Back to Journals » Drug Design, Development and Therapy » Volume 14
Activation of Cholinergic Anti-Inflammatory Pathway in Peripheral Immune Cells Involved in Therapeutic Actions of α-Mangostin on Collagen-Induced Arthritis in Rats
Authors Yin Q, Wu Y, Pan S, Wang D, Tao M, Pei W, Zuo J ![]()
Received 14 February 2020
Accepted for publication 25 April 2020
Published 22 May 2020 Volume 2020:14 Pages 1983—1993
DOI https://doi.org/10.2147/DDDT.S249865
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tuo Deng
Qin Yin, 1,* Yi-jin Wu, 1,* Shu Pan, 1 Dan-dan Wang, 2, 3 Meng-qing Tao, 2, 3 Wei-ya Pei, 2 Jian Zuo 2– 4
1Department of Pharmacy, The Second Affiliated Hospital of Wannan Medical College, Wuhu 241000, People’s Republic of China; 2Department of Traditional Chinese Medicine, Yijishan Hospital of Wannan Medical College, Wuhu 241000, People’s Republic of China; 3Research Center of Integration of Traditional Chinese and Western Medicine, Wannan Medical College, Wuhu 241000, People’s Republic of China; 4Key Laboratory of Non-Coding RNA Transformation Research of Anhui Higher Education Institution, Wannan Medical College, Wuhu 241000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jian Zuo Tel +86 15056477665
Email [email protected]
Background: Studies have shown that α-mangostin (MG) could exert anti-rheumatic effects in vivo by restoring immunity homeostasis, and have indicated that activation of the choline anti-inflammatory pathway (CAP) may contribute to this immunomodulatory property. The current study was designed to further investigate the effects of MG on the CAP in peripheral immune cells and clarify its relevance to the potential anti-rheumatic actions.
Methods: The catalytic activity of acetylcholinesterase (AChE) and expression of α 7-nicotinic cholinergic receptor (α 7nAChR) in peripheral blood mononuclear cells (PBMCs) from rats with collagen-induced arthritis (CIA) or human volunteers were evaluated after MG treatment. Consequent influences on the immune environment were assessed by flow cytometry and ELISA analyses. Indirect effects on joints resulting from these immune changes were studied in a co-culture system comprised of fibroblast-like synoviocytes (FLSs) and PBMCs.
Results: MG promoted α 7nAChR expression in PBMCs both in vivo and in vitro, and inhibited the enzymatic activity of AChE simultaneously. Activation of the CAP was accompanied by a significant decrease in Th17 cells (CD4+IL-17A+), while no obvious changes concerning the distribution of other T-cell subsets were noticed upon MG treatment. Meanwhile, MG decreased the secretion of TNF-α and IL-1β under inflammatory conditions. PBMCs from MG-treated CIA rats lost the potential to stimulate NF-κB activation and pro-inflammatory cytokine production of FLSs in the co-culture system.
Conclusion: Overall, the evidence suggested that MG can improve the peripheral immune milieu in CIA rats by suppressing Th17-cell differentiation through CAP activation, and achieve remission of inflammation mediated by FLSs.
Keywords: rheumatoid arthritis, α-mangostin, choline anti-inflammatory pathway, α 7-nicotinic cholinergic receptor, Th17/Treg
Corrigendum for this paper has been published
Introduction
Rheumatoid arthritis (RA) is an autoimmune disease characterized by chronic inflammation in joints together with synovial hyperplasia and bone/cartilage destruction.1 There is accumulating evidence suggesting that the development of RA is the result of lymphocyte abnormalities. In particular, imbalance of CD4+ T-cell subsets plays a dominant role in the etiology of RA.2 According to this consensus, biological therapies targeting cytokines involved in pro-inflammatory T-cell-related immune responses, such as TNF-α and IL-1β, have been applied successfully to RA treatments. However, these regimens are expensive and usually accompanied by a high risk of side effects.3
In recent years, great progress has been made in understanding the association of cholinergic systems with immunity and inflammation. The central nervous system is believed to have a rapid neurotransmitter-like regulatory effect on inflammatory responses, through the cholinergic anti-inflammatory pathway (CAP).4 The molecular basis for CAP activation is the stimulation of the α7-nicotinic cholinergic receptor (α7nAChR) by autonomic nervous system-released acetylcholine (ACh).4 Up-regulation of the CAP affects a variety of immune cell functions and eventually produces anti-inflammatory effects in the periphery.4,5 The CAP in RA patients is generally in a state of inhibition, and an agonist of α7nAChR was proved to be capable of controlling inflammation in experimental arthritis.6 Considering the profound influences of the CAP on immunity and inflammation, the prospects of CAP-targeting anti-rheumatic therapies are promising, although no such drugs are available so far. Identification of CAP agonists from conventional drugs or natural products is a feasible approach to achieve breakthroughs in RA treatments in this regard.
Previously, we found that xanthone derivatives (XANs) can restore the cytokine network balance, alleviate oxidative stresses, inhibit hyperproliferation of fibroblast-like synoviocytes (FLSs), and thereby have good therapeutic effects on experimental arthritis.7–13 Among investigated compounds, the anti-rheumatic potential of α-mangostin (MG, an isoprenyl-substituted xanthone isolated from mangosteen) is especially notable. It not only significantly improved systematic inflammation but also greatly reduced joint damage in rats with experimental arthritis.12,13 Our previous study found that MG induced cholinergic nerve excitation in rats by promoting α7nAChR expression, and consequently reduced the severity of LPS-induced acute lung injuries. As it has been reported that XANs are ideal candidates as acetylcholinesterase (AChE) inhibitors, we assume that MG could cure RA by manipulating the CAP in various ways.14
In the course of RA, infiltrated immune cells produce a large amount of pro-inflammatory cytokines in synovial tissue. They, together with FLSs, initiate and perpetuate pathological changes in joints.1,2 Therefore, it is of great significance to study the effects of MG on immunity through CAP regulation in the case of RA. In this study, we further validated the stimulatory effects of MG on the CAP in peripheral immune cells, and preliminarily clarified its relevance to the anti-rheumatic actions in vivo.
Materials and Methods
Materials and Reagents
MG (purity >98%) and carboxyl methyl cellulose sodium (CMC-Na) were obtained from SanHerb Bioscience (Chengdu, Sichuan, People’s Republic of China) and Aladdin (Shanghai, People’s Republic of China), respectively. Lipopolysaccharide (LPS, from the Gram-negative bacterium E. coli055:B5), nicotine (Nic, α7nAChR agonist), and methyllycaconitine (MLA, α7nAChR specific inhibitor) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Dulbecco’s modified Eagle’s medium (DMEM) and Roswell Park Memorial Institute (RPMI) 1640 medium were bought from Hyclone (Logan City, UT, USA). Lymphocyte separation solution was purchased from Solarbio (Beijing, People’s Republic of China). Anti-human CD4-APC, IL-17A-FITC, CD25-FITC, and CD127-PE-CY7 antibodies were obtained from BioLegend (San Diego, CA, USA). ELISA kits (rat TNF-α, IL-1β; human TNF-α, IL-1β, IL-10) were supplied by Boster Biotech (Wuhan, People’s Republic of China). Anti-rat p65, p-p65, and α7nAChR antibodies were products of Cell Signaling Technology (Boston, MA, USA). Biotin-conjugated secondary antibodies were purchased from Beyotime Biotech (Nantong, Jiangsu, People’s Republic of China).
Animals and Volunteers
Male Sprague–Dawley (SD) rats (180±20 g) were purchased from Qinglongshan Experimental Animal Company (Nanjing, People’s Republic of China). The animals were acclimated for 7 days prior to experimental procedures. The animal experimental protocols were approved by the Ethical Committee of Yijishan Hospital of Wannan Medical College (no. YJS 2018-8-004), strictly in accordance with the Guidelines for the Care and Use of Laboratory Animals (United States National Research Council, 2011). All volunteers were patients without RA or other immune diseases recruited from the Second Affiliated Hospital of Wannan Medical College, and signed an informed consent agreement for blood sample donation.
Induction of Collagen-Induced Arthritis (CIA) in Rats and Treatments
Bovine type II collagen (Chondrex, Redmond, WA, USA) was dissolved in 0.1 M acetic acid (the final concentration of collagen was 2 g/L). The solution was then mixed into incomplete Freund’s adjuvant (IFA) dropwise, and fully emulsified in an ice-water bath to obtain a milky homogeneous emulsion. Then, 0.2 mL of the emulsion was intradermally injected into back and tail roots of rats at multiple sites. At day 7, a booster injection was carried out. After the first injection, 12 immunized rats were randomly assigned into model control and MG treatment groups, and another six healthy rats were adopted as normal controls. MG was dissolved in 0.5% CMC-Na with the aid of ethanol and Tween 40, and given to rats by gavage at a dose of 40 mg/kg daily for 45 consecutive days. Normal and CIA model controls were given an equal volume of CMC-Na instead.13 For the duration of the experiment, the arthritis index was evaluated periodically (every 3 days) from the onset of CIA (day 17).
Histopathological and Immunohistochemical Evaluation
The rats were killed on day 45, then the hind ankle joints and spleen were fixed in 4% formaldehyde solution. Then, specimens were subjected to paraffin embedding, sectioning, and hematoxylin–eosin (HE) staining procedures. With the aid of a light microscope, pathological changes in joints and spleen were observed and scored with the same criteria defined previously.15–17
Certain samples were dewaxed and dehydrated in xylene and gradient ethanol solutions, respectively. Processed specimens were then treated with 0.5% Triton X-100, and soaked in citrate buffer (0.01 M) for antigen retrieval with the aid of heating. After a further treatment with 3% H2O2, the sections were incubated with α7nAChR primary antibody (1:100, at 37°C for 60 min) and biotin-conjugated secondary antibody (1:1000, at room temperature for 20 min) in turn, and extensive washing with PBS was carried out in every interval. The signal was then visualized by diaminobenzidine (DAB) chromogenic solution and hematoxylin (Zsbio, Beijing, People’s Republic of China). Stained sections were finally dehydrated, sealed, and observed under a light microscope.16
Cell Harvest and Culture
FLSs were obtained from synovial tissue of healthy rats. The synovium of rats was washed repeatedly with D-Hank’s solution (containing 200 mg/L streptomycin and 200 U/mL penicillin), then deliberately cut, seeded on the bottom of cell culture flasks, and cultured in DMEM containing 20% fetal bovine serum at 37°C in 5% CO2.18 When 80% confluence had been reached, a passage was carried out at the ratio of 1:2. Cells from passages 3–6 were used in the following studies. Peripheral blood mononuclear cells (PBMCs) from rats in different experimental groups were prepared by taking the following steps. Fresh blood was diluted with an equal volume of prechilled PBS. The resulting cell suspension was transferred into 15 mL centrifuge tubes containing the same amount of separation solution. After the gradient centrifugation, a translucent cell layer appeared in the centrifuge tube. The cells were subsequently sucked out, and washed three times with RPMI 1640 medium. After further centrifugation at 250 g for 10 min, PBMCs were collected for immunoblotting assays or other experiments.
Co-Culture of FLSs with PBMCs
PBMCs obtained from rats receiving different treatments were seeded into Transwell chambers together with FLSs from healthy rats (PBMCs were located in the upper chamber and FLSs in the lower chamber). The density was 2×105/well for both of them, and RPMI 1640 medium containing 20% fetal bovine serum was adopted in the co-culture system. After 24 h, culture media from the both upper and lower chambers were collected separately, and the levels of IL-1β and TNF-α were quantified by an ELISA method. FLSs were then harvested for protein extraction and immunoblotting assay.
Evaluation of AChE Activity in PBMCs Under MG Treatments
Prior to the assay, PBMCs obtained from healthy rats were treated with MG at various concentrations (2.5, 5, and 10 μg/mL) for 2 h. The catalytic activity of AChE in treated cells was assessed by a commercially available detection kit purchased from Solarbio (Beijing, People’s Republic of China). In brief, PBMCs were lysed using the extraction solution in the kit (1 mL of extraction solution for every 5×106 cells) in an ice bath with the aid of supersonic treatment. The lysate was then centrifuged at 8000 g at 4°C for 10 min, and the supernatant was collected. After reaction with the provided reagents, the optical absorbance of the tested samples was read at 412 nm, and the readings were used for AChE activity calculation based on the quantitative formula given in the instructions.
Western Blot Assay
Protein expression in PBMCs and FLSs from rats was investigated by the Western blot method. First, qualified protein samples (10 μg/well, based on the BCA test) were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Separated proteins were transferred onto polyvinylidene fluoride (PVDF) membranes, which were then incubated with primary antibodies (α7nAChR anti-rabbit, 1:1000; p65 anti-rabbit, 1:1000; p-p65anti-rabbit, 1:1000; GAPDH anti-rat, 1:5000) overnight at 4°C, followed by blocking with 5% skim milk for 2 h at room temperature. Subsequently, the specific antigen of the primary antibody was reacted with secondary antibodies (anti-rabbit IgG, HRP-linked antibody, 1:10,000; anti-rat IgG, HRP-linked antibody, 1:10,000). Signals were finally developed with an enhanced chemiluminescence (ECL) substrate kit. The bands were photographed and analyzed using ImageJ software.
Flow Cytometry
Human PBMCs were isolated from whole blood donated by volunteers using the same protocol described in Cell Harvest and Culture, and then seeded into 24-well plates at a density of 5×106 cells/well. Experimental arrangements were as follows: normal control, LPS (2 nM), LPS + MG (5 μg/mL), LPS + Nic (10 μM), LPS + MLA (5 nM), LPS + MG + MLA, and LPS + Nic + MLA. After stimulation for 24 h, the cells were harvested for two different analytical purposes. One portion of the cells was stained with different fluorescein-labeled CD4, CD25, and CD127 flow antibodies in the dark for 30 min; the other portion was stained with fluorescein-labeled CD4 and IL-17A flow antibodies in turn, with proper fixing and permeabilizing processes during the interval. The stained cells were subsequently fed to a flow cytometer (FC500; Beckman) for quantitative analysis. Obtained data were analyzed by CytExpert (version 2.1). CD4+CD25+CD127low/– and CD4+IL-17A+ cells were identified as Treg and Th17 cells, respectively.
Data Analysis
The data were expressed as mean ± SD. Data analysis was performed using SPSS 16.0 software, and statistical differences among different groups were compared by one-way analysis of variance (ANOVA) coupled with t-tests. Values of P<0.05 and P<0.01 were considered to be statistically significant.
Results
MG Alleviated the Severity of CIA in Rats
Generally speaking, this study further validated the therapeutic effects of MG on experimental arthritis.12,13 As shown in Figure 1A, CIA progressed quickly in rats during the period between days 17 and 28, and then was ameliorated spontaneously. MG significantly reduced the arthritis index, mainly in the late stage of CIA (after day 28). This provided basic evidence for the anti-rheumatic properties of MG in vivo.
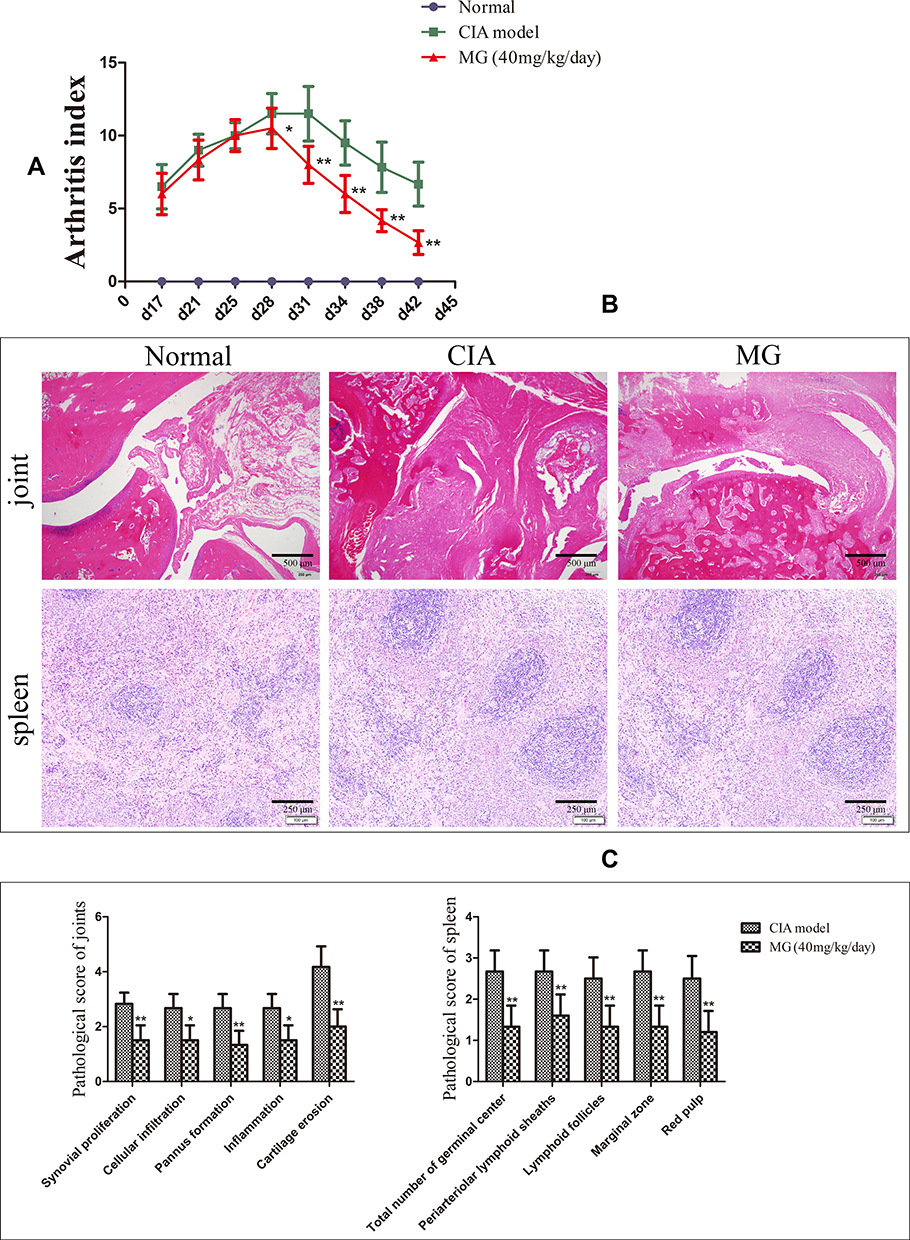 |
Figure 1 Therapeutic effects of MG on CIA rats. (A) Arthritis index changes in rats; (B) histological examination of joint and spleen; (C) histopathological scores of joint and spleen. Data are presented as mean ± SD (n=6). *P<0.05 and **P<0.01 compared with CIA rats. |
Compared with normal controls, obvious synovial hyperplasia, as well as inflammatory infiltration, articular cartilage erosion, and pannus formation, were found in CIA rats. MG ameliorated all the pathological changes effectively (Figure 1B and C). Besides, MG notably alleviated abnormal changes in the spleen of CIA rats, including an increase in lymphocyte density around arteries in the germinal center, white pulp hyperplasia, and boundary blurring of the red pulp (Figure 1B and C).
Possible Stimulatory Effects of MG on the CAP in CIA Rats
α7nAChR was mainly expressed on lymphocytes located in the medullary part of the spleen (Figure 2A). Its expression in CIA rats was largely decreased, indicating the deficiency of the CAP under such pathological circumstances. In comparison with CIA rats, MG treatment restored this decline efficiently (Figure 2B). This result demonstrates the positive effects of MG on the CAP in the immune system in vivo.
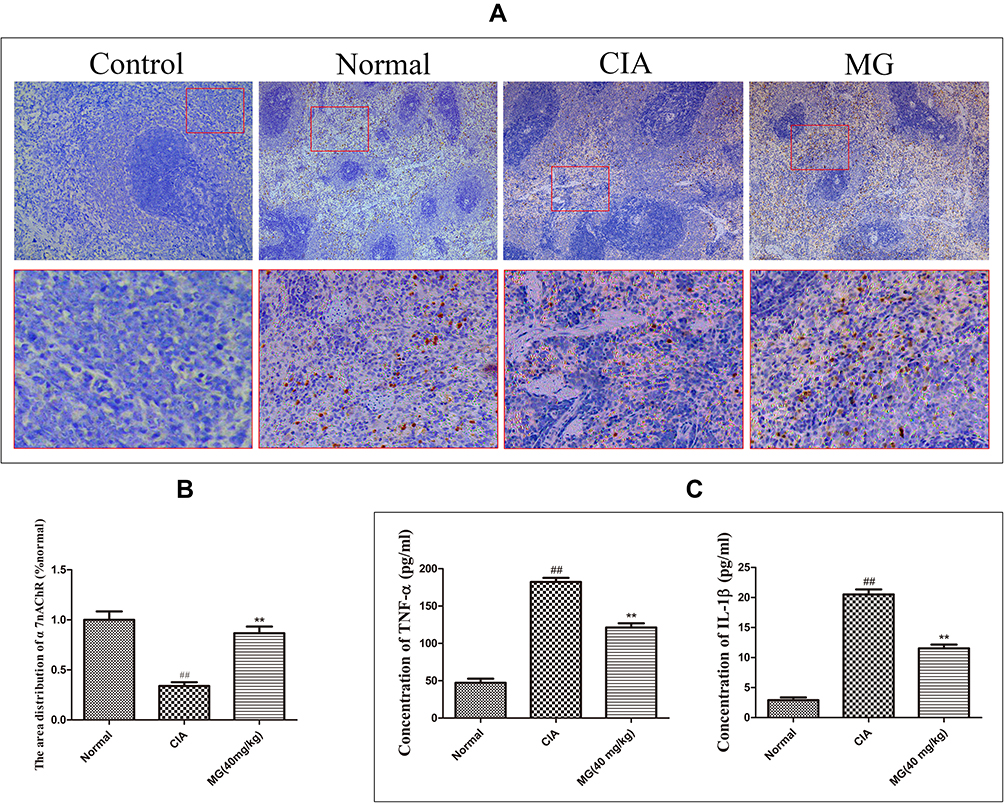 |
Figure 2 Possible effects of MG on CAP in CIA rats. (A) Local expression of α7nAChR in spleen; isotype controls were processed with routine immunohistochemical procedures but incubated with normal serum instead of specific primary antibody; (B) quantification results of the immunohistochemical experiment analyzed by ImageJ software based on optical density; (C) levels of TNF-α and IL-1β in rat serum determined by ELISA. Data are presented as mean ± SD (n=6). ##P<0.01 compared with normal healthy rats; **P<0.01 compared with CIA rats. |
Consistent with the anti-inflammatory functions of the CAP, the decrease in α7nAChR in CIA rats was accompanied by obvious increases in circulating TNF-α and IL-1β. It seems that up-regulation of α7nAChR was favorable to the remission of inflammation, as MG treatment induced significant decreases in the two pro-inflammatory cytokines simultaneously (Figure 2C).
MG Activated the CAP in PBMCs
Although MG promoted α7nAChR expression in the spleen, it cannot simply be deduced that MG would rebalance the disrupted immune system in CIA rats through CAP activation, because lymphocytes in the spleen are mainly comprised of B cells, whereas the pathology of RA/CIA is mainly mediated by peripheral T cells. Therefore, we investigated the effects of MG on the CAP in PBMCs in the following experiments, because PBMCs are largely comprised of T cells, and they contribute significantly to the local inflammation in RA patients, as the direct resource of infiltrated immune cells in joints. First, we examined the catalytic activity of AChE in PBMCs, and found that the enzymatic activity was inhibited by MG at 5 μg/mL shortly after treatment (Figure 3A). This change would inevitably lead to the accumulation of ACh and consequent CAP activation in peripheral tissues. Since there is a considerable amount of monocytes in PBMCs, which affect T-cell differentiation greatly and initiate malicious immune feedback in many inflammatory diseases, we stimulated PBMCs with LPS and investigated the effects of MG on the overall immune milieu. As shown in Figure 3B, LPS stimulation resulted in an obvious decrease in α7nAChR in PBMCs, which was significantly restored by MG at 5 μg/mL. Taken together, these changes show that MG has a potential stimulatory effect on the impaired CAP in the peripheral immune system under CIA/RA circumstances.
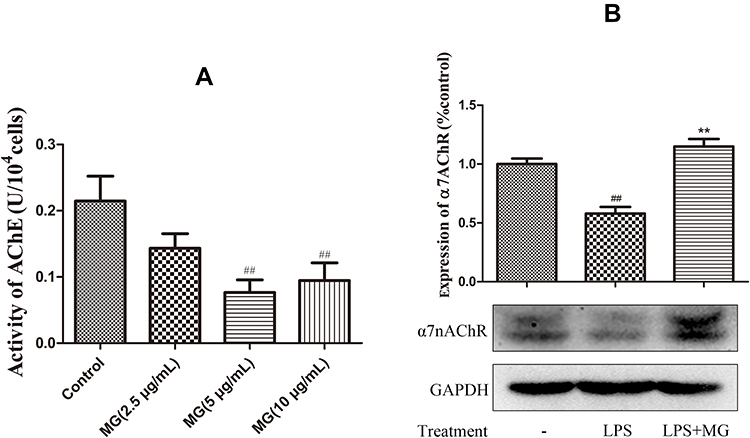 |
Figure 3 MG activated CAP in PBMCs from rats. (A) Inhibitory effect of MG on enzymatic activity of AChE in PBMCs; data are presented as mean ± SD (n=5); (B) MG reduced α7nAChR expression in LPS-pretreated PBMCs (double bands revealed in the immunoblotting assay were caused by unspecified modification and degradation of α7nAChR). Data are presented as mean ± SD (n=3). ##P<0.01 compared with normal controls, **P<0.01 compared with LPS-pretreated cells. |
To test whether CAP-related changes substantially altered the differentiation profile of T cells, thereby having profound implications for RA treatment, we performed flow cytometry and ELISA analyses on human PBMCs. Because imbalances of Th17/Treg as well as Th1/Th2 play important roles in the occurrence and development of RA, we investigated the distribution of these subsets in LPS-pretreated PBMCs under MG treatment in combination with an α7nAChR antagonist or agonist.19 As anticipated, LPS augmented the Th17 subset population (Figure 4A). Both MG and Nic significantly reduced its distribution, from 10.0% to 5.6% and 4.7%, respectively. Although the selective α7nAChR inhibitor MLA alone seemed to have no effect, it reversed the MG-induced decrease in the Th17 subset. A similar antagonistic effect between MLA and Nic was also observed (Figure 4B). These findings indicate that activation of the CAP by either MG or Nic substantially hampered the differentiation of Th17 cells. To our disappointment, none of the treatments increased Treg distribution (Figure 4). Nonetheless, MG down-regulated the Th17/Treg ratio substantially in LPS-pretreated PBMCs owing to the significant decrease in Th17 cells. The involvement of monocytic polarization in Th17-cell differentiation under these treatments was preliminarily proved by ELISA. Activation of the CAP by MG and Nic led to decreased production of TNF-α and IL-1β, which was then antagonized by MLA (Figure 5). This suggests that CAP activation could abrogate pro-inflammatory phenotype (M1) polarization of monocytes induced by LPS, and consequently have a profound influence on disturbed T cells. Meanwhile, MG treatment reduced both Th1 and Th2 cells slightly, but the ratio of Th1/Th2 was barely affected (data not provided). Considering the well-recognized immunosuppressive functions of Th2 cells, this phenomenon is confusing, and its clinical implications are not totally understood. Consistent with the results from flow cytometric analyses, none of these treatments had a significant effect on the secretion of IL-10 (Figure 5).
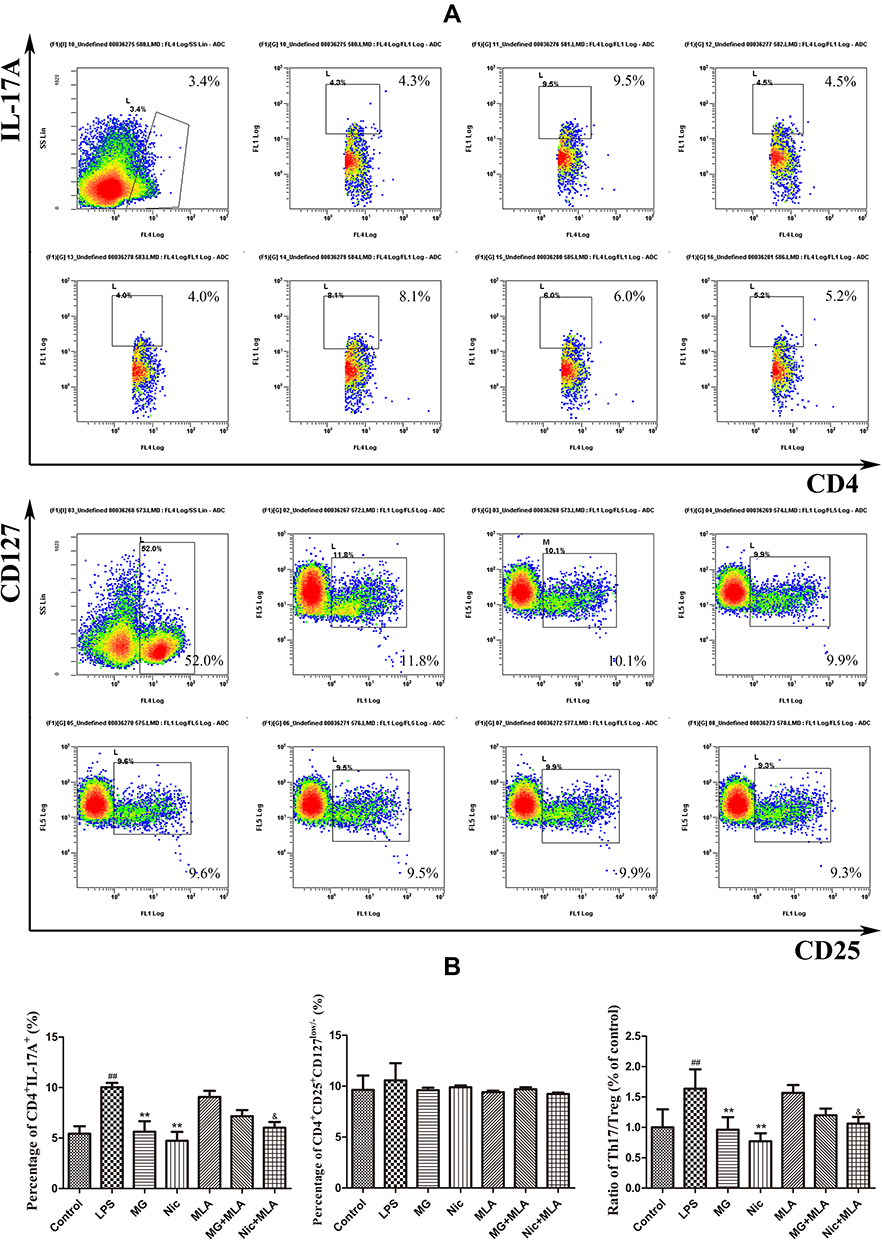 |
Figure 4 Effects of MG on T-cell subset distribution in human PBMCs under LPS stimulus investigated by flow cytometry. (A) Representative raw data indicating Th17 andTreg cells distribution; (B)quantification results of flow cytometry experiments. All cells in treatment groups were pretreated with LPS. Data are presented as mean ± SD (n=3). ##P<0.01 compared with normal controls, **P<0.01 compared with LPS-pretreated cells, &P<0.05 compared with MLA-treated cells. |
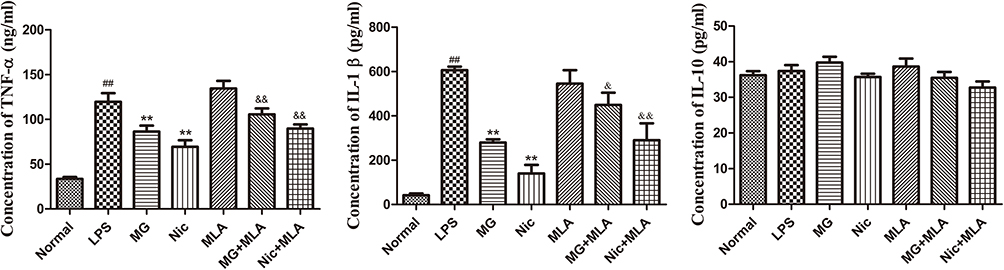 |
Figure 5 Effects of MG on cytokine production of human PBMCs under LPS stimulus evaluated by ELISA. All cells in treatment groups were pretreated with LPS. Data are presented as mean ± SD (n=3). ##P<0.01 compared with normal controls, **P<0.01 compared with LPS-pretreated cells, &P<0.05 and &&P<0.01 compared with MLA-treated cells. |
MG-Induced CAP Activation in PBMCs Alleviated FLS-Mediated Inflammation
Th17 cells and other pro-inflammatory immune cells play important pathological roles in tissue degradation and inflammation in joints through interactions with FLSs. Therefore, in theory, immune improvement characterized by a decrease in Th17 cells under MG treatment would be beneficial for the alleviation of FLS-mediated inflammation, and we tested this assumption using a co-culture system. Compared with the controls, PBMCs obtained from CIA rats promoted the secretion of both TNF-α and IL-1β in normal FLSs (Figure 6A). In contrast, PBMCs obtained from MG-treated rats lost the ability to stimulate the release of these cytokines. In addition, the accelerated phosphorylation of p65 in normal FLSs induced by PBMCs from CIA rats was suppressed by MG treatment (Figure 6B). As hyperactivation of nuclear factor-κB (NF-κB) is deeply implicated in synovitis, this change was essential to the alleviation of arthritic manifestations in CIA/RA.
 |
Figure 6 Effects of PBMCs obtained from rats receiving different treatments on FLS-mediated inflammation in co-culture system. (A) Levels of TNF-α and IL-1β in culture medium of FLS determined by ELISA; (B) expression of p-p65 and p65 in FLS investigated by immunoblotting. Data are presented as mean ± SD (n=3). ##P<0.01 compared with normal controls, **P<0.01 compared with samples co-cultured with CIA rat-derived PBMCs. |
Discussion
MG is a naturally occurring polyphenol, which is mainly distributed in the inedible pericarp of mangosteen. Because of its significant bioactivity and high abundance, exploitation of its medicinal potential is not only clinically meaningful but also economically efficient.20 Previous studies suggested that MG could have promising applications in RA treatment.12,13 Our preliminary observations showed that MG-enriched extract of mangosteen could activate the CAP in CIA rats, which was accompanied by significantly altered T-cell differentiation in both immune organs and peripheral tissues.21 It is highly plausible that CAP activation contributed directly toward the improved immune milieu upon such treatment. To prove this hypothesis, the priority was to clarify the effects of MG on the CAP in the immune system. From this point of view, the results of this study are very meaningful and inspiring. On one hand, MG treatment greatly inhibited the catalytic activity of AChE in PBMCs. The impaired hydrolysis will eventually result in slowed ACh clearance, and elicit fast anti-inflammatory effects in the short term due to prolonged CAP activation. On the other hand, MG treatment promoted α7nAChR expression in immune cells both in vivo and in vitro, which was crucial for the sustained CAP activation, according to the current understanding of signal transduction mechanisms in this pathway.
Although it is widely accepted that the pathological changes of RA are mainly controlled by lymphocytes, especially CD4+ T cells, the role of monocytes/macrophages in RA cannot be overlooked. As indispensable components of innate immunity, these cells sense pathogens first, and confer the signals to the adaptive immune system. Furthermore, prominent M1 polarization is deeply implicated in arthritic changes, including persistent inflammation and progressive tissue damages in joints, mainly through immune responses. From these perspectives, imbalanced polarization of monocytes/macrophages could be largely responsible for the deteriorated immune milieu in RA patients. We found that MG could substantially ease monocyte-mediated inflammation.22 Our earlier study suggested that this phenomenon was closely related to CAP activation. As the pro-inflammatory phenotype of monocytes/macrophages promotes the differentiation of Th1/17 cells, the inhibition of MG on M1 polarization through CAP activation will benefit the rebalancing of disrupted CD4+ T-cell subset distribution under RA/CIA conditions.23 Differently from previous findings, MG decreased only Th17 cells in this study, but had little effect on other CD4+ subsets.21 This discrepancy could be caused by the different experimental strategies adopted. Previously, all results were obtained from CIA rats. In this study, we mainly focused on the effects of MG on pro-inflammatory phenotype monocyte-mediated T-cell differentiation through CAP activation in human PBMCs under LPS stimulus. More accurate effects of MG on T-cell differentiation involved in CAP activation will be investigated in future research. Nonetheless, the significant decrease in Th17 cells partly explained the positive influence of MG on peripheral immunity by activating the CAP.
Because of their aggressive and pro-inflammatory nature, FLSs are believed to be important in the development of arthritic manifestations, and an ideal target for RA treatment. We have already noticed that the effective protective effect of XANs in experimental arthritis was closely related to their inhibitory effects on RA-FLSs.9–12 MG and other XANs are capable of down-regulating the NF-κB pathway in RA-FLSs both in vivo and in vitro, and therefore have great potential in treating synovitis-related symptoms.10–12 It should be noted that the abnormal functions of RA-FLSs result largely from immune responses of infiltrated immune cells. Consequently, improvement of the immune environment in joints is especially meaningful to ease FLS-mediated pathological changes. In this study, we co-cultured PBMCs from rats receiving different treatments with normal FLSs. It was found that CIA rat-derived PBMCs potently initiated inflammatory reactions in FLSs, and MG showed an indirect anti-inflammatory effect on RA-FLSs, as PBMCs from MG-treated CIA rats lost the ability to stimulate the secretion of TNF-α and IL-1β and phosphorylation of p65 in co-cultured FLSs. These findings suggest that the restoration of immune homeostasis under MG treatments could be the foundation for the overall therapeutic effects on experimental arthritis.
Conclusion
This study revealed the potential of MG in activating the CAP in peripheral immune cells by inhibiting AChE catalytic activity and promoting α7nAChR expression. These changes consequently suppressed Th17 cell differentiation through immune responses with monocytes under inflammatory circumstances, and alleviated FLS-mediated pathological changes in joints. In summary, we demonstrated that MG can improve the peripheral immune milieu through CAP activation, and ameliorate the severity of CIA.
Acknowledgments
This work was supported by the Key Project of Natural Science Foundation of Anhui Province for College Scholar (KJ2018A0249, KJ2019A0416), National Natural Science Foundation of China (81973828), Talent Cultivation and International Academic Visiting Project for College Scholar of Anhui Province (gxfxZD2016163), and Funding from the “Peak” Training Program for Scientific Research of Yijishan Hospital, Wannan Medical College (GF2019J01).
Disclosure
The authors declare no conflict of interest.
References
1. Littlejohn EA, Monrad SU. Early diagnosis and treatment of rheumatoid arthritis. Prim Care. 2018;45(2):237–255. doi:10.1016/j.pop.2018.02.010
2. Lina C, Conghua W, Nan L, Ping Z. Combined treatment of etanercept and MTX reverses Th1/Th2, Th17/Treg imbalance in patients with rheumatoid arthritis. J Clin Immunol. 2011;31(4):596–605. doi:10.1007/s10875-011-9542-6
3. Venkatesha SH, Dudics S, Acharya B, Moudgil KD. Cytokine-modulating strategies and newer cytokine targets for arthritis therapy. Int J Mol Sci. 2014;16(1):887–906. doi:10.3390/ijms16010887
4. Tracey KJ. Physiology and immunology of the cholinergic antiinflammatory pathway. J Clin Invest. 2007;117(2):289–296. doi:10.1172/JCI30555
5. de Jonge WJ, Ulloa L. The alpha7 nicotinic acetylcholine receptor as a pharmacological target for inflammation. Br J Pharmacol. 2007;151(7):915–929. doi:10.1038/sj.bjp.0707264
6. van Maanen MA, Vervoordeldonk MJ, Tak PP. The cholinergic anti-inflammatory pathway: towards innovative treatment of rheumatoid arthritis. Nat Rev Rheumatol. 2009;5(4):229–232. doi:10.1038/nrrheum.2009.31
7. Zuo J, Xia Y, Li X, Chen JW. Xanthones from Securidaca inappendiculata exert significant therapeutic efficacy on adjuvant-induced arthritis in mice. Inflammation. 2014;37(3):908–916. doi:10.1007/s10753-014-9810-8
8. Zuo J, Xia Y, Mao KJ, Li X, Chen JW. Xanthone-rich dichloromethane fraction of Securidaca inappendiculata, the possible antirheumatic material base with anti-inflammatory, analgesic, and immunodepressive effects. Pharm Biol. 2014;52(11):1367–1373. doi:10.3109/13880209.2014.892143
9. Zuo J, Xia Y, Li X, Ou-Yang Z, Chen JW. Selective modulation of MAPKs contribute to the anti-proliferative and anti-inflammatory activities of 1,7-dihydroxy-3,4-dimethoxyxanthone in rheumatoid arthritis-derived fibroblast-like synoviocyte MH7A cells. J Ethnopharmacol. 2015;168:248–254. doi:10.1016/j.jep.2015.03.069
10. Zuo J, Dou DY, Wang HF, Zhu YH, Li Y, Luan JJ. Reactive oxygen species mediated NF-κB/p38 feedback loop implicated in proliferation inhibition of HFLS-RA cells induced by 1,7-dihydroxy-3,4-dimethoxyxanthone. Biomed Pharmacother. 2017;94:1002–1009. doi:10.1016/j.biopha.2017.07.164
11. Ji CL, Jiang H, Tao MQ, Wu WT, Jiang J, Zuo J. Selective regulation of IKKβ/NF-κB pathway involved in proliferation inhibition of HFLS-RA cells induced by 1,7-dihydroxyl-3,4-dimethoxylxanthone. Kaohsiung J Med Sci. 2017;33(10):486–495. doi:10.1016/j.kjms.2017.06.015
12. Zuo J, Yin Q, Wang YW, et al. Inhibition of NF-κB pathway in fibroblast-like synoviocytes by α-mangostin implicated in protective effects on joints in rats suffering from adjuvant-induced arthritis. Int Immunopharmacol. 2018;56:78–89. doi:10.1016/j.intimp.2018.01.016
13. Yang K, Yin Q, Mao Q, et al. Metabolomics analysis reveals therapeutic effects of α-mangostin on collagen-induced arthritis in rats by down-regulating nicotinamide phosphoribosyltransferase. Inflammation. 2019;42(2):741–753. doi:10.1007/s10753-018-0932-2
14. Ryu HW, Oh SR, Curtis-Long MJ, Lee JH, Song HH, Park KH. Rapid identification of cholinesterase inhibitors from the seedcases of mangosteen using an enzyme affinity assay. J Agric Food Chem. 2014;62(6):1338–1343. doi:10.1021/jf405072e
15. Germolec DR, Nyska A, Kashon M, et al. Extended histopathology in immunotoxicity testing: interlaboratory validation studies. Toxicol Sci. 2004;78(1):107–115. doi:10.1093/toxsci/kfh049
16. Liu Y, Zhang L, Wu Y, et al. Therapeutic effects of TACI-Ig on collagen-induced arthritis by regulating T and B lymphocytes function in DBA/1 mice. Eur J Pharmacol. 2011;654(3):304–314. doi:10.1016/j.ejphar.2011.01.002
17. Shu JL, Zhang XZ, Han L, et al. Paeoniflorin-6ʹ-O-benzene sulfonate alleviates collagen-induced arthritis in mice by downregulating BAFF-TRAF2-NF-κB signaling: comparison with biological agents. Acta Pharmacol Sin. 2019;40(6):801–813. doi:10.1038/s41401-018-0169-5
18. Jia XY, Chang Y, Wei F, et al. CP-25 reverses prostaglandin E4 receptor desensitization-induced fibroblast-like synoviocyte dysfunction via the G protein-coupled receptor kinase 2 in autoimmune arthritis. Acta Pharmacol Sin. 2019;40(8):1029–1039. doi:10.1038/s41401-018-0196-2
19. Lubberts E. Role of T lymphocytes in the development of rheumatoid arthritis. Implications for treatment. Curr Pharm Des. 2015;21(2):142–146. doi:10.2174/1381612820666140825122247
20. Obolskiy D, Pischel I, Siriwatanametanon N, Heinrich M. Garcinia mangostana L.: a phytochemical and pharmacological review. Phytother Res. 2009;23(8):1047–1065. doi:10.1002/ptr.2730
21. Zuo J, Yin Q, Wang L, et al. Mangosteen ethanol extract alleviated the severity of collagen-induced arthritis in rats and produced synergistic effects with methotrexate. Pharm Biol. 2018;56(1):455–464. doi:10.1080/13880209.2018.1506939
22. Tao M, Jiang J, Wang L, et al. α-Mangostin alleviated lipopolysaccharide induced acute lung injury in rats by suppressing NAMPT/NAD controlled inflammatory reactions. Evid-Based Compl Alt. 2018;2018:5470187. doi:10.1155/2018/5470187
23. Harris KM. Monocytes differentiated with GM-CSF and IL-15 initiate Th17 and Th1 responses that are contact-dependent and mediated by IL-15. J Leukoc Biol. 2011;90(4):727–734. doi:10.1189/jlb.0311132
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.