Back to Journals » International Journal of Women's Health » Volume 17
A Two-Sample Mendelian Randomization Study of Basophil Count and Risk of Gestational Diabetes Mellitus
Authors Mao J, Gan Y, Tan X, He Y, Jing Q, Shi Q
Received 12 October 2024
Accepted for publication 22 January 2025
Published 26 February 2025 Volume 2025:17 Pages 517—527
DOI https://doi.org/10.2147/IJWH.S500632
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Vinay Kumar
Jing Mao,* Yanqiong Gan,* Xinlin Tan, Yuhan He, Qiao Jing, Qi Shi
Affiliated Hospital of North Sichuan Medical College, Nanchong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Qi Shi, Affiliated Hospital of North Sichuan Medical College, No. 1, Maoyuan South Road, Shunqing District, Nanchong, Sichuan, People’s Republic of China, Tel +8615983789947, Email [email protected]
Objective: High basophil count levels are associated with an increased risk of gestational diabetes mellitus (GDM). We used two-sample Mendelian randomisation (MR) to explore a potential causal relationship. It also aims to offer genetic evidence supporting the link between basophil count and the development of gestational diabetes mellitus while addressing the potential issues of confounding and reverse causality commonly encountered in observational studies.
Methods: We utilized publically accessible summary information obtained from genome-wide association studies (GWAS) for conducting a two-sample Mendelian randomization (TSMR) study. The major analysis method employed was inverse variance weighted (IVW), whereas the other four methods, namely weighted median method, MR-Egger regression, simple model and weighted model, were used as supplemental analyses. We also investigated the relationship between GDM and basophil count in the opposite direction using directional validation of MR analysis. Furthermore, the R package “ClusterProfiler” to conduct an analysis of Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways and Gene Ontology (GO) terms was used. Additionally, with the help of the STRING database, we have constructed a network of protein–protein interactions (PPIs).
Results: The Inverse Variance Weighted (IVW) method revealed a significant causal association between basophil count and gestational diabetes mellitus (OR, 0.84; 95% CI; 0.74– 0.96; P, 0.01). A sensitivity analysis was performed to assess the reliability of the results, indicating no indication of pleiotropy or heterogeneity, hence strengthening the validity of the findings. The reverse causation of GDM predisposition on basophil counts was not supported by the results of the directional validation of the MR analysis.
Conclusion: The results of this study showed a causal relationship between high basophil counts and increased risk of GDM but did not support a causal relationship between genetic susceptibility to GDM and basophil counts.
Keywords: basophil count, gestational diabetes mellitus, GDM, Mendelian randomization
Introduction
Gestational diabetes mellitus (GDM) is defined as any degree of glucose intolerance that begins or is first detected during pregnancy, regardless of the degree of hyperglycemia.1 It is a prevalent medical issue during pregnancy in our nation. The occurrence of GDM differs significantly based on demographic factors such as maternal age, ethnicity, or body mass index (BMI), as well as the methods utilized for screening and the criteria used for diagnosis, including the glucose threshold.2,3 The International Diabetes Federation (IDF) 2021 Global Diabetes Map reported that GDM affected 14% of pregnant women globally;4 and in China, the incidence of GDM ranged from 17.5% to 18.9%, and there was a noticeable tendency for it to affect younger patients.5 GDM can result in various unfavorable consequences for pregnant mothers and their offspring.6 Pregnant women may experience adverse perinatal outcomes such as pre-eclampsia, urinary tract/vaginal infection, caesarean delivery, postpartum hemorrhage, additionally prolonged type 2 diabetes mellitus (T2DM) and cardiovascular disease. Adverse consequences for offspring encompass stillbirth, death shortly after birth, premature birth, low blood sugar in newborns, high levels of bilirubin in newborns, excessive birth weight, difficulty during birth due to shoulder obstruction, and the long-term development of metabolic syndrome.
Gestational diabetes mellitus is caused by multiple causative factors. GDM has major risk factors for its development, which are overweight or obesity, advanced pregnancy, history of polycystic ovary syndrome, history of hypothyroidism, history of GDM, family history of type 2 diabetes mellitus, multiple pregnancies, smoking and race.3,7
Although we still do not completely understand the pathophysiology of Gestational Diabetes, there are a range of potentially contributing factors, includingβ-Cell Dysfunction, Chronic Insulin Resistance, Neurohormonal (Leptin and Adiponectin) Networks, Adipose Tissue Energy Storage and Adipose Tissue Inflammation, Liver, Skeletal and Cardiac Muscle, Gut Microbiome, Oxidative Stress and Placental Transport.8 Recent research9 has discovered that chronic inflammation is also involved in GDM pathogenesis. Low-grade chronic inflammation is a process of immune system activation and increased circulating cytokines and chemokines, with adipose tissue appearing to be a major source of pro-inflammatory factors.10 Chronic inflammation may contribute to the onset of GDM by increasing insulin resistance, interfering with insulin signalling and promoting the dysfunction of beta cells.11–13 Several inflammatory markers may reflect the low-grade chronic inflammatory process, such as white blood cell (WBC) and its subtype, tumour necrosis factor-α, and interleukin-6.13–15 Of these, WBC is routinely measured in clinical practice. Circulating WBCs include granulocytes (neutrophils, eosinophils and basophils), lymphocytes and monocytes, which may be biologically linked to GDM through inflammation and insulin resistance. Briefly, basophils constitute fewer than 1% of the leukocytes in the bloodstream and spleen,16 and are an important non-specific immune cell type. Basophils are responsible for initiating and recruiting other inflammatory cells and produce cytokines and chemokines that promote chronic inflammation, insulin resistance and the development of diabetes.17–19 Although several observational studies have attempted to clarify the relationship between basophil count and gestational diabetes mellitus, the interpretation of their results may be influenced by unanticipated confounding or reverse causality, making it difficult to establish definitive causal relationships. For instance, Tian Songbai et al20 observed that pregnant women with GDM had noticeably higher basophil levels than those with normal pregnancies. However, in a study conducted by Yonggang Zhang et al,21 a group of 1860 pregnant women were examined. The researchers discovered that pregnant women with gestational diabetes had notably lower basophil counts compared to pregnant women without the condition.
Mendelian randomisation (MR) studies have developed as a novel approach to causal inference in recent years.22 The method employs genetic variation as instrumental variables (IVs), similar to that of randomised controlled trials, to offer dependable evidence of causal connections between exposure and illness outcomes. There have been no documented magnetic resonance (MR) studies investigating the relationship between basophil count and GDM risk. In this study, two-sample MR analysis was performed using genome-wide association study (GWAS) summary statistics from IEU Open GWAS to investigate the causal effect between basophil count and gestational diabetes mellitus.
Materials and Methods
Study Design
First, we examined the effects of genetic variants strongly associated with basophil count as IV to investigate their effects on GDM and glycemic traits. We also performed the directional validation of MR analysis to assess the effect of GDM predisposition on basophil counts to rule out the possibility of reverse causation.
In a TSMR study, the IV should fulfill three fundamental requirements:23 (i) It is expected that a robust association will be exhibited with regard to the exposure; (ii) It is essential that the variables in question are not confounded by any factors that could potentially impact the link between exposure and outcome; (iii) It is only through exposure that they should have any impact on the outcome (Figure 1). The relationship between basophil count (exposure factor) and the incidence of gestational diabetes mellitus (GDM) was assessed using an instrumental variable, a single nucleotide polymorphism (SNP).
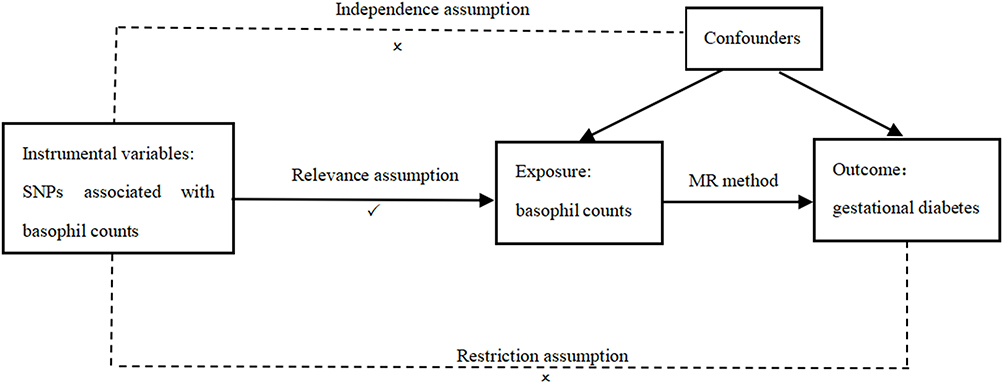 |
Figure 1 Schematic of the assumptions of the two-sample Mendelian randomization study. |
After confirming the cause-and-effect connection between basophils and gestational diabetes, Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses and Protein-Protein Interaction (PPI) networks were used to gain a deeper understanding of their impact on disease.
Data Sources
Selection of Appropriate IVs for MR Analysis from Two Different GWASs
Relevant GWAS datasets were collected from the IEU Open GWAS project website (https://gwas.mrcieu.ac.uk). The two sets of data ultimately come from the same population of European women. The dataset on basophil counts included a sample size of 170,143, with 109 SNPs. The dataset for GDM comprised 123,579 samples, with each sample including 109 SNPs. The study was a secondary analysis based on publicly available genome-wide association study (GWAS) data. No additional ethical approval was required (Data Availability: The datasets generated and analyzed during the current study are available on the IEU Open GWAS database (https://gwas.mrcieu.ac.uk/). They provide the Platform summary data from genome-wide association studies (GWAS Data). The study was a secondary analysis based on publicly available genome-wide association study (GWAS) data. This study is exempt from approval based on Article 32, Item 1, of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects, issued by China on 18 February 2023. This study did not require access to any individual level data and therefore, does not require ethical approval and informed consent. Ethical approval for all the included studies was sought by the investigators in the original studies and is reported at the referenced publications).
Instrumental Variable Selection
The instrumental variable (IV) is selected using the following steps: First, single nucleotide polymorphisms (SNPs) with significant associations in GWAS were extracted as instrumental variables (p < 5×10−8). Subsequently, using a clustering method based on linkage disequilibrium (LD), we screened for LD-independent SNPs (r2 < 0.01, physical distance = 10,000 kb). SNPs associated with confounding factors and SNPs with an opposite effect were removed. We also performed directional validation of the MR analysis to assess the effect of GDM predisposition on basophil counts to rule out the possibility of reverse causation. Finally, the F statistic was computed to evaluate the efficacy of the SNPs with the formula beta2/se2; F-statistic ≥10 shows an association between the instrumental variable and exposure.24 SNPs with less statistical power (F-statistics < 10) were removed in the final analyses.
Two-Sample Mendelian Randomization Analysis
The inverse variance weighted (IVW) method use inverse variance to weight connections between SNPs and outcomes, making it an effective technique for estimating causal relationships between exposures and outcomes. Hence, we used the IVW method to evaluate the causal association between gestational diabetes mellitus (outcomes) and basophil counts (exposures). Mendelian Randomization-Egger regression method (MR-Egger regression method), weighted median method (WM method), simple mode method and weighted mode method were employed as complementary approaches to assess the connection. Furthermore, to assess the influence of individual SNPs, a leave-one-out sensitivity analysis was conducted. We also performed the Cochran’s Q statistic for MR-inverse-variance weighted and MR Egger analyses to test for heterogeneity. The MR-Egger regression method was used to test for horizontal pleiotropy.
These statistical methods were estimated using the Two Sample MR and Mendelian Randomisation R packages in R Studio 4.2.1. Statistical significance was defined as P <0.05.
GO, KEGG Enrichment and PPI Network
To analyse inflammatory cell function in relation to gestational diabetes, we used the Metascape database (https://metascape.org/gp/index.html#/main/step1) for KEGG pathway and GO term analysis. Statistical significance was considered to be P <0.05.
A PPI network is constructed using the STRING database (http://string-db.org/cgi/input.pl) to further evaluate the inflammatory cellular protein network associated with gestational diabetes. We then identified key targets and assessed their accuracy using Cytoscape software.
Results
Selection of Instruments
SNPs were used as IVs for basophil counts according to the variable screening criteria of this research tool, and 109 SNPs were ultimately selected as IVs. The F statistics ranged from 29.72 to 1108.21, indicating that the causal estimations were not biased by the weak IVs. Detailed information on the IVs used in the MR analyses was displayed in Table S1.
The Results of a Two-Sample Mendelian Randomization Study
We aimed to investigate the causal effect of basophil count levels on GDM incidence. According to IVW estimates, the odds ratios (ORs) of developing GDM for a 1 standard deviation (SD) increase in genetic basophil count were 0.84 (95% CI: 0.74–0.96, P = 0.01). Other results: MR-Egger method: OR = 0.85, 95% CI: 0.65–1.12, P 0.25; weighted median method: OR = 0.85, 95% CI: 0.69 ~1.05, P 0.13; simple model: OR = 0.77, 95% CI: 0.51–1.15, P 0.20; weighted model: OR = 0.84, 95% CI: 0.67–1.06, P 0.15. See Figure 2
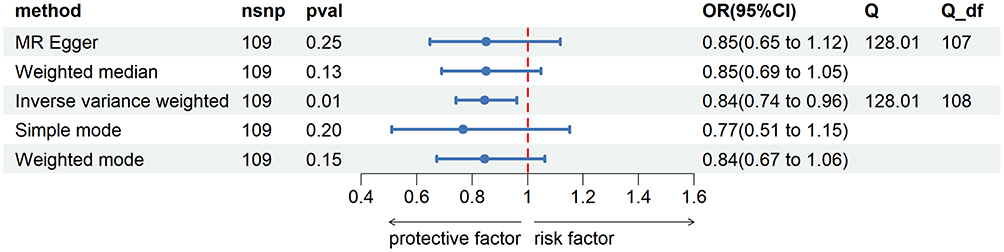 |
Figure 2 Estimate of the MR analysis for the basophil count and gestational diabetes mellitus. Note: The odds ratio (OR) and the corresponding 95% confidence interval (CI) illustrate the change in the standard deviation (SD) of basophil count with each incremental change in log odds related to gestational diabetes mellitus (GDM). |
There was no heterogeneity between IVs using Cochran’s Q test for MR-Egger regression and the IVW method (p > 0.05), see Table 1. The intercept term of the MR-Egger regression was close to 0, while P > 0.05 (Egger intercept/b=−0.161, P = 0.249), indicating that IVs do not have horizontal pleiotropy, see Table 2. The scatter plots, forest plots, and funnel plots are shown in Figure 3A–C. The results of the leave-one-out analysis indicated that no SNP significantly altered the IVW association, see Figure 3D.
 |
Table 1 Results of the MR Heterogeneity Analysis |
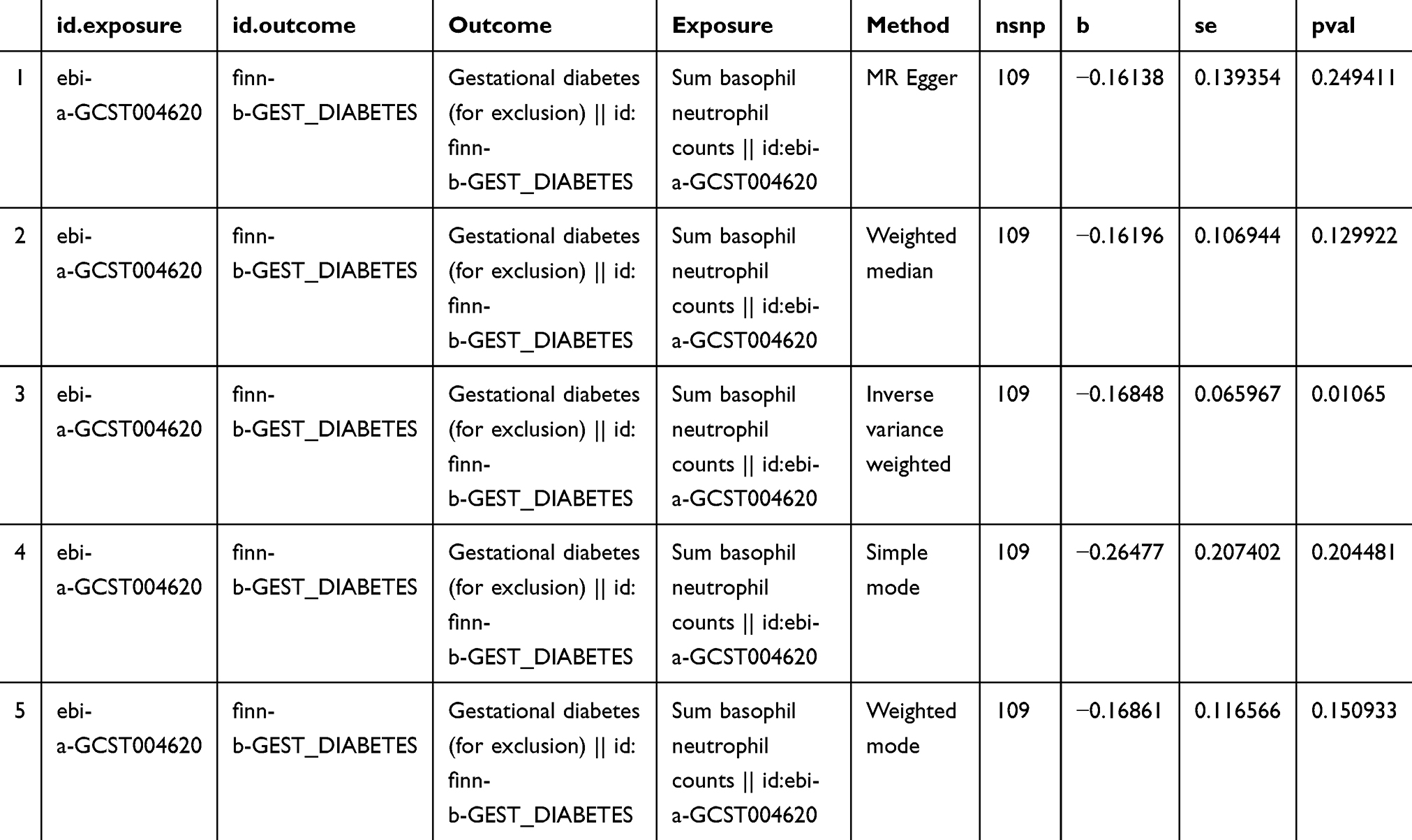 |
Table 2 Results of the MR Pleiotropy Analysis |
 |
Figure 3 Scatter plot, Forest plot, funnel plot, and leave-one-out analysis of MR analysis for the causal effect of basophil count on gestational diabetes mellitus. (A) Scatter plot, (B) forest plot, (C) funnel plot, and (D) Leave-one-out analysis. |
To further understand the source of observational associations between basophil count and gestational diabetes, we also explored the relationship of GDM with basophil count in the reverse direction using the directional validation of MR analysis. Overall, the results of directional validation of MR analysis did not support the reverse causation of GDM predisposition on the levels of basophil count. Detailed information on the results of the directional validation of the MR analysis is given in Table S2.
GO, KEGG Enrichment and PPI Network
In GO enrichment analysis, the top five biological process terms were “endocytosis”, “response to chemokines”, “positive regulation of cell adhesion”, “cholesterol efflux”, and “leukocyte homeostasis”. The four biological components that showed the highest enrichment were “late endosomal membrane”, “protein complexes involved in cell adhesion”, “side of membrane”, and “fiber center”. The top five enriched terms in molecular functions were predominantly related to “peptidase activator activity”, “cytokine receptor binding”, “protein homodimerization activity”, “glycosaminoglycan binding”, and “integrin binding” (Figure 4A–C).
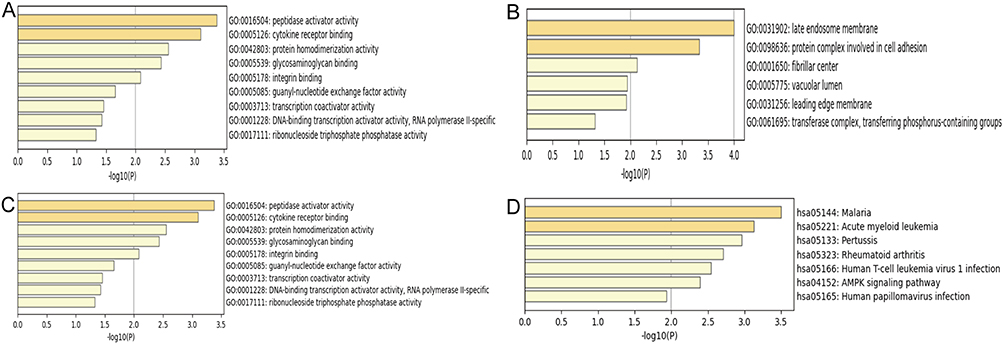 |
Figure 4 Bar graphs of GO and KEGG analysis. (A) GO biological processes, (B) GO cellular components, (C) GO molecular functions, and (D) KEGG pathway enrichment. |
Additionally, the KEGG pathway enrichment study revealed five pathways exhibiting considerable enrichment: “malaria”, “acute myeloid leukemia”, “rheumatoid arthritis”, “human T-cell leukemia virus type 1 infection”, and “AMPK signaling pathway” (Figure 4D).
Following the upload of the inflammatory cells linked to gestational diabetes onto the STRING online database, the protein–protein interaction network was generated using Cytoscape software (Figure 5A). The top 5 hub genes were then identified with the help of CytoHubba software, namely FLT3, CXCL12, HNF4A, TERT, and CDK6 (Figure 5B and C).
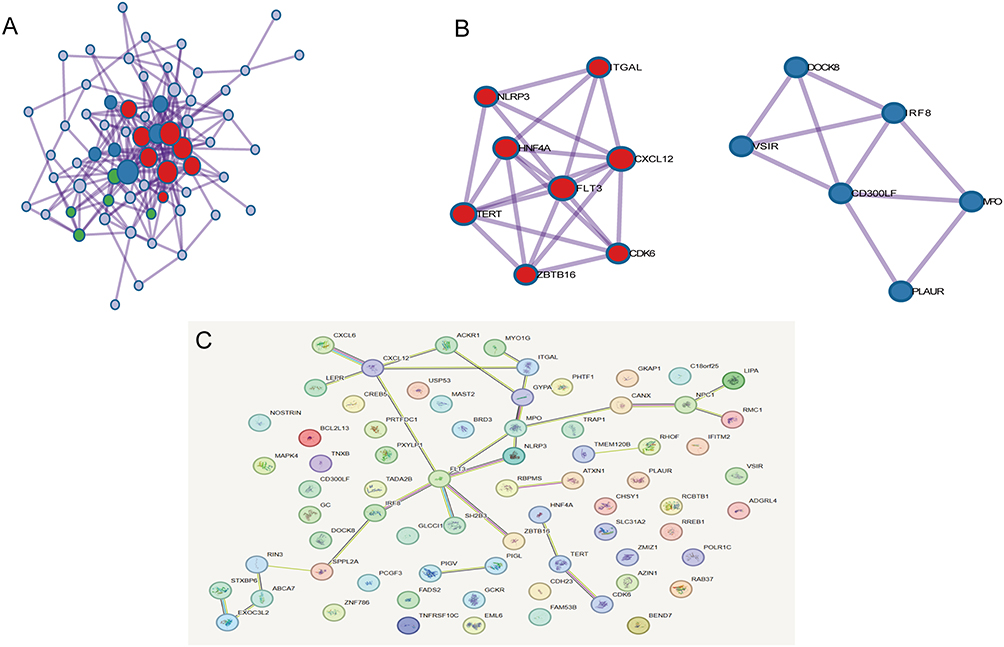 |
Figure 5 PPI network of inflammatory factors associated with gestational diabetes mellitus. Notes: (A) PPI network for the inflammatory cytokines associated with GDM. (B and C) The most significant top 5 inflammatory cytokines in the PPI network. |
Discussion
Based on the findings of this investigation, high basophil counts are associated with an increased risk of gestational diabetes mellitus (GDM). The results of the directional validation of the MR analysis did not support the reverse causation of GDM predisposition on basophil count levels. These findings suggest that basophil imbalance may lead to GDM developing.
Conclusions of this investigation agree with those of An Yan et al,25 who conducted a study in China involving 446 pregnant women (including 112 were GDM and 334 were healthy). The basophil levels of the women in the GDM group were significantly higher than those in the healthy group, and higher basophil counts in the early stages of pregnancy were linked to the development of GDM. Jin Xiaohui et al26 additionally discovered a positive relationship between basophils and GDM in during early and mid-pregnancy.
The etiology of GDM remains incompletely elucidated. Recent research indicates that insulin resistance and pancreatic β cell dysfunction are fundamental to its development.27 Meanwhile, the body’s inflammatory response is strongly linked to the development and progression of insulin resistance, making it an important factor in the occurrence of GDM.12 Lekva et al11 and Lorenzo et al12 have found that chronic inflammation may promote the onset of GDM by increasing insulin resistance, affecting insulin signaling, and promoting beta cell dysfunction. Basophils are one of the markers of inflammation and can reflect chronic inflammatory processes. Several studies17–19 have shown that basophils have the function of initiating and recruiting other inflammatory cells and producing cytokines and chemokines, thus playing an important role in promoting the development of chronic inflammation, insulin resistance and diabetes. Inflammatory responses can lead to local and systemic insulin resistance through several pathways, including the SMAD pathway,28,29 the AMPK pathway30 and the NF-κB pathway.31 Basophils are also involved in the SMAD pathway,32 the AMPK pathway,33 and the NF-κB pathway,34 so basophils may be biologically linked to GDM through inflammation and insulin resistance.
Meanwhile, genomics has identified five central genes associated with gestational diabetes, namely FLT3, CXCL12, HNF4A, TERT and CDK6.
FLT3 was a receptor-type tyrosine kinase that was typically found in blood stem or initial cells and was activated by ligands on the cell membrane. FLT3 ligand bound to and activated FLT3 and promoted cell survival, proliferation, and differentiation through PI3K, RAS, and STAT signaling pathways.35 It has also been shown that FLT3 is involved in the SMAD36 and AMPK pathways.37 Those signaling routes played a part in how insulin was used and signaling transduction, which further caused GDM.38–41 Therefore, it could be inferred that FLT3 may induce gestational diabetes mellitus through the above signaling pathways. Recent research has shown that giving the Flt3 ligand to NOD mice can greatly slow the development of insulitis and diabetes.42 We think that FLT3 might be connected to the inflammation response and insulin resistance signaling pathways through basophils, which could eventually lead to GDM. But because there is not a lot of research out there right now, no one knows for sure what part FLT3 plays in the pathological process of GDM by basophils. More research is needed.
CXCL12, also known as stromal-derived cell factor-1, is a member of the chemokine family that is expressed in various tissues and cells and is constitutively expressed by bone marrow stromal cells.43 Multiple studies had shown44,45 that CXCL12 was closely related to the development of GDM. Previous studies have suggested that CXCL12 may be important for basophil recruitment and activation.46,47 It can be speculated that CXCL12 regulates the inflammatory response through the recruitment and activation of basophils, which in turn leads to GDM. However, this has not yet been researched and it remains necessary to investigate further.
HNF4α (hepatocyte nuclear factor 4α),48 a transcription factor belonging to the nuclear hormone receptor superfamily, is expressed in the liver, kidney, intestines, and pancreas.48–50 The study by Latisha Love-Gregory et al51 suggested that HNF 4A might influence insulin secretion by affecting the activity of KATP channels. Ren et al’s study52 demonstrates that SIRT2 overexpression in hepatocytes enhances HNF4α protein expression and positively influences hepatic steatosis, insulin resistance, and inflammatory responses in obese subjects. The research conducted by Wang et al53 indicates that HNF4α significantly influences the regulation of several liver-specific genes; HNF4α may be involved in regulating the cytokine-induced inflammatory response. Mohlke and Boehnke et al54 discovered that variations in the HNF4α gene correlate with an increased risk of acquiring type 2 diabetes. However, Shaat et al55 conducted genotyping on 648 women diagnosed with GDM and 1232 non-diabetic pregnant women in Sweden. They could not find any connection between GDM and variations of the HNF4A gene. We hypothesize that HNF4α initiates an inflammatory response by modulating basophil expression, thus contributing to the onset of gestational diabetes. Nevertheless, no studies have currently established a direct correlation between HNF4α and basophils, necessitating more investigation
TERT was the primary catalytic subunit of the telomerase enzyme, extensively present in different types of cells, and was an important structure for maintaining telomeres and chromosome stability. Research indicates that chronic diseases including type 1 and type 2 diabetes are associated with telomere instability or dysregulation of telomerase activity.56 Liu et al57 demonstrated that TERT had been linked to impaired pancreatic β-cell activity in individuals diagnosed with T2D. The catalytic subunit of telomerase, TERT, may modulate NF-κB-dependent inflammation through its interaction with p65 and by possessing NF-κB target genes.58 Basophils also initiate inflammatory responses via the NF-κB pathway.34 Since GDM and T2D shared common pathophysiological mechanisms,59,60 we could infer that TERT may influence basophil expression by altering the NF-κB pathway, thereby initiating an inflammatory response and insulin resistance, which ultimately contributes to the onset of gestational diabetes. However, no studies have demonstrated a direct relationship between TERT and basophils, indicating the need for additional research.
Cyclin-Dependent Kinase 6 (CDK6) is an important regulatory protein of the cell cycle and metabolism,61 controlling cell growth, development, metabolism, inflammation and apoptosis.62 CDK6 and CDK4 frequently collaborate to promote the transition from G1 to S phase of the cell cycle.63 It had been shown that CDK4 deficiency led to defective pancreatic cell proliferation, resulting in insulin-deficient diabetes,64 while aberrantly expressed or activated CDK6 can lead to chronic inflammation via NF-κB.65–67 In addition, its overexpression is associated with reprogramming of glucose metabolism via alternative pathways and apoptosis.62 We hypothesise that CDK6 may ultimately trigger the development of GDM through an inflammatory response induced by the NF-κB pathway involved in cellular glucose metabolism and basophil involvement, but further studies are needed to prove this.
This is the first study that we know of that uses publicly available GWAS summary data for a TSMR analysis to look into the link between basophil counts and gestational diabetes. The following are the advantages of this study: First, MR can infer causal links between exposures and effects and is not affected by confounding variables. Second, the investigation utilized GWAS data collected from an extensive population sample, hence enhancing the credibility of the findings. Finally, this study conducted sensitivity analyses based on multiple methods to ensure the credibility of the results. Despite the compelling results of this study, some limitations remain. Firstly, there may be some unknown confounders in the dataset used for the study, which could have some effect on the results. Secondly, the findings of this investigation may be difficult to apply to other populations, as the study population was mainly concentrated in European populations, with significant differences in gene distribution between different ethnic groups. Therefore, future studies need to combine more genetic markers and clinical data to fully understand the pathogenesis of GDM.
Overall, our study employed two-sample MR analyses to describe the cause-and-effect chain between basophil counts and GDM, focusing on filling gaps in knowledge about this causal chain. Based on the MR analysis, there is strong proof that basophil counts are linked to gestational diabetes. However, more large-scale investigations are necessary to validate our findings and to further the search for new strategies to prevent gestational diabetes by targeting basophils.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. American Diabetes Association Professional Practice Committee. 2. Diagnosis and classification of diabetes: standards of care in diabetes-2024. Diabetes Care. 2024;47(Suppl 1):S20–S42. doi:10.2337/dc24-S002
2. Huvinen E, Eriksson JG, Koivusalo SB, et al. Heterogeneity of gestational diabetes (GDM) and long-term risk of diabetes and metabolic syndrome: findings from the RADIEL study follow-up. Acta Diabetol. 2018;55(5):493–501. doi:10.1007/s00592-018-1118-y
3. Lee KW, Ching SM, Ramachandran V, et al. Prevalence and risk factors of gestational diabetes mellitus in Asia: a systematic review and meta-analysis. BMC Pregnancy Childbirth. 2018;18(1):494. doi:10.1186/s12884-018-2131-4
4. Wang H, Li N, Chivese T, et al. IDF diabetes atlas: estimation of global and regional gestational diabetes mellitus prevalence for 2021 by International Association of Diabetes in Pregnancy Study Group’s Criteria. Diabet Res Clin Pract. 2022;183:109050. doi:10.1016/j.diabres.2021.109050
5. Wei YM, Yang HX. Development and prospect of research on gestational diabetes mellitus in China. Chin J Perinat Med. 2018;21(4):218–220.
6. Sweeting A, Wong J, Murphy HR, Ross GP. A clinical update on gestational diabetes mellitus. Endocr Rev. 2022;43(5):763–793. doi:10.1210/endrev/bnac003
7. Giannakou K, Evangelou E, Yiallouros P, et al. Risk factors for gestational diabetes: an umbrella review of meta-analyses of observational studies. PLoS One. 2019;14(4):e0215372. doi:10.1371/journal.pone.0215372
8. Plows JF, Stanley JL, Baker PN, Reynolds CM, Vickers MH. The pathophysiology of gestational diabetes mellitus. Int J Mol Sci. 2018;19(11):3342. doi:10.3390/ijms19113342
9. Yang H, Zhu C, Ma Q, Long Y, Cheng Z. Variations of blood cells in prediction of gestational diabetes mellitus. J Perinat Med. 2015;43(1):89–93. doi:10.1515/jpm-2014-0007
10. Kawai T, Autieri MV, Scalia R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am J Physiol Cell Physiol. 2021;320(3):C375–c391. doi:10.1152/ajpcell.00379.2020
11. Lekva T, Norwitz ER, Aukrust P, Ueland T. Impact of systemic inflammation on the progression of gestational diabetes mellitus. Curr Diab Rep. 2016;16(4):26. doi:10.1007/s11892-016-0715-9
12. Lorenzo PI, Martín-Montalvo A, Cobo Vuilleumier N, Gauthier BR. Molecular modelling of Islet β-Cell adaptation to inflammation in pregnancy and gestational diabetes mellitus. Int J Mol Sci. 2019;20(24):6171. doi:10.3390/ijms20246171
13. Germolec DR, Shipkowski KA, Frawley RP, Evans E. Markers of Inflammation. Methods Mol Biol. 2018;1803:57–79.
14. Herrero-Cervera A, Soehnlein O, Kenne E. Neutrophils in chronic inflammatory diseases. Cell Mol Immunol. 2022;19(2):177–191. doi:10.1038/s41423-021-00832-3
15. Miyake K, Ito J, Karasuyama H. Role of basophils in a broad spectrum of disorders. Front Immunol. 2022;13:902494. doi:10.3389/fimmu.2022.902494
16. Siracusa MC, Comeau MR, Artis D. New insights into basophil biology: initiators, regulators, and effectors of type 2 inflammation. Ann N Y Acad Sci. 2011;1217:166–177. doi:10.1111/j.1749-6632.2010.05918.x
17. Nakashima C, Otsuka A, Kabashima K. Recent advancement in the mechanism of basophil activation. J Dermatol Sci. 2018;91(1):3–8. doi:10.1016/j.jdermsci.2018.03.007
18. Kanazawa I, Tanaka S, Sugimoto T. The association between osteocalcin and chronic inflammation in patients with type 2 diabetes mellitus. Calcif Tissue Int. 2018;103(6):599–605. doi:10.1007/s00223-018-0460-y
19. Mzimela NC, Ngubane PS, Khathi A. The changes in immune cell concentration during the progression of pre-diabetes to type 2 diabetes in a high-fat high-carbohydrate diet-induced pre-diabetic rat model. Autoimmunity. 2019;52(1):27–36. doi:10.1080/08916934.2019.1575820
20. Tian SB, Sui Z, Zhao XQ, Shao CH, Yan H. Correlation of white blood cell count and its classification with gestational diabetes mellitus. China Maternal Child Health. 2017;32(18):4336–4338.
21. Zhang Y, Zhang Y, Zhao L, Shang Y, He D, Chen J. Distribution of complete blood count constituents in gestational diabetes mellitus. Medicine. 2021;100(23):e26301. doi:10.1097/MD.0000000000026301
22. Walker VM, Zheng J, Gaunt TR, Smith GD. Phenotypic causal inference using genome-wide association study data: Mendelian randomization and beyond. Annu Rev Biomed Data Sci. 2022;5:1–17. doi:10.1146/annurev-biodatasci-122120-024910
23. Emdin CA, Khera AV, Kathiresan S. Mendelian randomization. JAMA. 2017;318(19):1925–1926. doi:10.1001/jama.2017.17219
24. Burgess S, Thompson SG. Avoiding bias from weak instruments in Mendelian randomization studies. Int J Epidemiol. 2011;40(3):755–764. doi:10.1093/ije/dyr036
25. An Y, Guo YY, Sun S. Predictive value of fasting blood glucose and white blood cell count in early pregnancy for gestational diabetes mellitus. China Maternal Child Health. 2018;33(12):2704–2707.
26. Jin XJ. Analysis of changes in routine blood parameters during the third trimester of pregnancy and their correlation with gestational diabetes mellitus and subclinical nail function abnormalities[D/OL]. Shenyang: China Medical University; 2023. Available from: https://kns.cnki.net/kcms2/article/abstract?v=01ddXewXOSCgoAlRlKAvTcQKTytCWwxlzeor-FCzG7GIKuISH26VyMe_3in5AGYyL35FfM7kWY7oBXoGp82fGm1j000mSvtTIIlCfnayGrtF0NFC9nbNig==&uniplatform=NZKPT&language=gb.
27. Powe CE, Allard C, Battista MC, et al. Heterogeneous contribution of insulin sensitivity and secretion defects to gestational diabetes mellitus. Diabetes Care. 2016;39(6):1052–1055. doi:10.2337/dc15-2672
28. El-Gohary Y, Tulachan S, Wiersch J, et al. A smad signaling network regulates islet cell proliferation. Diabetes. 2014;63(1):224–236. doi:10.2337/db13-0432
29. Wang HL, Wang L, Zhao CY, Lan HY. Role of TGF-beta signaling in beta cell proliferation and function in diabetes. Biomolecules. 2022;12(3). doi:10.3390/biom12030373
30. Liong S, Lappas M. Activation of AMPK improves inflammation and insulin resistance in adipose tissue and skeletal muscle from pregnant women. J Physiol Biochem. 2015;71(4):703–717. doi:10.1007/s13105-015-0435-7
31. Nguyen-Ngo C, Jayabalan N, Salomon C, Lappas M. Molecular pathways disrupted by gestational diabetes mellitus. J Mol Endocrinol. 2019;63(3):R51–r72. doi:10.1530/JME-18-0274
32. Dou L, Wang W, Wang J, et al. miR-3934 regulates the apoptosis and secretion of inflammatory cytokines of basophils via targeting RAGE in asthma. Allergy Asthma Clin Immunol. 2022;18(1):66. doi:10.1186/s13223-022-00704-z
33. Iype J, Odermatt A, Bachmann S, Coeudevez M, Fux M. IL-1β promotes immunoregulatory responses in human blood basophils. Allergy. 2021;76(7):2017–2029. doi:10.1111/all.14760
34. Dera A, Rajagopalan P, Ahmed I, Alfhili M, Alsughayyir J, Chandramoorthy HC. Thymoquinone attenuates IgE-mediated allergic response via pi3k-Akt-NFκB pathway and upregulation of the Nrf2-HO1 axis. J Food Biochem. 2020;44(6):e13216. doi:10.1111/jfbc.13216
35. Grafone T, Palmisano M, Nicci C, Storti S. An overview on the role of FLT3-tyrosine kinase receptor in acute myeloid leukemia: biology and treatment. Oncol Rev. 2012;6(1):e8. doi:10.4081/oncol.2012.e8
36. Yoon JH, Bae E, Nagafuchi Y, et al. Repression of SMAD3 by STAT3 and c-Ski induces conventional dendritic cell differentiation. Life Sci Alliance. 2024;7(9):e201900581. doi:10.26508/lsa.201900581
37. Ma W, Liang F, Zhan H, et al. Activated FMS-like tyrosine kinase 3 ameliorates angiotensin II-induced cardiac remodelling. Acta Physiol. 2020;230(2):e13519. doi:10.1111/apha.13519
38. Zhang Y, Liu Y, Shi Y, et al. Upregulation of MMPs in placentas of patients with gestational diabetes mellitus: involvement of the PI3K/Akt pathway. Heliyon. 2024;10(12):e32518. doi:10.1016/j.heliyon.2024.e32518
39. Das S, Chattopadhyay D, Chatterjee SK, et al. Increase in PPARγ inhibitory phosphorylation by Fetuin-A through the activation of Ras-MEK-ERK pathway causes insulin resistance. Biochim Biophys Acta Mol Basis Dis. 2021;1867(4):166050. doi:10.1016/j.bbadis.2020.166050
40. Li L, Zhang Z, Li H, et al. Research progress on the STAT signaling pathway in pregnancy and pregnancy-associated disorders. Front Immunol. 2023;14:1331964. doi:10.3389/fimmu.2023.1331964
41. Bao D, Zhuang C, Jiao Y, Yang L. The possible involvement of circRNA DMNT1/p53/JAK/STAT in gestational diabetes mellitus and preeclampsia. Cell Death Discov. 2022;8(1):121. doi:10.1038/s41420-022-00913-w
42. Chilton PM, Rezzoug F, Fugier-Vivier I, et al. Flt3-ligand treatment prevents diabetes in NOD mice. Diabetes. 2004;53(8):1995–2002. doi:10.2337/diabetes.53.8.1995
43. Pizzolo F, Castagna A, Olivieri O, et al. Basophil blood cell count is associated with enhanced factor II plasma coagulant activity and increased risk of mortality in patients with stable coronary artery disease: not only neutrophils as prognostic marker in ischemic heart disease. J Am Heart Assoc. 2021;10(5):e018243. doi:10.1161/JAHA.120.018243
44. Shi SW, Sun Y, Wei JY. Relationship between serum sFkn and CXCL12 levels and gestational diabetes mellitus and perinatal outcome in high-risk pregnant women. Int J Lab Med. 2024;45:603–607.
45. Liu H, Liu A, Kaminga AC, McDonald J, Wen SW, Pan X. Chemokines in Gestational Diabetes Mellitus. Front Immunol. 2022;13:705852. doi:10.3389/fimmu.2022.705852
46. Jinquan T, Jacobi HH, Jing C, et al. Chemokine stromal cell-derived factor 1alpha activates basophils by means of CXCR4. J Allergy Clin Immunol. 2000;106(2):313–320. doi:10.1067/mai.2000.108108
47. Yamaguchi M, Koketsu R, Suzukawa M, Kawakami A, Iikura M. Human basophils and cytokines/chemokines. Allergol Int. 2009;58(1):1–10. doi:10.2332/allergolint.08-RAI-0056
48. Sladek FM, Zhong WM, Lai E, Darnell JE. Liver-enriched transcription factor HNF-4 is a novel member of the steroid hormone receptor superfamily. Genes Dev. 1990;4(12b):2353–2365. doi:10.1101/gad.4.12b.2353
49. Miquerol L, Lopez S, Cartier N, Tulliez M, Raymondjean M, Kahn A. Expression of the L-type pyruvate kinase gene and the hepatocyte nuclear factor 4 transcription factor in exocrine and endocrine pancreas. J Biol Chem. 1994;269(12):8944–8951. doi:10.1016/S0021-9258(17)37059-X
50. Taraviras S, Monaghan AP, Schütz G, Kelsey G. Characterization of the mouse HNF-4 gene and its expression during mouse embryogenesis. Mech Dev. 1994;48(2):67–79. doi:10.1016/0925-4773(94)90017-5
51. Love-Gregory L, Permutt MA. HNF4A genetic variants: role in diabetes. Curr Opin Clin Nutr Metab Care. 2007;10(4):397–402. doi:10.1097/MCO.0b013e3281e3888d
52. Ren H, Hu F, Wang D, et al. Sirtuin 2 prevents liver steatosis and metabolic disorders by deacetylation of hepatocyte nuclear factor 4α. Hepatology. 2021;74(2):723–740. doi:10.1002/hep.31773
53. Wang Z, Bishop EP, Burke PA. Expression profile analysis of the inflammatory response regulated by hepatocyte nuclear factor 4α. BMC Genomics. 2011;12:128. doi:10.1186/1471-2164-12-128
54. Mohlke KL, Boehnke M. The role of HNF4A variants in the risk of type 2 diabetes. Curr Diab Rep. 2005;5(2):149–156. doi:10.1007/s11892-005-0043-y
55. Shaat N, Karlsson E, Lernmark A, et al. Common variants in MODY genes increase the risk of gestational diabetes mellitus. Diabetologia. 2006;49(7):1545–1551. doi:10.1007/s00125-006-0258-8
56. Liew CW, Holman A, Kulkarni RN. The roles of telomeres and telomerase in beta-cell regeneration. Diabetes Obes Metab. 2009;11(Suppl 4):21–29. doi:10.1111/j.1463-1326.2009.01103.x
57. Liu C, QiNan W, XiaoTian L, et al. TERT and Akt are involved in the Par-4-dependent apoptosis of Islet β cells in type 2 diabetes. J Diabetes Res. 2018;2018:7653904. doi:10.1155/2018/7653904
58. Cai Y, Sukhova GK, Wong HK, et al. Rap1 induces cytokine production in pro-inflammatory macrophages through NFκB signaling and is highly expressed in human atherosclerotic lesions. Cell Cycle. 2015;14(22):3580–3592. doi:10.1080/15384101.2015.1100771
59. Pervjakova N, Moen GH, Borges MC, et al. Multi-ancestry genome-wide association study of gestational diabetes mellitus highlights genetic links with type 2 diabetes. Hum Mol Genet. 2022;31(19):3377–3391. doi:10.1093/hmg/ddac050
60. Vounzoulaki E, Khunti K, Abner SC, Tan BK, Davies MJ, Gillies CL. Progression to type 2 diabetes in women with a known history of gestational diabetes: systematic review and meta-analysis. BMJ. 2020;369:m1361. doi:10.1136/bmj.m1361
61. Wang H, Nicolay BN, Chick JM, et al. The metabolic function of cyclin D3-CDK6 kinase in cancer cell survival. Nature. 2017;546(7658):426–430. doi:10.1038/nature22797
62. Yousuf M, Shamsi A, Anjum F, et al. Effect of pH on the structure and function of cyclin-dependent kinase 6. PLoS One. 2022;17(2):e0263693. doi:10.1371/journal.pone.0263693
63. Qi J, Ouyang Z. Targeting CDK4/6 for Anticancer Therapy. Biomedicines. 2022;10(3):685. doi:10.3390/biomedicines10030685
64. Rane SG, Dubus P, Mettus RV, et al. Loss of Cdk4 expression causes insulin-deficient diabetes and Cdk4 activation results in beta-islet cell hyperplasia. Nat Genet. 1999;22(1):44–52. doi:10.1038/8751
65. Buss H, Handschick K, Jurrmann N, et al. Cyclin-dependent kinase 6 phosphorylates NF-κB P65 at serine 536 and contributes to the regulation of inflammatory gene expression. PLoS One. 2012;7(12):e51847. doi:10.1371/journal.pone.0051847
66. Handschick K, Beuerlein K, Jurida L, et al. Cyclin-dependent kinase 6 is a chromatin-bound cofactor for NF-κB-dependent gene expression. Mol Cell. 2014;53(2):193–208. doi:10.1016/j.molcel.2013.12.002
67. Schmitz ML, Kracht M. Cyclin-dependent kinases as coregulators of inflammatory gene expression. Trends Pharmacol Sci. 2016;37(2):101–113. doi:10.1016/j.tips.2015.10.004
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.