusion
In summary, the development of AMPs for caries management has shifted from a purely bactericidal focus to a broader paradigm of ecological regulation. These next-generation agents are designed not merely to eliminate pathogens but to modulate microbial communities and provide multifunctional protection, thereby advancing the concept of ecological caries prevention. Broadly, these innovative peptides categorize into four distinct classes: specifically targeted AMPs, bioresponsive AMPs, antibiofilm peptides, and bifunctional anticaries peptides. The evolution of this field represents a convergence of diverse disciplines—spanning dentistry, microbiology, biochemistry, and biomaterials—reflecting the immense translational potential of these therapeutic agents.106–108 Notably, emerging evidence links dental erosion to the microbial proteolysis of protective salivary proteins within the acquired enamel pellicle,109 suggesting a novel therapeutic avenue: the development of AMPs capable of inhibiting bacterial proteases.
Despite significant progress and promising experimental data, several scientific and technological hurdles remain. A substantial proportion of novel anticaries AMPs have been validated predominantly in vitro using single-species biofilm models, leaving a gap in verification within complex, multi-species environments and in vivo systems. In contrast to the heavy emphasis placed on the antimicrobial, ecological, and anti-caries properties of AMPs, their safety profiling is often simplistic or overlooked. Considering their intended clinical application, these safety evaluations warrant significantly greater attention. Parallel to these safety concerns, while robust proteolytic resistance is equally critical for AMPs to function in the complex oral environment, independent stability assessments are largely absent in most studies. This omission likely stems from a retrospective assumption: once AMPs demonstrate efficacy in bacterial or in vivo models, researchers tend to implicitly assume adequate stability. This prevalent oversight highlights a critical blind spot in current methodologies and underscores the absolute necessity of incorporating dedicated stability assays in future research. Furthermore, many engineered peptides possess intricate architectures and extended amino acid sequences, factors that inevitably escalate production costs and hinder clinical scalability. Consequently, future research must prioritize four key areas: (1) the establishment of physiologically relevant in vitro and in vivo models that better mimic the oral microenvironment; (2) the optimization of peptide design to achieve structural simplicity while maintaining high potency at reduced concentrations; (3) the rigorous evaluation of AMP safety and stability within the complex oral environment to ensure rational and compliant clinical translation; and (4) the standardization of research protocols to facilitate reproducible pre-clinical assessment and the rapid transition of promising candidates into clinical trials.
Acknowledgment
This work was funded by the National Natural Science Foundation of China (No. 82572783 and 82100995), Shandong Provincial Natural Science Foundation (No. ZR2024QH186 and ZR2025MS1350), Youth Creative Team Program of Shandong Province (2025KJJ014), Weifang Kite Capital Scholars Program (No.ydxz2025001) College Students’ Innovative Entrepreneurial Training Plan Program (S202510438056, X2025204, and X2025202) and the University-Sponsored Domestic Visiting Scholar Program of Shandong Second Medical University.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no competing interests.
References
1. Bawaskar HS, Bawaskar PH. Oral diseases: a global public health challenge. Lancet. 2020;395(10219):185–21. doi:10.1016/s0140-6736(19)33016-8
2. Ahmad P, Hussain A, Siqueira WL. Mass spectrometry-based proteomic approaches for salivary protein biomarkers discovery and dental caries diagnosis: a critical review. Mass Spectrom Rev. 2024;43(4):826–856. doi:10.1002/mas.21822
3. Zeng Z, Shaffer JR, Wang X, et al. Genome-wide association studies of pit-and-fissure- and smooth-surface caries in permanent dentition. J Dent Res. 2013;92(5):432–437. doi:10.1177/0022034513481976
4. Moores CJ, Kelly SAM, Moynihan PJ. Systematic review of the effect on caries of sugars intake: ten-year update. J Dent Res. 2022;101(9):1034–1045. doi:10.1177/00220345221082918
5. Kim D, Barraza JP, Arthur RA, et al. Spatial mapping of polymicrobial communities reveals a precise biogeography associated with human dental caries. Proc Natl Acad Sci U S A. 2020;117(22):12375–12386. doi:10.1073/pnas.1919099117
6. He J, Cheng L. The oral microbiome: a key determinant of oral health. Adv Exp Med Biol. 2025;1472:133–149. doi:10.1007/978-3-031-79146-8_9
7. Gao X, Jiang S, Koh D, Hsu C-YS. Salivary biomarkers for dental caries. Periodontol 2000. 2016;70(1):128–141. doi:10.1111/prd.12100
8. Min KH, Kim KH, Ki MR, Pack SP. Antimicrobial peptides and their biomedical applications: a review. Antibiotics. 2024;13(9). doi:10.3390/antibiotics13090794
9. Bertrand B, Hernandez-Adame PL, Munoz-Garay C. How useful are antimicrobial peptide properties for predicting activity, selectivity, and potency? Curr Protein Pept Sci. 2025;26(1):22–40. doi:10.2174/0113892037317887240625054710
10. Florin T, Maracci C, Graf M, et al. An antimicrobial peptide that inhibits translation by trapping release factors on the ribosome. Nat Struct Mol Biol. 2017;24(9):752–757. doi:10.1038/nsmb.3439
11. Loth K, Vergnes A, Barreto C, et al. The ancestral N-terminal domain of big defensins drives bacterially triggered assembly into antimicrobial nanonets. mBio. 2019;10(5). doi:10.1128/mBio.01821-19
12. Zhang H, Lv J, Ma Z, Ma J, Chen J. Advances in antimicrobial peptides: mechanisms, design innovations, and biomedical potential. Molecules. 2025;30(7). doi:10.3390/molecules30071529
13. Zhang OL, Niu JY, Yu OY, Mei ML, Jakubovics NS, Chu CH. Peptide designs for use in caries management: a systematic review. Int J Mol Sci. 2023;24(4). doi:10.3390/ijms24044247
14. Xu B, Shaoyong W, Wang L, et al. Gut-targeted nanoparticles deliver specifically targeted antimicrobial peptides against clostridium perfringens infections. Sci Adv. 2023;9(39):eadf8782. doi:10.1126/sciadv.adf8782
15. Lei M, Jayaraman A, Van Deventer JA, Lee K. Engineering selectively targeting antimicrobial peptides. Annu Rev Biomed Eng. 2021;23(1):339–357. doi:10.1146/annurev-bioeng-010220-095711
16. Zhang OL, Niu JY, Yin IX, Yu OY, Mei ML, Chu CH. Growing global research interest in antimicrobial peptides for caries management: a bibliometric analysis. J Funct Biomater. 2022;13(4):210. doi:10.3390/jfb13040210
17. Niu JY, Yin IX, Wu WKK, Li Q-L, Mei ML, Chu CH. Antimicrobial peptides for the prevention and treatment of dental caries: a concise review. Arch Oral Biol. 2021;122:105022. doi:10.1016/j.archoralbio.2020.105022
18. Philip N, Suneja B, Walsh LJ. Ecological approaches to dental caries prevention: paradigm shift or shibboleth? Caries Res. 2018;52(1–2):153–165. doi:10.1159/000484985
19. Koo H, Allan RN, Howlin RP, Stoodley P, Hall-Stoodley L. Targeting microbial biofilms: current and prospective therapeutic strategies. Nat Rev Microbiol. 2017;15(12):740–755. doi:10.1038/nrmicro.2017.99
20. Zhang JS, Chu C-H, Yu OY. Oral microbiome and dental caries development. Dent J. 2022;10(10). doi:10.3390/dj10100184
21. Drayton M, Kizhakkedathu JN, Straus SK. Towards robust delivery of antimicrobial peptides to combat bacterial resistance. Molecules. 2020;25(13):3048. doi:10.3390/molecules25133048
22. Tabarzad M, Torshabi M, Haeri A, Fathi F, Mortazavi SM. Peptide-based antibiotics: structure-driven strategies to tackle toxicity and resistance of antimicrobial peptides. Bioorg Med Chem. 2026;133:118486. doi:10.1016/j.bmc.2025.118486
23. Dad N, Elsawy MA, Humphreys G, Pluen A, Lu JR, McBain AJ. A critical view of antimicrobial peptides: exploring their potential and the barriers to realization. J Appl Microbiol. 2025;136(5). doi:10.1093/jambio/lxaf087
24. Chen CH, Lu TK. Development and challenges of antimicrobial peptides for therapeutic applications. Antibiotics. 2020;9(1). doi:10.3390/antibiotics9010024
25. Jiang W, Xie Z, Huang S, et al. Targeting cariogenic pathogens and promoting competitiveness of commensal bacteria with a novel pH-responsive antimicrobial peptide. J Oral Microbiol. 2023;15(1):2159375. doi:10.1080/20002297.2022.2159375
26. Lu J, Xu H, Xia J, et al. D- and unnatural amino acid substituted antimicrobial peptides with improved proteolytic resistance and their proteolytic degradation characteristics. Front Microbiol. 2020;11:563030. doi:10.3389/fmicb.2020.563030
27. Ciren D, Xia X, Zhu Y, Hui S, Hong L. The role and progress of antimicrobial peptides in managing oral biofilms: a narrative review. International Dental Journal. 2026;76(1):104022. doi:10.1016/j.identj.2025.104022
28. Eckert R, Qi F, Yarbrough DK, He J, Anderson MH, Shi W. Adding selectivity to antimicrobial peptides: rational design of a multidomain peptide against pseudomonas spp. Antimicrob Agents Chemother. 2006;50(4):1480–1488. doi:10.1128/aac.50.4.1480-1488.2006
29. Sztukowska MN, Roky M, Demuth DR. Peptide and non-peptide mimetics as potential therapeutics targeting oral bacteria and oral biofilms. Mol Oral Microbiol. 2019;34(5):169–182. doi:10.1111/omi.12267
30. Eckert R, He J, Yarbrough DK, Qi F, Anderson MH, Shi W. Targeted killing of Streptococcus mutans by a pheromone-guided “smart” antimicrobial peptide. Antimicrob Agents Chemother. 2006;50(11):3651–3657. doi:10.1128/aac.00622-06
31. Guo L, McLean JS, Yang Y, et al. Precision-guided antimicrobial peptide as a targeted modulator of human microbial ecology. Proc Natl Acad Sci U S A. 2015;112(24):7569–7574. doi:10.1073/pnas.1506207112
32. Huo L, Huang X, Ling J, Liu H, Liu J. Selective activities of STAMPs against Streptococcus mutans. Exp Ther Med. 2018;15(2):1886–1893. doi:10.3892/etm.2017.5631
33. Kaplan CW, Sim JH, Shah KR, Kolesnikova-Kaplan A, Shi W, Eckert R. Selective membrane disruption: mode of action of C16G2, a specifically targeted antimicrobial peptide. Antimicrob Agents Chemother. 2011;55(7):3446–3452. doi:10.1128/aac.00342-11
34. Li L-N, Guo L-H, Lux R, et al. Targeted antimicrobial therapy against Streptococcus mutans establishes protective non-cariogenic oral biofilms and reduces subsequent infection. Int J Oral Sci. 2010;2(2):66–73. doi:10.4248/ijos10024
35. Sullivan R, Santarpia P, Lavender S, et al. Clinical efficacy of a specifically targeted antimicrobial peptide mouth rinse: targeted elimination of Streptococcus mutans and prevention of demineralization. Caries Res. 2011;45(5):415–428. doi:10.1159/000330510
36. Ting DSJ, Beuerman RW, Dua HS, Lakshminarayanan R, Mohammed I. Strategies in Translating the Therapeutic Potentials of Host Defense Peptides. Front Immunol. 2020;11:983. doi:10.3389/fimmu.2020.00983
37. Mishra R, Panda AK, De Mandal S, Shakeel M, Bisht SS, Khan J. Natural anti-biofilm agents: strategies to control biofilm-forming pathogens. Front Microbiol. 2020;11:566325. doi:10.3389/fmicb.2020.566325
38. Li X, Jiang Y, Lin Y. Production of antimicrobial peptide arasin-like Sp in Escherichia coli via an ELP-intein self-cleavage system. J Biotechnol. 2022;347:49–55. doi:10.1016/j.jbiotec.2022.02.010
39. Mejía-Pitta A, Broset E, de la Fuente-Nunez C. Probiotic engineering strategies for the heterologous production of antimicrobial peptides. Adv Drug Deliv Rev. 2021;176:113863. doi:10.1016/j.addr.2021.113863
40. Montesinos L, Bundó M, Izquierdo E, et al. Production of biologically active cecropin a peptide in rice seed oil bodies. PLoS One. 2016;11(1):e0146919. doi:10.1371/journal.pone.0146919
41. Pandi A, Adam D, Zare A, et al. Cell-free biosynthesis combined with deep learning accelerates de novo-development of antimicrobial peptides. Nat Commun. 2023;14(1):7197. doi:10.1038/s41467-023-42434-9
42. Chunxiao C, Keyu J, Yuanyuan M, et al. Biological characteristics of a human specifically targeted antimicrobial peptide C16LL-37 against Streptococcus mutans. Hua Xi Kou Qiang Yi Xue Za Zhi. 2016;34(3):295–301. doi:10.7518/hxkq.2016.03.016
43. He J, Yarbrough DK, Kreth J, Anderson MH, Shi W, Eckert R. Systematic approach to optimizing specifically targeted antimicrobial peptides against Streptococcus mutans. Antimicrob Agents Chemother. 2010;54(5):2143–2151. doi:10.1128/aac.01391-09
44. Lyu B, Zhou H, Wang H, et al. Empowering healthcare with phage display technology. ACS Biomater Sci Eng. 2025;11:3228–3248. doi:10.1021/acsbiomaterials.5c00218
45. Konakbayeva D, Karlsson AJ. Strategies and opportunities for engineering antifungal peptides for therapeutic applications. Curr Opin Biotechnol. 2023;81:102926. doi:10.1016/j.copbio.2023.102926
46. Xu W, Peng L, Li C, et al. A novel fully human recombinant antibody neutralizing α-hemolysin of Staphylococcus aureus. Apmis. 2022;130(9):578–589. doi:10.1111/apm.13258
47. Chu JJK, Poh WH, Hasnuddin NTB, et al. Novel phage lysin Abp013 against Acinetobacter baumannii. Antibiotics. 2022;11(2). doi:10.3390/antibiotics11020169
48. Kupper MB, Huhn M, Spiegel H, et al. Generation of human antibody fragments against Streptococcus mutansusing a phage display chain shuffling approach. BMC Biotechnol. 2005;5(1):4. doi:10.1186/1472-6750-5-4
49. Alam MK, Zheng L, Liu R, Papagerakis S, Papagerakis P, Geyer CR. Synthetic antigen-binding fragments (Fabs) against S. mutans and S. sobrinus inhibit caries formation. Sci Rep. 2018;8(1):10173. doi:10.1038/s41598-018-28240-0
50. Lee J, Shin W-R, Kim Y-H, Ahn J-Y, Chae S, Min J. Targeted inhibition of oral biofilm formation using phage-derived high-affinity peptides. J Biotechnol. 2025;402:51–58. doi:10.1016/j.jbiotec.2025.03.009
51. Luo J, Wang Y, Wang K, Jiang W, Li X, Zhang L. Comparative proteomic analysis on acquired enamel pellicle at two time points in caries-susceptible and caries-free subjects. J Dent. 2020;94:103301. doi:10.1016/j.jdent.2020.103301
52. Luo J, Feng Z, Jiang W, et al. Novel lactotransferrin-derived synthetic peptides suppress cariogenic bacteria in vitro and arrest dental caries in vivo. J Oral Microbiol. 2021;13(1):1943999. doi:10.1080/20002297.2021.1943999
53. Feng Z, Luo J, Lyu X, Chen Y, Zhang L. Selective antibacterial activity of a novel lactotransferrin-derived antimicrobial peptide LF-1 against Streptococcus mutans. Arch Oral Biol. 2022;139:105446. doi:10.1016/j.archoralbio.2022.105446
54. Zhang B, Zhao M, Tian JA, Lei L, Huang RZ. Novel antimicrobial agents targeting the Streptococcus mutans biofilms discovery through computer technology. Front Cell Infect Mi. 2022;12. doi:10.3389/fcimb.2022.1065235
55. Yang SR, Zhang J, Yang R, Xu X. Small molecule compounds, a novel strategy against Streptococcus mutans. Pathogens. 2021;10(12):1540. doi:10.3390/pathogens10121540
56. Zeng H, Liu J, Ling J. Efflux inhibitor suppresses Streptococcus mutans virulence properties. FEMS Microbiol Lett. 2017;364(7). doi:10.1093/femsle/fnx033
57. Sekiya M. Proton pumping ATPases: rotational catalysis, physiological roles in oral pathogenic bacteria, and inhibitors. Biol Pharm Bull. 2022;45(10):1404–1411. doi:10.1248/bpb.b22-00396
58. Duprez W, Premkumar L, Halili MA, et al. Peptide inhibitors of the escherichia coli DsbA oxidative machinery essential for bacterial virulence. J Med Chem. 2015;58(2):577–587. doi:10.1021/jm500955s
59. Zhou L, Liu Q, Fang Z, Li QL, Wong HM. Targeted antimicrobial self-assembly peptide hydrogel with in situ bio-mimic remineralization for caries management. Bioact Mater. 2025;44:428–446. doi:10.1016/j.bioactmat.2024.10.022
60. Jiang S, Zha Y, Zhao T, et al. Temporin-derived peptides disrupt the exopolysaccharide matrix of Streptococcus mutans to prevent related dental caries. J Agric Food Chem. 2024;72(47):26236–26247. doi:10.1021/acs.jafc.4c09488
61. Wei H, Xie Z, Tan X, et al. Temporin-like peptides show antimicrobial and anti-biofilm activities against Streptococcus mutans with reduced hemolysis. Molecules. 2020;25(23):5724. doi:10.3390/molecules25235724
62. Luo J, Feng Z, Lyu X, Zhang L. Novel lactotransferrin-derived antimicrobial peptide LF-1 inhibits the cariogenic virulence factors of Streptococcus mutans. Antibiotics. 2023;12(3). doi:10.3390/antibiotics12030563
63. Jiang W, Luo J, Wang Y, et al. The pH-responsive property of antimicrobial peptide GH12 enhances its anticaries effects at acidic pH. Caries Res. 2021;55(1):21–31. doi:10.1159/000508458
64. Jiang W, Wang Y, Luo J, et al. Antimicrobial peptide GH12 prevents dental caries by regulating dental plaque microbiota. Appl Environ Microbiol. 2020;86(14). doi:10.1128/aem.00527-20
65. Johansson I, Witkowska E, Kaveh B, Lif Holgerson P, Tanner AC. The microbiome in populations with a low and high prevalence of caries. J Dent Res. 2016;95(1):80–86. doi:10.1177/0022034515609554
66. Meriç E, Bolgül B, Duran N, Ay E. Evaluation of oral streptococci in saliva of children with severe early childhood caries and caries-free. Eur J Paediatr Dent. 2020;21(1):13–17. doi:10.23804/ejpd.2020.21.01.03
67. Simon-Soro A, Sherriff A, Sadique S, et al. Combined analysis of the salivary microbiome and host defence peptides predicts dental disease. Sci Rep. 2018;8(1):1484. doi:10.1038/s41598-018-20085-x
68. AlEraky DM, Madi M, El Tantawi M, et al. Predominance of non-Streptococcus mutans bacteria in dental biofilm and its relation to caries progression. Saudi J Biol Sci. 2021;28(12):7390–7395. doi:10.1016/j.sjbs.2021.08.052
69. Spatafora G, Li Y, He X, Cowan A, Tanner ACR. The evolving microbiome of dental caries. Microorganisms. 2024;12(1). doi:10.3390/microorganisms12010121
70. Wang Y, Fan Y, Zhou Z, et al. De novo synthetic short antimicrobial peptides against cariogenic bacteria. Arch Oral Biol. 2017;80:41–50. doi:10.1016/j.archoralbio.2017.03.017
71. Tu H, Fan Y, Lv X, Han S, Zhou X, Zhang L. Activity of synthetic antimicrobial peptide GH12 against oral Streptococci. Caries Res. 2016;50(1):48–61. doi:10.1159/000442898
72. Wang Y, Wang X, Jiang W, et al. Antimicrobial peptide GH12 suppresses cariogenic virulence factors of Streptococcus mutans. J Oral Microbiol. 2018;10(1):1442089. doi:10.1080/20002297.2018.1442089
73. Jiang W, Wang Y, Luo J, et al. Effects of antimicrobial peptide GH12 on the cariogenic properties and composition of a cariogenic multispecies biofilm. Appl Environ Microbiol. 2018;84(24). doi:10.1128/aem.01423-18
74. Calinsky R, Levy Y. Histidine in proteins: pH-dependent interplay between π–π, Cation–π, and CH–π interactions. J Chem Theory Comput. 2024;20(15):6930–6945. doi:10.1021/acs.jctc.4c00606
75. Wang Z, Li Q, Li J, et al. pH-responsive antimicrobial peptide with selective killing activity for bacterial abscess therapy. J Med Chem. 2022;65(7):5355–5373. doi:10.1021/acs.jmedchem.1c01485
76. Zhang P, Wu S, Li J, et al. Dual-sensitive antibacterial peptide nanoparticles prevent dental caries. Theranostics. 2022;12(10):4818–4833. doi:10.7150/thno.73181
77. Wang ZJ, Shen Y, Haapasalo M. Antibiofilm peptides against oral biofilms. J Oral Microbiol. 2017;9:1327308. doi:10.1080/20002297.2017.1327308
78. Chen DR, Li J, Pan T, Wu RX, Tao Y, Lin HC. The broad-spectrum antibiofilm activity of amyloid-forming hexapeptides. Microb Biotechnol. 2021;14(2):656–667. doi:10.1111/1751-7915.13721
79. Huang Y, Wang T, Chen Y, Lin H, Chen D. Amyloid hexapeptide prevent dental caries by antibiofilm formation. J Dent. 2023;135:104596. doi:10.1016/j.jdent.2023.104596
80. Kokilakanit P, Koontongkaew S, Roytrakul S, Utispan K. A novel non-cytotoxic synthetic peptide, Pug-1, exhibited an antibiofilm effect on Streptococcus mutans adhesion. Lett Appl Microbiol. 2020;70(3):151–158. doi:10.1111/lam.13265
81. Guo JM, Chan EWC, Chen S, Zeng Z. Development of a novel quantum dots and graphene oxide based FRET assay for rapid detection of invA gene of salmonella. Front Microbiol. 2017;8:8. doi:10.3389/fmicb.2017.00008
82. Xu VW, Yin IX, Niu JY, Yu OY, Nizami MZI, Chu CH. Antibacterial and mineralising properties of copper tetraamine fluoride on dentine caries: a laboratory study. J Dent. 2024;151:105444. doi:10.1016/j.jdent.2024.105444
83. Cheng YH, Liu BQ, Cui B, et al. Alanine substitution to determine the effect of LR5 and YR6 rice peptide structure on antioxidant and anti-inflammatory activity. Nutrients. 2023;15(10):2373. doi:10.3390/nu15102373
84. Pan Q, Lin S, Li Y, et al. A novel GLP-1 and FGF21 dual agonist has therapeutic potential for diabetes and non-alcoholic steatohepatitis. EBioMedicine. 2021;63:103202. doi:10.1016/j.ebiom.2020.103202
85. Wongchai M, Wongkaewkhiaw S, Kanthawong S, Roytrakul S, Aunpad R. Dual-function antimicrobial-antibiofilm peptide hybrid to tackle biofilm-forming Staphylococcus epidermidis. Ann Clin Microbiol Antimicrob. 2024;23(1):44. doi:10.1186/s12941-024-00701-7
86. Wang X, Wang Y, Wang K, et al. Bifunctional anticaries peptides with antibacterial and remineralizing effects. Oral Dis. 2019;25(2):488–496. doi:10.1111/odi.12990
87. Niu JY, Yin IX, Wu WKK, Li Q-L, Mei ML, Chu CH. A novel dual-action antimicrobial peptide for caries management. J Dent. 2021;111:103729. doi:10.1016/j.jdent.2021.103729
88. Niu JY, Yin IX, Wu WKK, Li Q-L, Mei ML, Chu CH. Remineralising dentine caries using an artificial antimicrobial peptide: an in vitro study. J Dent. 2021;111:103736. doi:10.1016/j.jdent.2021.103736
89. Niu JY, Yin IX, Wu WKK, Li Q-L, Mei ML, Chu CH. Efficacy of the dual-action GA-KR12 peptide for remineralising initial enamel caries: an in vitro study. Clin Oral Investig. 2022;26(3):2441–2451. doi:10.1007/s00784-021-04210-1
90. Gagat P, Ostrówka M, Duda-Madej A, Mackiewicz P. Enhancing antimicrobial peptide activity through modifications of charge, hydrophobicity, and structure. Int J Mol Sci. 2024;25(19):10821. doi:10.3390/ijms251910821
91. Huang Z-B, Shi X, Mao J, Gong S-Q. Design of a hydroxyapatite-binding antimicrobial peptide with improved retention and antibacterial efficacy for oral pathogen control. Sci Rep. 2016;6(1):38410. doi:10.1038/srep38410
92. Yu Q, Wu Z, Chen H. Dual-function antibacterial surfaces for biomedical applications. Acta Biomater. 2015;16:1–13. doi:10.1016/j.actbio.2015.01.018
93. Zhou L, Wong HM, Zhang YY, Li QL. Constructing an antibiofouling and mineralizing bioactive tooth surface to protect against decay and promote self-healing. ACS Appl Mater Interfaces. 2020;12(2):3021–3031. doi:10.1021/acsami.9b19745
94. Zhou L, Li QL, Wong HM. A novel strategy for caries management: constructing an antibiofouling and mineralizing dual-bioactive tooth surface. ACS Appl Mater Interfaces. 2021;13(26):31140–31152. doi:10.1021/acsami.1c06989
95. Hsueh HT, Chou RT, Rai U, et al. Machine learning-driven multifunctional peptide engineering for sustained ocular drug delivery. Nat Commun. 2023;14(1):2509. doi:10.1038/s41467-023-38056-w
96. Diener C, Garza Ramos Martínez G, Moreno Blas D, et al. Effective design of multifunctional peptides by combining compatible functions. PLoS Comput Biol. 2016;12(4):e1004786. doi:10.1371/journal.pcbi.1004786
97. Wang J, Zhang R, Zhao X, et al. Hybridization design and high-throughput screening of peptides with immunomodulatory and antioxidant activities. Int J Mol Sci. 2025;26(2). doi:10.3390/ijms26020505
98. Xie Z, Dai C, Gao X, et al. Bioinspired multifunctional peptides integrated with anti-infective, immunomodulatory and osteogenic activities for infectious bone regeneration. Chem Eng J. 2025;503:158256. doi:10.1016/j.cej.2024.158256
99. Thomadaki K, Helmerhorst EJ, Tian N, et al. Whole-saliva proteolysis and its impact on salivary diagnostics. J Dent Res. 2011;90(11):1325–1330. doi:10.1177/0022034511420721
100. Makambi WK, Chiu VL, Kasper L, Hube B, Karlsson AJ. Role of amino acid substitutions on proteolytic stability of histatin 5 in the presence of secreted aspartyl proteases and salivary proteases. Protein Sci. 2025;34(1):e70011. doi:10.1002/pro.70011
101. Cavaco M, Valle J, Flores I, Andreu D, Arbc M. Estimating peptide half-life in serum from tunable, sequence-related physicochemical properties. Clin Transl Sci. 2021;14(4):1349–1358. doi:10.1111/cts.12985
102. Mendes B, Castelletto V, Hamley IW, Barrett G. D-amino acid substitution and cyclization enhance the stability and antimicrobial activity of arginine-rich peptides. Microbiology. 2026;172(2). doi:10.1099/mic.0.001657
103. Mwangi J, Kamau PM, Thuku RC, Lai R. Design methods for antimicrobial peptides with improved performance. Zool Res. 2023;44(6):1095–1114. doi:10.24272/j.issn.2095-8137.2023.246
104. Brango-Vanegas J, Leite ML, Macedo MLR, Cardoso MH, Franco OL. Capping motifs in antimicrobial peptides and their relevance for improved biological activities. Front Chem. 2024;12:1382954. doi:10.3389/fchem.2024.1382954
105. Yeh J-C, Hazam PK, Hsieh C-Y, et al. Rational design of stapled antimicrobial peptides to enhance stability and in vivo potency against polymicrobial sepsis. Microbiol Spectr. 2023;11(2):e0385322. doi:10.1128/spectrum.03853-22
106. Moussa DG, Aparicio C, Kreth J. Targeting the oral plaque microbiome with immobilized anti-biofilm peptides at tooth-restoration interfaces. PLoS One. 2020;15(7):e0235283. doi:10.1371/journal.pone.0235283
107. Moussa DG, Kung RW, Tse JS, Siqueira WL. Mechanistic insights into bioengineered antibiofilm enamel pellicles. J Dent Res. 2023;102(7):743–751. doi:10.1177/00220345231162336
108. Pfeifer CS, Lucena FS, Logan MG, Nair D, Lewis SH. Current approaches to produce durable biomaterials: trends in polymeric materials for restorative dentistry applications. Dent Mater. 2024;40(12):2122–2134. doi:10.1016/j.dental.2024.10.004
109. Cleaver LM, Carda-Diéguez M, Moazzez R, Carpenter GH. Novel bacterial proteolytic and metabolic activity associated with dental erosion-induced oral dysbiosis. Microbiome. 2023;11(1):69. doi:10.1186/s40168-023-01514-0
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.