Back to Journals » Clinical Interventions in Aging » Volume 11
Relationships between FTO rs9939609, MC4R rs17782313, and PPARγ rs1801282 polymorphisms and the occurrence of selected metabolic and hormonal disorders in middle-aged and elderly men – a preliminary study
Authors Rotter I ![]() , Skonieczna-Żydecka K, Kosik-Bogacka D
, Skonieczna-Żydecka K, Kosik-Bogacka D ![]() , Adler G, Rył A, Laszczyńska M
, Adler G, Rył A, Laszczyńska M ![]()
Received 20 August 2016
Accepted for publication 5 October 2016
Published 23 November 2016 Volume 2016:11 Pages 1723—1732
DOI https://doi.org/10.2147/CIA.S120253
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Richard Walker
Iwona Rotter,1 Karolina Skonieczna-Żydecka,2 Danuta Kosik-Bogacka,3 Grażyna Adler,2 Aleksandra Rył,4 Maria Laszczyńska4
1Department of Medical Rehabilitation, 2Department of Gerontobiology, 3Department of Biology and Medical Parasitology, 4Department of Histology and Developmental Biology, Pomeranian Medical University, Szczecin, Poland
Purpose: Metabolic disorders, including MetS, obesity, and lipid disorders, may be related to genetic factors. Metabolic disorders are associated with decreased TS levels in aging men. The aim of this study was to evaluate the relationship between FTO rs9939609, MC4R rs17782313, and PPARɣ rs1801282 polymorphisms and the presence of MetS and its components, the concurrent lipid disorders, as well as sex hormone concentrations.
Subjects and methods: This study involved 272 men of Caucasian descent aged 50–75 years. Lipid profile, including TCh, LDL, HDL, and TG, was evaluated by spectrophotometric method. Anthropometric measurements concerned WC and blood pressure. MetS was diagnosed according to the criteria of the IDF. Sex hormone profile, including TST, FTS, E2, DHEAS, and SHBG, was examined using enzyme-linked immunosorbent assay. Polymorphisms within FTO, MC4R, and PPARɣ genes were identified using polymerase chain reaction-restriction fragments length polymorphism.
Results: This study did not show links between the analyzed genetic polymorphisms and the presence of MetS, T2DM, HT, and obesity. However, higher concentrations of TCh and LDL were found in men with the FTO rs9939609 polymorphism in the recessive mode of inheritance (P=0.03 and P=0.05, respectively). Lower WC was found to be associated with MC4R rs17782313 gene inherited in the same model (P=0.005).
Conclusion: FTO rs9939609, MC4R rs17782313, and PPARɣ rs1801282 polymorphisms seem to have little effect on the incidence of metabolic malfunctions and no effect on androgen-related disorders in the examined middle-aged and elderly men.
Keywords: aging men, metabolic syndrome, sex hormones, genes, FTO, MC4R, PPARɣ
Introduction
Although the incidence of MetS is known to depend on lifestyle, diet, and physical activity,1,2 it is also suggested that genetic factors may play an important role. For example, the risk for MetS may be increased by the presence of common SNPs within the FTO-associated gene, the MC4R gene, and the PPAR gene. As the expression profile of the corresponding proteins is strictly dependent on the genetic information within their genes, functional implications of these three polymorphisms seem to be significant. However, their exact mechanisms of action are still not clear.
The FTO gene is located on chromosome 16 and encodes 2-oxoglutarate-dependent nucleic acid demethylase. The gene is expressed in many tissues, especially in the hypothalamus, responsible for the control of energy balance.3,4 Increases in the hypothalamic expression of FTO are associated with the regulation of energy intake but not with the feeding reward.5 Few SNPs within FTO gene were found to play a role in obesity and T2DM.6–9 To date, three SNPs (rs9939609, rs1121980, and rs1558902) have been associated with obesity and BMI in Caucasian and Hispanic Americans.3,6,7 The association of rs9939609 (A>T variant) with obesity was found to be the strongest, especially in the dominant model of inheritance.10 In addition, a link between the FTO rs9939609 polymorphism and obesity and MetS has been confirmed in many studied populations, including in Europe.11–16
The MC4R and leptin genes are essential in the hypothalamic regulation of appetite.17,18 The MC4R gene plays an essential role in the maintenance of energy balance and is stimulated by endogenous melanocortins.19 Its mutations account for 2.4%–4% of morbidly obese people; the polymorphisms of this gene have also been associated with obesity.20 Common variants within the MC4R gene have been reported as the second strongest association signal for common obesity in the GWAS.21 In adult Europeans, the MC4R rs17782313 (C>T variant) polymorphism has been widely studied and found to predispose to obesity.21 With regard to MetS, the studied association with the MC4R rs17782313 polymorphism disappears after adjusting for WC, indicating that the association with MetS is driven by the association with this factor.22 Moreover, studies on males proved that the rs17782313 polymorphism is associated with lower HDL concentration23 and elevated DBP.14,24
PPARs are nuclear receptors, participate in adipogenesis and lipid metabolism, regulate insulin sensitivity, and participate in transformations in the energy system.25 Of the three types of PPARs (α, β, and γ), the best known is PPARγ, with two isoforms – PPARγ1, which is expressed in many tissues, and PPARγ2, which is expressed almost exclusively in adipocytes.26 The PPARγ activation plays an important role in adipocyte differentiation and maturation, lipid metabolism and transport, and improving insulin sensitivity.27,28 The PPARγ gene is located on the short arm of chromosome 3 at 3p25 band. The most common PPARγ gene polymorphism is rs1801282 (C>G variant), resulting from proline being replaced with alanine.29 Some literature data indicate a relationship between the rs1801282 polymorphism and metabolic disorders.30 Healthy nonsmoking men carrying the mutant allele of rs1801282 polymorphism are at a high risk for MetS and insulin resistance.30 In a large study of the Caucasians, the rs1801282 was linked to WC in patients with T2DM.31 However, it must be remembered that other researchers found no such associations.32 In addition, PPARγ is involved in the activation of androgen receptor (AR), which may affect the action of androgens.33
Literature studies on men show significant relationships between MetS and hormonal changes. It is emphasized that a decrease in FTS and TST promotes the development of metabolic disorders, while disorders of carbohydrate metabolism, lipid metabolism and obesity contribute to hypogonadism.34–37 This indicates that these pathologies are interrelated.
Both cross-sectional and longitudinal epidemiological studies indicate that the level of SHBG is lower in men with MetS.38–40 Other researchers emphasize, however, that the relationships between MetS and either TS or SHBG are stronger in younger men.34,35 Dehydroepiandrosterone does not seem to play a significant role in the development of MetS in men.34,37,41 Finally, the conclusions of the EMAS support the supposition that E2 levels in aging men are not related to the development of MetS.42
Despite some knowledge on the interrelationships between metabolic and hormonal disorders in men, we still do not know which factors trigger them. It remains to be solved whether they are environmental or lifestyle factors, or genetic factors, and what comes first – metabolic or hormonal changes. Regarding the genetic factors that with a high probability are responsible for MetS and associated disorders, it seems important to find any potential links with hormonal disturbances.
We hypothesized that the selected genetic polymorphisms (FTO rs9939609, MC4R rs17782313, and PPARγ rs1801282) may contribute to the occurrence and increase in metabolic and androgen disorders in aging men. We aimed to examine the relationships between the FTO rs9939609, MC4R rs17782313, and PPARγ rs1801282 and the prevalence of MetS and related disorders, such as HT, T2DM, and obesity. In addition, we aimed to analyze the relationships between selected polymorphisms and the parameters of lipid profile, including TCh, LDL, HDL, TG, sex hormones, including TST, FTS, E2, DHEAS, SHBG, and anthropometric parameters, including BMI, WC, and ABP in middle-aged and elderly men, which may contribute to increased knowledge about the mutual relationships between metabolic and sex hormone disorders in aging men.
Subjects and methods
Study population
This study involved 272 Caucasian men aged 50–75 years (mean age 62±6.4 years) who voluntarily signed up after receiving information about the study from their doctors at primary health-care centers in the city of Szczecin (Poland). The exclusion criteria in this study included cancer treatment, receiving steroids (including T and dehydroepiandrosterone), thyroid disease, and receiving neuroleptics or antidepressants. The medical questionnaire showed that none of the participants exceeded a daily alcohol intake of 40 g. We excluded patients on slimming diets or showing above-average physical activity. The research material was collected from June 2013 to July 2014. Patients participating in the study filled in an original questionnaire (questions about lifestyle including nutritional habits, smoking, alcohol intake, demographics status, physical activity, diseases, and drug intake). The study was performed in accordance with the Declaration of Helsinki and approved by the Bioethics Committee of the Pomeranian Medical University in Szczecin (KB-0012/159/12). The men in the study were informed about the details of the research project and expressed their written consent to participate in the study. They also signed a separate agreement for storing their genetic material.
Among the respondents, we conducted a survey on demographic data and the presence of chronic diseases. Surveys revealed that 150 people suffered from HT (55.1%), 50 people had T2DM (18.4%), and statins were taken by 30 of the men (11.0%). None of the patients received fibrates.
MetS was diagnosed according to the 2005 IDF criteria for European men (WC ≥94 cm and at least two of the following: FPG ≥100 mg/dL or T2DM treatment; ABP ≥130/85 mmHg or HT treatment; HDL <40 mg/dL or dyslipidemia treatment; and TG ≥150 mg/dL or dyslipidemia treatment).
Anthropometric and blood pressure (BP) measurements
In order to accurately measure WC, a physician located patients’ upper hip bone and then placed a tape measure around bare stomach just above the hip bone. Each time, the measuring tape was parallel to the floor and adjacent to the patient’s body. During measurement, all the patients were asked to relax the abdomen and exhale. The measurements were performed under identical environmental conditions and by one person. BMI was calculated; we assumed that BMI in the range of 18.5–24.99 denotes normal weight, of 25–29.99 indicates overweight, and of ≥30 means obesity.
To assess the SBP and DBP, the sphygmomanometer was used. The smallest cuff size covering ~2/3 of the right upper arm and encircling the entire arm completely, was selected. BP was measured in a supine position after 15 minutes rest, and only one of the physicians performed all of the BP measurements using a standardized protocol.
Blood sampling
Blood was taken from the tested men on an empty stomach from an ulnar vein, between 7.30 am and 9.00 am. For the biochemical and hormonal assays, blood was drawn into a tube with a coagulator and gel separator and then centrifuged. For the genetic assays, blood was collected into tubes with ethylenediaminetetraacetic acid (anticoagulant). The sera were stored at −70°C.
In the blood serum, we determined TCh, LDL, HDL, TG, and FPG by spectrophotometric method, with the use of reagent kits (BIOLABO; Aqua-Med, Lodz, Poland). FPG was determined only in the nondiabetic men. MetS was diagnosed according to the criteria of the IDF from 2005.43 Enzyme-linked immunosorbent assay was used to determine serum concentrations of DHEAS, TST, FTS, E2, and SHBG with the use of reagent kits (DRG Medtek, Warsaw, Poland). Measurements were performed in triplets, and arithmetic mean (AM) was calculated. The measurements were rejected when the coefficient of variation was >10%.
Genotyping of the FTO, MC4R, and PPARγ
Genetic investigations were carried out in a laboratory of the Department of Gerontobiology, Pomeranian Medical University, Szczecin, Poland. Genomic DNA from peripheral blood leukocytes was extracted using an extraction kit (High Pure PCR Template preparation Kit; Roche, Mannheim, Germany) according to the manufacturer’s instructions. DNA samples were stored at 4°C for further analyses. Protocols for polymorphisms FTO rs9939609, MC4R rs17782313, and PPARγ rs1801282 followed previously published polymerase chain reaction-restriction fragments length polymorphism techniques,44–46 which were performed in a Hightech Thermocycler Cycler-Technology for Life (SensoQuest, Gottingen, Germany). The genotypes were determined by 2%–3% agarose gel electrophoresis (Agarose; Sigma-Aldrich, Munich, Germany) stained with DNA-star dye (Lonza, Inc, Rockland, ME, USA). The correctness of results was verified by repeating each mutated sample from PCR onward. All results were found to be reliable. For each of the loci, we performed analyses in overdominant models of inheritance followed by the adopted recessive models of inheritance (FTO: AA + TA vs TT; MC4R: CC + CT vs TT; and PPARγ: GG + CG vs CC).
Statistical analysis
Statistical analysis was performed using the StatView software, Version 5.0 (SAS Institute Inc., Cary, NC, USA). Continuous variables (ie, BMI, ABP, TST, FTS, SHGB, E2, DHEAS, TG, TCh, LDL, HDL, and FPG) were described by AM with standard deviation, median, and range. In the description of qualitative variables (ie, genotype, T2DM, HT, MetS, overweight and obesity, and statin treatment), we presented the number (n), which is also expressed as a percentage.
First, χ2 test was used to verify whether genotype frequencies fit to the Hardy–Weinberg (H–W) equilibrium. Then, an analysis of variance test was used to assess the associations between the genotypes FTO rs9939609, MC4R rs17782313, and PPARγ rs1801282 polymorphisms and the anthropometric indicators, hormonal and metabolic parameters. In the next step, the assessment of relationships between genotypes and the qualitative variables was performed using a χ2 test of independence. To evaluate whether the studied polymorphisms determined any of the anthropometric and metabolic indices, which were found to be significant in previous analyses, we used the logistic regression analysis for the determination of odds ratio (OR) and 95% confidence intervals. The level of significance was set at P≤0.05.
Results
The characteristics of the study group and descriptive analysis of the metabolic and hormonal parameters are listed in Table 1. Normal body weight was found in 65 patients (23.9%), overweight was found in 130 patients (47.8%), and obesity was found in 77 (28.3%). MetS was diagnosed in 144 patients (52.9%).
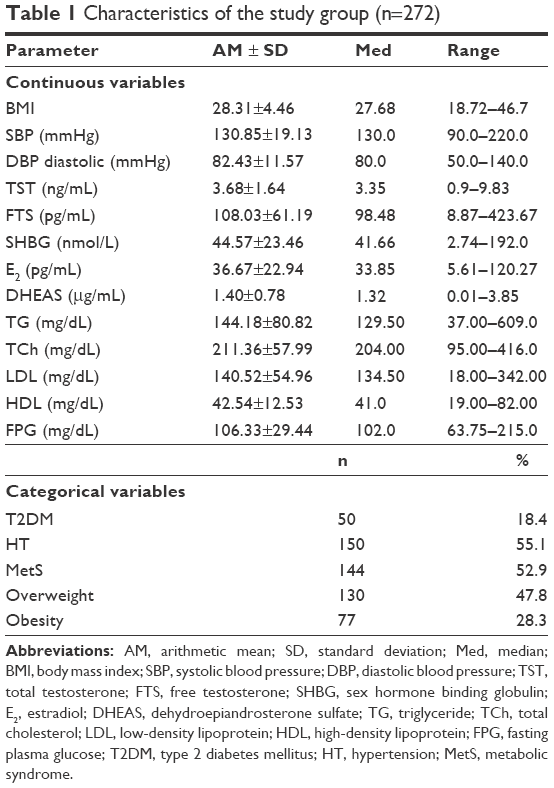 | Table 1 Characteristics of the study group (n=272) |
Genotype and allele distribution is listed in Table 2. They did not differ from H–W distribution (P>0.05).
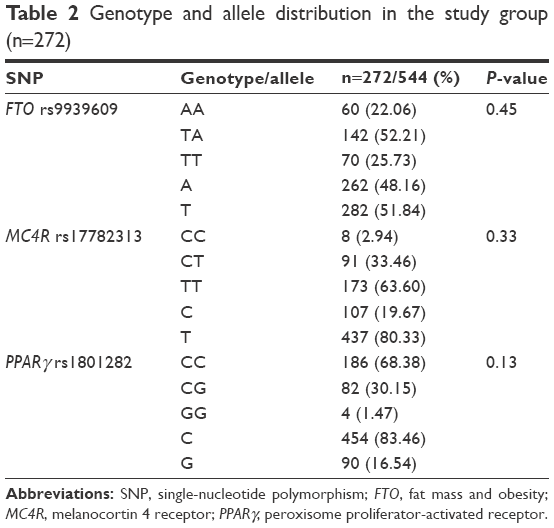 | Table 2 Genotype and allele distribution in the study group (n=272) |
We found no associations between FTO rs9939609, MC4R rs17782313, and PPARγ rs1801282 polymorphisms and the qualitative variables (MetS, T2DM, HT, obesity, and overweight), and also in the overdominant and recessive models of inheritance (Table 3).
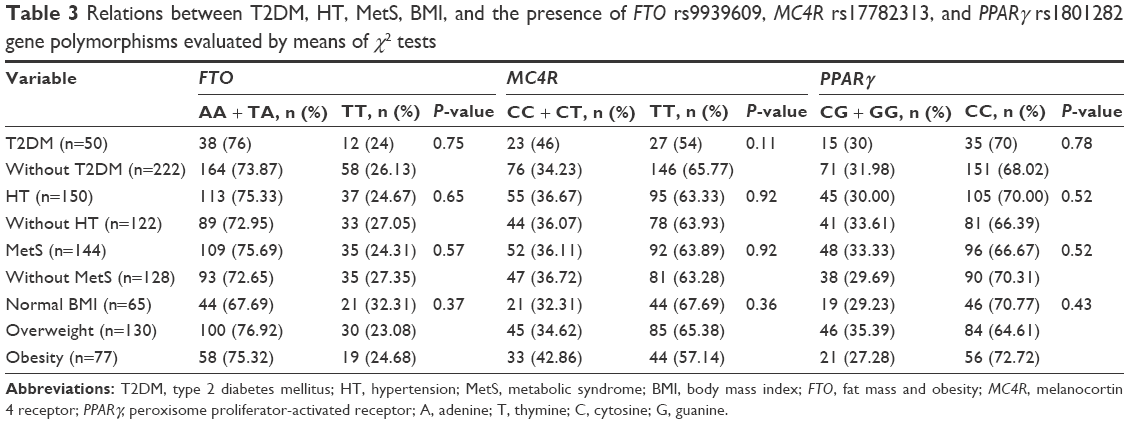 | Table 3 Relations between T2DM, HT, MetS, BMI, and the presence of FTO rs9939609, MC4R rs17782313, and PPARγ rs1801282 gene polymorphisms evaluated by means of χ2 tests |
In the case of continuous variables, we found that FTO rs9939609 polymorphism inherited only in the recessive pattern, which was associated with LDL concentration; the difference in LDL concentration was statistically significant (AA + AT: 136.18±51.57 mg/dL vs TT: 153.04±62.48 mg/dL; P=0.03). We also found that the presence of two mutant alleles was associated with higher TCh level (TT: 223.07±64.94 mg/dL and AA + AT: 207.3±54.9 mg/dL; P=0.05). There was no other evidence of links between the analyzed continuous variables and the FTO rs9939609 polymorphism in any of the analyzed models of inheritance. However, the association between the recessive genotype of rs9939609 polymorphism and SBP was close to statistical significance (P=0.07).
Analyzing the relationship between the continuous variables and the MC4R rs17782313 polymorphism, we demonstrated that men with the CC and CT genotypes had a significantly greater WC (CC + CT: 103.91±12.61 cm vs TT: 99.77±10.88 cm; P=0.005) (Table 4). For other continuous variables, we found no statistically significant relationships with the MC4R rs17782313 polymorphism in both models of inheritance.
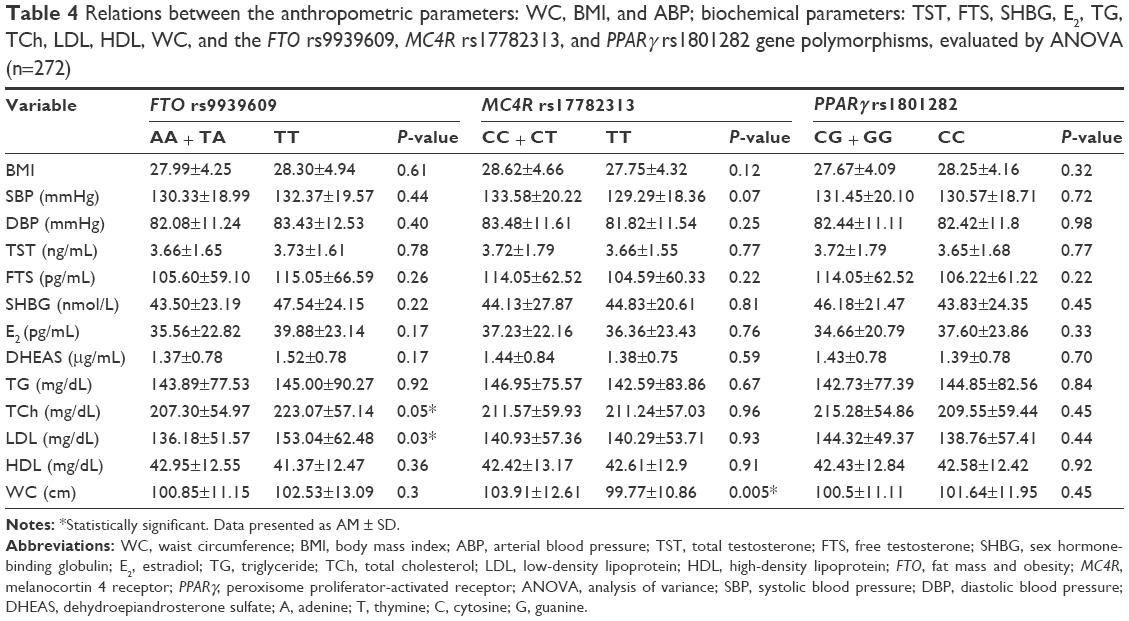 | Table 4 Relations between the anthropometric parameters: WC, BMI, and ABP; biochemical parameters: TST, FTS, SHBG, E2, TG, TCh, LDL, HDL, WC, and the FTO rs9939609, MC4R rs17782313, and PPARγ rs1801282 gene polymorphisms, evaluated by ANOVA (n=272) |
This study showed no connection between the PPARγ (rs1801282) polymorphism and the studied continuous variables.
In the final step of our experiments, we aimed to establish whether recessive genotypes of the polymorphisms determined any of the clinical conditions (T2DM, MetS, HT, overweight, and obesity), or continuous variables – anthropometric, hormonal, and metabolic indices – by means of logistic regression. We found no statistical significance with regard to the relationship between FTO rs9939609, MC4R rs17782313, and PPARγ (rs1801282) polymorphisms and any of the clinical conditions – MetS, T2DM, HT, overweight, and obesity (P>0.05). On the other hand, two mutant alleles of FTO rs9939609 polymorphism were found to be related to a minimal elevation of LDL, and the recessive genotype (TT homozygotes) of MC4R rs17782313 polymorphism was associated with a reduction in WC. The remaining anthropometric and biochemical parameters showed no statistical significance in the logistic regression model. The results are presented in Table 5.
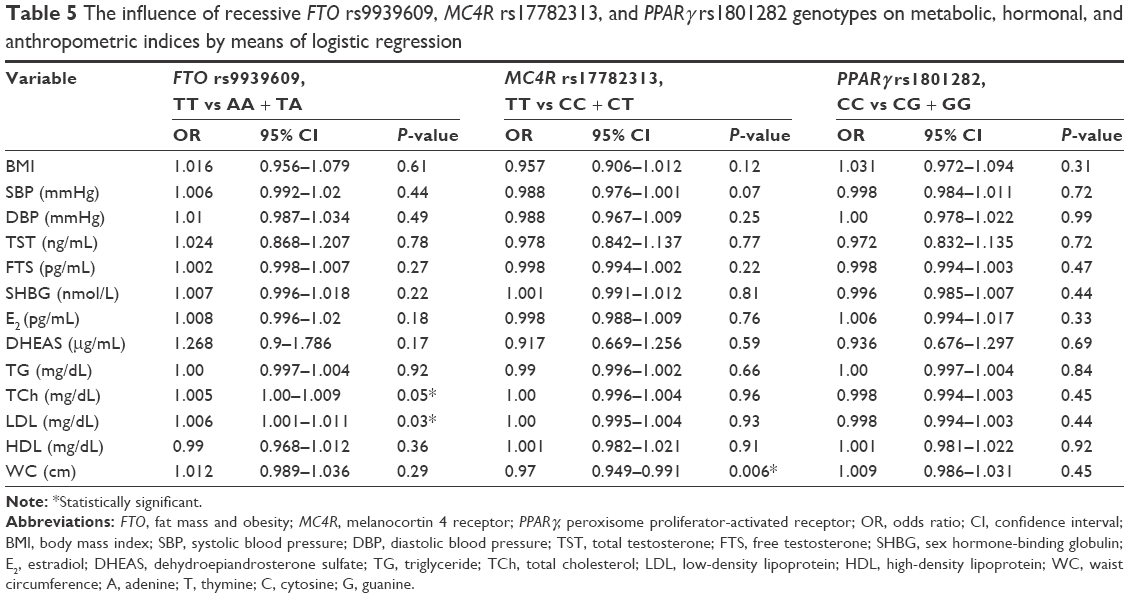 | Table 5 The influence of recessive FTO rs9939609, MC4R rs17782313, and PPARγ rs1801282 genotypes on metabolic, hormonal, and anthropometric indices by means of logistic regression |
Discussion
In this study, we found no relationship between the presence of FTO rs9939609, MC4R rs17782313, and PPARγ rs1801282 polymorphisms and the incidence of T2DM, HT, MetS, and obesity in Caucasian men aged 50–75 years, living in the city of Szczecin (Poland). We also found no links between the presence of these polymorphisms and the levels of the tested hormones (TST, FTS, E2, and DHEAS) and SHBG. With regard to biochemical parameters, we found a statistically significant association between the levels of TCh and LDL and the FTO rs9939609 polymorphism in the recessive model of inheritance. We proved that mutant homozygotes had higher TCh and LDL concentrations in comparison to other genotypes. In the group of anthropometric parameters, WC was statistically significantly associated with the MC4R rs17782313 polymorphism. Possessing two mutant alleles was found to be associated with lower WC in comparison to wild-type homozygotes and heterozygotes for the analyzed SNP.
Our results on the FTO rs9939609 polymorphism stand in contrast to a study in which non-Caucasian subjects demonstrated links between the FTO rs9939609 polymorphism and the occurrence of MetS.47 Moreover, that study also reported that an increased risk for MetS, especially in men, is associated with the presence of at least one wild-type allele. This regularity is also mentioned in a meta-analysis by Freathy et al,48 where allele A frequency correlated with lower HDL and higher TG concentrations and other components of MetS, but after adjusting for BMI, this relationship became statistically insignificant. In this study, we found no statistically significant relationships between either HDL or TG level and the FTO rs9939609 polymorphism. However, we did observe a relationship between the presence of the FTO rs9939609 polymorphism and both LDL and TCh concentrations. Our results indicated that the men with at least one allele of wild type had significantly lower levels of TCh and LDL compared to the men with the TT (mutant) genotype. Probably, lipid concentrations result primarily from the lifestyle of middle-aged and older men, including a high-calorie diet and low physical activity.
Carlos et al15 concluded that rs9939609 variant predisposes to obesity in Portuguese population aged 18–50 years. Woehning et al16 found that the polymorphism is linked to initial body weight, based on the results of their cross-sectional study in adults aged 18–72 years. Liguori et al49 proved that in a population of obese subjects of Italian origin, TA heterozygotes of rs9939609 are more prone to develop MetS (OR 2.53). A study by Jacobsson et al50 finds no relationship between the FTO rs9939609 polymorphism and BMI in elderly men from Sweden, indicating that its contribution to the incidence of obesity decreases with age. Zavatarri et al51 showed no association between the FTO rs9939609 polymorphism and the parameters of lipid metabolism and stressed that AA homozygotes are particularly predisposed to obesity. Population studies conducted by Gustavsson et al52 on Gothenburg residents confirmed that AA and TA genotypes have a greater WC and a higher BMI compared to TT genotype.
All the aforementioned results stand in contrast to our findings, showing the lack of relationship between metabolic parameters and the FTO rs9939609 polymorphism. It may have been due to the relatively small size of the group and the differences in selection of the participants in terms of age and gender. Our male population was aged between 50 and 75 years, while there is evidence that in people aged >65 years, the relationship between the FTO rs9939609 polymorphism and obesity is reduced, especially in men.53
With respect to the MC4R rs17782313 polymorphism, some researchers emphasize the role of C (wild-type) allele being associated with a higher risk for insulin resistance, T2DM, and HT incidence, independently of BMI.54 The relation between the MC4R rs17782313 polymorphism and obesity was also described by Xi et al.55 With regard to MetS, the association with the MC4R rs17782313 polymorphism disappears after adjusting for WC, indicating that the association with MetS is driven by the link with this factor,13 which shows that C allele may predispose to abdominal obesity. This is confirmed by our results where men with at least one C allele had a significantly higher WC, in addition to a higher BMI, although that difference was not statistically significant. Therefore, it can be assumed that the wild-type homozygous and heterozygous genotypes of MC4R rs17782313 polymorphism promote the accumulation of visceral fat. On the other hand, Marcadenti et al14 found that MC4R rs17782313 is positively associated with neck circumference and BMI in women and negatively associated with BP in men. However, the study was conducted in non-Caucasians, which makes the comparisons complicated.
In this study, we found no statistically significant associations between the PPARγ rs1801282 polymorphism and MetS, T2DM, HT, overweight, obesity, and anthropometric and biochemical parameters in men aged between 50 and 75 years. On the other hand, Tellechea et al30 found that in men from Argentina, with the exception of Caucasians, the rs1801282 polymorphism showed a relation with the occurrence of MetS diagnosed using the criteria of the National Cholesterol Education Program/Adult Treatment Panel III and with insulin resistance, especially in nonsmokers. However, those studies were conducted among younger men, so it can be assumed that environmental influences were lower. As in this study, Milewicz et al56 did not observe any association between the studied PPARγ rs1801282 polymorphism and MetS and metabolic parameters in postmenopausal women from Poland. Also studies conducted on an ethnically diverse group of people (Caucasian, South Asian, and African American) did not show any association between the PPARγ rs1801282 polymorphism and MetS.32 Dytfeld and Horst-Sikorska29 highlighted the role of diet in metabolic disorders in patients with the PPARγ rs1801282 polymorphism and modification of the genotype by environmental factors, which can lead to diverse phenotypes. On the other hand, Kruzliak et al31 conducted analyses in a group of >1000 Caucasians with T2DM and proved that the homozygous Slovene females with wild-type alleles had lower WC in comparison with homozygous females with mutated alleles. All in all, these findings were not replicated in this study, probably due to gender and age differences.
In this study on men, we found no relationship between the FTO rs9939609 polymorphism and the concentration of hormones DHEAS, E2, TST, FTS, and SHBG. In the available literature, we found no reports on the relationships between the FTO rs9939609 and MC4R rs17782313 polymorphisms and the levels of sex hormones in men. In this study, we also found no such relations.
Hainerová et al,57 based on the results of studies in young obese men with hypogonadism and G181D mutation in the MC4R gene, reported that obesity caused by the mutation in the MC4R rs17782313 polymorphism indirectly contributes to hormonal changes, as the adipose tissue is an active endocrine organ. In this study, we found no association between the PPARγ rs1801282 polymorphism and TST, FTS, E2, DHEAS, and SHBG concentrations, but the study included men aged >50 years, in whom metabolic disorders are mainly the result of lifestyle. Studies of Park et al58 in men have shown that the PPARγ polymorphisms play a significant role in tumorigenesis as a tumor suppressor and PPARγ agonists may have a beneficial effect in the treatment of prostate cancer. It is, therefore, necessary to conduct further studies on the possible associations between the PPARγ rs1801282 polymorphism and hypogonadism, prostatic hyperplasia, and prostate cancer.
Some studies59,60 have demonstrated the importance of dietary factors and physical activity level for the development of MetS. Lifestyle modification is effective in resolving MetS and reducing the severity of related abnormalities (FPG, WC, SBP, DBP, and TG) in patients with MetS.59 Although it does not necessarily influence any given risk factor as much as dedicated drugs, its benefit lies in a moderate reduction in all the metabolic risk factors.
Genetic factors in MetS are still under investigation. Our results and other researchers’ findings show that metabolic disorders in middle-aged and elderly people, including MetS and its components, are mainly the effect of lifestyle, including many years of improper eating habits and low physical activity, while genetic predisposition plays a much less significant role. Also in case of a singular SNP within the gene, it may be difficult to fully explain the risk for metabolic disorders and related traits.61 This phenomenon may be linked to potential gene–gene interactions. In case of singular SNP, other modest-risk variants may be the origin of the risk or control the expression of the particular risk factor.62
On the other hand, if an SNP may be an independent risk factor for a given clinical event, its effect can be observed with the passage of time. In complex diseases such as metabolic disorders, the effect of biallelic polymorphisms is most visible in the elderly, as the majority of independent environmental factors (eg, eating habits) remain at relatively stable levels. That is why this period of life is the most appropriate to observe the direct influence of genetic variation on the value of a given clinical parameter. Patients aged >50 years, ie, those we studied in this research, are therefore the optimal group in this context. However, it must be remembered that this study had significant limitations, as we included patients who reported voluntarily and we did not exclude patients with extreme obesity.
Due to the multitude of statistically insignificant results, we performed a post hoc power analysis using the G* Power software.63 The power of tests, which showed no statistical significance, was below the recommended level of 0.8.64 On the other hand, tests showing statistical significance had an acceptable power, reaching as high as 0.84. In our opinion, the small power of tests may be responsible for the great number of statistically insignificant results of the analysis. In order to increase the power of the tests, the sizes of the study groups would have to increase significantly. Besides, in a genetic association study, a reliable assessment of the prevalence of alleles increases with sample size (which means a greater number of alleles at a given locus). One should also take into account additional factors, such as the adopted model of inheritance, and especially the frequency of risk alleles at a given locus and the risk for the disease in a given population. In conclusion, a limited number of statistically significant results in this study may have been caused by the insufficient number of patients in the study, and thus the low power of statistical tests.
Conclusion
The FTO rs9939609 and MC4R rs17782313 gene polymorphisms have little significant relationships with metabolic health problems (T2DM, HT, overweight and obesity, and lipid disorders) and do not result in androgen disorders in aging men. However, our results extend the knowledge on genotype susceptibility for metabolic disturbances in relation to a specific geographical area of residence.
Abbreviations
MetS, metabolic syndrome; SNP, single-nucleotide polymorphism; FTO, fat mass and obesity-associated gene; MC4R, melanocortin 4 receptor gene; PPARγ, peroxisome proliferator-activated receptor gene; GWAS, Genome-Wide Association Study; T2DM, type 2 diabetes mellitus; HT, hypertension; TS, testosterone; TST, total testosterone; FTS, free testosterone; E2, estradiol; DHEAS, dehydroepiandrosterone sulfate; SHBG, sex hormone-binding globulin; TG, triglyceride; LDL, low-density lipoprotein; HDL, high-density lipoprotein; TCh, total cholesterol; ABP, arterial blood pressure; SBP, systolic blood pressure; DBP, diastolic blood pressure; FPG, fasting plasma glucose; WC, waist circumference; BMI, body mass index; EMAS, European Male Aging Study; IDF, International Diabetes Federation.
Acknowledgment
The study was financed as research project no FSN (stimulation fund research) WNoZ 321-10/13 by the Pomeranian Medical University in Szczecin, Poland.
Author contributions
IR designed the study and collected the data. IR, KS-Z, GA, AR, and ML conducted the data analysis. KS-Z conducted the statistical analysis. IR, KS-Z, and DK-B drafted the article. All authors contributed toward data analysis, drafting and critically revising the paper and agree to be accountable for all aspects of the work. All authors have read and approved the final version submitted for publication.
Disclosure
The authors report no conflicts of interest in this work.
References
Eckel RH, Grundy SM, Zimmet PZ. The metabolic syndrome. Lancet. 2005;365(9468):1415–1428. | ||
Fappa E, Yannakoulia M, Pitsavos C, Skoumas I, Valourdou S, Stefanadis C. Lifestyle intervention in the management of metabolic syndrome: could we improve adherence issues? Nutrition. 2008;24(3):286–291. | ||
Dina C, Meyre D, Gallina S, et al. Variation in FTO contributes to childhood obesity and severe adult obesity. Nat Genet. 2007;39(6): 724–726. | ||
Frayling TM, Timpson NJ, Weedon MN, et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science. 2007;316(5826):889–894. | ||
Olszewski PK, Fredriksson R, Olszewska AM, et al. Hypothalamic FTO is associated with the regulation of energy intake not feeding reward. BMC Neurosci. 2009;10:129. | ||
Hinney A, Nguyen TT, Scherag A, et al. Genome wide association (GWA) study for early onset extreme obesity supports the role of fat mass and obesity associated gene (FTO) variants. PLoS One. 2007;2(12):e1361. | ||
Scuteri A, Sanna S, Chen WM, et al. Genome-wide association scan shows genetic variants in the FTO gene are associated with obesity-related traits. PLoS Genet. 2007;3(7):e115. | ||
Sandholt CH, Hansen T, Pedersen O. Beyond the fourth wave of genome-wide obesity association studies. Nutr Diabetes. 2012;2:e37. | ||
Loos RJ, Yeo GS. The bigger picture of FTO: the first GWAS-identified obesity gene. Nat Rev Endocrinol. 2014;10(1):51–61. | ||
Tercjak-Rećko M, Luczyński W, Bernatowicz P, et al. Polymorphism rs9939609 of FTO gene is related to the body mass index in children from Podlaskie voievodship. Med Wieku Rozwoj. 2012;16(1):53–60. | ||
Luczynski W, Zalewski G, Bossowski A. The association of the FTO rs9939609 polymorphism with obesity and metabolic risk factors for cardiovascular diseases in Polish children. J Physiol Pharmacol. 2012;63(3):241–248. | ||
Dušátková L, Zamrazilová H, Sedláčková B, et al. Association of obesity susceptibility gene variants with metabolic syndrome and related traits in 1,443 Czech adolescents. Folia Biol (Praha). 2013;59(3):123–133. | ||
Povel CM, Boer JM, Reiling E, Feskens EJ. Genetic variants and the metabolic syndrome: a systematic review. Obes Rev. 2011;12(11): 952–967. | ||
Marcadenti A, Fuchs FD, Matte U, Sperb F, Moreira LB, Fuchs SC. Effects of FTO RS9939906 and MC4R RS17782313 on obesity, type 2 diabetes mellitus and blood pressure in patients with hypertension. Cardiovasc Diabetol. 2013;12:103. | ||
Carlos F, Silva-Nunes J, Flores O, et al. Association of FTO and PPARG polymorphisms with obesity in Portuguese women. Diabetes Metab Syndr Obes. 2013;6:241–245. | ||
Woehning A, Schultz JH, Roeder E, et al. The A-allele of the common FTO gene variant rs9939609 complicates weight maintenance in severe obese patients. Int J Obes (Lond). 2013;37(1):135–139. | ||
Mutch D, Clément K. Genetics of human obesity. Best Pract Res Clin Endocrinol Metab. 2006;20(4):647–664. | ||
Sikaris K. The clinical biochemistry of obesity. Clin Biochem Rev. 2004;25(3):165–181. | ||
Siljee JE, Unmehopa UA, Kalsbeek A, Swaab DF, Fliers E, Alkemade A. Melanocortin 4 receptor distribution in the human hypothalamus. Eur J Endocrinol. 2013;168(3):361–369. | ||
Męczekalski B, Czyżyk A, Warenik-Szymankiewicz A. The role of genes in pathogenesis of obesity. Contemporary view, pathogenesis, clinical aspects. Endocrinol Otyłość. 2008;5(1):27–37. | ||
Loos RJ, Lindgren CM, Li S, et al. Common variants near MC4R are associated with fat mass, weight and risk of obesity. Nat Genet. 2008;40(6):768–775. | ||
Povel CM, Boer JM, Onland-Moret NC, Dollé ME, Feskens EJ, Van der Schouw YT. Single nucleotide polymorphisms (SNPs) involved in insulin resistance, weight regulation, lipid metabolism and inflammation in relation to metabolic syndrome: an epidemiological study. Cardiovasc Diabetol. 2012;11:133. | ||
Kring SI, Holst C, Toubro S, et al. Common variants near MC4R in relation to body fat, body fat distribution, metabolic traits and energy expenditure. Int J Obes (Lond). 2010;34(1):182–189. | ||
Vogel CI, Boes T, Reinehr T, et al. Common variants near MC4R: exploring gender effects in overweight and obese children and adolescents participating in a lifestyle intervention. Obes Facts. 2011;4(1): 67–75. | ||
Winczyk K. The role of peroxisome proliferators-activated receptors (PPARγ) in neoplasms of endocrine glands. Endokrynol Pol. 2008;59(2):156–166. | ||
Braissant O, Foufelle F, Scotto C, Dauça M, Wahli W. Differential expression of peroxisome proliferator-activated receptors (PPARs): tissue distribution of PPAR-alpha, -beta, and -gamma in the adult rat. Endocrinology. 1996;137(1):354–366. | ||
Houseknecht KL, Cole BM, Steele PJ. Peroxisome proliferator-activated receptor gamma (PPARgamma) and its ligands: a review. Domest Anim Endocrinol. 2002;22(1):1–23. | ||
Desvergne B, Wahli W. Peroxisome proliferator-activated receptors: nuclear control of metabolism. Endocr Rev. 1999;20(5):649–688. | ||
Dytfeld J, Horst-Sikorska W. Peroxisome proliferator activated receptors γ (PPAR-γ) in human physiology and pathology. Prz Kardiodiabetol. 2009;4:187–191. | ||
Tellechea ML, Aranguren F, Pérez MS, Cerrone GE, Frechtel GD, Taverna MJ. Pro12Ala polymorphism of the peroxisome proliferatoractivated receptor-gamma gene is associated with metabolic syndrome and surrogate measures of insulin resistance in healthy men: interaction with smoking status. Circ J. 2009;73(11):2118–2124. | ||
Kruzliak P, Haley AP, Starcevic JN, Gaspar L, Petrovic D. Polymorphisms of the peroxisome proliferator-activated receptor-γ (rs1801282) and its coactivator-1 (rs8192673) are associated with obesity indexes in subjects with type 2 diabetes mellitus. Cardiovasc Diabetol. 2015;28(14):42. | ||
Miller M, Rhyne J, Chen H, et al. POC3 promoter polymorphisms C-482T and T-455C are associated with the metabolic syndrome. Arch Med Res. 2007;38(4):444–451. | ||
Moss PE, Lyles BE, Stewart LV. The PPARγ ligand ciglitazone regulates androgen receptor activation differently in androgen-dependent versus androgen-independent human prostate cancer cells. Exp Cell Res. 2010;316(20):3478–3488. | ||
Haring R, Völzke H, Felix SB, et al. Prediction of metabolic syndrome by low serum testosterone levels in men: results from the study of health in Pomerania. Diabetes. 2009;58(9):2027–2031. | ||
Brand JS, Van der Tweel I, Grobbee DE, Emmelot-Vonk MH, Van der Schouw YT. Testosterone, sex hormone-binding globulin and the metabolic syndrome: a systematic review and meta-analysis of observational studies. Int J Epidemiol. 2011;40(1):189–207. | ||
Corona G, Monami M, Rastrelli G, et al. Testosterone and metabolic syndrome: a meta-analysis study. J Sex Med. 2011;8(1):272–283. | ||
Herman WA, Krzoska A, Pawliczak E, Roszak M, Łacka K. Evaluation of relationships between plasma androgens level and clinical signs of hipoandrogenism according to age and comorbidity of metabolic syndrome in men. Pol Merkur Lekarski. 2014;36(211):16–21. | ||
Chubb SA, Hyde Z, Almeida OP, et al. Lower sex hormone-binding globulin is more strongly associated with metabolic syndrome than lower total testosterone in older men: the Health in Men Study. Eur J Endocrinol. 2008;158(6):785–792. | ||
Hong D, Kim YS, Son ES, et al. Total testosterone and sex hormone-binding globulin are associated with metabolic syndrome independent of age and body mass index in Korean men. Maturitas. 2013;74(2):148–153. | ||
Yang YH, Zhao MJ, Zhou SJ, et al. Is serum sex hormone-binding globulin a dominant risk factor for metabolic syndrome? Asian J Androl. 2015;17(6):991–995. | ||
Rotter I, Kosik-Bogacka D, Dołęgowska B, Skonieczna-Żydecka K, Pawlukowska W, Laszczyńska M. Analysis of relationships between the concentrations of total testosterone and dehydroepiandrosterone sulfate and the occurrence of selected metabolic disorders in aging men. Aging Male. 2015;18(2):249–255. | ||
Antonio L, Wu FC, O’neill TW, et al; EMAS Study Group. Associations between sex steroids and the development of metabolic syndrome: a longitudinal study in European men. J Clin Endocrinol Metab. 2015;100(4):1396–1404. | ||
Alberti KG, Zimmet P, Shaw J. The metabolic syndrome-a new worldwide definition. Lancet. 2005;366(9491):1059–1062. | ||
Shahid A, Rana S, Saeed S, Imran M, Afzal N, Mahmood S. Common variant of FTO gene, rs9939609, and obesity in Pakistani females. Biomed Res Int. 2013;2013:324093. | ||
Petry CJ, Lopez-Bermejo A, Díaz M, et al. Association between a common variant near MC4R and change in body mass index develops by two weeks of age. Horm Res Paediatr. 2010;73(4):275–280. | ||
Yen CJ, Beamer BA, Negri C, et al. Molecular scanning of the human peroxisome proliferator activated receptor gamma (hPPAR gamma) gene in diabetic Caucasians: identification of a Pro12Ala PPAR gamma 2 missense mutation. Biochem Biophys Res Commun. 1997;241(2):270–274. | ||
Al-Attar SA, Pollex RL, Ban MR, et al. Association between the FTO rs9939609 polymorphism and the metabolic syndrome in a non-Caucasian multi-ethnic sample. Cardiovasc Diabetol. 2008;7:5. | ||
Freathy RM, Timpson NJ, Lawlor DA, et al. Common variation in the FTO gene alters diabetes-related metabolic traits to the extent expected given its effect on BMI. Diabetes. 2008;57(5):1419–1426. | ||
Liguori R, Labruna G, Alfieri A, et al. The FTO gene polymorphism (rs9939609) is associated with metabolic syndrome in morbidly obese subjects from southern Italy. Mol Cell Probes. 2014;28(4):195–199. | ||
Jacobsson JA, Risérus U, Axelsson T, Lannfelt L, Schiöth HB, Fredriksson R. The common FTO variant rs9939609 is not associated with BMI in a longitudinal study on a cohort of Swedish men born 1920–1924. BMC Med Genet. 2009;10:131. | ||
Zavattari P, Loche A, Pilia S, et al. rs9939609 in the FTO gene is associated with obesity but not with several biochemical parameters in Sardinian obese children. Ann Hum Genet. 2011;75(6):648–654. | ||
Gustavsson J, Mehlig K, Leander K, et al. FTO genotype, physical activity, and coronary heart disease risk in Swedish men and women. Circ Cardiovasc Genet. 2014;7(2):171–177. | ||
Qi L, Kang K, Zhang C, et al. Fat mass-and obesity-associated (FTO) gene variant is associated with obesity: longitudinal analyses in two cohort studies and functional test. Diabetes. 2008;57(11):3145–3151. | ||
Qi L, Kraft P, Hunter DJ, Hu FB. The common obesity variant near MC4R gene is associated with higher intakes of total energy and dietary fat, weight change and diabetes risk in women. Hum Mol Genet. 2008;17(22):3502–3508. | ||
Xi B, Chandak GR, Shen Y, Wang Q, Shou D. Association between common polymorphism near the MC4R gene and obesity risk: a systematic review and meta analysis. PLoS One. 2012;7(9):e45731. | ||
Milewicz A, Tworowska-Bardzińska U, Dunajska K, Jędrzejuk D, Lwow F. Relationship of PPARgamma2 polymorphism with obesity and metabolic syndrome in postmenopausal Polish women. Exp Clin Endocrinol Diabetes. 2009;117(10):628–632. | ||
Hainerová IA, Zamrazilová H, Sedláčková D, Hainer V. Hypogonadotropic hypogonadism in a homozygous MC4R mutation carrier and the effect of sibutramine treatment on body weight and obesity-related health risks. Obes Facts. 2011;4(4):324–328. | ||
Park HK, Kim H, Kim HG, et al. Expression of peroxisome proliferator activated receptor gamma in prostatic adenocarcinoma. J Korean Med Sci. 2015;30(5):53. | ||
Kazue Y, Toshiro T. Effects of lifestyle modification on metabolic syndrome: a systematic review and meta-analysis. BMC Med. 2012; 10:138. | ||
Lakka TA, Laaksonen DE, Lakka HM, et al. Sedentary lifestyle, poor cardiorespiratory fitness, and the metabolic syndrome. Med Sci Sports Exerc. 2003;35(8):1279–1286. | ||
Liu YJ, Liu PY, Long J, et al. Linkage and association analyses of the UCP3 gene with obesity phenotypes in Caucasian families. Physiol Genomics. 2005;22(2):197–203. | ||
Rosmond R. Association studies of genetic polymorphisms in central obesity: a critical review. Int J Obes Relat Metab Disord. 2003;27(10):1141–1151. | ||
Faul F, Erdfelder E, Lang AG, Buchner A. G*Power 3 a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav Res Method. 2007;39(2):175–191. | ||
Cohen J. Statistical Power Analysis for the Behavioral Sciences. 2nd ed. Hillsdale, NJ: Lawrence Earlbaum Associates; 1988. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.