Back to Journals » Psychology Research and Behavior Management » Volume 19
Adolescent Insomnia Disorder – Problematic Internet Use Nexus: A Convergent Developmental Trajectory – Bridging Neuroimaging Biomarkers, Molecular Susceptibility, and Environmental-Neurocognitive Cascades
Authors Zeng Z, Jiang J, Wang M, Huang Y, Maes M, Li J
Received 14 November 2025
Accepted for publication 26 February 2026
Published 4 March 2026 Volume 2026:19 581502
DOI https://doi.org/10.2147/PRBM.S581502
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Igor Elman
Zixuan Zeng,1 Jiajun Jiang,2 Min Wang,2 Yi Huang,2 Michael Maes,1,3 Jing Li1– 3
1Sichuan Provincial Center for Mental Health, Sichuan Provincial People`s Hospital, School of Medicine, University of Electronic Science and Technology of China, Chengdu, 610072, People’s Republic of China; 2Department of Psychiatry, The School of Psychiatry, North Sichuan Medical College, Nanchong, Sichuan, 637000, People’s Republic of China; 3Key Laboratory of Psychosomatic Medicine, Chinese Academy of Medical Sciences, Chengdu, 610072, People’s Republic of China
Correspondence: Michael Maes, Sichuan Provincial Center for Mental Health Sichuan Provincial People’s Hospital, School of Medicine, University of Electronic Science and Technology of China, Chengdu, 610072, People’s Republic of China, Email [email protected] Jing Li, Sichuan Provincial Center for Mental Health Sichuan Provincial People’s Hospital, School of Medicine, University of Electronic Science and Technology of China, Chengdu, 610072, People’s Republic of China, Email [email protected]
Abstract: Adolescent Insomnia Disorder (ID) and Problematic Internet Use (PIU) represent a pressing global public health challenge, frequently co-occurring and synergistically impacting mental and physical well-being. Despite their high comorbidity, a comprehensive integrative model elucidating their shared pathophysiology is lacking. This review is the first to propose a Convergent Developmental Trajectory Model, which posits that ID and PIU in adolescents are sustained by a self-reinforcing cycle driven by the dynamic interplay of three core components: (a) a shared biological vulnerability stemming from adolescent-specific neurodevelopment and molecular genetics; (b) common environmental triggers, such as academic stress and adverse childhood experiences; and (c) neurocognitive reinforcement through impaired inhibitory control and dysregulated reward processing. We synthesize converging evidence from neuroimaging, molecular studies, and environmental psychology to demonstrate how these components interact within the adolescent developmental context to perpetuate a vicious cycle. The model underscores that effective interventions must target these shared pathways, for instance, by developing transdiagnostic Cognitive Behavioral Therapy (CBT) protocols specifically designed for ID-PIU comorbidity. Future research should prioritize longitudinal neurodevelopmental studies and multi-omics profiling to enable early risk stratification and personalized approaches. Addressing this complex issue requires interdisciplinary collaboration to formulate precision therapies and evidence-based public health policies.
Keywords: insomnia disorder, problematic internet use, adolescent, comorbidity, developmental model
Introduction
Definitions and Diagnostic Criteria
According to the Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition (DSM-5), Insomnia Disorder (ID) is defined as dissatisfaction with sleep quantity or quality, characterized by difficulty initiating sleep, maintaining sleep, or early morning awakening with inability to return to sleep, despite adequate sleep opportunities. The sleep disturbance must cause clinically significant distress or impairment in daytime functioning, occur at least three nights per week, and persist for a minimum of three months.1 The World Health Organization (WHO) defines adolescents as individuals aged 10 to 19 years.
While the DSM-5 explicitly delineates Internet Gaming Disorder (IGD) as a condition marked by excessive and compulsive engagement in electronic games leading to significant impairment or distress over a 12-month period—manifesting in adverse outcomes such as academic decline, physical health issues, and social isolation1—the International Classification of Diseases, 11th Revision (ICD-11) employs the term Gaming Disorder (GD) to describe a behavioral pattern characterized by impaired control over gaming, where gaming increasingly takes precedence over other life activities and persists despite negative consequences.2 Notably, neither classification provides formal clinical diagnostic criteria for broader patterns of Problematic Internet Use (PIU). However, beyond IGD/GD, a substantial body of research indicates that other internet-related activities—such as excessive social networking, internet pornography consumption, or online shopping—can engender significant behavioral dysregulation and functional impairment. In such cases, these patterns may be conceptualized under the ICD-11 category of “Other Specified Disorders Due to Addictive Behaviors”.3 In the absence of formal diagnostic criteria, the research community has operationalized the construct of PIU to investigate these maladaptive behaviors. It is typically defined by a pattern of compulsive use of digital networks, marked by excessive preoccupation, loss of control, and cravings, which results in significant functional impairments and psychological distress. This construct is commonly measured using validated scales such as the Compulsive Internet Use Scale (CIUS),4 which reflects a shift away from older frameworks like Young’s Internet Addiction Test (IAT). This review focuses on the characteristics of PIU among adolescents, particularly as it relates to gaming, social networking, and online content.
Epidemiology and Public Health Burden
Insomnia is one of the prevalent neuropsychological issues during adolescence. Research has revealed that adolescence represents a critical phase for brain development and functional reorganization.5,6 Adolescent insomnia is associated with diminished health-related quality of life and elevated risks of somatic, interpersonal, and psychiatric problems.7,8 Variations in diagnostic criteria and assessment methodologies across studies have led to heterogeneous epidemiological findings on insomnia disorders in children and adolescents.
Adolescence is a period of heightened vulnerability for the onset of both sleep disorders and maladaptive digital behaviors. Insomnia Disorder (ID) is prevalent in this population, with estimates varying by diagnostic criteria but consistently pointing to a significant burden; for instance, a large Norwegian study found a prevalence of 18.5% using DSM-5 criteria,9 while a Chinese meta-analysis reported a 26% pooled prevalence of sleep disturbances.10 Concurrently, PIU has emerged as a global concern, with meta-analytic data suggesting a pooled prevalence of Internet Gaming Disorder, a core component of PIU, as high as 9.9% among adolescents and young adults.11 The co-occurrence of ID and PIU is frequent and pernicious, associated with diminished quality of life, academic decline, and exacerbated psychiatric symptoms.7,8,12 This confluence underscores the urgent need to move beyond studying these conditions in isolation and to understand the interconnected mechanisms that drive their comorbidity.
Historical Evolution and the Need for an Integrative Model
The investigation into ID and PIU has followed distinct historical pathways, necessitating a contemporary synthesis. Initial research phases (early 2000s) primarily characterized ID and PIU as independent entities. Seminal work established the epidemiology and diagnostic criteria of ID in adolescents,7,9,13 while parallel research pioneered the conceptualization of PIU, often adapting frameworks from substance addiction.4
A second phase (circa 2010s) witnessed growing clinical and epidemiological recognition of their co-occurrence,12 prompting initial theories about bidirectional relationships. Research began to suggest that internet engagement could displace sleep time and disrupt circadian rhythms,12–18 while sleep deprivation could impair executive function, increasing vulnerability to compulsive internet use.19,20 However, these explorations remained largely siloed within psychological or behavioral domains.
The current era is defined by technological advances enabling the interrogation of shared neurobiological and molecular substrates. Converging evidence from neuroimaging,21–24 genetics,25 and immunology26 has revealed common neural circuits (eg, DMN-SN-CEN dysregulation) and biological pathways (eg, dopaminergic dysregulation) underlying both disorders. This accumulating yet fragmented evidence creates a critical imperative: to move beyond describing comorbidity and toward explaining it through a unified framework that integrates these multi-level findings. This review answers that call by proposing the Convergent Developmental Trajectory Model, which synthesizes neurobiological, molecular, and environmental-neurocognitive evidence into a cohesive narrative of shared vulnerability and reciprocal reinforcement, specifically within the adolescent neurodevelopmental context.
Theoretical Frameworks Linking Both Conditions
Building upon this historical context, contemporary evidence robustly supports a bidirectional relationship between adolescent ID and PIU.12 Internet engagement (including social media, smartphone usage, and online gaming) profoundly impacts sleep quality in adolescents.14–18
In developing the theoretical underpinnings for the present model, we build upon the influential I-PACE (Interaction of Person-Affect-Cognition-Execution) model by Brand et al, which provides a comprehensive, process-based framework for understanding the development and maintenance of various internet-use disorders.27 The I-PACE model astutely conceptualizes how predisposing core characteristics (P) interact with affective and cognitive responses to trigger and perpetuate addictive behaviors, moderated by executive control (E).
The Convergent Developmental Trajectory Model proposed here does not seek to replace the foundational I-PACE framework. Instead, it offers a focused specification and extension tailored to the specific clinical context of adolescent ID-PIU comorbidity. This model extends the I-PACE framework by: (a) specifying the shared neurodevelopmental vulnerabilities, detailing the particular neurobiological and molecular substrates involved; (b) elaborating a specific, mutually reinforcing cycle that captures how sleep disruption and compulsive internet use become locked together during adolescence; and (c) integrating evidence across neuroimaging, genetics, and environmental psychology to construct an integrative, multi-level explanatory framework. By doing so, it systematically elucidates the mechanisms underlying their comorbidity. By concentrating on this specific comorbidity during the critical window of adolescence, our model aims to provide a more granular, mechanism-driven heuristic that can generate testable hypotheses and guide targeted, transdiagnostic interventions for this particular population.
Through a comprehensive review of literature linking adolescent ID and PIU, coupled with the convergence of neuroimaging and molecular mechanisms, we propose a potential vicious cycle model characterized by:
a) Biological predisposition: biological and neurodevelopmental vulnerabilities increase susceptibility to initial sleep dysregulation;
b) Environmental triggers: psychosocial stressors (eg, academic pressure, social isolation) drive maladaptive internet use as maladjusted emotional coping strategies;
c) Neurocognitive reinforcement: dysfunctional prefrontal-striatal circuitry and dopaminergic dysregulation impair inhibitory control, perpetuating both sleep disturbances and compulsive internet behaviors.
Building upon this perspective, the subsequent review will systematically examine the interplay between:
(i) Neurobiological convergence in neuroimaging – focusing on functional/structural brain alterations across insomnia and Problematic Internet Use phenotypes;
(ii) Molecular predisposition Convergence – focusing on the biological factor effects and predictive functions of both;
(iii) The impact of Environmental-Neurocognitive Cascades -focusing on the environmental t riggers and neurocognitive reinforcement in adolescent of both.
Scope and Synthesis of This Theoretical Review
This narrative theoretical review proposes a theoretical model—the Convergent Developmental Trajectory Model—to explain the comorbidity of Insomnia Disorder and Problematic Internet Use in adolescents. As such, it is not a systematic review but a narrative synthesis and theoretical integration of existing evidence. The model was constructed based on the authors’ expertise and a comprehensive examination of the literature in the fields of adolescent sleep medicine, behavioral addictions, developmental neuroscience, and molecular psychiatry. We prioritized seminal works and high-impact recent studies that illuminate the shared biological, environmental, and neurocognitive pathways between ID and PIU. Our aim is not to provide an exhaustive catalog of all related studies, but to synthesize a coherent and testable framework that can guide future hypothesis-driven research. Therefore, the evidence cited is illustrative of the proposed mechanisms rather than the product of a systematic search and screening process.
Neuroimaging Biomarkers
Neuroimaging Aberrations in Insomnia Disorder
The default mode network (DMN) represents a set of brain regions exhibiting high activity during rest, with core components encompassing the posterior cingulate cortex/precuneus (PCC/Precuneus), medial prefrontal cortex (mPFC), middle temporal lobe (MTL), angular gyrus (AG), parahippocampal gyrus (PHC), and temporoparietal junction (TPJ).19 Recent investigations have revealed significant associations between DMN functional abnormalities and the pathophysiology of ID. Accumulating evidence demonstrates that brain regions constituting the DMN fail to deactivate appropriately during rest in insomnia patients. Neuroimaging analyses of intra-DMN connectivity demonstrate enhanced functional coupling between core DMN regions (eg, PCC and mPFC) in individuals with insomnia. This hyperconnectivity may contribute to resting-state hyperarousal and ruminative thinking, clinically manifesting as sleep initiation difficulties and nocturnal awakenings.21
Concurrently, research indicates functional dysregulation between the DMN and other large-scale networks in insomnia. The salience network (SN), responsible for detecting environmentally salient stimuli, normally suppresses DMN activity during task engagement. In insomnia patients, disrupted competitive regulation between the DMN and SN impairs effective switching to external task-processing states, exacerbating excessive self-referential processing and heightened activity in primary limbic structures associated with anxiety, potentially mediating emotional dysregulation and hyperarousal during insomnia. Abnormal coordination is also observed between the DMN and central executive network (CEN), the latter being crucial for attentional focus and task execution. Reduced anticorrelation between the DMN and CEN in insomnia patients correlates with daytime attentional deficits and cognitive decline.22
Structural neuroimaging studies further identify gray matter (GM) volume alterations in DMN-related regions. Increased GM volumes in the right fusiform gyrus, primary auditory cortex (superior temporal gyrus), and supplementary motor area (BA6) may reflect compensatory enhancements in sensory processing and motor preparation under chronic hyperarousal states.28 Notably, reduced prefrontal cortex (PFC) GM volume in adolescent insomnia patients emerges as a key neuroimaging finding. As the PFC orchestrates executive functions, emotional regulation, and sleep-wake cycle modulation, its structural abnormalities may directly contribute to insomnia pathogenesis.29 Similar frontal volume reductions across age groups substantiate the PFC’s critical role in sleep maintenance.30
Adolescent-specific mechanisms involve hormonal influences on DMN maturation. Post-pubertal DMN development demonstrates nonlinear progression in segregation efficiency and network integration, particularly within attention and task-control networks.31 This neurodevelopmental pattern may partially explain adolescents’ heightened vulnerability to insomnia.
The SN, comprising the anterior insular cortex (AIC), anterior cingulate cortex (ACC), and subcortical nodes, exhibits hyperactivation and strengthened functional connectivity in insomnia patients, potentially reflecting hypervigilance to sleep-related threats.32,33 Right anterior insula (rAI), a key SN hub, shows abnormal effective connectivity with the PCC in insomnia populations.34
The amygdala (AMG), a core emotional processing region, demonstrates significant associations with affective dysregulation and hyperarousal states.35,36 Chronic insomnia patients exhibit localized amygdala atrophy, with specific amygdalar subregions correlating with both insomnia severity and comorbid anxiety symptoms.35 Notably, insomnia patients demonstrate significant alterations in functional connectivity between the AMG and anterior insula during resting wakefulness. Furthermore, resting-state functional connectivity between the amygdala and supramarginal gyrus shows significant correlations with EEG beta power during sleep.37
The striatum, a critical hub for reward and motivation regulation, exhibits reduced functional activity and aberrant connectivity in adolescent insomnia. Non-depressed adolescents with insomnia demonstrate significantly diminished activation in the ventral striatum (particularly the nucleus accumbens) compared to healthy controls during reward-based tasks.38 Li et al propose that chronic pain-induced insomnia may be mediated by ACC pyramidal neurons and their dopaminergic projections to the dorsal medial striatum.39
Neuroimaging Aberrations in Problematic Internet Use
DMN demonstrates both functional and anatomical connectivity abnormalities in PIU patients. Addictive behaviors disrupt DMN connectivity through multiple pathways, including altered interactions between the DMN and medial temporal lobe (MTL) as well as dorsomedial prefrontal cortex (dMPFC).40 Neuroimaging analyses reveal elevated local functional connectivity density (lFCD) in PIU patients within the right dorsolateral prefrontal cortex (DLPFC), left parahippocampal gyrus (PHG), cerebellum, bilateral mid-cingulate cortex (MCC), and superior temporal pole (STP), alongside reduced lFCD in the right inferior parietal lobule (IPL) and bilateral calcarine/lingual gyri.41 Investigations of internet gaming disorder (IGD) identify diminished dynamics in prefrontal-striatal and striatal-DMN neural circuits, potentially serving as biomarkers for predicting addiction severity.42
Structural alterations persist in DMN regions of PIU patients. Individuals with higher IGD severity exhibit increased gray matter (GM) volume in midline DMN components (ie, mPFC and precuneus). Progressive IGD severity correlates with volumetric expansion of DMN structures and weakened structural associations with visuospatial attention and motivational craving regions.43 Resting-state fMRI studies in adolescent PIU cohorts demonstrate: Reduced interhemispheric functional connectivity in the right frontoparietal network (FPN); Enhanced intrahemispheric connectivity in the left FPN; Decreased mPFC functional connectivity; Diminished functional connectivity between the salience network (SN) and DMN.23 Abnormal functional connectivity between SN/DMN and the left posterior superior temporal sulcus (pSTS) further suggests dysregulated integration of social brain networks in adolescent IGD.24
The anterior cingulate cortex (ACC), a critical SN node mediating conscious behavioral control, shows structural impairments in the dorsal ACC (dACC) of IGD patients.40 ACC GM volume negatively correlates with impulsivity in IGD populations, with concurrent GM reductions observed in the supplementary motor area (SMA), left ventrolateral prefrontal cortex (VLPFC), left inferior parietal lobule (IPL), and left anterior temporal lobe (ATL). Notably, VLPFC volume inversely associates with gaming duration.44
Amygdala functional-structural connectivity abnormalities characterize PIU pathophysiology: Attenuated negative functional connectivity between the amygdala and DLPFC; Enhanced negative connectivity with the precuneus/superior occipital gyrus (SOG); Reduced positive connectivity strength in amygdala-ACC pathways; Amplified positive connectivity in amygdala-thalamic circuits.45 Systematic reviews propose prolonged electronic screen exposure during adolescence may induce amygdala dysregulation.46
Striatal dysfunction emerges in IGD pathophysiology,47 with ventral striatal reward anticipation mechanisms potentially explaining symptoms of internet pornography addiction.48 The nucleus accumbens (NAc), a ventral striatal component of basal ganglia circuitry, plays pivotal roles in reward processing, motivation, addiction, and emotional regulation.49 PIU patients exhibit aberrant connectivity patterns involving: Left NAc-right cerebellar crus II; Right cerebellar lobule VI.46 IGD cohorts additionally demonstrate abnormal functional connectivity between the right hippocampus (rHip) and NAc.50
Neurobiological Convergence: Shared Functional/Structural Brain Alterations
Figure 1 illustrates the key aberrant brain regions in ID and PIU, with red indicating ID-specific abnormalities, blue representing PIU-specific alterations, and purple denoting overlapping regions.
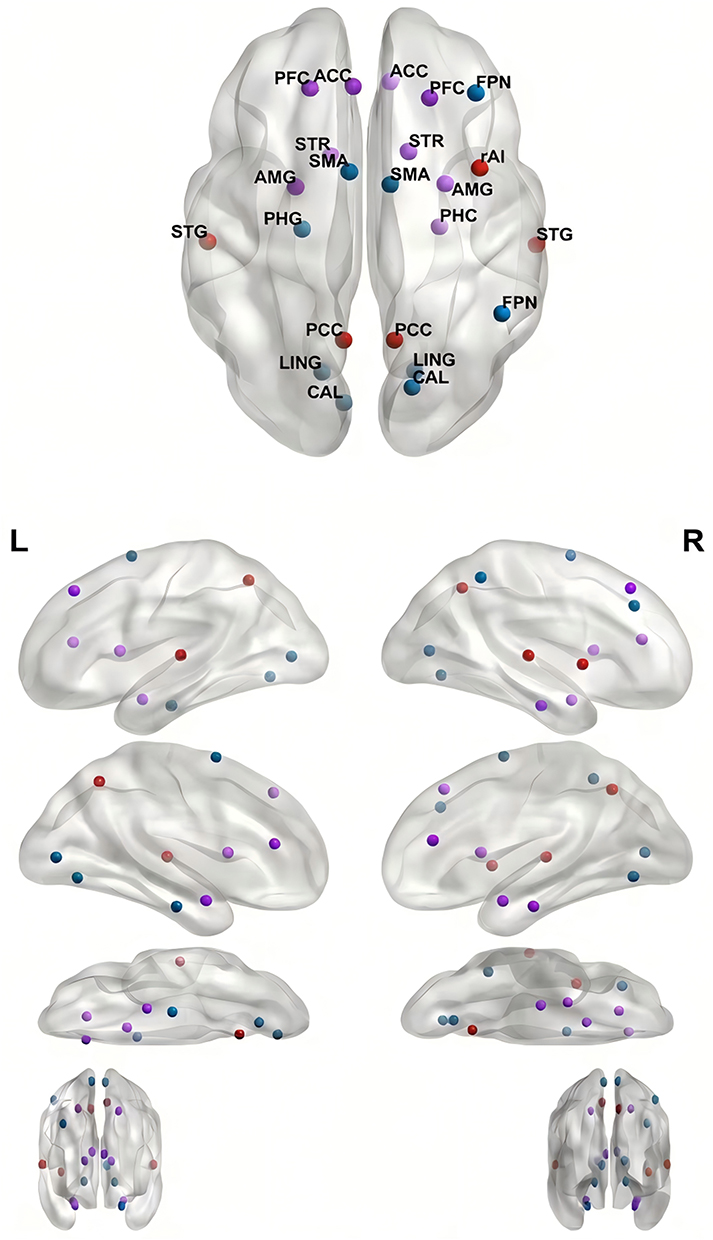 |
Figure 1 Spatial Distribution of Key Aberrant Brain Regions in Adolescent Insomnia Disorder and Internet Addiction. |
Emerging evidence reveals transdiagnostic convergence in DMN-SN-CEN dysfunction across ID and PIU, despite divergent clinical phenotypes. Critically, both disorders exhibit impaired SN-mediated network switching: ID manifests as DMN hyperconnectivity (eg, pathological PCC-mPFC coupling) with attenuated SN suppression,21,22 while PIU shows DMN-MTL/dmPFC dysregulation and compromised SN-DMN anticorrelation.23,40 This shared failure in dynamic network reconfiguration underlies persistent hyperarousal states in ID21,22 and may contribute to cue reactivity and loss of control in PIU.23,40 Thus, despite differing phenotypic presentations, both disorders exhibit a fundamental impairment in the balance between internally-focused (DMN) and externally-focused/executive (CEN) networks.22,42 Mechanistically, SN dysfunction mediates maladaptive allocation between internal (DMN) and external (CEN) cognition,22,23 establishing DMN-SN-CEN decoupling as a common neural substrate. Recent findings extend this convergence to PIU-related sleep disturbances, mediated by specific DMN subnetwork alterations (eg, rPrcu-lMTG connectivity impairment).51
Parallel dysregulation is also evident in the prefrontal-limbic system, which plays a pivotal role in cognition, emotional regulation, and behavioral control.52,53 ID and PIU demonstrate shared vulnerabilities within this circuitry. ID is characterized by structural PFC deficits, particularly reduced gray matter (GM) volume in adolescents,29 likely impairing top-down inhibition of limbic hyperactivity. Conversely, PIU manifests as ventrolateral PFC (VLPFC) atrophy correlating with impulsivity44 and dorsal anterior cingulate cortex (dACC) conflict monitoring deficits.54 Both disorders exhibit amygdala-related dysregulation: ID patients display amygdala-prefrontal decoupling (eg, weakened amygdala-DLPFC connectivity37) and localized amygdalar atrophy associated with anxiety,35 while PIU cohorts demonstrate altered amygdala-ACC connectivity45 and screen exposure-induced amygdalar hyperreactivity.46 These findings underscore a shared vulnerability in prefrontal-limbic circuitry, where combined PFC structural compromise (ID:;29 PIU:44) and ACC functional impairments (ID:;29 PIU:40) disrupt emotional homeostasis, exacerbating disorder-specific symptoms (eg, sleep-related anxiety in ID, impulsive internet use in PIU).
Reward processing represents another domain of convergent yet divergent dysregulation. The reward system, centered on the nucleus accumbens (NAc) and amygdala, governs motivation and reinforcement learning.55 Both disorders exhibit dysregulation with opposing phenotypes. Chronic sleep deprivation in ID induces ventral striatal hyporesponsiveness, exemplified by blunted NAc activation during reward tasks,38 reflecting diminished motivation for natural reinforcers. In contrast, PIU is characterized by ventral striatal hyperactivity, manifesting as enhanced NAc reactivity to gaming cues48 and aberrant NAc-cerebellar/hippocampal connectivity50,56 that drive compulsive internet-seeking behaviors. Despite phenotypic divergence, both conditions implicate dopaminergic dysregulation: ID involves ACC-driven dopaminergic projections to the dorsomedial striatum,39 while PIU pathophysiology correlates with NAc-centered reward prediction errors.48 This bidirectional striatal dysfunction—hypoactivity in ID versus hyperactivity in PIU—highlights a shared perturbation of the brain’s reward-aversion balance. Notably, adolescents with ID or PIU exhibit amplified striatal vulnerability due to immature prefrontal-striatal circuitry (ID:;38 PIU:56), suggesting asynchronous maturation of reward-control systems during puberty as a shared risk factor.31,46
These shared neural vulnerabilities are compounded by the unique neurodevelopmental context of adolescence. Previous studies have demonstrated that adolescents exhibit age-dependent increases in functional connectivity strength within the DMN, particularly between midline regions such as the PCC and mPFC. While these spatial patterns progressively approximate adult-like configurations—indicative of enhanced network integration capacity—mature DMN connectivity profiles remain underdeveloped.55 Furthermore, longitudinal PIU exposure induces structural brain alterations, as evidenced by significant correlations between internet addiction duration and gray matter volume reductions in the DLPFC, rACC, SMA, as well as white matter fractional anisotropy (FA) changes in the left posterior limb of the internal capsule (PLIC) among adolescent PIU populations.57
Adolescent vulnerability to both ID and PIU may be exacerbated by adverse childhood experiences (ACEs). Critically, ACEs correlate with aberrant spontaneous activity within hubs of the DMN (eg, heightened fractional amplitude of low-frequency fluctuations (fALFF) in the PCC) and accelerated leukocyte telomere shortening. Crucially, telomere shortening and DMN alterations are mechanistically linked: stress-induced proinflammatory states (eg, elevated IL-6) concurrently promote telomere attrition and aberrant DMN connectivity. Moreover, the quality of the family environment modulates the DMN-telomere relationship (as evidenced by the FES*LTL interaction at the mPFC), confirming their shared biological embedding. Thus, telomere shortening serves as a marker capturing the longitudinal sequelae of chronic stress beyond acute biomarkers (eg, cortisol, cytokines), while DMN dysfunction provides the neural substrate for sustained hyperarousal implicated in the pathogenesis of ID and PIU.58 We hypothesize that this ACE-related cascade (stress → inflammation → telomere attrition/DMN dysfunction) may represent a shared environmental-biological pathway that increases susceptibility to both ID and PIU during adolescence.58 These convergent DMN-centered neurobiological alterations are summarized in Table 1.
Molecular Susceptibility
Biological Abnormalities in Insomnia Disorder
Dopamine (DA) is a key modulator of learning and motivation.59 Early studies have demonstrated profound links between DA and sleep-wake regulation,60 suggesting that impaired DA function in humans may contribute to various sleep disorders, such as restless legs syndrome and REM sleep behavior disorder.61 Notably, DA-deficient mice exhibit a marked reduction in rapid eye movement (REM) sleep. Given that REM sleep typically follows deep non-rapid eye movement (NREM) sleep, this reduction may result from fragmented NREM sleep in DA-deficient mice.62 However, large-scale clinical studies investigating the high correlation between DA and ID remain scarce.
Gamma-aminobutyric acid (GABA), the primary inhibitory neurotransmitter in the central nervous system, promotes sleep through activation of GABA(A) receptors.63 Studies have shown that upregulating GABA expression in insomnia-prone mice alleviates sleep disturbances.64 Evidence indicates that GABAergic neuronal excitation induces NREM sleep while suppressing REM sleep, with REM suppression leading to sustained wakefulness.65 Current research reveals a significant positive correlation between ID and peripheral blood expression levels of GABA(A) receptor α1 and α2 subunit mRNAs in ID patients. This suggests that GABA(A) receptors, particularly α1-GABA(A) and/or α2-GABA(A) subtypes, may play critical roles in the pathophysiology of insomnia.66 Mainstream pharmacological treatments for insomnia, such as benzodiazepines and non-benzodiazepine drugs, exert sedative, anxiolytic, muscle relaxant, and hypnotic effects by targeting GABA receptor sites.67
The orexin system is hypothesized to be a key stabilizer of wakefulness.68 Previous studies found that aged mice exhibit a decline in orexin neuron numbers with age, while remaining neurons become hyperexcitable, leading to reduced arousal thresholds during sleep and increased sleep fragmentation. This may reflect age-related reductions in sleep demand.69 Orexin-targeting medications have been developed to improve insomnia, with evidence supporting their efficacy.70
Melatonin (N-acetyl-5-methoxytryptamine), primarily synthesized and secreted nocturnally by the pine pineal gland under normal environmental conditions, induces drowsiness and helps maintain circadian rhythms aligned with the light-dark cycle.71,72 A meta-analysis demonstrated that melatonin significantly increases total sleep time (with robust evidence), shortens sleep onset latency (though with substantial heterogeneity across studies), and exhibits clearer efficacy in children and adolescents with insomnia—potentially due to physiological melatonin deficiency (with robust evidence). Limited effects in adults may stem from reduced melatonin receptor sensitivity or insufficient dosing.73 Despite its close association with insomnia, some studies argue that melatonin lacks clinical value in managing psychophysiological insomnia.74
Current research identifies systemic inflammatory markers, including elevated C-reactive protein (CRP) and interleukin-6 (IL-6) levels, as significantly correlated with insomnia, with IL-6 showing stronger effects.26 These findings suggest that inflammatory cytokines may serve as predictive biomarkers for insomnia. Additionally, studies on gut microbiota and insomnia propose that bidirectional gut-brain interactions may trigger complex inflammatory mechanisms that exacerbate or initiate sleep disturbances.75
Genetic alterations are also tightly linked to insomnia. A large-scale genome-wide association study (GWAS) identified MEIS1 (chromosome 2), BTBD9 (chromosome 6), and MTNR1B (melatonin receptor) as key insomnia risk genes, alongside core circadian regulators such as CLOCK/BMAL1 and period genes PER2/PER3.25 Recent research has further uncovered more novel genes associated with sleep-related traits.76 Sleep-related genetic factors implicated in circadian rhythm regulation and insomnia vulnerability are summarized in Table 2.
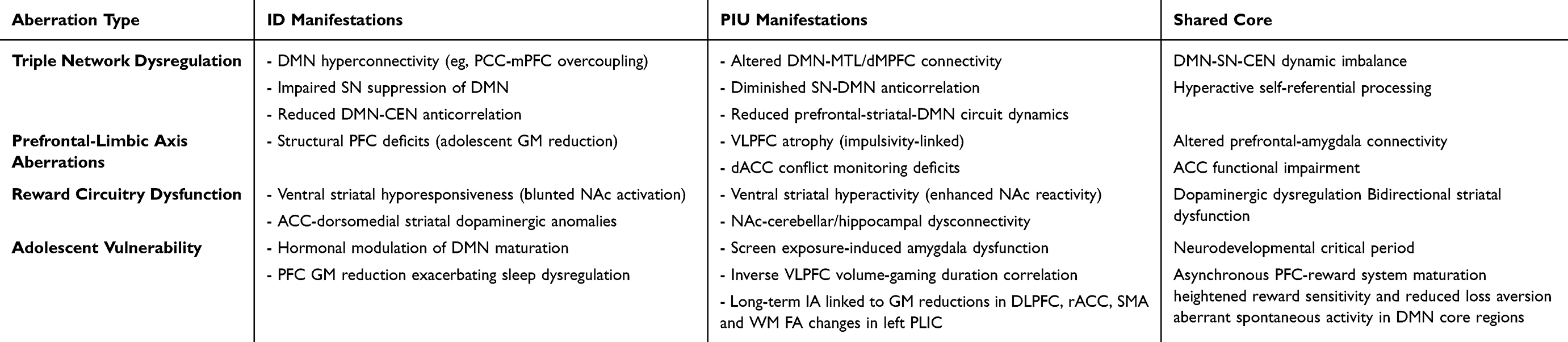 |
Table 1 Comparative & Convergent Neuroimaging Abnormalities in Adolescent Insomnia Disorder and Problematic Internet Use |
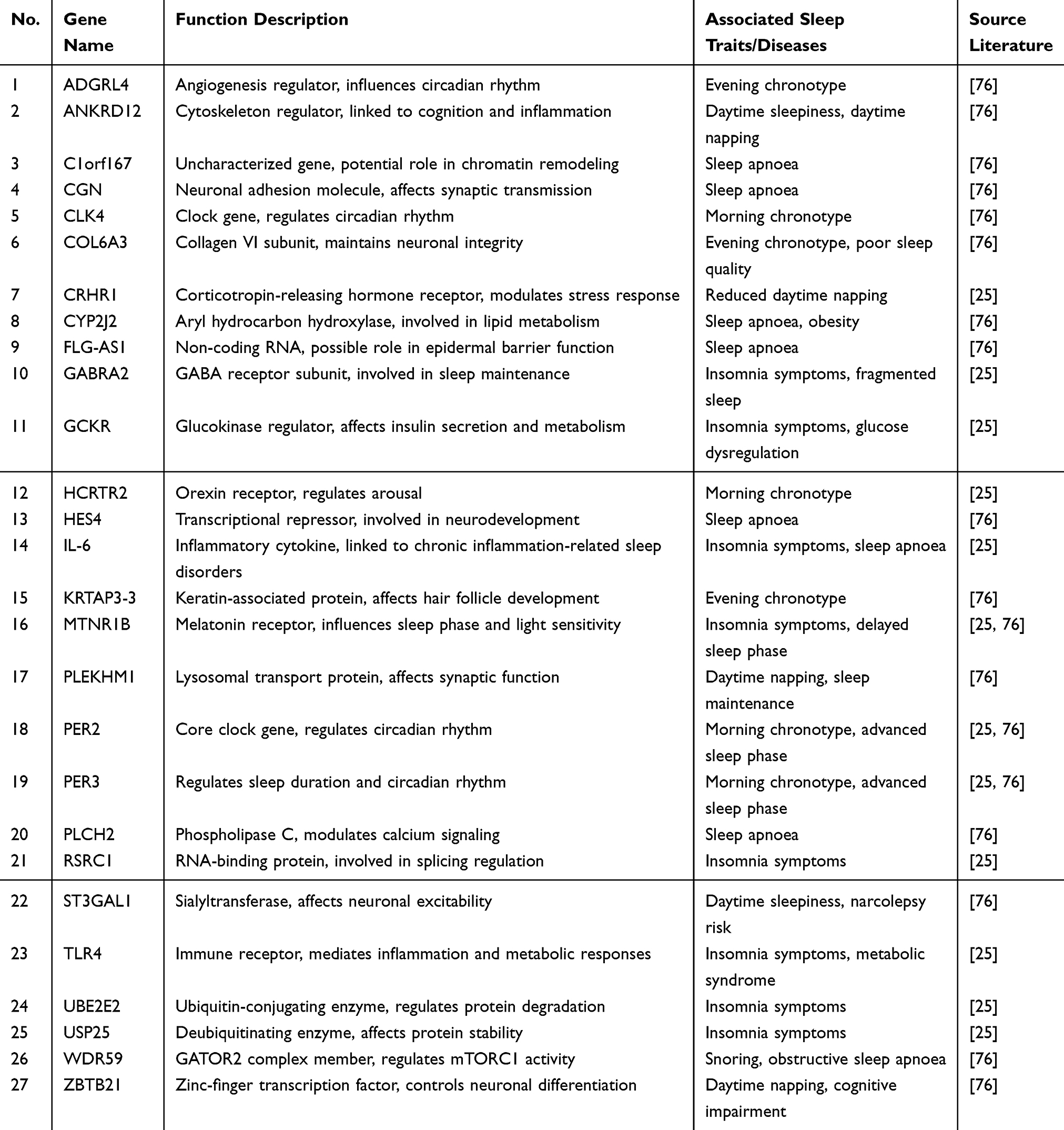 |
Table 2 Integrated List of Sleep-Related Genes |
 |
Table 3 Molecular Susceptibility Comparison Between Insomnia Disorder and Problematic Internet Use |
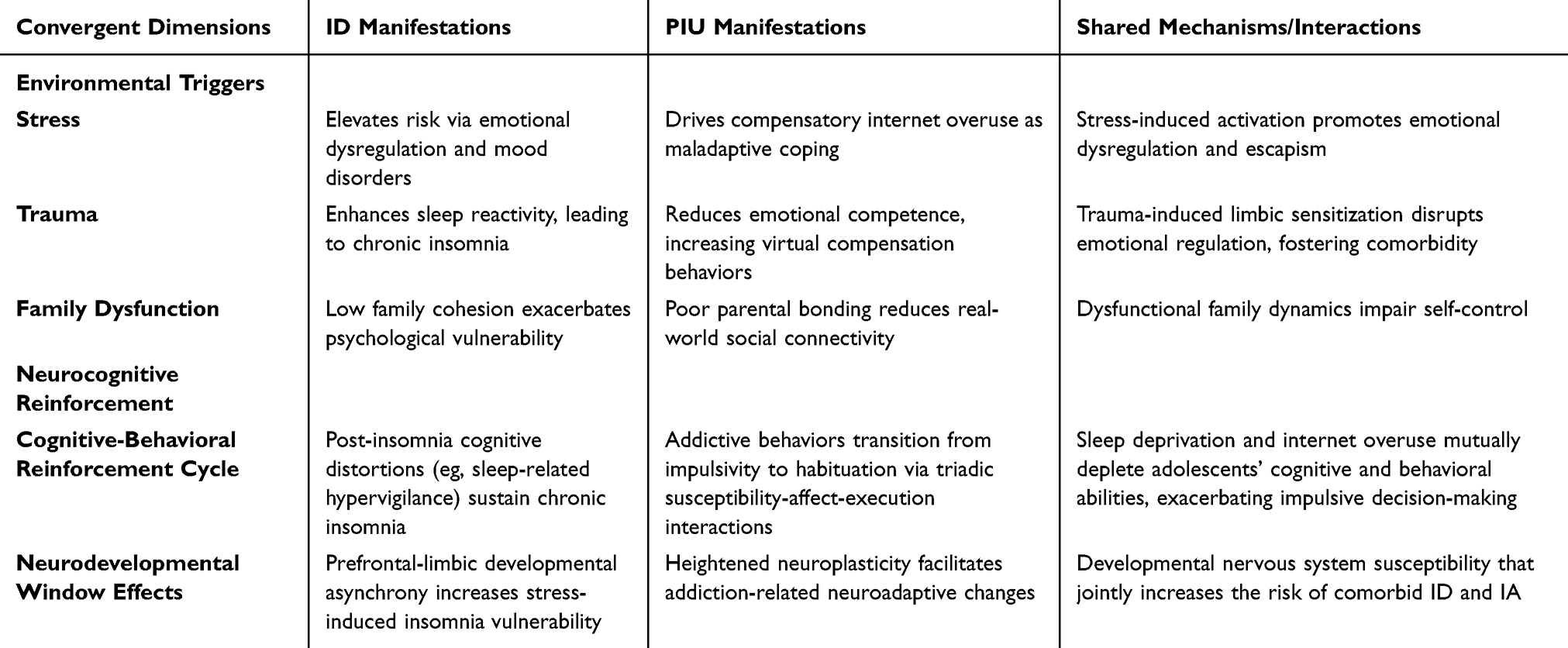 |
Table 4 Convergence of ID and PIU Environmental Triggers and Neurocognitive Reinforcement Mechanisms |
Biological Abnormalities in Problematic Internet Use
Dopamine (DA) is intricately linked to addiction. Nearly all addictive substances (eg, cocaine, alcohol, nicotine) elevate extracellular DA levels by activating the mesolimbic dopamine system, thereby reinforcing reward-seeking behaviors.77 Concurrently, evidence supports a strong association between downregulation of dopamine receptors and addiction susceptibility.78 Recent studies report that individuals with internet gaming disorder (IGD) exhibit significantly reduced striatal dopamine D2 receptor (D2R) density, which inversely correlates with addiction severity. Furthermore, post-gaming-task reductions in striatal dopamine transporter (DAT) levels in IGD patients suggest dysregulated reward circuitry.79 A study on adolescent PIU also identified elevated plasma DA levels in some patients.80 These findings underscore the critical role of DA in PIU pathophysiology.
Serotonin (5-HT) is indispensable for motor, cognitive, reward, and affective functions, with its primordial role hypothesized to counteract hyperactivation of neuromodulators such as norepinephrine and dopamine.81,82 Emerging research highlights its involvement in addictive behaviors.83,84 For instance, the 5-HTTLPR genotype elevates PIU risk by modulating serotonergic system activity.85 However, direct causal evidence linking 5-HT to PIU remains elusive.
In PIU, orexin A levels may rise to hyperactivate reward circuits.86 Additionally, IGD is associated with reduced peripheral glutamate levels, as aberrant glutamate transmission may impair learning- and memory-related functions, promoting compulsive gaming behaviors. This dysregulation may indirectly modulate dopaminergic reward pathways to drive PIU development.87 Excessive screen exposure further disrupts circadian rhythms by suppressing melatonin secretion.88
While genome-wide association studies (GWAS) on PIU are scarce, several candidate genes have been implicated. Polymorphisms in HTR2A (eg, rs6313 T allele [T102C]) are linked to PIU risk, potentially via shared mechanisms with substance use disorders, depression, and impaired cognitive control.89 In young males, NTRK3 gene polymorphisms increase IA susceptibility through neurodevelopmental impacts on reward processing.90 ANKK1 and DRD2 indirectly contribute to problematic internet use by altering hormone levels (eg, prolactin, 5-HT, DA, estradiol).90 Individuals carrying the MAOA TT genotype (rs1137070) exhibit substantially higher IGD risk than C allele carriers.91 Epigenetic mechanisms also play a role: methylation patterns in DAT1’s 5’-UTR and 3’-UTR regions influence PIU risk by regulating dopamine signaling, impulsivity, and attachment behaviors.92 The OXTR rs53576 AA genotype exacerbates IGD susceptibility in adolescents exposed to peer bullying.93 The T variant (CC genotype) of CHRNA4 rs1044396 significantly elevates PIU incidence.94 DRD2 rs6277 CT/TT genotypes may increase problematic video game use (PVGU) risk by reducing striatal D2 receptor density/affinity, impairing prefrontal cortex function, and exacerbating impulsivity and social deficits.95 Finally, the NTRK3 rs2229910 SNP (a synonymous mutation) correlates with reduced IGD susceptibility, possibly by altering mRNA splicing/stability to modulate NTRK3 expression, myelination, and dopaminergic signaling.96
Molecular Predisposition Convergence of Insomnia Disorder and Problematic Internet Use
Dopamine (DA) serves as a pivotal mediator in both ID and PIU. In ID, impaired DA function is linked to sleep-wake cycle disruptions and fragmented NREM-REM transition,57–60 while in PIU, DA hyperactivation via the mesolimbic pathway reinforces addictive reward-seeking behaviors.77–80 Notably, striatal D2 receptor (D2R) downregulation and dopamine transporter (DAT) dysfunction are observed in both conditions, suggesting overlapping dopaminergic deficits in reward processing and arousal modulation.
The orexin system stabilizes wakefulness in ID but is paradoxically hyperactivated in PIU to drive reward-seeking behaviors.68–70,86 Additionally, both disorders exhibit circadian rhythm disturbances: ID is linked to melatonin deficiency and delayed secretion,71–74 while PIU-related screen exposure suppresses melatonin, exacerbating sleep-phase delays.88 These findings suggest that orexin and melatonin dysregulation may serve as a shared biological substrate for sleep and addiction pathologies.
In addition, elevated levels of IL-6 and CRP are associated with chronic insomnia, suggesting that inflammatory markers could serve as biomarkers in ID patients.26 This also implies potential pathophysiological connections between ID and systemic inflammation. Notably, PIU patients exhibit gastrointestinal inflammatory responses, and gut-brain axis interactions may activate inflammatory pathways associated with both sleep disturbances and addictive behaviors.75,87 However, conclusive evidence for this hypothesis remains lacking, necessitating further investigation. The proposed link between gut-brain axis inflammation and both ID and PIU remains speculative and illustrates the kind of cross-disciplinary hypothesis generated by our integrative model, warranting future investigation.
Genetic evidence points to overlapping risk. Circadian genes (MEIS1, MTNR1B) are risk factors for ID25 and may influence PIU susceptibility via downstream effects on sleep and dopamine.88,95 Polymorphisms in dopaminergic (DRD2) and serotonergic (HTR2A, SLC6A4) genes are associated with traits (eg, impulsivity, reward sensitivity) central to both disorders.85,89,90,95
During adolescence, significant biological changes occur. Adolescents experience a phase delay in circadian rhythms, primarily driven by physiological adjustments in the circadian system, including delayed melatonin secretion onset, which shifts sleep-wake cycles toward later bedtimes.97 Compounding this, studies show that adolescents exhibit significantly later nocturnal sleep onset compared to childhood, while morning wake times are forcibly advanced due to school schedules, leading to cumulative sleep debt.98 This mismatch between sleep demand and actual sleep duration may predispose adolescents to insomnia. The hypothalamic-pituitary-adrenal (HPA) axis influences sleep through multiple pathways.99 Adolescence is a period of heightened stress sensitivity, during which chronic stress suppresses the growth hormone/insulin-like growth factor-1 (GH/IGF-1) axis, thyroid axis, and gonadal axis,100 reflecting the fragility of HPA axis stability in adolescents77 and its subsequent impact on sleep.
The adolescent developmental period amplifies these molecular vulnerabilities. This critical window is characterized by a confluence of DA system remodeling, circadian phase delay, and heightened HPA axis sensitivity.97–102 It is the synergistic interaction of these normative adolescent changes with the aforementioned genetic and molecular susceptibilities that likely creates a perfect storm, significantly elevating the risk for the co-emergence of ID and PIU.In this context, Collectively, these alterations across neurotransmitter, hormonal, inflammatory, and genetic systems suggest partially overlapping molecular vulnerability profiles for ID and PIU, as summarized in Table 3.
Environmental-Neurocognitive Cascades
For adolescents, entering puberty signifies not only a transition into a new developmental stage characterized by physical growth, but also the onset of a distinct phase of cognitive and psychological transformation. During this period, although individuals demonstrate marked improvements in logical reasoning, working memory, and planning capacities, their actual behaviors are frequently driven by emotional and social influences even when they cognitively recognize associated risks.103 Concurrently, research has demonstrated the profound impact of familial and peer relationships on adolescent development.104 Furthermore, this population exhibits particular susceptibility to various mental disorders.105 Regarding ID and PIU, substantial evidence highlights the contributory roles of social, psychological, and comorbid factors. These determinants can be systematically categorized into two principal dimensions: Environmental Triggers and Neurocognitive Reinforcement factors.
Environmental Triggers in Adolescence
Adolescents are more susceptible to insomnia due to various socio-environmental factors. Foremost among these is stress, which may be strongly associated with ID in adolescents.8 Studies indicate that 27% of adolescents experience high stress levels, with school being one of the most common stressors. School-related stress emerges as a particularly prominent risk factor among adolescents with insomnia, potentially contributing to ID by influencing emotional states, dietary behaviors, and increasing susceptibility to mood disorders such as anxiety and depression.106–109 Adverse childhood experiences also constitute significant contributors to ID. Childhood traumatic events may increase vulnerability to ID, with effects persisting for decades into adulthood.110 Repeated emotional and physical abuse during childhood may predispose individuals to later insomnia by enhancing sleep reactivity.111 Moreover, such maltreatment may persist as an ongoing stressor, further exacerbating sleep disturbances.112 Concurrently, studies have identified potential positive correlations between adolescent insomnia and high caffeine intake as well as electronic device usage.113,114 Prolonged exposure to electronic screens and outdoor artificial lighting extends adolescents’ light exposure duration, which can delay both bedtime and waking times, potentially leading to ID and other psychiatric disorders.115–119
Similarly, stress may predispose adolescents to PIU, with studies demonstrating a positive correlation between stress and PIU.120 Childhood trauma is significantly associated with adolescent PIU, as traumatic experiences may reduce familial warmth, diminish self-esteem and social competence, and amplify negative emotions (eg, depression, anxiety, loneliness), thereby driving adolescents to seek emotional compensation through virtual environments.121,122 Family and school factors also critically influence PIU susceptibility. Prior research identifies reduced maternal care, increased maternal control, heightened paternal autonomy, and elevated parental rejection as risk factors for adolescent PIU.123 Adolescents exhibiting lower levels of family cohesion, poorer teacher-student relationships, weaker peer connections, negative academic attitudes, and reduced school engagement demonstrate greater susceptibility to PIU.124,125 Genetics further indirectly modulate adolescents` environmental susceptibility to PIU. For instance, interactions between the 5-HTTLPR genotype and environmental factors (eg, family functioning) suggest serotonin’s potential role in PIU development.85 The DAT1`s 5’-UTR and 3’-UTR regions may mediate impulsive behaviors that heighten PIU vulnerability,92 while the OXTR rs53576 AA genotype reveals associations between bullying exposure and adolescent IGD.93
Mental health issues also contribute to adolescent susceptibility to both ID and PIU. Psychological well-being serves as a critical foundation for healthy sleep,126 and mental health factors similarly play a significant role in adolescent insomnia. Psychological conditions such as depression and anxiety exhibit positive correlations with insomnia.127 Notably, childhood maltreatment may impair individuals` ability to identify and express emotions, heightening attention to external stimuli and fostering the development of alexithymia, which in turn promotes ID through mental health pathways.128 Divergent parenting styles may further shape personality traits, generating varied psychological vulnerabilities that predispose individuals to insomnia during development.129 Additionally, adverse psychosocial experiences, including racial discrimination and non-racial forms of discrimination (eg, based on gender, age, and height/weight), have been linked to ID occurrence in adolescents.130
The interplay between psychiatric comorbidities and sleep disturbances reveals critical risk patterns for ID. Other psychiatric disorders (eg, mood disorders) constitute significant risk factors for ID, primarily including depressive disorders,131 anxiety disorders,9,132 and attention-deficit disorders.133 Studies suggest a bidirectional relationship between adolescent ID and depressive disorders, where sleep disruptions may intensify ruminative thinking patterns, perpetuating further sleep disturbances and potentially evolving into depression over time.107,113 Adolescents’heightened vulnerability to anxiety symptoms134 further compounds this dynamic, as anxiety disorders may exacerbate ID symptoms. Concurrently, insomnia amplifies the burden of anxiety-related hyperarousal and social dysfunction, creating a self-reinforcing cycle.109,135,136
In parallel to these psychiatric comorbidities in ID, research underscores distinct psychosocial correlates of PIU. Adolescent PIU demonstrates significant associations with diminished well-being, reduced self-esteem, and elevated impulsivity and egocentrism.137 Emotional and behavioral disturbances indirectly shape internet use patterns through academic underperformance, establishing a cascading “psychological distress → academic difficulties → internet dependency” pathway.138 This mechanism reinforces the bidirectional interplay between compulsive internet use and mental health deterioration.139 Social anxiety emerges as a salient risk factor for PIU, with maladaptive social behaviors exacerbating addictive internet engagement.140 Notably, self-esteem operates as a protective buffer, with higher self-esteem correlating markedly with reduced PIU susceptibility.120
Emerging evidence highlights reciprocal environmental triggers between ID and PIU in adolescents. Shared socio-environmental risk factors, such as chronic stress and childhood trauma, may concurrently predispose individuals to both conditions through shared pathways. For instance, school-related stress not only elevates ID risk via emotional dysregulation106–109 but also drives compensatory internet overuse as a maladaptive coping mechanism.120 Similarly, childhood maltreatment demonstrates dual impacts—enhancing sleep reactivity to perpetuate ID111,112 while simultaneously reducing emotional competence, thereby increasing PIU susceptibility through virtual compensation behaviors.121,122
Environmental mediators further reinforce this interconnection. Prolonged electronic device exposure, while directly contributing to ID via circadian disruption,113–119 simultaneously facilitates PIU development through habitual internet engagement. Evidence demonstrates that PIU-related behaviors such as pre-sleep social media engagement12 and cyberbullying18 show strong associations with adolescent ID. Family dynamics exhibit bidirectional influences: poor parental bonding and low family cohesion amplify ID risk through psychological vulnerabilities128,129 while concurrently fostering PIU via reduced real-world social connectivity.123–125 Notably, mental health comorbidities create synergistic effects—anxiety and depressive symptoms not only exacerbate ID through hyperarousal mechanisms9,132,135 but also promote PIU via the “psychological distress → internet dependency” pathway.138,139 Crucially, these environmental triggers may operate through shared neurobiological substrates, as genetic polymorphisms influencing serotonin and dopamine systems modulate susceptibility to both sleep disturbances85 and impulsive internet use.92 Emerging research further reveals that sleep deprivation exacerbates addicts’ cravings and emotional instability through elevated stress hormones (eg, cortisol), while impairing decision-making capacity, thereby heightening addiction-related risks.141 This complex interplay underscores the necessity for integrated interventions addressing shared environmental determinants in adolescent ID and PIU comorbidity.
Neurocognitive Reinforcement in Adolescence
Cognitive-behavioral factors profoundly influence adolescent susceptibility to both ID and PIU. Adolescence represents a critical neurodevelopmental window, characterized by dynamic brain maturation that mediates both adaptive functioning and psychopathology. During this phase, gene-environment interactions may facilitate rapid cognitive growth. However, the prefrontal cortex’s inhibitory control functions, which undergo accelerated development in early adolescence, exhibit marked interindividual variability. In contexts of prefrontal functional immaturity, motivational efforts may become dysregulated, contributing to impulsive control deficits in some adolescents. Concurrently, heightened midbrain dopaminergic system activity during adolescence increases reward sensitivity, driving risk-taking behaviors. These neurocognitive dynamics render adolescents particularly vulnerable to ID and PIU due to cognitive-behavioral predispositions.142–144
The persistence of ID and PIU further perpetuates cognitive impairments in adolescents. Post-insomnia cognitive distortions related to sleep (eg, maladaptive beliefs, unrealistic expectations, attentional biases) play a pivotal role in sustaining chronic insomnia.145 Addiction research proposes a triadic interaction mechanism involving susceptibility, affective reactivity, and executive functioning, wherein addictive behaviors transition from impulsive actions to habitual patterns.27 The pursuit of reward fulfillment and hedonic experiences in addiction may reinforce behavioral persistence,146 a pattern evident in adolescent PIU.147 Adolescents’neurodevelopmental traits predispose them to such neurocognitive reinforcement processes,148 wherein addictive behaviors and sleep disturbances mutually exacerbate cognitive dysregulation.Together, these environmental stressors and neurocognitive reinforcement processes form a self-perpetuating cycle that sustains and amplifies the comorbidity between ID and PIU during adolescence, as summarized in Table 4.
Discussion
The intersection of adolescent ID and PIU represents a critical public health challenge, driven by overlapping neurobiological, molecular, and environmental vulnerabilities. Previous studies have identified significantly shortened sleep duration among individuals with excessive internet engagement.149 Due to adolescents’ unique physiological and psychological developmental factors,19,20,55,57,58,97–99,101–105 ID and PIU exhibit more pronounced convergent pathological alterations and shared risk factors in this population. This review synthesizes evidence supporting the bidirectional relationship between ID and PIU and proposes a vicious cycle model rooted in biological susceptibility, environmental triggers, and neurocognitive reinforcement. Figure 2 illustrates the mechanistic diagram of this model.
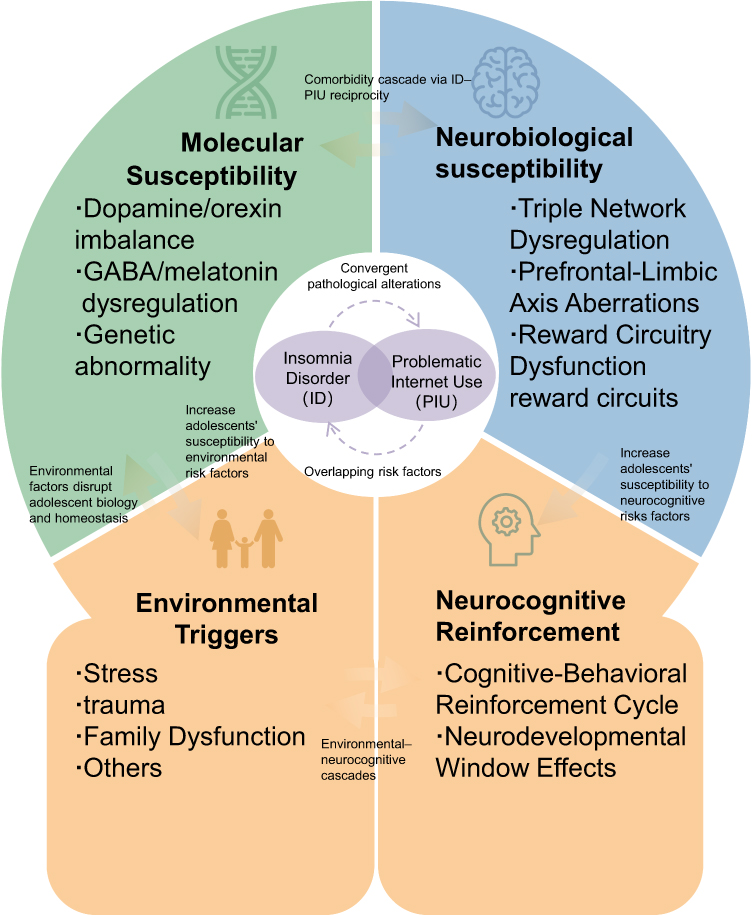 |
Figure 2 The Vicious Cycle Model. |
The proposed model posits that ID and PIU co-emerge through three interconnected pathways: (a) biological susceptibility (molecular and neurobiological vulnerabilities), (b) environmental triggers, and (c) neurocognitive reinforcement. The shared biological susceptibility between ID and PIU may synergistically exacerbate disease progression and persistence, potentially creating a pathological positive feedback loop. Adolescents, situated in a unique developmental context, lack fully adaptive coping mechanisms for stressors, trauma, and familial environments,150 rendering them more vulnerable to disruptions in biological homeostasis triggered by these environmental factors.151 Pathological activation of biological mediators (eg, dopamine [DA], GABA) or genetic predispositions59–62,68–70,87,95 further amplifies adolescents’ susceptibility to environmental risks. Although adolescents’ brain development progresses toward structural and functional maturity, their reward circuitry and prefrontal cortex remain insufficiently developed to meet cognitive demands,19,20,29,44 creating a neurodevelopmental window of vulnerability for ID and PIU. Interactions between environmental triggers and neurocognitive reinforcement form environmental-neurocognitive cascades, wherein adolescents’ insufficient sleep literacy152 and limited intervention strategies for PIU153 perpetuate the ID-PIU vicious cycle. Thus, interventions targeting adolescent ID and PIU must prioritize shared pathways to disrupt this co-occurring maladaptive loop.
Above all, this review synthesizes converging evidence to propose a Convergent Developmental Trajectory Model for adolescent ID-PIU comorbidity. Our integration of neuroimaging, molecular, and environmental-neurocognitive findings suggeststhat ID and PIU may not be fully independent conditions but are potentiallylinked through shared mechanisms. These include common neurobiological vulnerabilities (eg, alterations in DMN-SN-CEN dynamics, prefrontal-limbic-striatal circuitry), overlapping molecular susceptibilities (eg, involvement of dopaminergic and circadian rhythm genes), and reinforcing environmental-neurocognitive cascades. The adolescent neurodevelopmental period is hypothesizedto be a critical window where these factors interact, possiblycreating a self-perpetuating cycle that poses significant challenges for intervention.
Implications for Future Research
The proposed model, while synthetic, highlights several promising and prioritized avenues for future research to move from correlation toward causation and personalized approaches:
Longitudinal Neurodevelopmental Tracking: There is a pressing needfor cohort studies that integrate multi-modal data (eg, neuroimaging, digital phenotyping, polysomnography, molecular assays) to track the temporal dynamics and progression of ID-PIU comorbidity, which could help elucidate causal pathways.
Multi-Omics Integration: Research could benefit frommoving beyond candidate gene approaches to integrate genomics, proteomics, and metabolomics data. This mayhelp identify biosignatures for early risk stratification and uncover novel molecular targets.
Cross-Cultural Studies: It would be valuableto investigate whether the proposed mechanisms are universal or vary across cultures, particularly in regions with high rates of PIU. Such research is essentialfor developing globally effective interventions.
Experimental Mechanistic Trials: Proof-of-concept trials using neuromodulation (eg, TMS targeting prefrontal or cingulate regions) mighthelp test the model’s predictions by examining whether modulating specific neural circuits concurrently affects both sleep and internet use behaviors.
Testable Predictions Derived from the Model: To guide empirical validation, future studies could test specific predictions, such as: (a) Adolescents with a specific polygenic risk score combining circadian (eg, MTNR1B) and dopaminergic (eg, DRD2) gene variants will show a higher incidence of comorbid ID-PIU onset than those with risk for either disorder alone; (b) Baseline DMN-SN decoupling measured via fMRI at age 12–13 will predict the trajectory of both insomnia severity and compulsive internet use over a 3-year follow-up, even after controlling for baseline symptoms.
Potential Clinical and Public Health Implications
The accumulated evidence points to the potential utility of a paradigm shift in approaching adolescent sleep and digital health, perhapsmoving from isolated treatments to more integrated, transdiagnostic strategies.
Exploring Integrated Interventions: The development of transdiagnostic CBT protocols that target shared mechanisms (eg, metacognitions, reward processing) appears warranted. Digital platforms could be exploredas a scalable medium for delivering such interventions.
Informing School-Based Policies: School systems represent a logical venuefor intervention. Evidence suggeststhat circadian-aligned school start times mayimprove sleep duration, while sleep and digital literacy education couldempower self-regulation.
Guiding Diagnostic and Clinical Guidelines: The findings provide a scientific basisfor considering formal criteria for ID-PIU comorbidity in future diagnostic manuals, which mightstimulate the development of evidence-based assessment and treatment guidelines.
Informing Public Health Guidelines: Healthcare policymakers may considerformulating Adolescent Digital Health Guidelines that encourage features like evening screen curfews and discourage the use of addictive design elements in platforms targeting youth.
Limitations and Future Directions
The proposed Convergent Developmental Trajectory Model is a novel, integrative and heuristic framework for understanding the complex comorbidity between ID and PIU in adolescents. However, as a narrative theoretical synthesis, several important limitations must be acknowledged, which underscore its hypothetical nature and the critical need for empirical validation. These limitations also serve to outline priorities for future research aimed at testing and refining the model.
First, regarding the construct of PIU, our model largely treats PIU as a unitary construct for the sake of parsimony and initial integration. In reality, PIU encompasses a heterogeneous range of online activities (eg, gaming, social media use, pornography consumption, online shopping). It is highly plausible that the proposed neurobiological, molecular, and cognitive mechanisms differ in their weighting or presentation across these distinct behavioral subtypes. For instance, reward circuitry hyperactivation may be more central to gaming-related PIU (IGD), while social media-related PIU might be more strongly linked to social anxiety and dysregulation of the social brain network.154 Future research must explicitly test whether the pathways outlined in our model apply equally across PIU subtypes or whether subtype-specific models are needed.
Second, the evidence synthesized is primarily derived from cross-sectional studies comparing individuals with ID or PIU to healthy controls. This precludes definitive causal inferences about the developmental trajectory. While our model posits a bidirectional, reinforcing cycle, the temporal precedence, causal strength, and directionality of the links between biological vulnerability, environmental triggers, initial sleep disruption, and the onset of compulsive internet use remain to be rigorously tested. Therefore, longitudinal cohort studies tracking adolescents from pre- to post-puberty are essential. Such studies should employ multi-modal assessments (neuroimaging, genetics, digital phenotyping, sleep monitoring) at multiple time points to establish the developmental sequence and causal pathways implied by the model.
Third, methodological considerations of this review must be noted. Our synthesis, while comprehensive, was conducted as a narrative theoretical review rather than a systematic review or meta-analysis. This approach was chosen to allow for broad, transdisciplinary integration across often-siloed fields. However, it may be subject to selection bias. Future work would benefit from a systematic mapping and quantitative meta-analysis of the literature to objectively assess the strength, consistency, and effect sizes of the evidence for each proposed pathway within the model.
Finally, we acknowledge that several mechanisms highlighted in our model, such as Default Mode Network dysregulation and prefrontal-striatal circuitry deficits, are transdiagnostic in nature and are implicated in a range of psychopathologies, including depression and anxiety. The specificity of our model lies not in proposing entirely unique mechanisms, but in articulating how these common risk factors interact in a specific, reinforcing configuration to drive the particular comorbidity of ID and PIU during adolescence. A key empirical question is whether this proposed configuration of interactions is more predictive of ID-PIU co-onset than of other common comorbidities.
Embracing these limitations clarifies that the current model is a heuristic framework designed to stimulate hypothesis-driven research. Its primary value is in moving the field from a descriptive understanding of comorbidity towards a mechanistic, integrative, and developmental perspective that generates testable predictions, which can ultimately inform precision prevention and intervention strategies for adolescents.
Conclusion
In summary, the comorbidity between ID and PIU in adolescents represents a significant public health concern that may be underpinned by a confluence of biological, environmental, and neurocognitive factors. Recognizing their interconnectedness is an important step. The present model offers a heuristic framework that could inform a shift toward more integrative and preventative strategies. By proposing a convergent developmental trajectory, this model advances the field by moving beyond descriptive comorbidity and delineating testable, multi-level mechanisms that cut across traditional diagnostic boundaries. This framework invites interdisciplinary research and calls for integrated, transdiagnostic intervention strategies tailored to the adolescent developmental window. Addressing this complex issue will likely require sustained interdisciplinary collaboration across neuroscience, clinical practice, education, and policy to effectively support adolescent well-being in the digital age.
Abbreviations
ACC, Anterior Cingulate Cortex; ACEs, Adverse Childhood Experiences; ADHD, Attention-Deficit/Hyperactivity Disorder; AMG, Amygdala; ATL, Anterior Temporal Lobe; CBT, Cognitive Behavioral Therapy; CEN, Central Executive Network; CHRNA4, Cholinergic Receptor Nicotinic Alpha 4 Subunit; CLOCK, Circadian Locomotor Output Cycles Kaput; CIUS, Compulsive Internet Use Scale; CRP, C-Reactive Protein; DA, Dopamine; DAT, Dopamine Transporter; DAT1, Dopamine Transporter Gene (SLC6A3); DLPFC, Dorsolateral Prefrontal Cortex; DMN, Default Mode Network; DSM-5, Diagnostic and Statistical Manual of Mental Disorders, 5th Edition; dMPFC, Dorsomedial Prefrontal Cortex; EEG, Electroencephalogram; FES, Family Environment Scale; FPN, Frontoparietal Network; GABA, Gamma-Aminobutyric Acid; GD, Gaming Disorder; GM, Gray Matter; GWAS, Genome-Wide Association Study; HPA, Hypothalamic-Pituitary-Adrenal; HTR2A, 5-Hydroxytryptamine Receptor 2A; IA, Internet Addiction; ICD-11, International Classification of Diseases, 11th Revision; IAT, Internet Addiction Test; ID, Insomnia Disorder; IGD, Internet Gaming Disorder; IL-6, Interleukin-6; IPL, Inferior Parietal Lobule; ISAI, Insomnia Self-Assessment Inventory; ISI, Insomnia Severity Index; LTL, Leukocyte Telomere Length; MCC, Mid-Cingulate Cortex; MAOA, Monoamine Oxidase A; mPFC, Medial Prefrontal Cortex; MTNR1B, Melatonin Receptor 1B; MTL, Middle Temporal Lobe or Medial Temporal Lobe; NAc, Nucleus Accumbens; NREM, Non-Rapid Eye Movement (Sleep); OXTR, Oxytocin Receptor; PCC, Posterior Cingulate Cortex; PER2/PER3, Period Circadian Regulator 2/3; PHC, Parahippocampal Cortex; PIU, Problematic Internet Use; PLIC, Posterior Limb of Internal Capsule; PFC, Prefrontal Cortex; PSQI, Pittsburgh Sleep Quality Index; rACC, Rostral Anterior Cingulate Cortex; REM, Rapid Eye Movement (Sleep); rAI, Right Anterior Insula; rHip, Right Hippocampus; SLC6A4, Solute Carrier Family 6 Member 4 (Serotonin Transporter Gene); SMA, Supplementary Motor Area; SN, Salience Network; SNP, Single Nucleotide Polymorphism; STP, Superior Temporal Pole; TPJ, Temporoparietal Junction; VLPFC, Ventrolateral Prefrontal Cortex; WHO, World Health Organization.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Sichuan Science and Technology Program [2025YFHZ0108] and Chengdu Science and Technology Program [2024-KP01-00024-SN] & [2024-YF05-01745-SN].
Disclosure
The authors report no conflicts of interest in this work.
References
1. First MB. Diagnostic and statistical manual of mental disorders, 5th edition, and clinical utility. J Nerv Ment Dis. 2013;201(9):727–23. doi:10.1097/NMD.0b013e3182a2168a
2. Wang HY, Cheng C. Psychometric Evaluation and Comparison of Two Gaming Disorder Measures Derived From the DSM-5 and ICD-11 Frameworks. Front Psychiatry. 2020;11:577366. doi:10.3389/fpsyt.2020.577366
3. Brand M, Rumpf HJ, Demetrovics Z, et al. Which conditions should be considered as disorders in the International Classification of Diseases (ICD-11) designation of “other specified disorders due to addictive behaviors”? J Behav Addict. 2020;11(2):150–159. doi:10.1556/2006.2020.00035
4. Kuss DJ, Lopez-Fernandez O. Internet addiction and problematic Internet use: a systematic review of clinical research. World J Psychiatry. 2016;6(1):143–176. doi:10.5498/wjp.v6.i1.143
5. Shaw P, Kabani NJ, Lerch JP, et al. Neurodevelopmental trajectories of the human cerebral cortex. J Neurosci. 2008;28(14):3586–3594. doi:10.1523/jneurosci.5309-07.2008
6. Gilmore JH, Knickmeyer RC, Gao W. Imaging structural and functional brain development in early childhood. Nat Rev Neurosci. 2018;19(3):123–137. doi:10.1038/nrn.2018.1
7. Johnson EO, Roth T, Schultz L, Breslau N. Epidemiology of DSM-IV insomnia in adolescence: lifetime prevalence, chronicity, and an emergent gender difference. Pediatrics. 2006;117(2):e247–56. doi:10.1542/peds.2004-2629
8. Roberts RE, Roberts CR, Duong HT. Chronic insomnia and its negative consequences for health and functioning of adolescents: a 12-month prospective study. J Adolesc Health. 2008;42(3):294–302. doi:10.1016/j.jadohealth.2007.09.016
9. Hysing M, Pallesen S, Stormark KM, Lundervold AJ, Sivertsen B. Sleep patterns and insomnia among adolescents: a population-based study. J Sleep Res. 2013;22(5):549–556. doi:10.1111/jsr.12055
10. Liang M, Guo L, Huo J, Zhou G, Taheri S. Prevalence of sleep disturbances in Chinese adolescents: a systematic review and meta-analysis. PLoS One. 2021;16(3):e0247333. doi:10.1371/journal.pone.0247333
11. Gao YX, Wang JY, Dong GH. The prevalence and possible risk factors of internet gaming disorder among adolescents and young adults: systematic reviews and meta-analyses. J Psychiatr Res. 2022;154:35–43. doi:10.1016/j.jpsychires.2022.06.049
12. MacKenzie MD, Scott H, Reid K, Gardani M. Adolescent perspectives of bedtime social media use: a qualitative systematic review and thematic synthesis. Sleep Med Rev. 2022;63:101626. doi:10.1016/j.smrv.2022.101626
13. Amaral O, Garrido A, Pereira C, Veiga N, Serpa C, Sakellarides C. Sleep patterns and insomnia among Portuguese adolescents: a cross-sectional study. Aten Primaria. 2014;46(5):191–194. doi:10.1016/s0212-6567(14)70090-3
14. Tashjian SM, Mullins JL, Galván A. Bedtime Autonomy and Cellphone Use Influence Sleep Duration in Adolescents. J Adolesc Health. 2019;64(1):124–130. doi:10.1016/j.jadohealth.2018.07.018
15. Levenson JC, Shensa A, Sidani JE, Colditz JB, Primack BA. The association between social media use and sleep disturbance among young adults. Prev Med. 2016;85:36–41. doi:10.1016/j.ypmed.2016.01.001
16. Garett R, Liu S, Young SD. The Relationship Between Social Media Use and Sleep Quality among Undergraduate Students. Inf Commun Soc. 2018;21(2):163–173. doi:10.1080/1369118x.2016.1266374
17. Lopez KN, Karlsten M, Bonaduce De Nigris F, et al. Understanding Age-based Transition Needs: perspectives from Adolescents and Adults with Congenital Heart Disease. Congenit Heart Dis. 2015;10(6):561–571. doi:10.1111/chd.12283
18. Fekih-Romdhane F, Lamloum E, Loch AA, Cherif W, Cheour M, Hallit S. The relationship between internet gaming disorder and psychotic experiences: cyberbullying and insomnia severity as mediators. BMC Psychiatry. 2023;23(1):857. doi:10.1186/s12888-023-05363-x
19. Chambers RA, Taylor JR, Potenza MN. Developmental neurocircuitry of motivation in adolescence: a critical period of addiction vulnerability. Am J Psychiatry. 2003;160(6):1041–1052. doi:10.1176/appi.ajp.160.6.1041
20. Dong G, Huang J, Du X. Enhanced reward sensitivity and decreased loss sensitivity in Internet addicts: an fMRI study during a guessing task. J Psychiatr Res. 2011;45(11):1525–1529. doi:10.1016/j.jpsychires.2011.06.017
21. Marques DR, Gomes AA, Caetano G, Castelo-Branco M. Insomnia Disorder and Brain’s Default-Mode Network. Curr Neurol Neurosci Rep. 2018;18(8):45. doi:10.1007/s11910-018-0861-3
22. Schiel JE, Holub F, Petri R, et al. Affect and Arousal in Insomnia: through a Lens of Neuroimaging Studies. Curr Psychiatry Rep. 2020;22(9):44. doi:10.1007/s11920-020-01173-0
23. Wang L, Shen H, Lei Y, et al. Altered default mode, fronto-parietal and salience networks in adolescents with Internet addiction. Addict Behav. 2017;70:1–6. doi:10.1016/j.addbeh.2017.01.021
24. Lee J, Lee D, Namkoong K, Jung YC. Aberrant posterior superior temporal sulcus functional connectivity and executive dysfunction in adolescents with internet gaming disorder. J Behav Addict. 2020;9(3):589–597. doi:10.1556/2006.2020.00060
25. Jansen PR, Watanabe K, Stringer S, et al. Genome-wide analysis of insomnia in 1,331,010 individuals identifies new risk loci and functional pathways. Nat Genet. 2019;51(3):394–403. doi:10.1038/s41588-018-0333-3
26. Irwin MR, Olmstead R, Carroll JE. Sleep Disturbance, Sleep Duration, and Inflammation: a Systematic Review and Meta-Analysis of Cohort Studies and Experimental Sleep Deprivation. Biol Psychiatry. 2016;80(1):40–52. doi:10.1016/j.biopsych.2015.05.014
27. Brand M, Müller A, Wegmann E, et al. Current interpretations of the I-PACE model of behavioral addictions. J Behav Addict. 2025;14(1):1–17. doi:10.1556/2006.2025.00020
28. Grau-Rivera O, Operto G, Falcón C, et al. Association between insomnia and cognitive performance, gray matter volume, and white matter microstructure in cognitively unimpaired adults. Alzheimers Res Ther. 2020;12(1):4. doi:10.1186/s13195-019-0547-3
29. Lapidaire W, Urrila AS, Artiges E, et al. Irregular sleep habits, regional grey matter volumes, and psychological functioning in adolescents. PLoS One. 2021;16(2):e0243720. doi:10.1371/journal.pone.0243720
30. Chao LL, Mohlenhoff BS, Weiner MW, Neylan TC. Associations between subjective sleep quality and brain volume in Gulf War veterans. Sleep. 2014;37(3):445–452. doi:10.5665/sleep.3472
31. Gracia-Tabuenca Z, Moreno MB, Barrios FA, Alcauter S. Development of the brain functional connectome follows puberty-dependent nonlinear trajectories. Neuroimage. 2021;229:117769. doi:10.1016/j.neuroimage.2021.117769
32. Chen MC, Chang C, Glover GH, Gotlib IH. Increased insula coactivation with salience networks in insomnia. Biol Psychol. 2014;97:1–8. doi:10.1016/j.biopsycho.2013.12.016
33. Sfera A, Thomas KA, Ogunjale IA, Jafri N, Bota PG. Insomnia in Forensic Detainees: is Salience Network the Common Pathway for Sleep, Neuropsychiatric, and Neurodegenerative Disorders? J Clin Med. 2024;13(6):1691. doi:10.3390/jcm13061691
34. Li C, Dong M, Yin Y, Hua K, Fu S, Jiang G. Aberrant Effective Connectivity of the Right Anterior Insula in Primary Insomnia. Front Neurol. 2018;9:317. doi:10.3389/fneur.2018.00317
35. Gong L, Liao T, Liu D, et al. Amygdala Changes in Chronic Insomnia and Their Association with Sleep and Anxiety Symptoms: insight from Shape Analysis. Neural Plast. 2019;2019:8549237. doi:10.1155/2019/8549237
36. Sergerie K, Chochol C, Armony JL. The role of the amygdala in emotional processing: a quantitative meta-analysis of functional neuroimaging studies. Neurosci Biobehav Rev. 2008;32(4):811–830. doi:10.1016/j.neubiorev.2007.12.002
37. Kweon W, Lee KH, Choi SH, et al. Amygdala resting-state functional connectivity alterations in patients with chronic insomnia disorder: correlation with electroencephalography beta power during sleep. Sleep. 2023;46(10):205. doi:10.1093/sleep/zsad205
38. Ling J, Lin X, Li X, et al. Neural response to rewards in youths with insomnia. Sleep. 2022;45(2):238. doi:10.1093/sleep/zsab238
39. Li YD, Luo YJ, Su WK, et al. Anterior cingulate cortex projections to the dorsal medial striatum underlie insomnia associated with chronic pain. Neuron. 2024;112(8):1328–1341.e4. doi:10.1016/j.neuron.2024.01.014
40. Zhang R, Volkow ND. Brain default-mode network dysfunction in addiction. Neuroimage. 2019;200:313–331. doi:10.1016/j.neuroimage.2019.06.036
41. Wang Y, Qin Y, Li H, et al. Abnormal Functional Connectivity in Cognitive Control Network, Default Mode Network, and Visual Attention Network in Internet Addiction: a Resting-State fMRI Study. Front Neurol. 2019;10:1006. doi:10.3389/fneur.2019.01006
42. Wang L, Zhang Z, Wang S, et al. Deficient dynamics of prefrontal-striatal and striatal-default mode network (DMN) neural circuits in internet gaming disorder. J Affect Disord. 2023;323:336–344. doi:10.1016/j.jad.2022.11.074
43. Chen S, Wang M, Dong H, et al. Internet gaming disorder impacts gray matter structural covariance organization in the default mode network. J Affect Disord. 2021;288:23–30. doi:10.1016/j.jad.2021.03.077
44. Lee D, Namkoong K, Lee J, Jung YC. Abnormal gray matter volume and impulsivity in young adults with Internet gaming disorder. Addict Biol. 2018;23(5):1160–1167. doi:10.1111/adb.12552
45. Cheng H, Liu J. Alterations in Amygdala Connectivity in Internet Addiction Disorder. Sci Rep. 2020;10(1):2370. doi:10.1038/s41598-020-59195-w
46. Marciano L, Camerini AL, Morese R. The Developing Brain in the Digital Era: a Scoping Review of Structural and Functional Correlates of Screen Time in Adolescence. Front Psychol. 2021;12:671817. doi:10.3389/fpsyg.2021.671817
47. Cai C, Yuan K, Yin J, et al. Striatum morphometry is associated with cognitive control deficits and symptom severity in internet gaming disorder. Brain Imaging Behav. 2016;10(1):12–20. doi:10.1007/s11682-015-9358-8
48. Brand M, Snagowski J, Laier C, Maderwald S. Ventral striatum activity when watching preferred pornographic pictures is correlated with symptoms of Internet pornography addiction. Neuroimage. 2016;129:224–232. doi:10.1016/j.neuroimage.2016.01.033
49. Floresco SB. The nucleus accumbens: an interface between cognition, emotion, and action. Annu Rev Psychol. 2015;66:25–52. doi:10.1146/annurev-psych-010213-115159
50. Qiu X, Han X, Wang Y, et al. Interaction effects between smoking and internet gaming disorder on resting-state functional connectivity of the ventral tegmental area and hippocampus. Front Neurosci. 2023;17:1270014. doi:10.3389/fnins.2023.1270014
51. Bai Y, Qu J, Li D, Yin H. Neural basis underlying the relation between internet addiction tendency and sleep quality: the intrinsic default-mode network connectivity pathways. Int J Psychophysiol. 2024;195:112264. doi:10.1016/j.ijpsycho.2023.112264
52. Braun K. The prefrontal-limbic system: development, neuroanatomy, function, and implications for socioemotional development. Clin Perinatol. 2011;38(4):685–702. doi:10.1016/j.clp.2011.08.013
53. Lu F, Cui Q, He Z, et al. Prefrontal-limbic-striatum dysconnectivity associated with negative emotional endophenotypes in bipolar disorder during depressive episodes. J Affect Disord. 2021;295:422–430. doi:10.1016/j.jad.2021.08.055
54. Hong T, Zhou H, Xi W, et al. Acting with awareness is positively correlated with dorsal anterior cingulate cortex glutamate concentration but both are impaired in Internet gaming disorder. Neuroscience. 2025;564:226–235. doi:10.1016/j.neuroscience.2024.11.054
55. Fan F, Liao X, Lei T, et al. Development of the default-mode network during childhood and adolescence: a longitudinal resting-state fMRI study. Neuroimage. 2021;226:117581. doi:10.1016/j.neuroimage.2020.117581
56. Ko M, Chi SH, Lee JH, Suh SI, Lee MS. Altered Functional Connectivity of the Nucleus Accumbens and Amygdala in Cyber Addiction: a Resting State Functional Magnetic Resonance Imaging Study. Clin Psychopharmacol Neurosci. 2023;21(2):304–312. doi:10.9758/cpn.2023.21.2.304
57. von Rhein D, Cools R, Zwiers MP, et al. Increased neural responses to reward in adolescents and young adults with attention-deficit/hyperactivity disorder and their unaffected siblings. J Am Acad Child Adolesc Psychiatry. 2015;54(5):394–402. doi:10.1016/j.jaac.2015.02.012
58. Rebello K, Moura LM, Xavier G, et al. Association between spontaneous activity of the default mode network hubs and leukocyte telomere length in late childhood and early adolescence. J Psychosom Res. 2019;127:109864. doi:10.1016/j.jpsychores.2019.109864
59. Berke JD. What does dopamine mean? Nat Neurosci. 2018;21(6):787–793. doi:10.1038/s41593-018-0152-y
60. Monti JM, Monti D. The involvement of dopamine in the modulation of sleep and waking. Sleep Med Rev. 2007;11(2):113–133. doi:10.1016/j.smrv.2006.08.003
61. Dauvilliers Y, Tafti M, Landolt HP. Catechol-O-methyltransferase, dopamine, and sleep-wake regulation. Sleep Med Rev. 2015;22:47–53. doi:10.1016/j.smrv.2014.10.006
62. Kashiwagi M, Kanuka M, Tanaka K, et al. Impaired wakefulness and rapid eye movement sleep in dopamine-deficient mice. Mol Brain. 2021;14(1):170. doi:10.1186/s13041-021-00879-3
63. Gottesmann C. GABA mechanisms and sleep. Neuroscience. 2002;111(2):231–239. doi:10.1016/s0306-4522(02)00034-9
64. Bowery NG, Hudson AL, Price GW. GABAA and GABAB receptor site distribution in the rat central nervous system. Neuroscience. 1987;20(2):365–383. doi:10.1016/0306-4522(87)90098-4
65. Yu X, Li W, Ma Y, et al. GABA and glutamate neurons in the VTA regulate sleep and wakefulness. Nat Neurosci. 2019;22(1):106–119. doi:10.1038/s41593-018-0288-9
66. Xiang T, Liao J, Cai Y, et al. Impairment of GABA inhibition in insomnia disorders: evidence from the peripheral blood system. Front Psychiatry. 2023;14:1134434. doi:10.3389/fpsyt.2023.1134434
67. Bollu PC, Kaur H. Sleep Medicine: insomnia and Sleep. Mo Med. 2019;116(1):68–75.
68. Saper CB, Scammell TE, Lu J. Hypothalamic regulation of sleep and circadian rhythms. Nature. 2005;437(7063):1257–1263. doi:10.1038/nature04284
69. Li SB, Damonte VM, Chen C, et al. Hyperexcitable arousal circuits drive sleep instability during aging. Science. 2022;375:
70. Muehlan C, Roch C, Vaillant C, Dingemanse J. The orexin story and orexin receptor antagonists for the treatment of insomnia. J Sleep Res. 2023;32(6):e13902. doi:10.1111/jsr.13902
71. Claustrat B, Brun J, Chazot G. The basic physiology and pathophysiology of melatonin. Sleep Med Rev. 2005;9(1):11–24. doi:10.1016/j.smrv.2004.08.001
72. Uchiyama M. Melatonin receptor agonist. Nihon Rinsho. 2015;73(6):1017–1022.
73. Choi K, Lee YJ, Park S, Je NK, Suh HS. Efficacy of melatonin for chronic insomnia: systematic reviews and meta-analyses. Sleep Med Rev. 2022;66:101692. doi:10.1016/j.smrv.2022.101692
74. Riemann D, Espie CA, Altena E, et al. The European Insomnia Guideline: an update on the diagnosis and treatment of insomnia 2023. J Sleep Res. 2023;32(6):e14035. doi:10.1111/jsr.14035
75. Chen HW, Zhou R, Cao BF, et al. The predictive, preventive, and personalized medicine of insomnia: gut microbiota and inflammation. Epma j. 2023;14(4):571–583. doi:10.1007/s13167-023-00345-1
76. Fei CJ, Li ZY, Ning J, et al. Exome sequencing identifies genes associated with sleep-related traits. Nat Hum Behav. 2024;8(3):576–589. doi:10.1038/s41562-023-01785-5
77. Wise RA, Koob GF. The development and maintenance of drug addiction. Neuropsychopharmacology. 2014;39(2):254–262. doi:10.1038/npp.2013.261
78. Wise RA, Robble MA. Dopamine and Addiction. Annu Rev Psychol. 2020;71:79–106. doi:10.1146/annurev-psych-010418-103337
79. Carpita B, Muti D, Nardi B, et al. Biochemical Correlates of Video Game Use: from Physiology to Pathology. A Narrative Review. Life. 2021;11(8):775. doi:10.3390/life11080775
80. Liu M, Luo J. Relationship between peripheral blood dopamine level and internet addiction disorder in adolescents: a pilot study. Int J Clin Exp Med. 2015;8(6):9943–9948.
81. Tops M, Russo S, Boksem MA, Tucker DM. Serotonin: modulator of a drive to withdraw. Brain Cogn. 2009;71(3):427–436. doi:10.1016/j.bandc.2009.03.009
82. Mohammad-Zadeh LF, Moses L, Gwaltney-Brant SM. Serotonin: a review. J Vet Pharmacol Ther. 2008;31(3):187–199. doi:10.1111/j.1365-2885.2008.00944.x
83. Cunningham KA, Anastasio NC. Serotonin at the nexus of impulsivity and cue reactivity in cocaine addiction. Neuropharmacology. 2014;760:460–478. doi:10.1016/j.neuropharm.2013.06.030
84. Elvig SK, McGinn MA, Smith C, Arends MA, Koob GF, Vendruscolo LF. Tolerance to alcohol: a critical yet understudied factor in alcohol addiction. Pharmacol Biochem Behav. 2021;204:173155. doi:10.1016/j.pbb.2021.173155
85. Cerniglia L, Cimino S, Marzilli E, Pascale E, Tambelli R. Associations Among Internet Addiction, Genetic Polymorphisms, Family Functioning, and Psychopathological Risk: cross-Sectional Exploratory Study. JMIR Ment Health. 2020;7(12):e17341. doi:10.2196/17341
86. Choi MR, Cho H, Chun JW, Yoo JH, Kim DJ. Increase of orexin A in the peripheral blood of adolescents with Internet gaming disorder. J Behav Addict. 2020;9(1):93–104. doi:10.1556/2006.8.2019.65
87. Paik SH, Choi MR, Kwak SM, Bang SH, Kim DJ. Decreased Serum Glutamate Levels in Male Adults with Internet Gaming Disorder: a Pilot Study. Clin Psychopharmacol Neurosci. 2018;16(3):276–281. doi:10.9758/cpn.2018.16.3.276
88. Chang AM, Aeschbach D, Duffy JF, Czeisler CA. Evening use of light-emitting eReaders negatively affects sleep, circadian timing, and next-morning alertness. Proc Natl Acad Sci U S A. 2015;112(4):1232–1237. doi:10.1073/pnas.1418490112
89. Dai Y, Zhang C, Zhang L, Wen C, Li H, Zhu T. Genetic polymorphism in HTR2A rs6313 is associated with internet addiction disorder. Front Psychiatry. 2024;15:1292877. doi:10.3389/fpsyt.2024.1292877
90. Rył A, Tomska N, Jakubowska A, Ogrodniczak A, Palma J, Rotter I. Genetic Aspects of Problematic and Risky Internet Use in Young Men-Analysis of ANKK1, DRD2 and NTRK3 Gene Polymorphism. Genes (Basel). 2024;15(2):169. doi:10.3390/genes15020169
91. Yen JY, Chou WP, Lin HC, Wu HC, Tsai WX, Ko CH. Roles of Hostility and Depression in the Association between the MAOA Gene Polymorphism and Internet Gaming Disorder. Int J Environ Res Public Health. 2021;18(13):6910. doi:10.3390/ijerph18136910
92. De Nardi L, Carpentieri V, Pascale E, et al. Involvement of DAT1 Gene on Internet Addiction: cross-Correlations of Methylation Levels in 5’-UTR and 3’-UTR Genotypes, Interact with Impulsivity and Attachment-Driven Quality of Relationships. Int J Environ Res Public Health. 2020;17(21):7956. doi:10.3390/ijerph17217956
93. Liang Q, Yu C, Xing Q, Chen P, Li S. Parent-Adolescent Conflict, Peer Victimization, and Internet Gaming Disorder Among Chinese Adolescents: the Moderating Effect of OXTR Gene rs53576 Polymorphism. Child Psychiatry Hum Dev. 2024;55(6):1634–1643. doi:10.1007/s10578-023-01523-y
94. Montag C, Kirsch P, Sauer C, Markett S, Reuter M. The role of the CHRNA4 gene in Internet addiction: a case-control study. J Addict Med. 2012;6(3):191–195. doi:10.1097/ADM.0b013e31825ba7e7
95. Tereshchenko SY, Afonicheva KV, Marchenko IV, Shubina MV, Smolnikova MV. Polymorphic variants of the dopamine receptor gene DRD2 (rs6277, rs1800497) in adolescents with problematic video game use. Vavilovskii Zhurnal Genet Selektsii. 2024;28(6):667–674. doi:10.18699/vjgb-24-74
96. Kim JY, Jeong JE, Rhee JK, et al. Targeted exome sequencing for the identification of a protective variant against Internet gaming disorder at rs2229910 of neurotrophic tyrosine kinase receptor, type 3 (NTRK3): a pilot study. J Behav Addict. 2016;5(4):631–638. doi:10.1556/2006.5.2016.077
97. Crowley SJ, Cain SW, Burns AC, Acebo C, Carskadon MA. Increased Sensitivity of the Circadian System to Light in Early/Mid-Puberty. J Clin Endocrinol Metab. 2015;100(11):4067–4073. doi:10.1210/jc.2015-2775
98. Illingworth G, Sharman R, Harvey CJ, Foster RG, Espie CA. The Teensleep study: the effectiveness of a school-based sleep education programme at improving early adolescent sleep. Sleep Med X. 2020;2:100011. doi:10.1016/j.sleepx.2019.100011
99. Nicolaides NC, Vgontzas AN, Kritikou I, Chrousos G. HPA Axis and Sleep. Feingold KR, Ahmed SF, Anawalt B, eds.. Endotext. MDText.com, Inc; 2000.
100. Mousikou M, Kyriakou A, Skordis N. Stress and Growth in Children and Adolescents. Horm Res Paediatr. 2023;96(1):25–33. doi:10.1159/000521074
101. Spear LP. Rewards, aversions and affect in adolescence: emerging convergences across laboratory animal and human data. Developmental Cognitive Neuroscience. 2011;1(4):392–400. doi:10.1016/j.dcn.2011.08.001
102. Wahlstrom D, Collins P, White T, Luciana M. Developmental changes in dopamine neurotransmission in adolescence: behavioral implications and issues in assessment. Brain Cogn. 2010;72(1):146–159. doi:10.1016/j.bandc.2009.10.013
103. Steinberg L. Cognitive and affective development in adolescence. Trends Cognit Sci. 2005;9(2):69–74. doi:10.1016/j.tics.2004.12.005
104. Malti T, Galarneau E, Peplak J. Moral Development in Adolescence. J Res Adolesc. 2021;31(4):1097–1113. doi:10.1111/jora.12639
105. Tor J, Dolz M, Sintes A, et al. Clinical high risk for psychosis in children and adolescents: a systematic review. Eur Child Adolesc Psychiatry. 2018;27(6):683–700. doi:10.1007/s00787-017-1046-3
106. de Zambotti M, Goldstone A, Colrain IM, Baker FC. Insomnia disorder in adolescence: diagnosis, impact, and treatment. Sleep Med Rev. 2018;(39):12–24. doi:10.1016/j.smrv.2017.06.009
107. Goldberg SG, Wagner K. American Psychological Association practice guidelines for psychopharmacology: ethical practice considerations for psychologists involving psychotropic use with children and adolescents. J Clin Psychol. 2019;75(3):344–363. doi:10.1002/jclp.22705
108. Bauducco SV, Özdemir M, Gradisar M, Boersma K, Bayram-özdemir S. Trajectories of insomnia symptoms and insufficient sleep duration in early adolescents: associations with school stress. Sleep Adv. 2022;3(1):zpac018. doi:10.1093/sleepadvances/zpac018
109. Zhang WJ, Yan C, Shum D, Deng CP. Responses to academic stress mediate the association between sleep difficulties and depressive/anxiety symptoms in Chinese adolescents. J Affect Disord. 2020;263:89–98. doi:10.1016/j.jad.2019.11.157
110. Oh CH, Wallace ML, Germain A. Childhood trauma and gender: synergistic and additive effects on sleep in healthy young adults. Sleep Health. 2022;8(5):498–504. doi:10.1016/j.sleh.2022.06.008
111. Reffi AN, Kalmbach DA, Cheng P, et al. Sleep reactivity as a potential pathway from childhood abuse to adult insomnia. Sleep Med. 2022;94:70–75. doi:10.1016/j.sleep.2022.03.026
112. Pfaff A, Schlarb AA. Consequences of child maltreatment: a glimpse at stress and sleep. J Sleep Res. 2022;31(2):e13456. doi:10.1111/jsr.13456
113. Koivusilta L, Kuoppamäki H, Rimpelä A. Energy drink consumption, health complaints and late bedtime among young adolescents. Int J Public Health. 2016;61(3):299–306. doi:10.1007/s00038-016-0797-9
114. Choi K, Son H, Park M, et al. Internet overuse and excessive daytime sleepiness in adolescents. Psychiatry Clin Neurosci. 2009;63(4):455–462. doi:10.1111/j.1440-1819.2009.01925.x
115. Gasperetti CE, Dolsen EA, Harvey AG. The influence of intensity and timing of daily light exposure on subjective and objective sleep in adolescents with an evening circadian preference. Sleep Med. 2021;79:166–174. doi:10.1016/j.sleep.2020.11.014
116. Paksarian D, Rudolph KE, Stapp EK, et al. Association of Outdoor Artificial Light at Night With Mental Disorders and Sleep Patterns Among US Adolescents. JAMA Psychiatry. 2020;77(12):1266–1275. doi:10.1001/jamapsychiatry.2020.1935
117. Ricketts EJ, Joyce DS, Rissman AJ, et al. Electric lighting, adolescent sleep and circadian outcomes, and recommendations for improving light health. Sleep Med Rev. 2022;64:101667. doi:10.1016/j.smrv.2022.101667
118. Hartley S, Royant-Parola S, Zayoud A, Gremy I, Matulonga B. Do both timing and duration of screen use affect sleep patterns in adolescents? PLoS One. 2022;17(10):e0276226. doi:10.1371/journal.pone.0276226
119. Perrault AA, Bayer L, Peuvrier M, et al. Reducing the use of screen electronic devices in the evening is associated with improved sleep and daytime vigilance in adolescents. Sleep. 2019;42(9):125. doi:10.1093/sleep/zsz125
120. Younes F, Halawi G, Jabbour H, et al. Internet Addiction and Relationships with Insomnia, Anxiety, Depression, Stress and Self-Esteem in University Students: a Cross-Sectional Designed Study. PLoS One. 2016;11(9):e0161126. doi:10.1371/journal.pone.0161126
121. Tang H, Li Y, Dong W, et al. The relationship between childhood trauma and internet addiction in adolescents: a meta-analysis. J Behav Addict. 2024;13(1):36–50. doi:10.1556/2006.2024.00001
122. Lin WH, Chiao C. Relationship between adverse childhood experiences and problematic internet use among young adults: the role of the feeling of loneliness trajectory. J Behav Addict. 2022;11(4):1080–1091. doi:10.1556/2006.2022.00074
123. Tom A, Thomas B, Sharma M, Joseph A. Parental rejection and control: potential risks for excessive Internet usage among adolescents. Int J Soc Psychiatry. 2023;69(8):2007–2017. doi:10.1177/00207640231185450
124. Chung S, Lee J, Lee HK. Personal Factors, Internet Characteristics, and Environmental Factors Contributing to Adolescent Internet Addiction: a Public Health Perspective. Int J Environ Res Public Health. 2019;16(23):4635. doi:10.3390/ijerph16234635
125. Trumello C, Vismara L, Sechi C, Ricciardi P, Marino V, Babore A. Internet Addiction: the Role of Parental Care and Mental Health in Adolescence. Int J Environ Res Public Health. 2021;18(24):12876. doi:10.3390/ijerph182412876
126. Palagini L, Hertenstein E, Riemann D, Nissen C. Sleep, insomnia and mental health. J Sleep Res. 2022;31(4):e13628. doi:10.1111/jsr.13628
127. Hsieh YP, Lu WH, Yen CF. Psychosocial Determinants of Insomnia in Adolescents: roles of Mental Health, Behavioral Health, and Social Environment. Front Neurosci. 2019;13:848. doi:10.3389/fnins.2019.00848
128. Li M, Yuan Y, Cheng X, Wang Y, Xu Z. Childhood maltreatment and insomnia in college students: the role of alexithymia and psychological distress. Acta Psychol. 2024;243:104149. doi:10.1016/j.actpsy.2024.104149
129. Brand S, Hatzinger M, Beck J, Holsboer-Trachsler E. Perceived parenting styles, personality traits and sleep patterns in adolescents. J Adolesc. 2009;32(5):1189–1207. doi:10.1016/j.adolescence.2009.01.010
130. Majeno A, Tsai KM, Huynh VW, McCreath H, Fuligni AJ. Discrimination and Sleep Difficulties during Adolescence: the Mediating Roles of Loneliness and Perceived Stress. J Youth Adolesc. 2018;47(1):135–147. doi:10.1007/s10964-017-0755-8
131. Alanko D. The Health Effects of Video Games in Children and Adolescents. Pediatr Rev. 2023;44(1):23–32. doi:10.1542/pir.2022-005666
132. Brown WJ, Wilkerson AK, Boyd SJ, Dewey D, Mesa F, Bunnell BE. A review of sleep disturbance in children and adolescents with anxiety. J Sleep Res. 2018;27(3):e12635. doi:10.1111/jsr.12635
133. Larsson I, Aili K, Lönn M, et al. Sleep interventions for children with attention deficit hyperactivity disorder (ADHD): a systematic literature review. Sleep Med. 2023;102:64–75. doi:10.1016/j.sleep.2022.12.021
134. Blank TS, Meyer BM, Wieser MK, Rabl U, Schögl P, Pezawas L. Brain morphometry and connectivity differs between adolescent- and adult-onset major depressive disorder. Depress Anxiety. 2022;39(5):387–396. doi:10.1002/da.23254
135. Nunes ML, Bruni O. Insomnia in childhood and adolescence: clinical aspects, diagnosis, and therapeutic approach. J Pediatr. 2015;91(6 Suppl 1):S26–35. doi:10.1016/j.jped.2015.08.006
136. Nyer M, Farabaugh A, Fehling K, et al. Relationship between sleep disturbance and depression, anxiety, and functioning in college students. Depress Anxiety. 2013;30(9):873–880. doi:10.1002/da.22064
137. Mei S, Yau YH, Chai J, Guo J, Potenza MN. Problematic Internet use, well-being, self-esteem and self-control: data from a high-school survey in China. Addict Behav. 2016;61:74–79. doi:10.1016/j.addbeh.2016.05.009
138. Guo L, Shi G, Du X, Wang W, Guo Y, Lu C. Associations of emotional and behavioral problems with Internet use among Chinese young adults: the role of academic performance. J Affect Disord. 2021;287:214–221. doi:10.1016/j.jad.2021.03.050
139. Ciarrochi J, Parker P, Sahdra B, et al. The development of compulsive internet use and mental health: a four-year study of adolescence. Dev Psychol. 2016;52(2):272–283. doi:10.1037/dev0000070
140. Pontes HM, Macur M. Problematic internet use profiles and psychosocial risk among adolescents. PLoS One. 2021;16(9):e0257329. doi:10.1371/journal.pone.0257329
141. López-Muciño LA, García-García F, Cueto-Escobedo J, Acosta-Hernández M, Venebra-Muñoz A, Rodríguez-Alba JC. Sleep loss and addiction. Neurosci Biobehav Rev. 2022;141:104832. doi:10.1016/j.neubiorev.2022.104832
142. Luciana M. Adolescent brain development in normality and psychopathology. Dev Psychopathol. 2013;25(4 Pt 2):1325–1345. doi:10.1017/s0954579413000643
143. Luciana M, Bjork JM, Nagel BJ, et al. Adolescent neurocognitive development and impacts of substance use: overview of the adolescent brain cognitive development (ABCD) baseline neurocognition battery. Dev Cogn Neurosci. 2018;32:67–79. doi:10.1016/j.dcn.2018.02.006
144. Dresp-Langley B, Hutt A. Digital Addiction and Sleep. Int J Environ Res Public Health. 2022;19(11):6910. doi:10.3390/ijerph19116910
145. Morin CM, Vallières A, Ivers H. Dysfunctional beliefs and attitudes about sleep (DBAS): validation of a brief version (DBAS-16). Sleep. 2007;30(11):1547–1554. doi:10.1093/sleep/30.11.1547
146. Wegmann E, Antons S, Brand M. The experience of gratification and compensation in addictive behaviors: how can these experiences be measured systematically within and across disorders due to addictive behaviors? Compr Psychiatry. 2022;117:152336. doi:10.1016/j.comppsych.2022.152336
147. Ko CH, Hsieh TJ, Chen CY, et al. Altered brain activation during response inhibition and error processing in subjects with Internet gaming disorder: a functional magnetic imaging study. Eur Arch Psychiatry Clin Neurosci. 2014;264(8):661–672. doi:10.1007/s00406-013-0483-3
148. Brand M, Young KS, Laier C. Prefrontal control and internet addiction: a theoretical model and review of neuropsychological and neuroimaging findings. Front Hum Neurosci. 2014;8:375. doi:10.3389/fnhum.2014.00375
149. Alimoradi Z, Lin CY, Broström A, et al. Internet addiction and sleep problems: a systematic review and meta-analysis. Sleep Med Rev. 2019;47:51–61. doi:10.1016/j.smrv.2019.06.004
150. Best O, Ban S. Adolescence: physical changes and neurological development. Br J Nurs. 2021;30(5):272–275. doi:10.12968/bjon.2021.30.5.272
151. Atrooz F, Salim S. Sleep deprivation, oxidative stress and inflammation. Adv Protein Chem Struct Biol. 2020;119:309–336. doi:10.1016/bs.apcsb.2019.03.001
152. Touitou Y, Touitou D, Reinberg A. Disruption of adolescents’ circadian clock: the vicious circle of media use, exposure to light at night, sleep loss and risk behaviors. J Physiol Paris. 2016;110(4 Pt B):467–479. doi:10.1016/j.jphysparis.2017.05.001
153. Ayub S, Jain L, Parnia S, et al. Treatment Modalities for Internet Addiction in Children and Adolescents: a Systematic Review of Randomized Controlled Trials (RCTs). J Clin Med. 2023;12(9):3345. doi:10.3390/jcm12093345
154. Liu C, Liu Y, Liu C, et al. The Moderated Mediating Effects of Social Media Identity and Loneliness on the Relationship Between Problematic Internet Use and Mental Health in China: nationwide Cross-Sectional Questionnaire Study. J Med Internet Res. 2025;27:e57907. doi:10.2196/57907
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.