Back to Journals » Pharmacogenomics and Personalized Medicine » Volume 15
Acyl-CoA Binding Domain Containing 4 Polymorphism rs4986172 and Expression Can Serve as Overall Survival Biomarkers for Hepatitis B Virus-Related Hepatocellular Carcinoma Patients After Hepatectomy
Authors Huang H, Liao X ![]() , Zhu G
, Zhu G ![]() , Han C, Wang X, Yang C
, Han C, Wang X, Yang C ![]() , Zhou X, Liang T, Huang K, Peng T
, Zhou X, Liang T, Huang K, Peng T ![]()
Received 14 November 2021
Accepted for publication 4 March 2022
Published 29 March 2022 Volume 2022:15 Pages 277—300
DOI https://doi.org/10.2147/PGPM.S349350
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Huasheng Huang,* Xiwen Liao,* Guangzhi Zhu, Chuangye Han, Xiangkun Wang, Chengkun Yang, Xin Zhou, Tianyi Liang, Ketuan Huang, Tao Peng
Department of Hepatobiliary Surgery, The First Affiliated Hospital of Guangxi Medical University, Nanning, 530021, Guangxi Zhuang Autonomous Region, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Tao Peng; Xiwen Liao, Department of Hepatobiliary Surgery, The First Affiliated Hospital of Guangxi Medical University, Shuang Yong Road 6, Nanning, 530021, Guangxi Zhuang Autonomous Region, People’s Republic of China, Tel +86-771-5356528, Fax +86-771-5350031, Email [email protected]; [email protected]
Background: The aim of our study was to evaluate the potential of expression and single nucleotide polymorphism of Acyl-CoA binding domain containing 4 (ACBD4) gene as prognosis biomarkers in patients with hepatitis B virus (HBV)-related hepatocellular carcinoma (HCC) after hepatectomy.
Methods: HBV-related HCC patients from the First Affiliated Hospital of Guangxi Medical University and GSE14520 were included in the current study, as well as The Cancer Genome Atlas (TCGA) HCC verification cohort. Prognostic analysis and multiple functional enrichment analysis methods were used to evaluate the prognostic value and potential biological functions of the ACBD4 gene in HBV-related HCC.
Results: We found that ACBD4 gene is highly expressed in normal liver tissues and markedly down-regulated in HBV-related HCC tissues. ACBD4 gene was significantly related to overall survival (OS) of HCC in TCGA and GSE14520 cohorts, and patients with low ACBD4 expression were markedly related to poor OS. Rs4986172 was observed as an OS biomarker after hepatectomy in the Guangxi HBV-related HCC cohort. The OS of rs4986172 GG genotype was worse than that of HCC patients with A allele (AA and AG genotypes). Multifunctional enrichment analysis suggested that ACBD4 gene is closely related to the metabolic, peroxisome proliferator-activated receptor and cytochrome P450 pathway. Through connectivity map, we also identified eight compounds that may be used as targeted therapeutic agents for ACBD4 gene in HBV-related HCC; these compounds were scopoletin, alfaxalone, bephenium hydroxynaphthoate, apramycin, 4,5-dianilinophthalimide, DL-thiorphan, aminohippuric acid and quinidine. Immune microenvironment analysis revealed that there were significant differences in immune scores of HBV-related HCC tumor tissues with different ACBD4 expression levels.
Conclusion: Our study reveals that ACBD4 expression and rs4986172 can be serve as biomarkers of OS in HBV-related HCC patients after hepatectomy.
Keywords: rs4986172, ACBD4, HBV-related HCC, overall survival, hepatectomy
Introduction
Liver cancer is the second leading cause of death among men worldwide, and about half of all liver cancers occur in China.1 Liver cancer is the fourth most common cancer in men in China, and the third leading mortality rate among both men and women.2 A majority of primary liver cancer is hepatocellular carcinomas (HCC). High exposure to aflatoxin and hepatitis B virus (HBV) are the main reasons for the high incidence of HCC in Guangxi.3–5 Occurrence of hepatocellular carcinoma is driven by both genetic and environmental factors. With the progression of high-throughput sequencing, many biomarkers related to the occurrence and prognosis of HCC have been discovered based on high-throughput sequencing databases such as The Cancer Genome Atlas (TCGA) and Gene Expression Omnibus (GEO). By screening the expression abundance of normal liver tissues from the Genotype-Tissue Expression Portal (GTEx) database, we found that Acyl-CoA binding domain containing 4 (ACBD4) gene is highly expressed in normal liver tissues. Through literature review, we know that ACBD4 is a peroxisomal protein, which is closely related to various metabolisms in vivo.6,7 Previous study has shown that ACBD4 gene is associated with cardiac arrhythmias.8 Liao et al. suggested that ACBD4 can be used as a target gene of p53 to be dysregulated in colorectal cancer cells induced by inauhzin, thereby exerting a tumor suppressor effect.9 Previous studies have not reported on the clinical value and molecular mechanism of ACBD4 gene in HCC. Therefore, we designed this study to conduct an in-depth investigation of ACBD4 gene in HBV-related HCC. The purpose of the current study was to assess the possibility of expression and single nucleotide polymorphism of Acyl-CoA binding domain containing 4 (ACBD4) gene as prognosis biomarkers in patients with HBV-related HCC after hepatectomy. Furthermore, we also used a variety of functional enrichment analysis methods to preliminarily identify the molecular mechanisms of ACBD4 in HBV-related HCC.
Materials and Methods
Data Acquisition
The expression distribution of ACBD4 gene in human organs was obtained from GTEx Portal (https://www.gtexportal.org/home/).10 The HBV-related HCC genome-wide expression dataset is derived from GSE14520 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE14520) of Gene Expression Omnibus database.11–17 All the patients had received radical resection in the GSE14520 cohort.16 The validation cohort for the relationship between ACBD4 expression level and HCC prognosis was derived from the TCGA cohort and was calculated by the Kaplan-Meier plotter (http://kmplot.com/analysis/index.php?p=service) online analysis tool.18 To further verify the expression difference of ACBD4 gene between HCC tumor and paracancerous tissues, we also used four GEO datasets as validation queues, including GSE25097,19–21 GSE36376,22–24 GSE54236,25–27 and GSE64041.28 At the same time, we also used the GEPIA online analysis tool (http://gepia.cancer-pku.cn/index.html) TCGA HCC cohort tumor samples and GTEx normal liver samples dataset to compare the expression difference of ACBD4 gene.29 All GEO datasets processing was normalized in R platform by limma software package. Multiple probes were combined to correspond to a gene, and their average value represented the expression level of this gene. Single nucleotide polymorphisms (SNPs) of ACBD4-rs4986172 were derived from whole exon array in The First Affiliated Hospital of Guangxi Medical University HBV-related HCC cohort, and the whole exon array is Human Exome Bead Chip 12v1-1 system (Illumina, Inc., San Diego, CA). Data on 474 new patients with HBV-related HCC who received hepatectomy in The First Affiliated Hospital of Guangxi Medical University from January 2001 to November 2013 were collected, and our study was approved by the Ethics Committee of The First Affiliated Hospital of Guangxi Medical University, the approval number is 2021 (KY-E-097). Written informed consent was obtained from all patients for the procedures in this study.
Survival Analysis
Kaplan-Meier method and Log rank test were applied to univariate survival analysis. Cox proportional hazard regression model was applied for multivariate survival analysis, hazard ratio (HR) and 95% confidence interval (CI) were used to assess the risk ratio for death or recurrence. Stratified survival analysis was applied to assess the prognostic value of ACBD4 gene in different clinical subgroups. Joint effect survival analysis was used to evaluate the combination of ACBD4 gene and traditional clinical parameters to improve the prediction of the HCC prognosis. The nomogram was used to assess the influence of various clinical parameters to the prognosis of HCC. The validation cohort is derived from the TCGA liver hepatocellular carcinoma (LIHC) cohort of the Kaplan-Meier Plotter online analysis tool (http://kmplot.com/analysis/index.php?p=serviceandcancer=liver_rnaseq).18
Functional Enrichment Analysis
In order to understand the role of ACBD4 in HBV-related HCC, genome-wide expression profile matrix of GSE14520 cohort was applied to identify the differentially expressed genes (DEGs) between the high- and low-ACBD4 expression groups and genome-wide co-expressed genes of ACBD4 gene in tumor tissues. The screening of ACBD4 co-expression protein coding genes (PCGs) hinge on Pearson correlation coefficient. DEGs’ selection criteria are as follows: |log2 fold change (FC)|>1, P value less than 0.05, as well as false discovery rate (FDR) value less than 0.05, which were confirmed by limma package. After DEGs and co-expressed genes were obtained, Database for Annotation, Visualization and Integrated Discovery (DAVID) v6.8 (https://david.ncifcrf.gov/home.jsp) was used for gene ontology (G O) and Kyoto Encyclopedia of Genes and Genomes (KEGG) functional annotation. The biological function differences between the high- and low-ACBD4 expression groups were further explored by gene set enrichment analysis (GSEA: http://software.broadinstitute.org/gsea/index.jsp).
Drug Prediction and Immune Microenvironment Analysis
We further used Connectivity Map (CMap: https://portals.broadinstitute.org/cmap/) to identify compounds through the DEGs of ACBD4 in GSE14520 cohort HBV-related HCC.30,31 The structures of these compounds are downloaded from the PubChem (https://pubchem.ncbi.nlm.nih.gov) online tool, the drug-gene networks were downloaded from the STITCH online analysis tool.32,33 Immune microenvironment analysis in GSE14520 cohort was done using Estimation of STromal and Immune cells in MAlignant Tumours using Expression data (ESTIMATE) package in R platform, and the composition of stromal cells and immune cells in the immune microenvironment can be evaluated.34
Statistical Analysis
GSEA results criteria are as follows: |Normalized Enrichment Score (NES)|>1, P value less than 0.05 and FDR value less than 0.25. Binary logistic regression model was applied to estimate the relationship between SNPs and clinical parameters, and odds ratio (OR) was used to evaluate the risk ratio. P value less than 0.05 was identified as statistically significant difference. All statistical analysis used R version 4.0.2 and SPSS version 22.0.
Results
Survival Analysis
Through the GTEx database, we found that ACBD4 gene expression was the highest in liver tissue compared with other normal tissues of the human body (Figure 1). Subsequently, we used the GSE14520 database to explore the prognostic value of ACBD4 gene. Since the GSE14520 dataset contained two different batches of expression profile chip datasets, HCC cohort of the Affymetrix HT Human Genome U133A Array chip dataset containing most patients was included into this study to avoid batch effect. After excluding non-HBV-related HCC patients and patients without survival data, we got 212 HBV-related HCC tumor samples and 204 paracancerous samples were fitted into subsequent analysis. The baseline information of GSE14520 is summarized in Table S1. Investigation of GSE14520 HCC cohort revealed that ACBD4 mRNA expression is markedly up-regulated in adjacent paracancerous tissues (P <0.0001, Figure 2A). The receiver operating characteristic (ROC) curve suggests that the ACBD4 gene can be used to distinguish tumor from adjacent paracancerous tissues, the area under the curve (AUC) is 0.8932, and the 95% CI is 0.8601–0.9263 (Figure 2B). To verify the difference in expression of ACBD4 gene between tumor and paracancerous tissues of HCC, we found that expression of ACBD4 gene was markedly down-regulated in HCC tumor samples in all expression profile datasets of five validation cohorts (TCGA, GSE25097, GSE36376, GSE54236 and GSE64041) and ROC analysis indicated that ACBD4 was effective in differentiating HCC tumor from paracancerous tissues (Figure S1A–I). Survival analysis suggested that low-ACBD4 expression in HBV-related HCC patients leads to a poor overall survival (OS) (log-rank P = 0.0016, adjusted P = 0.017, adjusted HR = 0.561, 95% CI = 0.348–0.903, Figure 2C and D, Table 1). Subsequently, we used the GSE14520 HBV-related HCC cohort to perform a stratification analysis of ACBD in OS. Stratified analysis showed that ACBD gene was markedly related to HBV-related HCC OS in the following clinical subgroups, including male, age less than or equal to 60 years or greater than 60 years, tumor size less than or equal to 5 cm or greater than 5 cm, single tumor, cirrhosis, BCLC A stage, serum AFP less than or equal to 300 ng/mL (Figure 3). We performed a joint effect survival analysis in HBV-related HCC using the clinical parameters related to OS combined with ACBD gene in GSE14520. Joint effect survival analysis suggests that ACBD gene combined with tumor size, BCLC stage, serum AFP and cirrhosis can significantly distinguish HBV-related HCC subgroups with different prognostic phenotypes, respectively (Figure 4A–D, Table 2). The nomogram analysis of GSE14520 cohort suggests that the contribution of ACBD gene to HBV-related HCC OS was second only to BCLC and liver cirrhosis (Figure 5). Subsequently, survival analysis of ACBD was verified using the TCGA HCC cohort in the Kaplan-Meier Plotter website (Figure 6A–D). We observed that expression of ACBD gene was significantly related to OS (log-rank P = 0.0064, HR = 0.62, 95% CI = 0.44–0.88, Figure 6A), progression-free survival (PFS, log-rank P = 0.0096, HR = 0.68, 95% CI = 0.51–0.91, Figure 6B) and disease-specific survival (DSS, log-rank P = 0.014, HR = 0.57, 95% CI = 0.37–0.90, Figure 6D) in HCC patients of TCGA cohort.
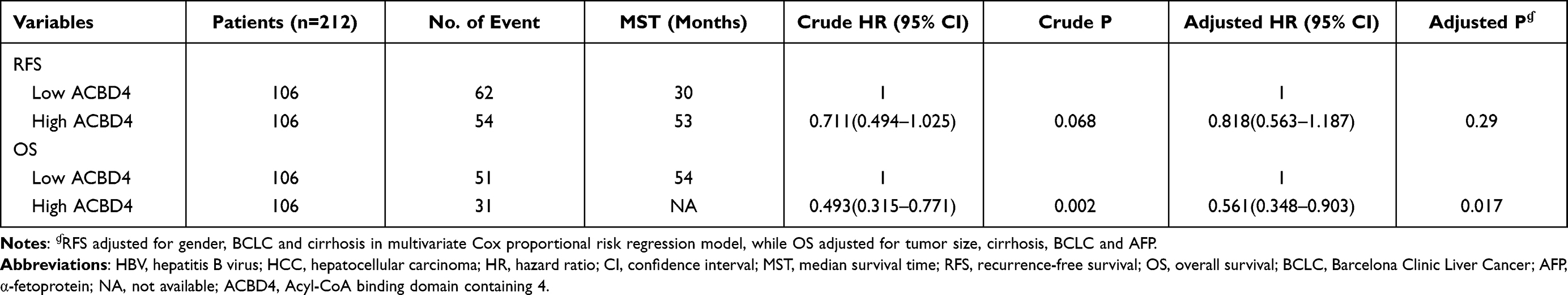 |
Table 1 Survival Analysis of ACBD4 Gene in GSE14520 HBV-Related HCC Cohort |
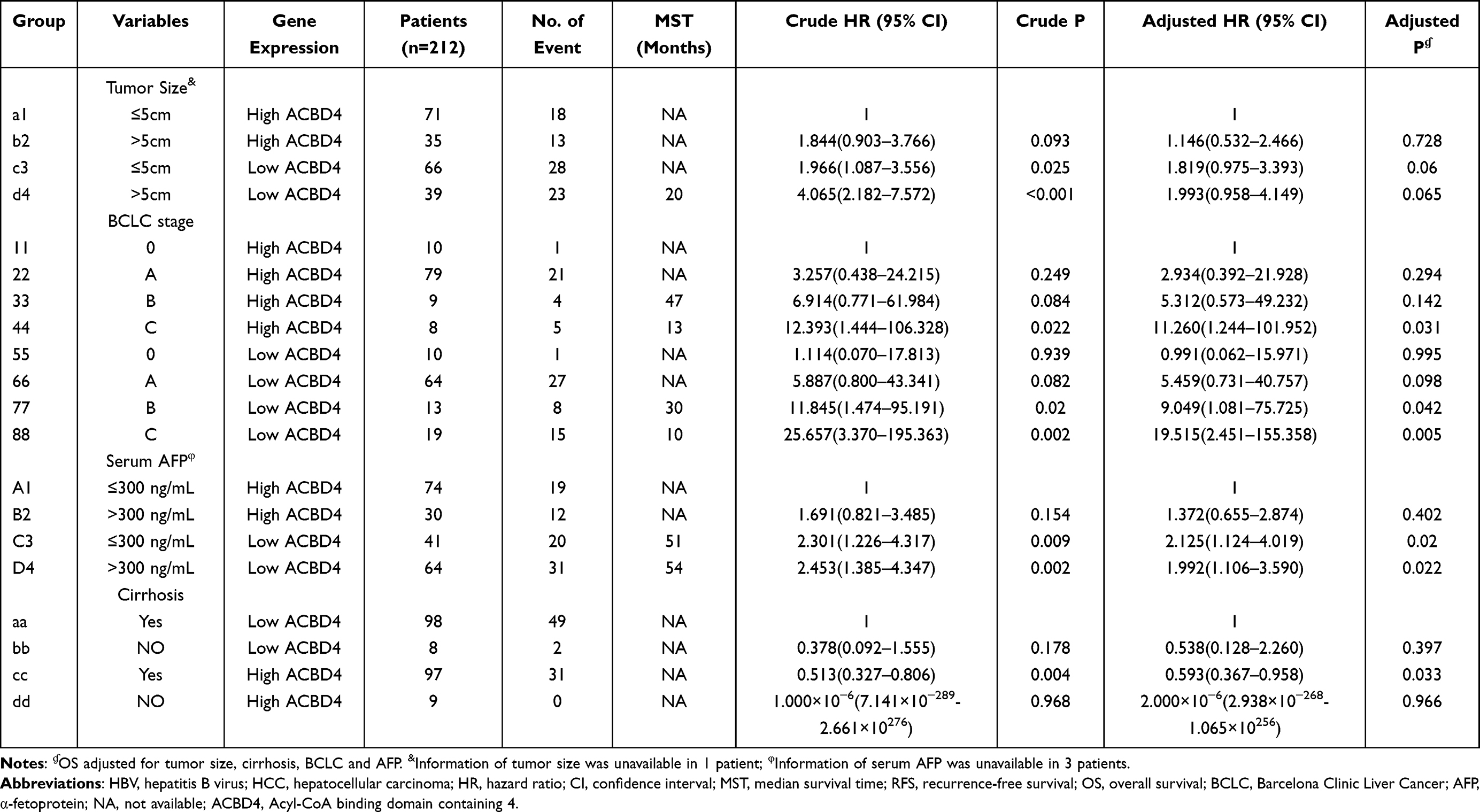 |
Table 2 Joint Effect Survival Analysis of ACBD4 Expression and Clinical Parameters in GSE14520 HBV-Related HCC Patients |
 |
Figure 1 Violin diagram of ACBD4 gene expression level distribution in normal human organ tissues from GTEx database. |
 |
Figure 2 The diagnostic and prognostic values of ACBD4 in GSE14520 HBV-related HCC cohort. (A) Expression distribution of ACBD4 in tumor and adjacent paracancerous tissues; (B) the ROC curve of ACBD4 gene in distinguished tumor and adjacent paracancerous tissues; (C) Kaplan-Meier curve of ACBD4 in HBV-related HCC recurrence-free survival time; (D) Kaplan-Meier curve of ACBD4 in HBV-related HCC overall survival time. |
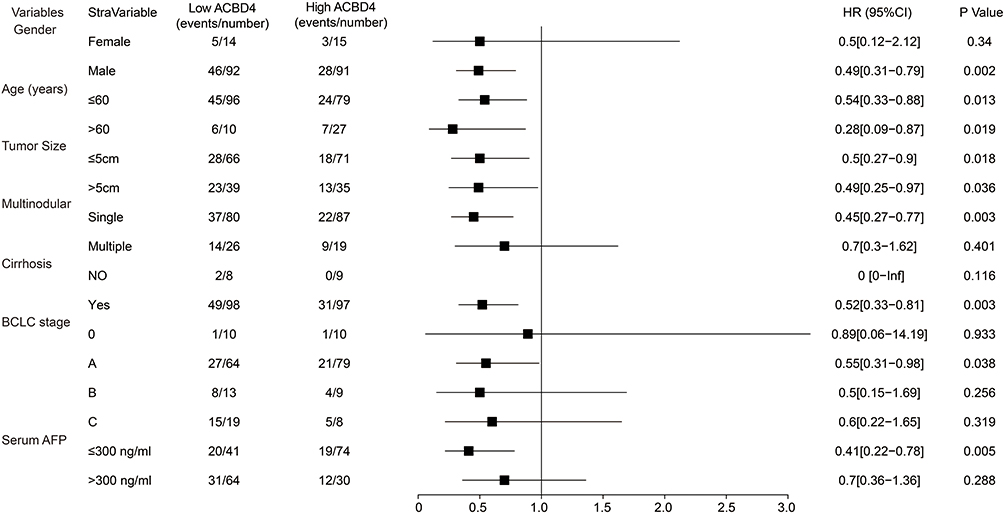 |
Figure 3 Stratified survival analysis results of ACBD4 gene in GSE14520 HBV-related HCC cohort. |
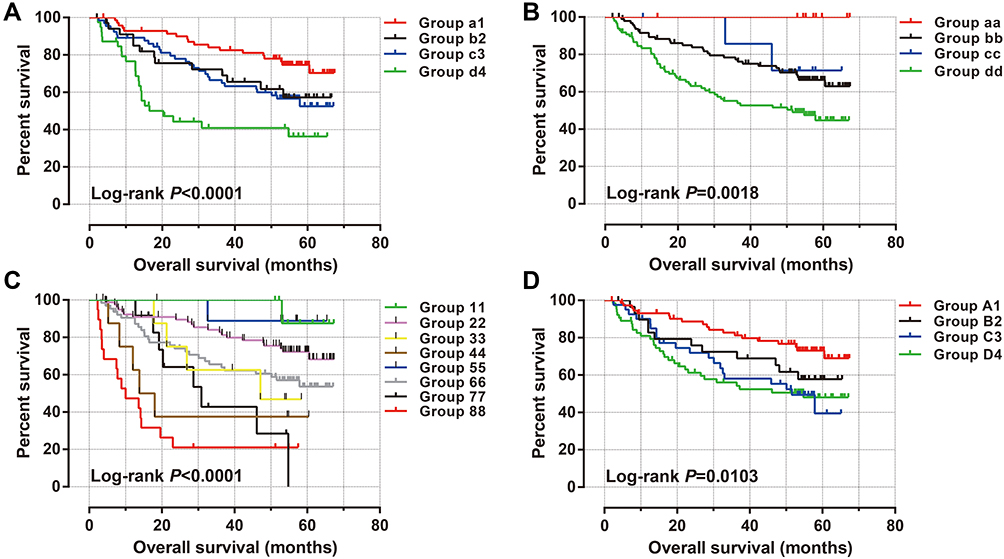 |
Figure 4 Joint effect survival analysis of ACBD4 expression and clinical parameters in GSE14520 HBV-related HCC patients. (A) Joint effect survival analysis of ACBD4 and tumor size; (B) Joint effect survival analysis of ACBD4 and cirrhosis; (C) Joint effect survival analysis of ACBD4 and BCLC stage; (D) Joint effect survival analysis of ACBD4 and serum AFP. Note: The detailed information on grouping is shown in Table 2. |
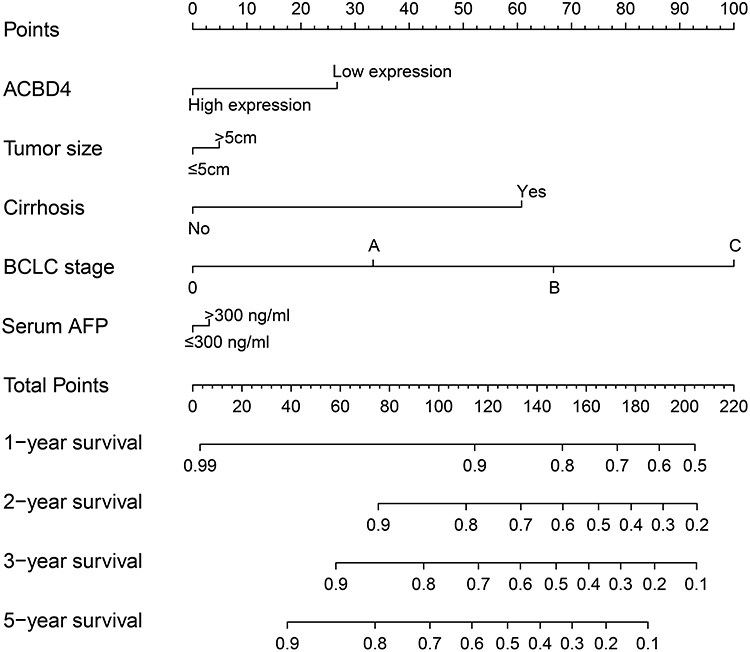 |
Figure 5 The nomogram of ACBD4 and clinical parameters in the GSE14520 HBV-related HCC cohort. |
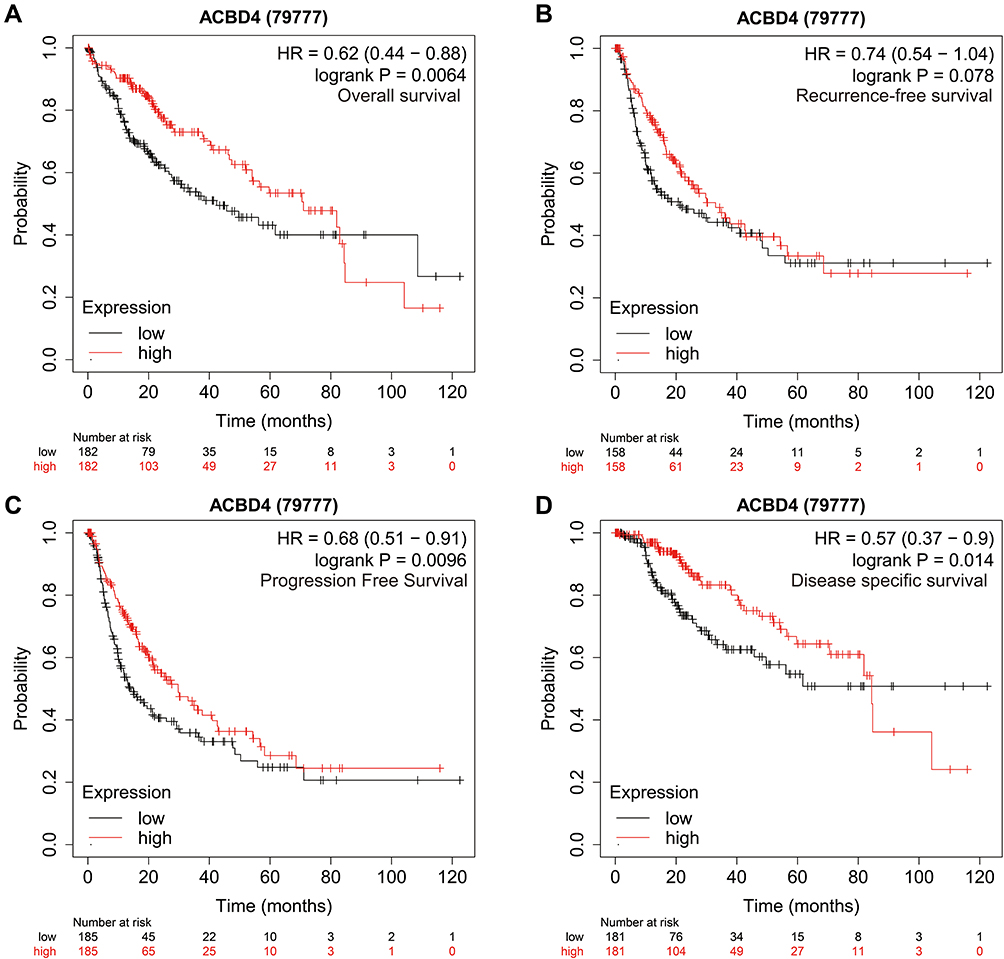 |
Figure 6 Prognostic value of ACBD4 gene in TCGA HCC cohort. (A) Kaplan-Meier curve of ACBD4 in HCC recurrence-free survival time; (B) Kaplan-Meier curve of ACBD4 in HCC overall survival time; (C) Kaplan-Meier curve of ACBD4 in HCC progression-free survival time; (D) Kaplan-Meier curve of ACBD4 in HCC disease-specific survival time. |
Clinical parameters of HBV-related HCC patients from The First Affiliated Hospital of Guangxi Medical University are shown in Table S2. Survival analysis revealed that patients with AG genotype of rs4986172 had better OS than those with GG genotype (adjusted P = 0.006, HR = 0.617, 95% CI = 0.438–0.870, Figure 7A, Table 3), while the difference in prognosis between AA genotype and GG genotype was not statistically significant. Subsequently, we combined patients with A allele (AA+AG) and found that HBV-related HCC patients with A allele had a better OS than those with GG genotype (log-rank P = 0.0015, adjusted P = 0.012, HR = 0.666, 95% CI = 0.486–0.913, Figure 7B, Table 3). We also compared the OS of AA genotype with that of HBV-related HCC patients carrying the G allele and found that the prognostic difference between the two groups was not statistically significant (log-rank P = 0.6165, adjusted P = 0.807, HR = 0.962, 95% CI = 0.705–1.312, Figure 7C, Table 3). Stratified prognostic analysis suggested that HBV-related HCC patients with A allele had better OS than those with GG genotype in each of the following subgroups: male, age less than or equal to 60 years, the Han nationality, both smoking or not, both drinking or not, body mass index (BMI) less than or equal to 25, serum alpha fetoprotein (AFP) less than or equal to 400 ng/mL, Child-Pugh A, BCLC C stage, single or multiple tumors, tumor size greater than 5 cm, cirrhosis, tumor with moderately differentiation, tumor with portal vein tumor thrombus (PVTT) and radical resection (Figure 8). Correlation analysis showed that GG genotype significantly increased the morbidity risk of BCLC C stage compared with patients with rs4986172 carrying the A allele (P = 0.032, OR = 1.707, 95% CI = 1.047–2.781, Table 4). In addition, GG genotype can significantly increase the morbidity risk of PVTT in HBV-related HCC patients (P = 0.012, OR = 1.944, 95% CI = 1.161–3.254, Table 4). We performed a joint effect survival analysis on these six clinical parameters significantly related to OS in the Guangxi HBV-related HCC cohort and the rs4986172 genotypes. We observed that the combination of rs4986172 genotypes and prognostic-related clinical parameters can more accurately distinguish HBV-related HCC patients with different prognostic subgroups (Figure 9A–F, Table 5). The nomogram indicates that contribution of rs4986172 genotypes to HBV-related HCC OS is second only to tumor size, PVTT and BCLC stage (Figure 10A and B).
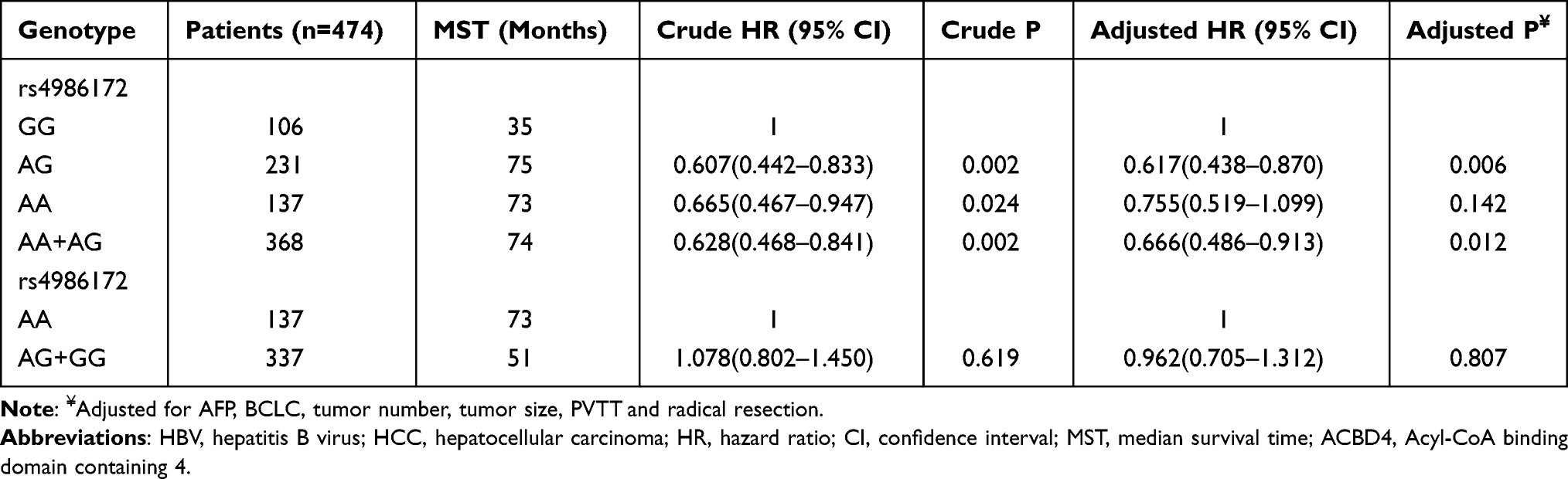 |
Table 3 Survival Analysis of ACBD4-rs4986172 in Guangxi HBV-Related HCC Patients |
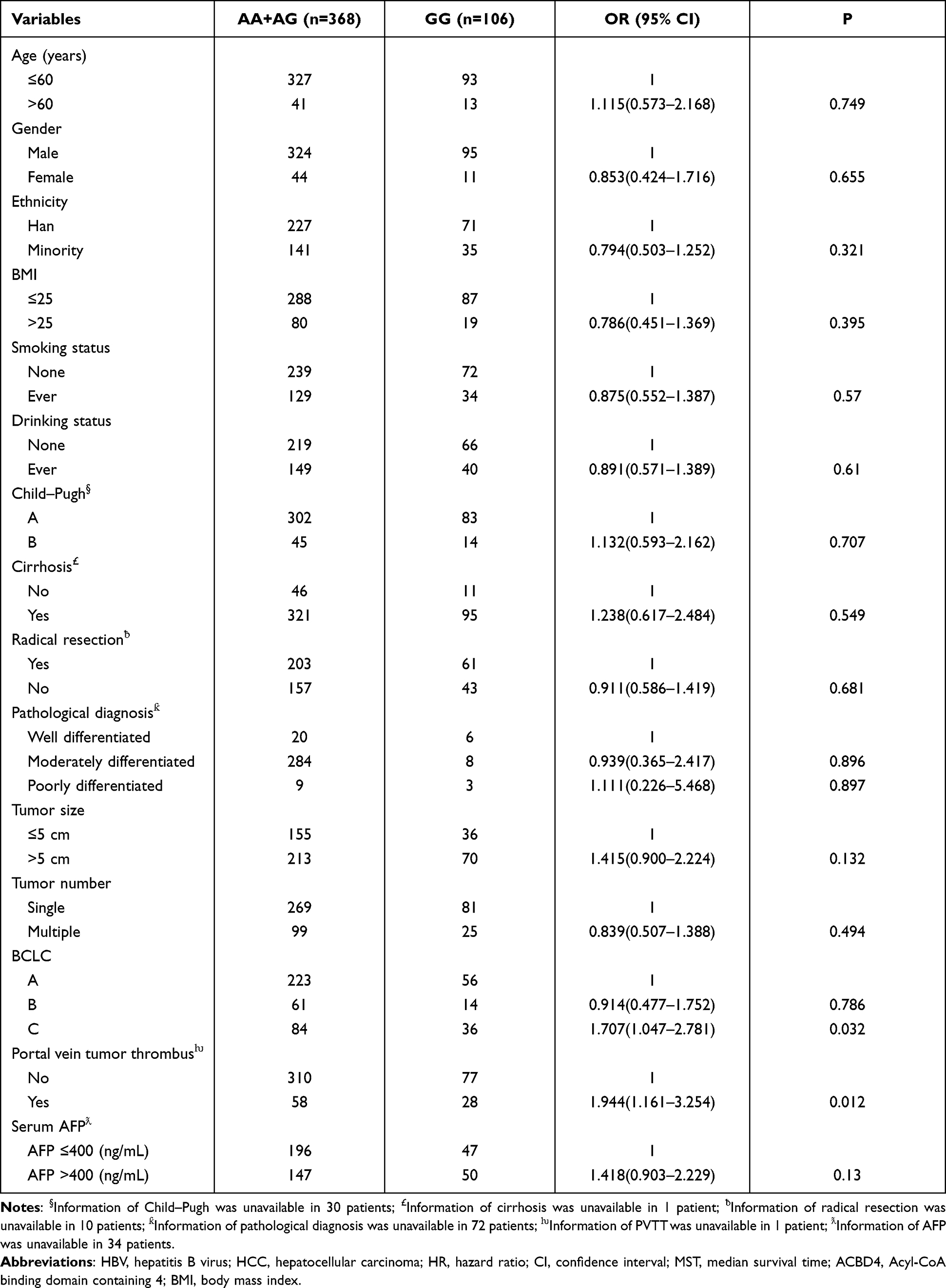 |
Table 4 Correlation Analysis Between ACBD4-rs4986172 and Clinical Parameters in Guangxi HBV-Related HCC Patients |
 |
Table 5 Joint Effect Survival Analysis of ACBD4-rs4986172 and Clinical Parameters in Guangxi HBV-Related HCC Patients |
 |
Figure 7 Prognostic value of ACBD4-rs4986172 in Guangxi HBV-related HCC cohort. (A) Kaplan-Meier curve of AA, AG and GG; (B) Kaplan-Meier curve of AA+AG and GG; (C) Kaplan-Meier curve of AA and AG+GG. |
 |
Figure 8 Stratified survival analysis results of ACBD4-rs4986172 in Guangxi HBV-related HCC cohort. |
 |
Figure 9 Joint effect survival analysis of ACBD4-rs4986172 and clinical parameters in Guangxi HBV-related HCC patients. (A) Joint effect survival analysis of rs4986172 and serum AFP; (B) Joint effect survival analysis of rs4986172 and BCLC stage; (C) Joint effect survival analysis of rs4986172 and PVTT; (D) Joint effect survival analysis of rs4986172 and radical resection. (E) Joint effect survival analysis of rs4986172 and tumor size; (F) Joint effect survival analysis of rs4986172 and tumor number. Note: The detailed information on grouping is shown in Table 5. |
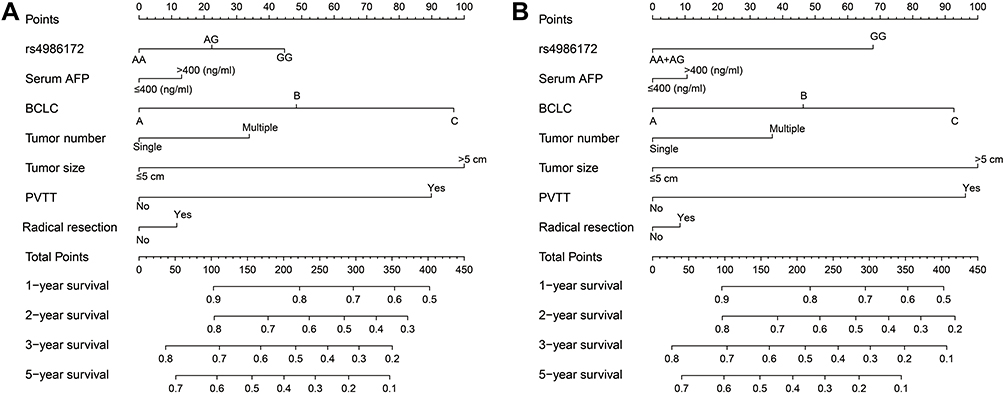 |
Figure 10 The nomogram of ACBD4-rs4986172 and clinical parameters in the Guangxi HBV-related HCC cohort. (A) AA, AG and GG genotypes in HBV-related HCC; (B) AA+AG and GG genotypes in HBV-related HCC. |
Functional Enrichment Analysis
To further understand the molecular mechanisms of ACBD4 gene in HBV-related HCC, three bioinformatics analysis methods, including whole genome co-expression analysis, DEG analysis and GSEA, were applied to comprehensively investigate the mechanism of ACBD4 gene. Co-expressed genes of ACBD4 are defined as Pearson correlation coefficient |r|≥0.4, and P value less than 0.05. Through whole genome co-expressed gene screening, we got 573 ACBD4 co-expressed genes in HBV-related HCC, of which 170 genes had negative correlation and 403 genes had positive correlation (Figure 11, Table S3). GO term enrichment suggests that ACBD4 and its co-expressed genes markedly participate in the following biological functions: omega-hydroxylase P450 pathway, epoxygenase P450 pathway, focal adhesion, mitotic nuclear division, liver development, DNA damage response, signal transduction by p53 class mediator resulting in cell cycle arrest, drug metabolic process, exogenous drug catabolic process, Hsp90 protein binding and other biological processes or functions (Table S4). KEGG analysis of ACBD4 and its co-expressed genes suggests that these genes are significantly involved in metabolic pathways, chemical carcinogenesis, peroxisome proliferator-activated receptor (PPAR) signaling pathway, adenosine monophosphate-activated protein kinase (AMPK) signaling pathway and other amino acid metabolism pathways (Table S5).
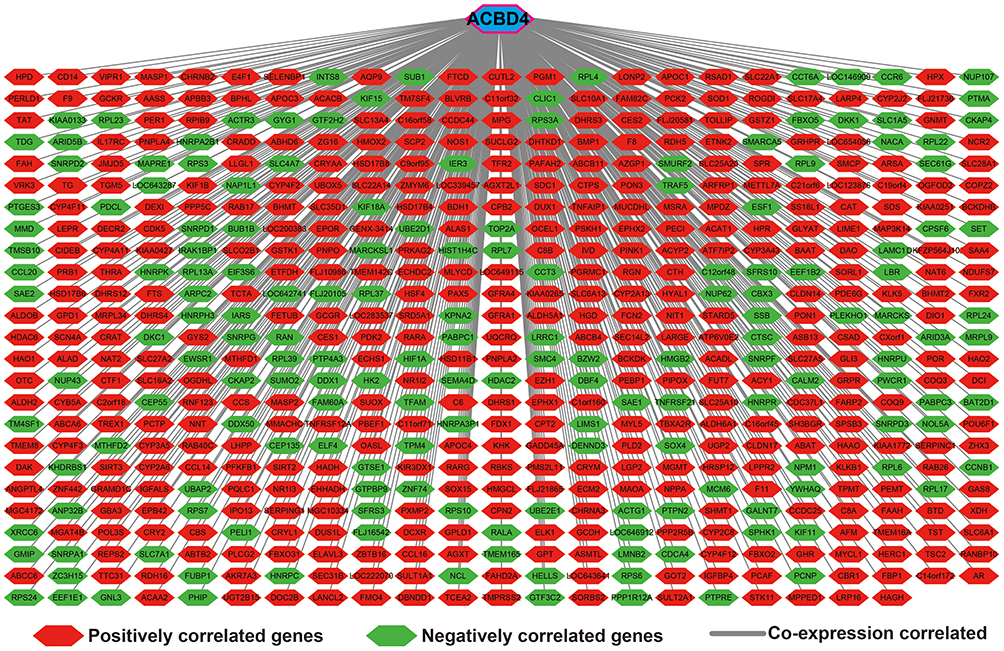 |
Figure 11 Genome-wide co-expression network of ACBD4 gene in HBV-related HCC tumor tissues of patients in GSE14520 cohort. |
DEGs between different ACBD4 expression groups suggests that the ACBD4 gene differed significantly between the two groups (log2FC = −1.25, P = 3.32×10−48, FDR = 4.33×10−44, Table S6). In addition, we identified 157 DEGs that were markedly down-regulated in HCC tumor samples with low ACBD4 expression, and 42 DEGs that were significantly up-regulated (Figure 12, Figure S2 and Table S). Functional enrichment of GO term implies that these DEGs can be markedly enriched in the following biological processes: drug metabolic process, epoxygenase P450 pathway, omega-hydroxylase P450 pathway, exogenous drug catabolic process, glucose metabolic process, cellular response to tumor necrosis factor and cellular response to interleukin-1 (Table S7). KEGG analysis suggests that DEGs of ACBD4 gene can be markedly enriched in the following pathways: metabolic pathways, chemical carcinogenesis, metabolism of xenobiotics by cytochrome P450, PPAR signaling pathway, drug metabolism-other enzymes, drug metabolism-cytochrome P450, AMPK signaling pathway, adipocytokine signaling pathway and glucagon signaling pathway (Table S8).
 |
Figure 12 Volcano plot of DEGs between low- and high-ACBD4 expression groups. |
GSEA was also used to further investigate the biological functional mechanisms that differed in patients in high- and low-ACBD4 expression groups. Using C2 (c2.all.v7.2.symbols.gmt) reference gene set base on the GSE14520 whole genome dataset, we found that ACBD4 gene plays a role in HBV-related HCC by participating in the regulation of the following signaling pathways: nuclear receptor transcription pathway, mTOR signaling up, liver cancer recurrence down, liver cancer poor survival down, PPAR signaling pathway, metabolism cytochrome P450, oxidation by cytochrome P450, bile acid and bile salt metabolism, stress pathway, fatty acid metabolism, tumor invasiveness up, and metastasis up (Figure 13A–L). Using C5 (c5.all.v7.2.symbols.gmt) reference gene set, we revealed that ACBD4 functions take part in the following biological processes: fatty acid metabolic process, polysaccharide catabolic process, bile acid metabolic process, alcohol catabolic process, lipid accumulation in hepatocytes, positive regulation of G protein coupled receptor signaling pathway, lipoprotein metabolic process, steroid metabolic process, bile acid and bile salt transport, drug metabolic process, and abnormality of hepatobiliary system physiology (Figure 14A–L).
 |
Figure 13 GSEA results between low- and high-ACBD4 expression groups by using c2 reference gene set. (A) nuclear receptor transcription pathway; (B) mTOR signaling up; (C) liver cancer recurrence down; (D) liver cancer poor survival down; (E) PPAR signaling pathway; (F) metabolism cytochrome P450; (G) oxidation by cytochrome P450; (H) bile acid and bile salt metabolism; (I) stress pathway; (J) fatty acid metabolism; (K) tumor invasiveness up; (L) metastasis up. |
 |
Figure 14 GSEA results between low- and high-ACBD4 expression groups by using c5 reference gene set. (A) fatty acid metabolic process; (B) polysaccharide catabolic process; (C) bile acid metabolic process; (D) alcohol catabolic process; (E) lipid accumulation in hepatocytes; (F) positive regulation of G protein coupled receptor signaling pathway; (G) fatty acid metabolic process; (H) lipoprotein metabolic process; (I) steroid metabolic process; (J) bile acid and bile salt transport; (K) drug metabolic process; (L) abnormality of hepatobiliary system physiology. |
Drug Prediction and Immune Microenvironment Analysis
We further used CMap to explore the potential targeting drugs of ACBD4 gene in HBV-related HCC. We have obtained a total of eight compounds that may be served as targeted drugs for ACBD4 gene in HBV-related HCC (Figure 15A–I), all of the mean connective score of compounds were less than −0.6 and P<0.05. Through STITCH, we also constructed a drug-gene interaction network of these compounds. The gene interacting with bephenium hydroxynaphthoate cannot be found in STITCH, while 4,5-dianilinophthalimide cannot be identified. Therefore, we finally constructed a drug-gene interaction network of the other six compounds. In this network, we found that some DEGs between the high- and low-ACBD4 expression groups interacted with these drugs (Figure 16). Differentially expressed genes KNG1 and NTS interact with DL-thiorphan, while ABAT, AGXT2L1 and RDH16 interact with apramycin. We also found that ABCB1, ABCB4, CYP2A6, TOP2A, CYP3A4, CYP2C9, SLC22A1, CYP2J2 and CYP2C8 interact with quinidine. ABCA8, ABCB1, SLCO1B1, SLC17A4, ABCA6 and ABCB4 interact with Aminohippuric acid. ABCB1, ABCB4, UGT1A4, UGT1A6, SLC1A1 and UGT2B15 interact with scopoletin. SLC1A1 interacts with alphaxalone. We further analyzed the relationship between ACBD4 gene and immune microenvironment. We found that there were significant differences in the immune score and ESTIMATE score between different ACBD4 expression subgroups, and the immune score and ESTIMATE score of HBV-related HCC patients with high-ACBD4 expression were markedly decreased (Figure 17). We did not observe a significant difference in stroma scores between high- and low-ACBD4 expression groups (Figure 17).
 |
Figure 15 CMap result of ACBD4 in HCC. (A) Chemical construction of scopoletin; (B) Chemical construction of alfaxalone; (C) Chemical construction of bephenium hydroxynaphthoate; (D) Chemical construction of apramycin; (E) Chemical construction of 4,5-dianilinophthalimide; (F) Chemical construction of DL-thiorphan; (G) Chemical construction of aminohippuric acid; (H) Chemical construction of quinidine; (I) Summary table of CMap results. |
 |
Figure 16 Drug-gene interaction network of compounds identified by CMap. |
 |
Figure 17 Score of ACBD4 gene in HBV-HCC immune microenvironment. |
Discussion
Through a literature search, we learned that the functional mechanism of ACBD4 gene in cancers is rarely reported. A previous study only found that ACBD4 gene produces a tumor suppressor effect as a target gene of p53 in colorectal cancer cell line.9 Based on the findings of this study, we know that ACBD4 gene has the highest expression in liver tissue among all normal human organ tissues. Therefore, we speculate that the main place of ACBD4 gene function in human body is liver tissue. We also observed significant differences in the distribution of ACBD4 gene between HBV-related HCC tumor and paracancerous tissues, and significant down-regulation in tumor tissues. We also performed a comprehensive prognostic analysis and found that ACBD4 expression and rs4986172 were significantly related to the OS of HBV-related HCC. Our study contributes to the exploration of the underlying clinical value of ACBD4 gene in HCC, and provides new evidence and theoretical basis for the application of ACBD4 gene in cancers. The findings of this study have important implications for translational medicine.
In this study, we screened DEGs and performed GSEA analysis between high- and low-ACBD4 expression subgroups, and genome-wide co-expression analysis of ACBD4 gene in HBV-related HCC tumor samples. We also found that ACBD4 gene was significantly associated with metabolic pathways and cytochrome P450 pathways, which is consistent with our previous understanding of the function of ACBD4 gene as a peroxisomal protein. We all know that P450 is an important enzyme in liver metabolism and is also an important enzyme related to HCC.35–38
In this study, we also identified eight targeted drugs of ACBD4 gene in HBV-related HCC. Through a review of the literature, we found that among the eight drugs, except bephenium hydroxynaphthoate and apramycin, the remaining six drugs have all been reported to play a certain role in cancers. Tian et al. revealed that scopoletin has an anti-cancer effect in human cervical cancer cell lines, which can inhibit the proliferation, invasion and cell cycle arrest of HeLa cells.39 Yuan et al. demonstrated the anti-cancer effect of lung cancer through the rat sarcoma virus (Ras)/rapidly accelerated fibrosarcoma (Raf)/mitogen-activated protein kinase (MEK)/extracellular signal-regulated kinase (ERK) pathway and the phosphatidylinositol 3-kinase (PI3K)// Akt pathway through network pharmacology and in vivo and in vitro experiments.40 Scopolamine can induce apoptosis of tumor lymphocytes, but can induce proliferation of normal lymphocytes.41 Genomics revealed that the activation of nuclear factor-kappa B (NF-κB) may be a key element of scopoletin resistance in cancers.42 Alfaxalone has been reported as an anticancer agent for glioma, inhibiting the migration and invasion of glioma cells without cytotoxicity.43 4,5-dianilinophthalimide is a new class of tyrosine kinase, epidermal growth factor receptor inhibitors. Previous studies suggest that dianilinophthalimide plays an anticancer effect in human transitional cell carcinoma and human A431 epidermoid carcinoma cells.44,45 Wu et al. initially identified DL-thiorphan as a potential targeted therapeutic agent for lung cancer based on GEO whole-genome expression profile datasets (GSE18842, GSE30219, GSE31210, GSE32863 and GSE40791) and CMAP analysis.46 Meanwhile, based on the similar bioinformatics analysis approach, DL-thiorphan is also considered as a possible candidate drug for type 2 diabetes, with an influence of reversing type 2 diabetes.47 Aminohippuric acid is a diagnostic drug, the study of Funai et al. suggests that Aminohippuric acid can be used as a diagnostic marker in lung cancer patients’ urine.48 P-glycoprotein (P-gp) is considered to be a key agent of multidrug resistance, and inhibition of P-gp in multidrug resistant cancers can be achieved by reverse targeting of quinidine-PEG conjugates, thereby improving tumor resistance.49 Quinidine has been used as a resistance medicament for epirubicin-resistant breast cancer patients, but clinical trials have shown that quinidine does not markedly alter toxicity, response rate, or survival after epirubicin chemotherapy in advanced breast cancer patients.50,51 However, there is a study suggesting that quinidine can exert a tumor suppressor effect by regulating the expression level of Myc protein.52 Quinidine, as a voltage-gated K+ channel blocker, inhibits the proliferation of glioma cell and induces cell apoptosis with a dose-dependent mode by misregulating multiple microRNA in glioma cell lines.53 The mechanism of quinidine antitumor cell proliferation in glioma may be through the inhibition of ODC activity.54 Quinidine can inhibit the proliferation of neurofibromatosis type 2-deficient human malignant mesothelioma cells by blocking the cell cycle, thus exhibiting the anticancer effect.55 Quinidine can reverse multidrug resistance of uterine sarcoma cells and enhance paclitaxel-induced cytotoxicity and apoptosis.56 In reviewing the above studies, we have not found any reports about the functions of these eight drugs screened in this study in HCC. However, many of them have anti-tumor effects in other cancers.
Through three bioinformatics function enrichment analysis pathways, we found that ACBD4 gene is closely related to a variety of metabolic pathways and P450 pathways, and that ACBD4 gene may play a role in HCC through PPAR pathway. PPAR regulates lipid metabolism, fat formation, expression of genes that maintain metabolic homeostasis and inflammation, and induces anticancer effects in a variety of human cancers, especially HCC.57 PPARs participates in the colonization and adaptation of intestinal microbes, has the function of regulating the intestinal flora, and has certain potential in the diagnosis and treatment of HCC.58 PPARγ plays an essential role in the PPAR signaling pathway. Overexpression of PPARγ in HCC cell lines can inhibit the proliferation, invasion and angiogenesis of HCC cells, and exert a tumor suppressor effect.59,60 At the same time, PPARγ is also regulated by miRNA, thus affecting the invasion and migration of HCC.61 The HCC clinical cohort analysis suggests that PPARγ is closely related to the MVI of HCC.60 PPARα, another key gene in the PPAR pathway, is significantly associated with poor prognosis, tumor size, TMN stage and vascular invasion in HCC, and can be used as a prognostic marker for HCC.62 In summary, the anticancer mechanism of PPAR signaling pathway in HCC has been clarified in previous studies. In this study, ACBD4 gene was preliminarily identified as a tumor suppressor gene of HCC, and the possible molecular mechanism was that ACBD4 exerted a tumor suppressor effect through PPAR signaling pathway. There has been increasing evidence that the HCC immune microenvironment is closely related to the prognosis of HCC.63,64 In this study, we found that HBV-related HCC patients with different ACBD4 expression levels have significant differences in tumor immune microenvironment, and their prognosis is also significantly different. We have reason to believe that the difference in prognosis between the two groups of HBV-related HCC patients may be influenced to some extent by the tumor immune microenvironment. This suggests that ACBD4 may be a potential target for HCC immunotherapy.
There are certain deficiencies in this study that need to be clarified. First of all, the main population included in our study was HCC patients with HBV infection who underwent HCC surgical resection, so the main conclusions of our study are only applicable to this subgroup of patients. Secondly, our current study has explored the biological function and molecular mechanism of ACBD4 gene in HBV-related HCC using a variety of bioinformatics approaches, but these findings lack in vivo and in vitro experimental verification. Thirdly, the sample size of our study population is not large enough, and the prognostic value of ACBD4 gene in HBV-related HCC still needs to be verified in future multi-center and large-sample studies. Although this study has some limitations, we are reporting for the first time that ACBD4 gene is dysregulated in liver cancer tissues, and it can be used as a prognostic marker of HCC in clinical application. At the same time, we also used the whole genome dataset to make preliminary identification of the potential function and mechanism of ACBD4 gene in HCC.
Conclusions
In conclusion, the present study reveals that ACBD4 gene was significantly expressed in normal human liver tissues, and was markedly down-regulated in HBV-related HCC tumor tissues. Survival analysis using GSE14520 cohort suggested that high ACBD4 expression was markedly related to poor OS in HBV-related HCC. The same result can also be verified in TCGA HCC cohort. The OS of rs4986172 GG genotype was worse than that of HCC patients with A allele (AA and AG genotypes). Functional enrichment analysis suggests that ACBD4 gene is involved in multiple metabolic, PPAR and cytochrome P450 pathways in HBV-related HCC. We also identified eight compounds that may be used as targeted therapeutic agents for ACBD4 gene in HBV-related HCC; these compounds were scopoletin, alfaxalone, bephenium hydroxynaphthoate, apramycin, 4,5-dianilinophthalimide, DL-thiorphan, aminohippuric acid and quinidine. Immune microenvironment analysis revealed that there were significant differences in immune and ESTIMATE scores of HBV-related HCC tumor tissues with different ACBD4 expression levels. As a result of some limitations in current study, our findings also require verification in future studies.
Abbreviations
ACBD4, Acyl-CoA binding domain containing 4; SNP, single nucleotide polymorphism; MST, median survival time; RFS, recurrence-free survival; OS, overall survival; HR, hazard ratio; CI, confidence interval; HBV, hepatitis B virus; HCC, hepatocellular carcinoma; TCGA, The Cancer Genome Atlas; AFP, α-fetoprotein; GO, gene ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; ROC, receiver operating characteristic; FDR, false discovery rate; GSEA, gene set enrichment analysis; DEGs, differentially expressed genes.
Acknowledgments
We acknowledge the support by the National Key Clinical Specialty Programs (General Surgery and Oncology) and the Key Laboratory of Early Prevention & Treatment for Regional High-Incidence-Tumor (Guangxi Medical University), Ministry of Education, China. The authors also thank the contributors of The Cancer Genome Atlas, GSE14520, GSE25097, GSE36376, GSE54236, and GSE64041 for sharing the HCC dataset on open access. In addition, we would like to acknowledge the helpful comments on this article received from our reviewers.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. Huasheng Huang and Xiwen Liao are co-first authors for this study.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108. doi:10.3322/caac.21262
2. Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66(2):115–132. doi:10.3322/caac.21338
3. Xu L, Qian G, Tang L, Su J, Wang JS. Genetic variations of hepatitis B virus and serum aflatoxin-lysine adduct on high risk of hepatocellular carcinoma in Southern Guangxi, China. J Hepatol. 2010;53(4):671–676. doi:10.1016/j.jhep.2010.04.032
4. Qi LN, Li LQ, Chen YY, et al. Genome-wide and differential proteomic analysis of hepatitis B virus and aflatoxin B1 related hepatocellular carcinoma in Guangxi, China. PLoS One. 2013;8(12):e83465. doi:10.1371/journal.pone.0083465
5. Zhou M, Wang H, Zeng X, et al. Mortality, morbidity, and risk factors in China and its provinces, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet. 2019;394(10204):1145–1158. doi:10.1016/S0140-6736(19)30427-1
6. Bishop A, Kamoshita M, Passmore JB, et al. Fluorescent tools to analyse peroxisome-ER interactions in mammalian cells. Contact. 2019;2:2515256419848641. doi:10.1177/2515256419848641
7. Costello JL, Castro IG, Schrader TA, Islinger M, Schrader M. Peroxisomal ACBD4 interacts with VAPB and promotes ER-peroxisome associations. Cell Cycle. 2017;16(11):1039–1045. doi:10.1080/15384101.2017.1314422
8. Adriaens ME, Lodder EM, Moreno-Moral A, et al. Systems genetics approaches in rat identify novel genes and gene networks associated with cardiac conduction. J Am Heart Assoc. 2018;7(21):e009243. doi:10.1161/JAHA.118.009243
9. Liao JM, Zeng SX, Zhou X, Lu H. Global effect of inauhzin on human p53-responsive transcriptome. PLoS One. 2012;7(12):e52172. doi:10.1371/journal.pone.0052172
10. Consortium GT, Thomas J, Salvatore M. The Genotype-Tissue Expression (GTEx) project. Nat Genet. 2013;45(6):580–585. doi:10.1038/ng.2653
11. Sun Y, Ji F, Kumar MR, et al. Transcriptome integration analysis in hepatocellular carcinoma reveals discordant intronic miRNA-host gene pairs in expression. Int J Biol Sci. 2017;13(11):1438–1449. doi:10.7150/ijbs.20836
12. Lu Y, Xu W, Ji J, et al. Alternative splicing of the cell fate determinant Numb in hepatocellular carcinoma. Hepatology. 2015;62(4):1122–1131. doi:10.1002/hep.27923
13. Chen S, Fang H, Li J, et al. Microarray analysis for expression profiles of lncRNAs and circRNAs in rat liver after brain-dead donor liver transplantation. Biomed Res Int. 2019;2019:5604843. doi:10.1155/2019/5604843
14. Zhao X, Parpart S, Takai A, et al. Integrative genomics identifies YY1AP1 as an oncogenic driver in EpCAM(+) AFP(+) hepatocellular carcinoma. Oncogene. 2015;34(39):5095–5104. doi:10.1038/onc.2014.438
15. Roessler S, Long EL, Budhu A, et al. Integrative genomic identification of genes on 8p associated with hepatocellular carcinoma progression and patient survival. Gastroenterology. 2012;142(4):957–966 e912. doi:10.1053/j.gastro.2011.12.039
16. Roessler S, Jia HL, Budhu A, et al. A unique metastasis gene signature enables prediction of tumor relapse in early-stage hepatocellular carcinoma patients. Cancer Res. 2010;70(24):10202–10212. doi:10.1158/0008-5472.CAN-10-2607
17. Wang Y, Gao B, Tan PY, et al. Genome-wide CRISPR knockout screens identify NCAPG as an essential oncogene for hepatocellular carcinoma tumor growth. FASEB J. 2019;33(8):8759–8770. doi:10.1096/fj.201802213RR
18. Menyhart O, Nagy A, Gyorffy B. Determining consistent prognostic biomarkers of overall survival and vascular invasion in hepatocellular carcinoma. R Soc Open Sci. 2018;5(12):181006. doi:10.1098/rsos.181006
19. Sung WK, Zheng H, Li S, et al. Genome-wide survey of recurrent HBV integration in hepatocellular carcinoma. Nat Genet. 2012;44(7):765–769. doi:10.1038/ng.2295
20. Srivastava S, Wong KF, Ong CW, et al. A morpho-molecular prognostic model for hepatocellular carcinoma. Br J Cancer. 2012;107(2):334–339. doi:10.1038/bjc.2012.230
21. Ivanovska I, Zhang C, Liu AM, et al. Gene signatures derived from a c-MET-driven liver cancer mouse model predict survival of patients with hepatocellular carcinoma. PLoS One. 2011;6(9):e24582. doi:10.1371/journal.pone.0024582
22. Lim HY, Sohn I, Deng S, et al. Prediction of disease-free survival in hepatocellular carcinoma by gene expression profiling. Ann Surg Oncol. 2013;20(12):3747–3753. doi:10.1245/s10434-013-3070-y
23. Cho YA, Choi S, Park S, Park CK, Ha SY. Expression of pregnancy up-regulated non-ubiquitous calmodulin kinase (PNCK) in hepatocellular carcinoma. Cancer Genomics Proteomics. 2020;17(6):747–755. doi:10.21873/cgp.20229
24. Ha SY, Sohn I, Hwang SH, Yang JW, Park CK. The prognosis of hepatocellular carcinoma after curative hepatectomy in young patients. Oncotarget. 2015;6(21):18664–18673. doi:10.18632/oncotarget.4330
25. Dituri F, Scialpi R, Schmidt TA, et al. Proteoglycan-4 is correlated with longer survival in HCC patients and enhances sorafenib and regorafenib effectiveness via CD44 in vitro. Cell Death Dis. 2020;11(11):984. doi:10.1038/s41419-020-03180-8
26. Zubiete-Franco I, Garcia-Rodriguez JL, Lopitz-Otsoa F, et al. SUMOylation regulates LKB1 localization and its oncogenic activity in liver cancer. EBioMedicine. 2019;40:406–421. doi:10.1016/j.ebiom.2018.12.031
27. Villa E, Critelli R, Lei B, et al. Neoangiogenesis-related genes are hallmarks of fast-growing hepatocellular carcinomas and worst survival. Results from a prospective study. Gut. 2016;65(5):861–869. doi:10.1136/gutjnl-2014-308483
28. Makowska Z, Boldanova T, Adametz D, et al. Gene expression analysis of biopsy samples reveals critical limitations of transcriptome-based molecular classifications of hepatocellular carcinoma. J Pathol Clin Res. 2016;2(2):80–92. doi:10.1002/cjp2.37
29. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017;45(W1):W98–W102. doi:10.1093/nar/gkx247
30. Lamb J, Crawford ED, Peck D, et al. The Connectivity Map: using gene-expression signatures to connect small molecules, genes, and disease. Science. 2006;313(5795):1929–1935. doi:10.1126/science.1132939
31. Lamb J. The connectivity map: a new tool for biomedical research. Nat Rev Cancer. 2007;7(1):54–60. doi:10.1038/nrc2044
32. Szklarczyk D, Santos A, von Mering C, Jensen LJ, Bork P, Kuhn M. STITCH 5: augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acids Res. 2016;44(D1):D380–384. doi:10.1093/nar/gkv1277
33. Kuhn M, von Mering C, Campillos M, Jensen LJ, Bork P. STITCH: interaction networks of chemicals and proteins. Nucleic Acids Res. 2008;36(Databaseissue):D684–688. doi:10.1093/nar/gkm795
34. Yoshihara K, Shahmoradgoli M, Martinez E, et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat Commun. 2013;4:2612. doi:10.1038/ncomms3612
35. Nekvindova J, Mrkvicova A, Zubanova V, et al. Hepatocellular carcinoma: gene expression profiling and regulation of xenobiotic-metabolizing cytochromes P450. Biochem Pharmacol. 2020;177:113912. doi:10.1016/j.bcp.2020.113912
36. Liu ZZ, Yan LN, Dong CN, et al. Cytochrome P450 family members are associated with fast-growing hepatocellular carcinoma and patient survival: an integrated analysis of gene expression profiles. Saudi J Gastroenterol. 2019;25(3):167–175. doi:10.4103/sjg.SJG_290_18
37. Wang JF, Chou KC. Molecular modeling of cytochrome P450 and drug metabolism. Curr Drug Metab. 2010;11(4):342–346. doi:10.2174/138920010791514180
38. Almazroo OA, Miah MK, Venkataramanan R. Drug Metabolism in the Liver. Clin Liver Dis. 2017;21(1):1–20. doi:10.1016/j.cld.2016.08.001
39. Tian Q, Wang L, Sun X, Zeng F, Pan Q, Xue M. Scopoletin exerts anticancer effects on human cervical cancer cell lines by triggering apoptosis, cell cycle arrest, inhibition of cell invasion and PI3K/AKT signalling pathway. J BUON. 2019;24(3):997–1002.
40. Yuan C, Wang MH, Wang F, et al. Network pharmacology and molecular docking reveal the mechanism of Scopoletin against non-small cell lung cancer. Life Sci. 2021;270:119105. doi:10.1016/j.lfs.2021.119105
41. Manuele MG, Ferraro G, Barreiro Arcos ML, Lopez P, Cremaschi G, Anesini C. Comparative immunomodulatory effect of scopoletin on tumoral and normal lymphocytes. Life Sci. 2006;79(21):2043–2048. doi:10.1016/j.lfs.2006.06.045
42. Seo EJ, Saeed M, Law BY, et al. Pharmacogenomics of scopoletin in tumor cells. Molecules. 2016;21(4):496. doi:10.3390/molecules21040496
43. Sun H, Zheng X, Zhou Y, et al. Alphaxalone inhibits growth, migration and invasion of rat C6 malignant glioma cells. Steroids. 2013;78(10):1041–1045. doi:10.1016/j.steroids.2013.06.008
44. Hagmann W, Borgers S. Requirement for epidermal growth factor receptor tyrosine kinase and for 12-lipoxygenase activity in the expression of 12-lipoxygenase in human epidermoid carcinoma cells. Biochem Pharmacol. 1997;53(7):937–942. doi:10.1016/S0006-2952(96)00833-7
45. Dinney CP, Parker C, Dong Z, et al. Therapy of human transitional cell carcinoma of the bladder by oral administration of the epidermal growth factor receptor protein tyrosine kinase inhibitor 4,5-dianilinophthalimide. Clin Cancer Res. 1997;3(2):161–168.
46. Wu Q, Zhang B, Sun Y, et al. Identification of novel biomarkers and candidate small molecule drugs in non-small-cell lung cancer by integrated microarray analysis. Onco Targets Ther. 2019;12:3545–3563. doi:10.2147/OTT.S198621
47. Wang Q, Zhao Z, Shang J, Xia W. Targets and candidate agents for type 2 diabetes treatment with computational bioinformatics approach. J Diabetes Res. 2014;2014:763936. doi:10.1155/2014/763936
48. Funai K, Honzawa K, Suzuki M, et al. Urinary fluorescent metabolite O-aminohippuric acid is a useful biomarker for lung cancer detection. Metabolomics. 2020;16(10):101. doi:10.1007/s11306-020-01721-y
49. Snyder S, Murundi S, Crawford L, Putnam D. Enabling P-glycoprotein inhibition in multidrug resistant cancer through the reverse targeting of a quinidine-PEG conjugate. J Control Release. 2020;317:291–299. doi:10.1016/j.jconrel.2019.11.027
50. Jones RD, Kerr DJ, Harnett AN, Rankin EM, Ray S, Kaye SB. A pilot study of quinidine and epirubicin in the treatment of advanced breast cancer. Br J Cancer. 1990;62(1):133–135. doi:10.1038/bjc.1990.244
51. Wishart GC, Bissett D, Paul J, et al. Quinidine as a resistance modulator of epirubicin in advanced breast cancer: mature results of a placebo-controlled randomized trial. J Clin Oncol. 1994;12(9):1771–1777. doi:10.1200/JCO.1994.12.9.1771
52. Melkoumian ZK, Martirosyan AR, Strobl JS. Myc protein is differentially sensitive to quinidine in tumor versus immortalized breast epithelial cell lines. Int J Cancer. 2002;102(1):60–69. doi:10.1002/ijc.10648
53. Ru Q, Tian X, Pi MS, et al. Voltagegated K+ channel blocker quinidine inhibits proliferation and induces apoptosis by regulating expression of microRNAs in human glioma U87MG cells. Int J Oncol. 2015;46(2):833–840. doi:10.3892/ijo.2014.2777
54. Weiger TM, Colombatto S, Kainz V, Heidegger W, Grillo MA, Hermann A. Potassium channel blockers quinidine and caesium halt cell proliferation in C6 glioma cells via a polyamine-dependent mechanism. Biochem Soc Trans. 2007;35(Pt 2):391–395. doi:10.1042/BST0350391
55. Utermark T, Alekov A, Lerche H, Abramowski V, Giovannini M, Hanemann CO. Quinidine impairs proliferation of neurofibromatosis type 2-deficient human malignant mesothelioma cells. Cancer. 2003;97(8):1955–1962. doi:10.1002/cncr.11275
56. Lee SY, Rhee YH, Jeong SJ, et al. Hydrocinchonine, cinchonine, and quinidine potentiate paclitaxel-induced cytotoxicity and apoptosis via multidrug resistance reversal in MES-SA/DX5 uterine sarcoma cells. Environ Toxicol. 2011;26(4):424–431. doi:10.1002/tox.20568
57. Kimura O, Kondo Y, Shimosegawa T. PPAR could contribute to the pathogenesis of hepatocellular carcinoma. PPAR Res. 2012;2012:574180. doi:10.1155/2012/574180
58. Yu Q, Wu L, Ji J, et al. Gut microbiota, peroxisome proliferator-activated receptors, and hepatocellular carcinoma. J Hepatocell Carcinoma. 2020;7:271–288. doi:10.2147/JHC.S277870
59. Wu CW, Farrell GC, Yu J. Functional role of peroxisome-proliferator-activated receptor gamma in hepatocellular carcinoma. J Gastroenterol Hepatol. 2012;27(11):1665–1669. doi:10.1111/j.1440-1746.2012.07213.x
60. Hsu HT, Sung MT, Lee CC, et al. Peroxisome proliferator-activated receptor gamma expression is inversely associated with macroscopic vascular invasion in human hepatocellular carcinoma. Int J Mol Sci. 2016;17(8):1226. doi:10.3390/ijms17081226
61. Tu K, Zheng X, Dou C, et al. MicroRNA-130b promotes cell aggressiveness by inhibiting peroxisome proliferator-activated receptor gamma in human hepatocellular carcinoma. Int J Mol Sci. 2014;15(11):20486–20499. doi:10.3390/ijms151120486
62. Xiao YB, Cai SH, Liu LL, Yang X, Yun JP. Decreased expression of peroxisome proliferator-activated receptor alpha indicates unfavorable outcomes in hepatocellular carcinoma. Cancer Manag Res. 2018;10:1781–1789. doi:10.2147/CMAR.S166971
63. Kurebayashi Y, Ojima H, Tsujikawa H, et al. Landscape of immune microenvironment in hepatocellular carcinoma and its additional impact on histological and molecular classification. Hepatology. 2018;68(3):1025–1041. doi:10.1002/hep.29904
64. Lim CJ, Lee YH, Pan L, et al. Multidimensional analyses reveal distinct immune microenvironment in hepatitis B virus-related hepatocellular carcinoma. Gut. 2019;68(5):916–927. doi:10.1136/gutjnl-2018-316510
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.