Back to Journals » Drug Design, Development and Therapy » Volume 9
Alpinetin inhibits lung cancer progression and elevates sensitization drug-resistant lung cancer cells to cis-diammined dichloridoplatium
Authors Wu L, Yang W, Zhang S, Lu J
Received 19 July 2015
Accepted for publication 10 September 2015
Published 16 November 2015 Volume 2015:9 Pages 6119—6127
DOI https://doi.org/10.2147/DDDT.S92702
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Wei Duan
Lin Wu, Wei Yang, Su-ning Zhang, Ji-bin Lu
Department of Thoracic Surgery, Sheng Jing Hospital of China Medical University, Shenyang, People’s Republic of China
Objective: Alpinetin is a novel flavonoid that has demonstrated potent antitumor activity in previous studies. However, the efficacy and mechanism of alpinetin in treating lung cancer have not been determined.
Methods: We evaluated the impact of different doses and durations of alpinetin treatment on the cell proliferation, the apoptosis of lung cancer cells, as well as the drug-resistant lung cancer cells.
Results: This study showed that the alpinetin inhibited the cell proliferation, enhanced the apoptosis, and inhibited the PI3K/Akt signaling in lung cancer cells. Moreover, alpinetin significantly increased the sensitivity of drug-resistant lung cancer cells to the chemotherapeutic effect of cis-diammined dichloridoplatium. Taken together, this study demonstrated that alpinetin significantly suppressed the development of human lung cancer possibly by influencing mitochondria and the PI3K/Akt signaling pathway and sensitized drug-resistant lung cancer cells.
Conclusion: Alpinetin may be used as a potential compound for combinatorial therapy or as a complement to other chemotherapeutic agents when multiple lines of treatments have failed to reduce lung cancer.
Keywords: alpinetin, cell proliferation and apoptosis, drug resistance reversal, PI3K/Akt, lung cancer
Introduction
Lung cancer has the highest incidence and mortality rate among malignant tumors and significantly impairs human health, especially in People’s Republic of China.1 According to the China Cancer Registry annual report in 2013, lung cancer had an incidence rate of 19.59% and was the highest among all malignant cancer types. Approximately 600,000 people in People’s Republic of China are diagnosed with lung cancer every year, which also has the highest mortality rate of 24.87%, causing 490,000 deaths each year.2 With the development of radiochemotherapy and the discovery of molecular targets, significant progress has been made in treating advanced-stage non-small-cell lung cancer and in improving the survival of patients. Nevertheless, the treatment of lung cancer is far from curative because the overall survival time of late-stage lung cancer remains considerably short despite these efforts.3 One of the primary causes of the poor prognosis of lung cancer is its tendency to develop multiple drug resistance.4 Alpinetin is a natural flavonoid primarily occurring in zingiberaceous plants. In the previous studies, potential antitumor activity of alpinetin toward gastric, liver, pancreatic, and breast cancers was demonstrated.5–8 However, the effectiveness of alpinetin for inhibiting lung cancer cells is unknown. In this study, we examined the antitumor efficacy and possible molecular mechanisms of action of alpinetin in the lung cancer.
Methods
Cell lines, reagents, and antibodies
Lung cancer cell lines A549, SK-MES-1, NCI-H292, and A549/cis-diammined dichloridoplatium (CDDP) were purchased from the Shanghai Institute of Cell Biology (Shanghai, People’s Republic of China), Chinese Academy of Science. The study was approved by the Ethics Committee of Shengjing Hospital of China.
Alpinetin was purchased from the National Institutes for Food and Drug Control (Beijing, People’s Republic of China). Fetal bovine serum, RPMI-1640 medium, and trypsin were purchased from Thermo Fisher Scientific, Waltham, MA, USA. Dimethyl sulfoxide, thiazolyl blue tetrazolium bromide (MTT), and Hoechst 33258 were purchased from Beyotime Institute of Biotechnology (Shanghai, People’s Republic of China). A dead cell apoptosis kit with annexin V-FITC and propidium iodide (PI) was purchased from Sigma-Aldrich Co., St Louis, MO, USA, and a transcription kit was purchased from Fermentas (Waltham, MA, USA).
Anti-Bcl-2, anti-Bcl-xL, anti-Bax, anti-XIAP, anticytochrome c, anti-PI3K, anti-Akt, anti-p-Akt, anti-MPR1, anti-MPR5, and anti-P-gp were all purchased from Bioworld (MN, USA). All of the secondary antibodies were purchased from Beijing Zhongshan Jinqiao Biotechnology Co., Ltd. (Beijing, People’s Republic of China), and an EPICS ALTRA flow cytometer was purchased from Beckman Coulter (Brea, FL, USA).
Cell culture
The lung cancer cells A549, SK-MES-1, NCI-H292, and A549/CDDP were cultured using RPMI-1640 containing 10% fetal bovine serum at 37°C and placed within an environment containing 5% CO2. To maintain drug resistance in the A549/CDDP cell media, 2 μg/mL CDDP was included in the cell media. All of the cell lines were passaged every 3–4 days.
MTT assay
Following digestion with 0.25% trypsin, A549, SK-MES-1, NCI-H292, and A549/CDDP cells were seeded into a 96-well plate at a density of 1×104/mL in a total volume of 100 μL. The culture medium contained a series of gradient concentrations of alpinetin (100 μL/well) according to the experimental plan for 24 hours. Then, the medium without alpinetin was added to the control wells. A total of 20 μL (0.5 mg/mL) of MTT was added into each well after 48 hours, and then the cells were returned to 37°C to continue culture for 4 hours. The cells in each well were treated with 200 μL dimethyl sulfoxide and shaken for 15 minutes. The absorbance of each well at 570 nm was then detected by a microplate reader.
Flow cytometry for apoptosis analysis
Following 48 hours of incubation, the A549 cells generally reached a density of 1×105 and were washed by phosphate-buffered saline (PBS) for three times. The cells were then resuspended with annexin V-FITC and PI staining solution, and the cells were gently mixed well. After incubating on ice in the dark for 15 minutes, the cell pellets were obtained by centrifuging for 5 minutes at 1,000 rpm and filtered through a 300 μm nylon mesh. The rate of cell apoptosis was determined by FACSCalibur flow cytometer (Becton-Dickinson, Franklin Lakes, NJ, USA). A total of 10,000 cells were examined.
Caspase activity analysis
A549 cells were cultured in a 96-well plate with 50 μL of RPMI-1640 containing 10% fetal bovine serum at 37°C in a humidified atmosphere of 5% CO2 and 95% air for 24 hours. The A549 cells were treated with different concentrations of alpinetin (0 μM, 50 μM, 100 μM, and 200 μM) 50 μL for 48 hours and then collected and tested for the viability of caspase-3, -8, and -9 using a caspase activity assay kit according to the manufacturer’s instructions (Beyotime Institute of Biotechnology).
Western blot analysis
A549 or A549/CDDP cells were lysed with ice-cold LaeμMli sodium dodecyl sulfate (SDS) sample buffer (pH 6.8), consisting of 25 μM Tris–HCl, 5% glycerol, 2.5% 2-mercaptoethanol, and 1% SDS, containing protease inhibitor cocktail. The lysates were incubated on ice for 30 minutes and centrifuged at 15,000 rpm for 20 minutes at 4°C. The supernatant was collected as a whole lysate. Equal amounts of protein (20 μg per lane) were loaded on a 10% SDS-polyacrylamide gel. The cell lysates were resolved by electrophoresis and transferred to membranes. Membranes were blocked with skim milk, probed with primary antibodies, washed, and then incubated with secondary antibody. Antibody dilutions used were as follows: anti-Bcl-2 (1/1,000), anti-Bcl-xL (1/500), anti-Bax (1/1,000), anti-XIAP (1/1,000), anticytochrome c (1/500), anti-PI3K (1/500), anti-Akt (1:500), anti-p-Akt (1/500), anti-MPR1 (1/1,000), anti-MPR5 (1/1,000), and anti-P-gp (1/500).
Xenograft assays in nude mice
Twenty-four male (5–6 weeks old) nude mice were purchased from the Animal Experiment Center of Guilin Medical University. The animal experimentation program was approved by the Medical Ethics Committee of Guilin Medical University. Lung cancer A549/CDDP cells were collected in the logarithmic log phase. After washing with PBS, the cells were suspended in a serum-free medium. Subsequently, 200 μL of the cell suspension (containing 2×107 cells) was injected subcutaneously into the right groin area of the mice. The mice were randomly divided into three groups of three mice each: control group (PBS injections weekly for 4 weeks), CDDP group (CDDP [2 mg/kg] injections weekly for 4 weeks), and CDDP + alpinetin group (CDDP [2 mg/kg] and alpinetin [50 mg/kg] injections weekly for 4 weeks). After tumors formed, the mice were observed for the growth of tumors at 7 days, 14 days, 21 days, and 28 days. Tumor volumes were calculated using the following formula: tumor volume (μM3) =0.5× length (μM) × width2 (μM2). The mice were sacrificed by cervical vertebral dislocation 4 weeks later. Tumors were excised and weighed to record the wet tumor weight. Subcutaneously transplanted tumor tissue was surgically removed under aseptic conditions for index analysis.
Statistical analysis
Differences among the groups used as qualitative data were analyzed using a chi-square test. The quantitative data were expressed as the mean ± SD. A t-test was used to compare the averages between the two groups, and one-way analysis of variance was used to analyze the differences between multiple groups. The least significant difference method was used to identify the pairwise differences among the groups. All of the statistical analyses were conducted using SPSS 17.0 statistical software, and a value of P<0.05 indicated that the difference was statistically significant.
Results
Alpinetin inhibited lung cancer cell growth
We treated A549, SK-MES-1, and NCI-H292 cells and the multidrug resistant A549/CDDP cells with gradually increasing doses of alpinetin (0 μM, 50 μM, 100 μM, 200 μM, and 400 μM) for 24 hours, 48 hours, and 72 hours. Cell survival was then evaluated by the MTT assay. The results showed that the proliferation of A549, SK-MES-1, and NCI-H292 cells was inhibited in a dose- and time-dependent manner by alpinetin treatment (Figure 1). The antiproliferative effect of alpinetin increased gradually over time and became particularly apparent at 72 hours. And the inhibition effect was obvious at a concentration of 200 μM. However, no growth inhibition was observed in the A549/CDDP cells. This result suggests that multidrug-resistant lung cancer cells are also resistant to the direct growth-inhibitory effect of alpinetin in vitro. The A549 and A549/CDDP cells were selected based on the goal of the experiment. To further investigate the mechanism of growth inhibition by alpinetin in these cells, A549 and A549/CDDP cells were treated with alpinetin at 0–200 μM concentrations for 72 hours.
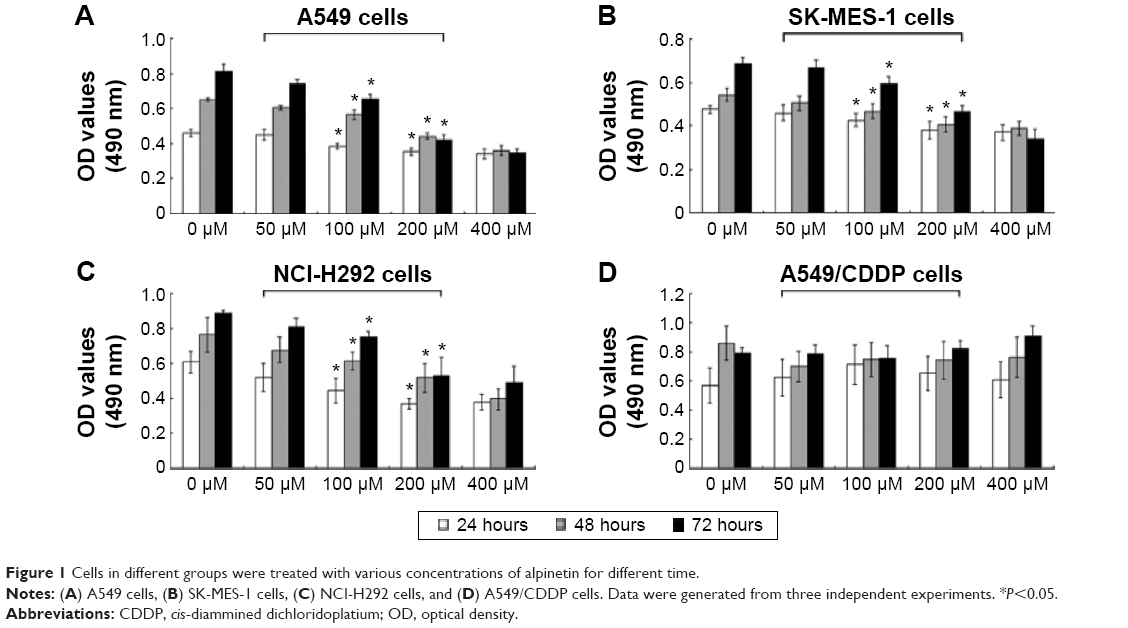 | Figure 1 Cells in different groups were treated with various concentrations of alpinetin for different time. |
Alpinetin induced lung cancer cell apoptosis and caspase activation
To determine whether the antiproliferative effects of alpinetin on A549 cells were a result of the induction of cell apoptosis, we treated A549 cells with various concentrations of alpinetin for 72 hours and evaluated cell apoptosis via annexin V-FITC/PI double staining coupled with flow cytometric analysis (Figure 2). The results indicated that the cell apoptotic rate was apparently increased by increasing the treatment dose of alpinetin and reached statistical significance at concentrations of 100 μM and 200 μM compared with the nontreated control group (P<0.05) (Figure 2A). In addition, the activation of caspase-3, -8, and -9 was examined (Figure 2B), and we observed that the activation of caspase-3 and caspase-9 was significantly increased in both the 100 μM and 200 μM alpinetin-treated A549 cells (Figure 2B) (P<0.05). In the 200 μM alpinetin-treated A549 cells, caspase-8 activation reached statistical significance (P<0.05). These data suggest that alpinetin induced dose-dependent A549 cell apoptosis via activating caspases and the caspase-dependent cell apoptotic pathway, thereby suppressing A549 cell proliferation.
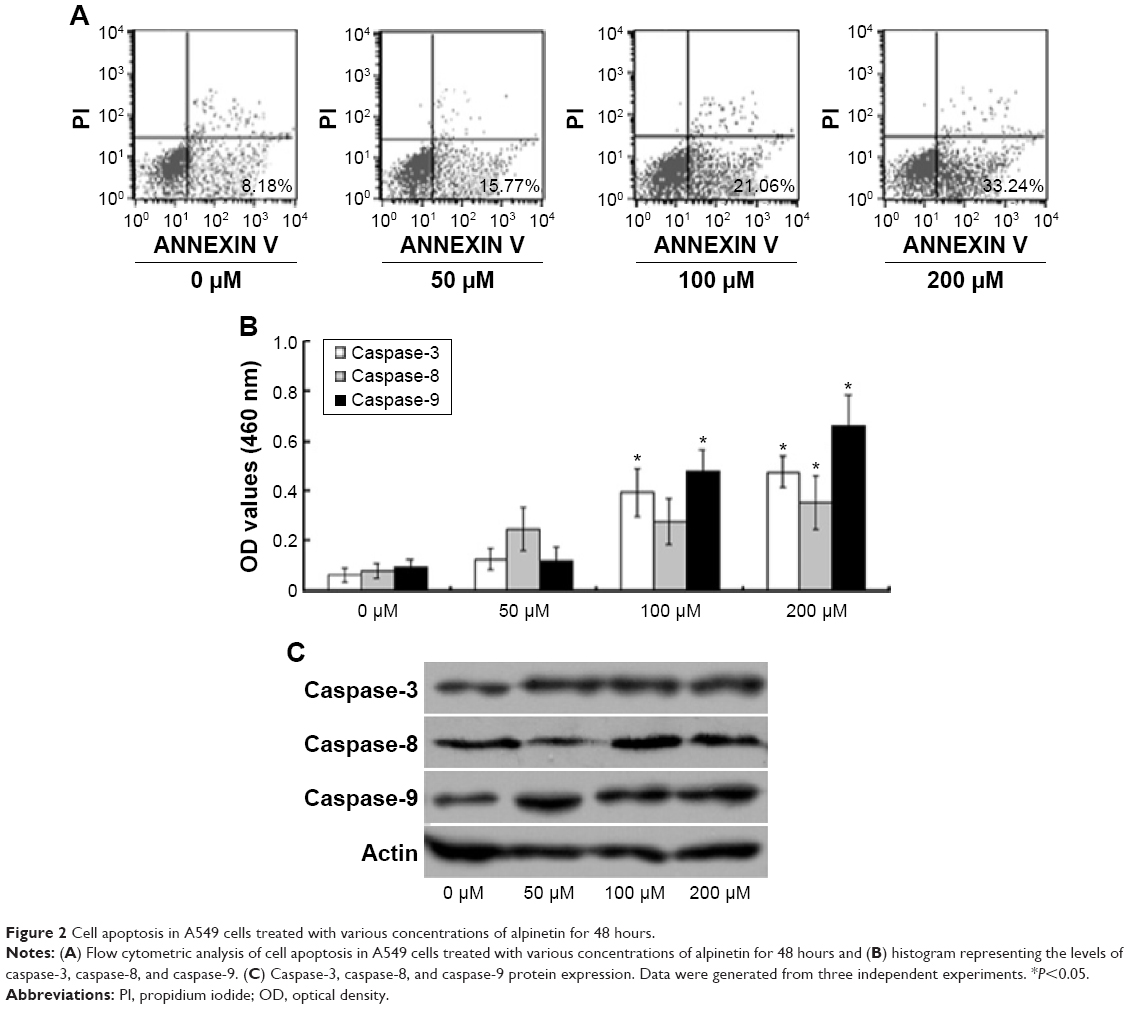 | Figure 2 Cell apoptosis in A549 cells treated with various concentrations of alpinetin for 48 hours. |
Alpinetin regulated the expression of Bcl-2 family member XIAP and cytochrome c
To explore the mechanism underlying alpinetin-mediated inhibition of cell proliferation and induction of cell apoptosis, we measured the expression changes in the Bcl-2 family member XIAP and cytochrome c in the cytosol and mitochondria in A549 cells treated with alpinetin for 72 hours (Figure 3). The data suggest that the distribution of cytochrome c was changed after alpinetin treatment, and the changes were significantly dose dependent. The changes of cytochrome c were significant at 200 μM, and cytochrome c protein expression was significantly decreased in the mitochondria (P<0.05) and increased in the cytosol (P<0.05) (Figure 3A and B). In addition, protein expression of the antiapoptotic proteins Bcl-2, Bcl-xL, and XIAP decreased in a dose-dependent manner by alpinetin treatment, whereas proapoptotic protein Bax increased in a dose-dependent manner in response to alpinetin. This difference reached statistical significance at 200 μM (P<0.05) (Figure 3C and D). These results indicate that alpinetin is able to activate the mitochondria-dependent cell apoptosis pathway and induce cell apoptosis and inhibit A549 cell proliferation via regulating the expression of the Bcl-2 family member XIAP and cytochrome c.
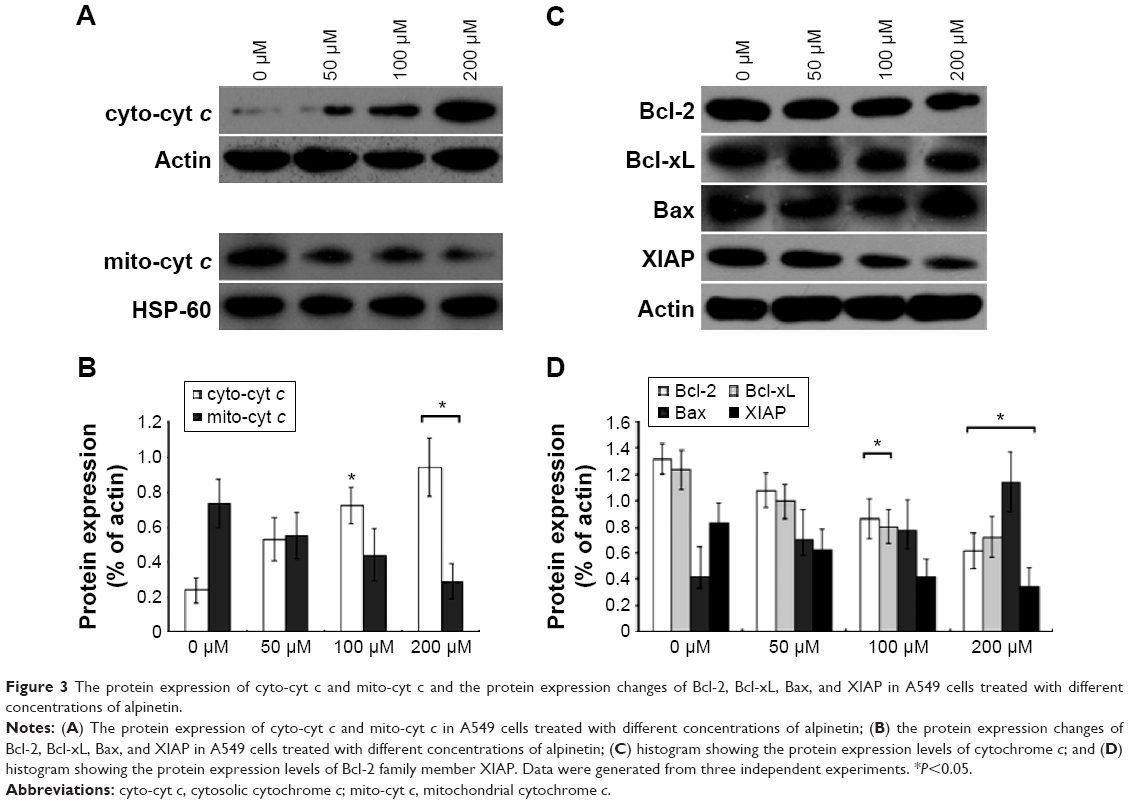 | Figure 3 The protein expression of cyto-cyt c and mito-cyt c and the protein expression changes of Bcl-2, Bcl-xL, Bax, and XIAP in A549 cells treated with different concentrations of alpinetin. |
Alpinetin regulated protein expression in the PI3K/Akt signaling pathway in lung cancer cells
To further examine the mechanism underlying the alpinetin-mediated antiproliferative and proapoptotic effects, Western blot analyses were used to evaluate the expression of proteins in the PI3K/Akt signaling pathway in A549 cells treated with alpinetin for 72 hours. The data showed that alpinetin induced a dose-dependent decrease in PI3K and p-Akt expressions, and this decrease was significant at a concentration of 200 μM (P<0.05) (Figure 4). In contrast, the expression of Akt did not exhibit significant differences in response to alpinetin treatment (P<0.05). These results suggest that alpinetin may reduce the antiapoptotic effects of the PI3K/Akt pathway through suppressing its activity, thereby inhibiting the cell proliferation of lung cancer cells.
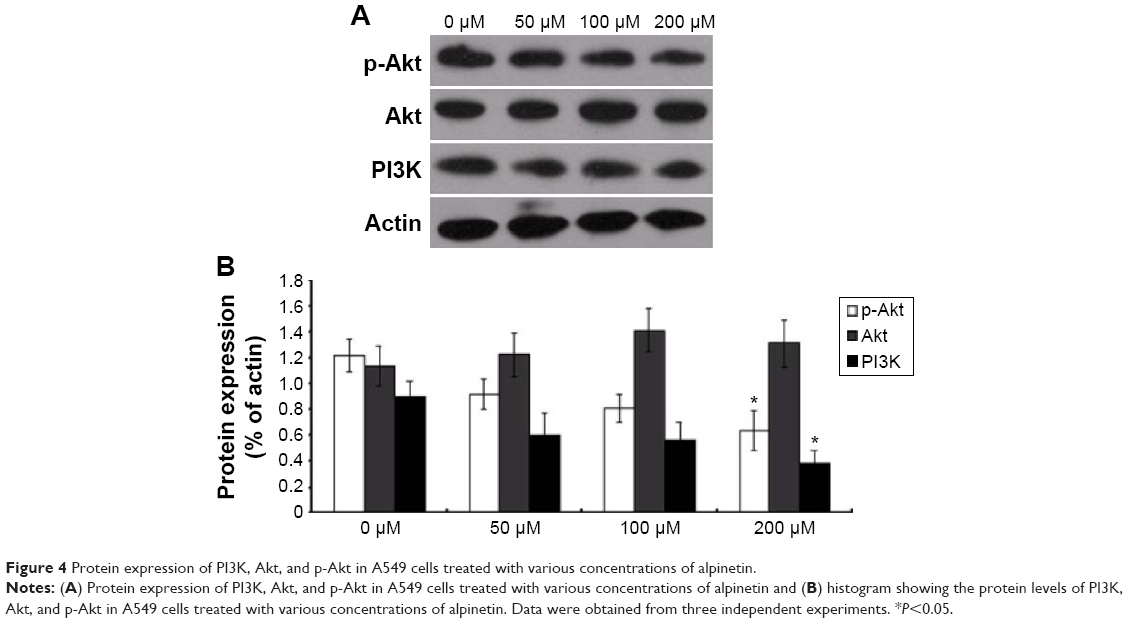 | Figure 4 Protein expression of PI3K, Akt, and p-Akt in A549 cells treated with various concentrations of alpinetin. |
Alpinetin reversed the resistance of A549/CDDP cells toward the chemotherapeutic effects of CDDP
When the CDDP-resistant A549/CDDP cells were pretreated with 200 μM alpinetin for 72 hours, the CDDP treatment inhibited the proliferation of A549/CDDP cells (P<0.05), indicating that A549/CDDP cells regained their sensitivity in the CDDP treatment and achieved a reversal of CDDP resistance in these cells (Figure 5A). The results suggest that the therapeutic dosage of alpinetin could reverse the CDDP resistance of A549/CDDP cells. Western blot analyses were used to further examine the protein expression levels of MPR1, MPR5, and P-gp, which are known to mediate drug resistance in A549/CDDP cells. The results showed that alpinetin induced a significant decrease in MPR1, MPR5, and P-gp protein expressions (P<0.05) (Figure 5B and C), which is an important mechanism underlying the alpinetin-mediated reversal of CDDP resistance in the A549/CDDP cells, in which the expression of drug resistance-related proteins was inhibited by alpinetin.
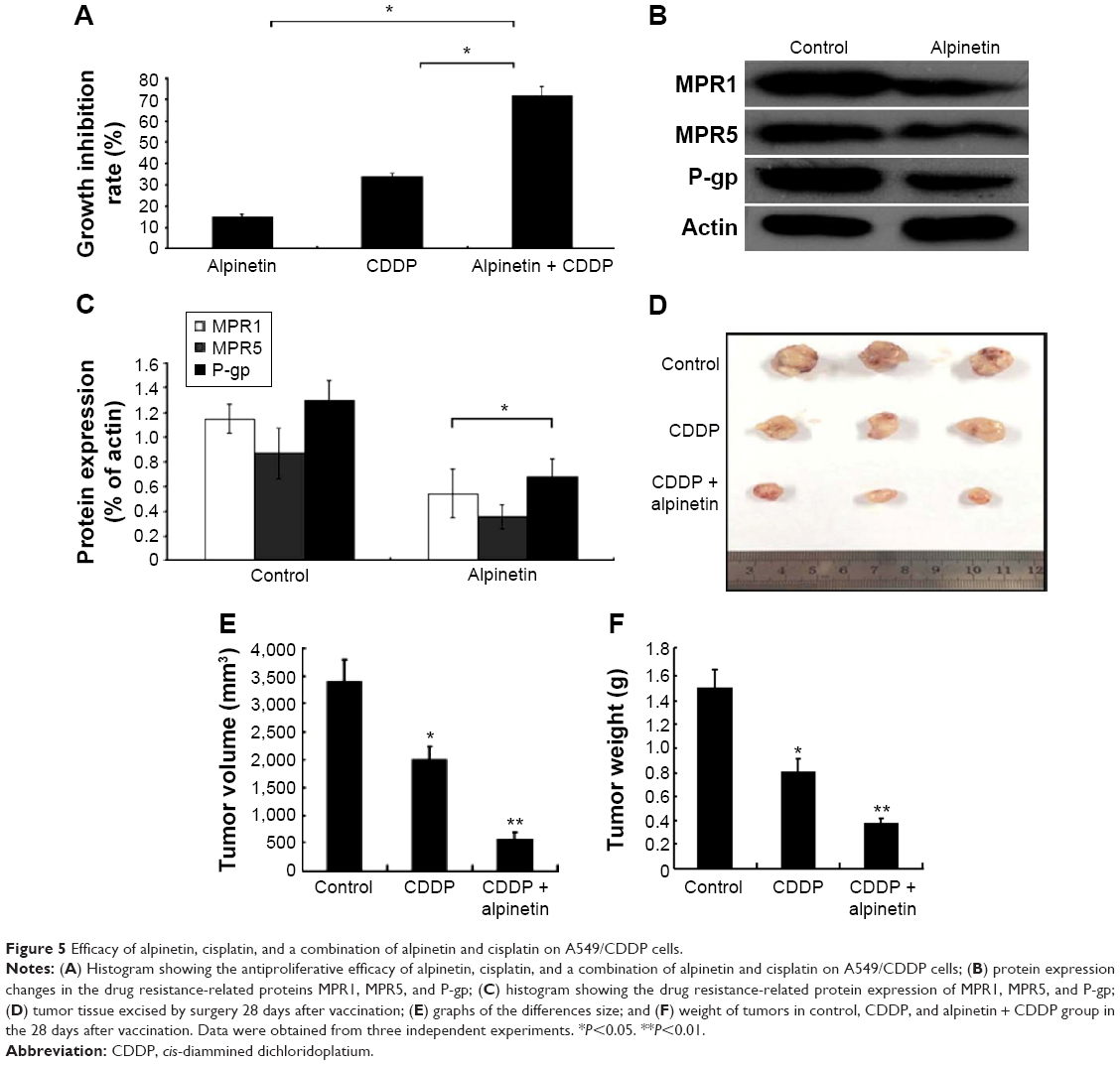 | Figure 5 Efficacy of alpinetin, cisplatin, and a combination of alpinetin and cisplatin on A549/CDDP cells. |
Alpinetin inhibited transplanted tumor growth in nude mice
We found that the tumor volume of alpinetin (50 mg/kg) injections + CDDP (2 mg/kg) group mice was consistently smaller than that of the control group or the CDDP (2 mg/kg) group mice from 14 days after vaccination. Additionally, in comparison with the control group and CDDP group, the tumor sizes and weight obtained by surgery in the 28 days after vaccination were significantly decreased (Figure 5D–F). These findings strongly indicate that alpinetin obviously inhibits the progression of lung cancer and sensitizes drug-resistant lung cancer cells toward chemotherapeutic treatment.
Discussion
Despite the development of combinatorial therapies, approximately 80%–85% of patients will experience recurrence and metastasis. During the second-line treatment, chemotherapeutic efficacy usually declines because of the emergence of drug resistance in cancer cells. Most chemotherapeutic drugs present multidrug resistance and decreased sensitivities in cells.9 There has been evidence suggesting that combining traditional Chinese medicine with chemotherapies enhanced the overall antitumor efficacy with some combinations having the potential to reverse cancer cell resistance to chemotherapy.10,11 Complex molecular mechanisms may be involved in the development of cisplatin resistance in cancer cells.12,13 Cisplatin-induced multidrug-resistant lung cancer cells have provided a representative model for characterizing these mechanisms. As a traditional Chinese medicine, alpinetin has demonstrated potent antiproliferative activity toward certain tumors. However, little is known of its utility in treating chemotherapy-resistant cancer cells.
The purpose of this study was to investigate the antitumor efficacy and potential mechanism of alpinetin in lung cancer cells, including A549, SK-MES-1, NCI-H292, and A549/CDDP cells. The results suggest that alpinetin inhibited the proliferation of A549, SK-MES-1, and NCI-H292 cells. Although alpinetin did not exhibit apparent growth-inhibitory effects in the multidrug resistant A549/CDDP cells, it restored the sensitivity of A549/CDDP cells toward the chemotherapy using CDDP by inhibiting the expression of drug resistance-related proteins. The flow cytometry analysis suggested that after 72 hours of treatment, alpinetin dose dependently increased the percentage of annexin V-positive and PI-negative cells, indicating that the growth-inhibitory effect of alpinetin is through the induction of cell apoptosis but not necrosis.
Cell apoptosis is an essential self-regulatory mechanism,14 which can be induced via two signaling processes: mitochondrial apoptotic pathway apoptosis and death receptor-mediated extrinsic apoptosis.15 Bcl-2 family members are important mediators of this extrinsic pathway and regulate cell apoptosis through activating caspase or cytochrome c.16,17 Caspase-3 is a crucial member of the caspase family of proteases and plays an important role during cell apoptosis.18 Caspase-8 usually occurs as an enzyme precursor and is activated during Fas-receptor- and TNFR-1-mediated apoptosis. Activated caspase-8 forms a dimer, which in turn activates the downstream effectors.19–23 Caspase-9 is an upstream regulator of the apoptotic pathway whose activation is regulated by phosphorylation.24–28 Following the mitochondrial release of cytochrome c, caspase-9 forms a complex with cytochrome c and Apaf-1 and becomes activated. Activated caspase-9 then activates the key enzyme caspase-3, which is the most important step for initiating cell apoptosis and elicits the subsequent apoptotic signals.29,30 Cytochrome c combines with the apoptotic factor Apaf-1, which activates the proapoptotic caspase-9 and initiates the caspase cascade reaction.31,32
In this study, alpinetin-treated lung cancer cells exhibited a dose-dependent increase in apoptotic rate. Apoptotic rates were observed at alpinetin potent concentrations of 200 μM compared with the control group. Caspase-3, -8, and -9 were significantly activated, suggesting the successful activation of caspase-dependent A549 cell apoptosis. Furthermore, we investigated the effect of alpinetin on the expression of the Bcl-2 protein family XIAP and cytochrome c in A549 cells. A Western blot analysis suggested that alpinetin dose dependently reduced the expression of the Bcl-2 subfamily proteins,33 including Bcl-2, Bcl-xL, and mitochondrial cytochrome c, in A549 cells. However, the Bax subfamily protein bax34 and the cytosolic cytochrome c exhibited increased expression in response to alpinetin in a dose-dependent manner. These data further suggest that alpinetin inhibits lung cancer proliferation by regulating the expression of XIAP in the Bcl-2 family, the release of cytochrome c, and the activation of caspase, thereby inducing cell apoptosis.
PI3K/Akt plays an important role in inhibiting cell apoptosis, possibly by regulating the activity of Bcl-2 family members. PI3K-dependent Akt activation can stimulate the phosphorylation of Bcl-2-associated death promoter. Bcl-2-associated death then dissociates from Bcl-2 or Bcl-xL and forms a complex with the antiapoptotic protein 14-3-3. The released Bcl-2 then exerts an antiapoptotic effect.35 In addition, the activation of the PI3K/Akt signaling pathway can increase the phosphorylation of Bax at Ser184, which renders Bax inactive and inhibits cell apoptosis.36 This study suggests that the PI3K/Akt signaling proteins were dose dependently downregulated by alpinetin with changes that are similar to the changes in Bcl-2, Bcl-xL, and XIAP expressions.
Moreover, drug resistance has become an inevitable issue for chemotherapeutic treatments with MPR1, MPR5, and P-gp playing important roles.37,38 The level of MPR1 has become an effective marker for the chemotherapeutic efficacy and overall survival rate of non-small-cell lung cancer.38 The expression of the drug resistance-related proteins MPR1, MPR5, and P-gp was determined by Western blot analysis. We found that the therapeutic doses of alpinetin reversed the drug resistance of A549/CDDP cells to CDDP by inhibiting the expression of MPR1, MPR5, and P-gp and restoring the sensitivity to CDDP, indicating that this effect is mediated by regulating the expression of drug resistance-related proteins.
Conclusion
In conclusion, this study demonstrated that alpinetin inhibited lung cancer cell proliferation by inducing the cell apoptotic response and reversed the drug resistance of A549/CDDP cells to CDDP.
Disclosure
The authors report no conflicts of interest in this work.
References
Ridge CA, McErlean AM, Ginsberg MS. Epidemiology of lung cancer. Semin Intervent Radiol. 2013;30:93–98. | ||
Chen W, Zhang S, Zeng H, Zheng R, Zou X, Zhao P. Report of cancer incidence and mortality in China. China Cancer. 2010;23:1–10. | ||
Warth A, Muley T, Meister M, Stenzinger A, Thomas M, Schirmacher P. The novel histologic International Association for the Study of Lung Cancer/American Thoracic Society/European Respiratory Society classification system of lung adenocarcinoma is a stage-independent predictor of survival. J Clin Oncol. 2012;30:1438–1446. | ||
Xin H, Kong Y, Jiang X, Wang K, Qin X, Miao Z. Multi-drug-resistant cells enriched from chronic myeloid leukemia cells by Doxorubicin possess tumor-initiating-cell properties. J Pharmacol Sci. 2013;20:299–304. | ||
Du J, Tang B, Wang J, Sui H, Jin X, Wang L. Antiproliferative effect of alpinetin in BxPC-3 pancreatic cancer cells. Int J Mol Med. 2012;29:607–612. | ||
Tang B, Du J, Wang J, Tan G, Gao Z, Wang Z. Alpinetin suppresses proliferation of human hepatoma cells by the activation of MKK7 and elevates sensitization to cis-diammined dichloridoplatium. Oncol Rep. 2012;27:1090–1096. | ||
Umehara K, Nemoto K, Kimijima K, Matsushita A, Terada E, Monthakantirat O. Estrogenic constituents of the heartwood of Dalbergia parviflora. Phytochemistry. 2008;69:546–552. | ||
Wang Z, Lu W, Li Y, Tang B. Alpinetin promotes Bax translocation, induces apoptosis through the mitochondrial pathway and arrests human gastric cancer cells at the G2/M phase. Mol Med Rep. 2013;7:915–920. | ||
Sun W, Lv C, Zhu T, Yang X, Wei S, Sun J. Ophiobolin-O reverses adriam;cin resistance via cell cycle arrest and apoptosis sensitization in adriamycin-resistant human breast carcinoma (MCF-7/ADR) cells. Mar Drugs. 2013;11:4570–4584. | ||
Fu Z, Han J, Zhang H. Effects of emodin on gene expression profile in small cell lung cancer NCI-H446 cell. Chin Med J. 2007;120:1710–1715. | ||
Clark AS, West K, Streicher S, Dennis PA. Constitutive and inducible Akt activity promotes resistance to chemotherapy, or tamoxifen in breast cancer cells. Mol Cancer Ther. 2002;1:707–717. | ||
Takano M, Kudo K, Goto T, Yamamoto K, Kita T, Kikuchi Y. Analyses by comparative genomic hybridization of genes relating with cisplatin-resistance in ovarian cancer. Hum Cell. 2001;14:267–271. | ||
Chen HY, Yu SL, Chen CH, Chang GC, Chen CY, Yuan A. A five-gene signature and clinical outcome in non-small-cell lung cancer. N Engl J Med. 2007;356:11–20. | ||
Meier P, Finch A, Evan G. Apoptosis in development. Nature. 2000;407:796–801. | ||
Arlt A, Müerköster SS, Schäfer H. Targeting apoptosis pathways in pancreatic cancer. Cancer Lett. 2013;332:346–358. | ||
Burlacu A. Regulation of apoptosis by bcl-2 family proteins. J Cell Mol Med. 2003;7:249–257. | ||
Kuwana T, Mackey MR, Perkins G, Ellisman MH, Latterich M, Schneiter R. Bid, Bax, and lipids cooperate to form supramolecular openings in the outer mitochondrial membrane. Cell. 2002;111:331–342. | ||
Shi Y. Mechanisms of caspase activation and inhibition during apoptosis. Mol Cell. 2002;9:459–470. | ||
Nicholson DW, Ali A, Thornberry NA, Vaillancourt JP, Ding CK, Gallant M. Identification and inhibition of the ICE/CED-3 protease necessary for mamlian apoptesis. Nature. 1995;376:37–43. | ||
Cepero E, King AM, Coffey LM, Perez RG, Boise LH. Caspase-9 and effector caspases have sequential and distinct effects on mitochondria. Oncogene. 2005;24:6354–6366. | ||
Gehring S, Rottmann S, Menkel AR, Mertsching J, Krippner-Heidenreich A, Lüscher B. Inhibition of proliferation and apoptosis by the transcriptional repressor Mad Repression of Fas induced caspase-8 activation. J Biol Chem. 2000;275:10413–10443. | ||
Scatena R, Bottoni P, Botta G, Martorana GE, Giardina B. The role of mitochondria in pharmacotoxicology: a reevaluation of an old, newly emerging topic. Am J Physiol Cell Physiol. 2007;293:12–21. | ||
Hacker G, Paschen SA. Therapeutic targets in the mitochondrial apoptotic pathway. Expert Opin Ther Targets. 2007;11:515–526. | ||
Yuan S, Yu X, Topf M, Ludtke SJ, Wang X, Akey CW. Structure of an apoptosome-procaspase-9 CARD complex. Structure. 2010;18:571–583. | ||
Malladi S, Challa-Malladi M, Fearnhead HO, Bratton SB. The Apaf-1 procaspase-9 apoptosome complex functions as a proteolytic-based molecular timer. EMBO J. 2009;8:1916–1925. | ||
Yamaguchi Y, Shinotsuka N, Nonomura K, Takemoto K, Kuida K, Yosida H. Live imaging of apoptosis in a novel transgenic mouse highlights its role in neural tube closure. J Cell Biol. 2011;195:1047–1060. | ||
Manns J, Daubrawa M, Driessen S, Paasch F, Hoffmann N. Löffler A: triggering of a novel intrinsic apoptosis pathway by the kinase inhibitor staurosporine: activation of caspase-9 in the absence of Apaf-1. FASEB J. 2011;25:3250–3261. | ||
Laussmann MA, Passante E, Düssmann H, Rauen JA, Würstle ML, Delgado ME. Proteasome inhibition can induce an autophagy-dependent apical activation of caspase-8. Cell Death Differ. 2011;18:1584–1597. | ||
Riedl SJ, Shi Y. Molecular mechanisms of caspase regulation during apoptosis. Nat Rev Mol Cell Biol. 2004;5:897–907. | ||
Chen M, Wang J. Initiator caspases in apoptosis signaling pathways. Apoptosis. 2002;7:313–319. | ||
Haebedem SL. Mitochondrial function in apoptotie neuronal cell death. Neurechem Res. 2004;29:521–530. | ||
Tian X, Zhang Z. Resveratrol promote permeability transifion pore opening mediated by Ca2+. Yao Xue Xue Bao. 2003;38:81–84. | ||
Sitailo LA, Jerome-Morais A, Denning MF. Mcl-1 functions as major epidermal survival protein required for proper keratinocyte differentiation. J Invest Dermatol. 2009;129:1351–1360. | ||
Theofilas P, Bedner P, Hüttmann K, Theis M, Steinhäuser C, Frank S. The proapoptotic Bcl-2 homologY domain 3-only protein Bim is not critiCal for acute excitotoxic cell death. J Neuropathol Exp Neurol. 2009;68:102–110. | ||
Henshall DC, Araki T, Schindler CK, Lan JQ, Tiekoter KL, Taki W. Activation of bcl-2 associated death protein and counter-response of Akt within cell populations during seizure-induced neuronal death. J Neurosci. 2002;22:8458–8465. | ||
Xin M, Deng X. Nicotine inactivation of the proapoptotic function of Bax through phosphorylation. J Biol Chem. 2005;280:10781. | ||
Hu T, To K, Wang L, Zhang L, Lu L, Shen J. Reversal of P-glycoprotein (P-gp) mediated multidrug resistance in colon cancer cells by cryptotanshinone and dihydrotanshinone of Salvia miltiorrhiza. Phytomedicine. 2014;21:1264–1272. | ||
Berger W, Setinek U, Hollaus P, Zidek T, Steiner E, Elbling L. Multidrug resistance markers P-glycoprotein, multidrug resistance protein 1, and lung resistance protein in non-small cell lung cancer: prognostic implications. J Cancer Res Clin Oncol. 2005;131:355–363. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.