Back to Journals » International Journal of Nephrology and Renovascular Disease » Volume 13
Functional Relevancies of Trans-Differentiation in the Juxtaglomerular Apparatus of Rat Kidney
Authors Razga Z ![]()
Received 17 January 2020
Accepted for publication 9 May 2020
Published 12 June 2020 Volume 2020:13 Pages 147—156
DOI https://doi.org/10.2147/IJNRD.S246476
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Pravin Singhal
Zsolt Razga
Department of Pathology, University of Szeged, Szeged, Hungary
Correspondence: Zsolt Razga
Department of Pathology, University of Szeged, Állomás U. 2, Szeged H 6725, Hungary
Tel/ Fax +36 62 545162
Email [email protected]
Abstract: Glomerular filtration rate is controlled by the contractile effect of angiotensin II on afferent and efferent arterioles. The renin positivity of the afferent arterioles depends on tubuloglomerular feedback via the macula densa (MD) and short loop feedback via the afferent arteriolar endothelia. The renin-producing cells are trans-differentiated from smooth muscle cells (SMCs) of mainly the afferent arterioles, the MD cells are trans-differentiated from the neighboring tubular cells, and the high-permeability endothelial cells are trans-differentiated from normal permeability endothelial cells facing the renin-negative part of the afferent arterioles. All of the trans-differentiations depend on the activity of the renin-angiotensin system (RAS). The distribution of AT1 receptors for angiotensin II expresses the contractile effects of angiotensin II on renin-negative SMCs and the negative effect on trans-differentiation of renin-positive SMCs and MD cells. The purpose of this review is to summarize the stereological data of molecules like angiotensin II AT1 receptors, L-type calcium channels, and renin receptors in the juxtaglomerular apparatus of normal and STZ-induced diabetic rat kidneys, thus showing their functional relevancies on trans-differentiation among the juxtaglomerular apparatus’ elements.
Keywords: angiotensin II, renin, trans-differentiation, kidney, arteriole, afferent arteriole, efferent arteriole, tubuloglomerular feedback, macula densa
Introduction
The elements of the juxtaglomerular apparatus (JGA) are well defined, while their stereological behaviors are not well characterized. The JGA is a functional unit, but it has no defined length, size or volume.1
The glomerular fluid flow and filtration pressure result in a glomerular filtration rate, which is controlled by the tone of the afferent and efferent arterioles. The contraction of afferent arterioles decreases glomerular flow and filtration pressure, in opposition to the contraction of efferent arterioles where the glomerular flow is decreased, but the glomerular filtration pressure is increased.2 The main agonist of arteriolar contraction is angiotensin II via the angiotensin II AT1 receptors.3 Glomerular fluid flow and filtration pressure are under the control of the renin-angiotensin system (RAS).
The main sources of renin for glomerular fluid flow and filtration pressure are the renin-granulated parts of the afferent arterioles. Actual renin granulation in afferent arterioles depends on the balance of trans-differentiation between the renin-positive and renin-negative SMCs. This trans-differentiation is determined by the balance of osmotic pressure in the JGA, thus it is under the control of the RAS.4,5
The osmotic balance in the JGA is dependent on the fluid flow in this area. Tubuloglomerular feedback via the macula densa (MD) and short loop feedback via the afferent arteriolar endothelium are the two pathways that affect the fluid flow characteristics of the JGA.4 Tubuloglomerular feedback depends on a balance of trans-differentiation between the MD cells and the neighboring tubular cells.1 Short loop feedback depends on a balance of trans-differentiation between the high-permeability endothelial cells facing the renin-positive area of the afferent arterioles and the normal permeability endothelial cells of the afferent arterioles. Both of the trans-differentiations are under the control of the RAS.5
The RAS control of trans-differentiation between renin-positive and renin-negative SMCs is expressed by the negative effects of angiotensin II on renin granulation. Treatment with angiotensin II AT1 receptor antagonists increases renin granulation.4,6
In diabetes, the balance of trans-differentiation between renin-positive and renin-negative SMCs exhibits elevated renin granulation in afferent arterioles, while the kidney tissue level of angiotensin II is also elevated.7,8 This discrepancy is a particularity of diabetes, in which the number of angiotensin II AT1 receptor subtypes are estimated using stereological principles in afferent and efferent arterioles as renin-positive SMCs and renin-negative SMCs makes out it.9
The purpose of this review is to summarize the stereological data of some molecules involved in the JGA. Levels of angiotensin II AT1 receptors, L-type calcium channels, and renin receptors are related to the trans-differentiation of high-permeability and normal permeability endothelial cells of arterioles, or the trans-differentiation of renin-positive and renin-negative SMCs, or the trans-differentiation of macula densa and neighboring tubular cells in normal and STZ-induced diabetic rat kidneys. The stereological data is used to show the functional relevancies of some molecules found in the JGA.
Method
Measurement of permeability of endothelia was performed on full length of afferent arterioles. The stereological method was used at TEM level for mapping the amount of tracers along the wall of arterioles.10 The experimental diabetes was performed with streptozotocin using standard protocol.9 Measurement of angiotensin II level was performed in blood and kidney tissue by ELISA.9 The BrdU test and Ki67 immunohistochemistry were performed to detect the proliferation.1 The same labeling system was used for different antibodies (AT1 receptors; L-type Ca channel (pro)renin receptors). Where the distribution of immunohistochemical signal along the arterioles were estimated by using the stereological principles as described at 2006.11
Discussion
Trans-Differentiation Between the Afferent Arteriolar High- and Normal Permeability Endothelia
Permeability of Afferent Arteriole
The afferent arterioles, defined as pre-glomerular arterioles from the glomerulus to first branching, consist of two types of endothelia. It exhibits fenestration facing the extra-glomerular mesangial cells.4 The permeability of the afferent arteriolar endothelium increases with closeness to the glomerulus. The location of this endothelium corresponds to the renin-positive part of the afferent arteriole. After treatment with the angiotensin II antagonist, candesartan, the length of the high-permeability portion of the endothelium along the afferent arteriole increased. When estimating the length of the higher permeability portion of the endothelium and the renin-positive part of the afferent arteriole, a correlation was found between endothelial permeability and renin-positivity.5 The higher permeability endothelia facing the renin-positive part of the afferent arteriole are trans-differentiated from the normal endothelia located on the renin-negative part. The trans-differentiation of endothelia is under the control of the RAS.5
The endothelial permeability is related to the Ca2+ level. Higher Ca2+ levels in the endothelial cell result in higher permeability of micro vessels.12 In normal kidneys, the L-type Ca2+ channels are downregulated in the endothelial cells facing the renin-positive part of the afferent arterioles. These downregulations are not present in STZ-induced diabetic rat kidneys.13 This regulation of L-type Ca2+ channels in afferent arterioles of normal and diabetic rats shows the Ca2+ channel may well be related to trans-differentiation between the high and normal permeability endothelia of afferent arterioles.13
Trans-Differentiation Between Renin-Positive and Renin-Negative SMCs of Afferent Arterioles
The afferent arterioles have two parts. The renin-positive SMCs and actin-negative SMCs are found close to the glomerulus, beyond this part there are the renin-negative SMCs and actin-positive SMCs. The lengths of the two can vary over a lifetime.14
Distribution of Angiotensin II AT1 Receptors in Kidney Arterioles of Normal Rats
Estimation of the immunohistochemical signals of angiotensin II AT1 receptor on afferent and efferent arteriole profiles does not show any differences in SMCs and endothelial cells along the arterioles. Nonetheless, between the renin-positive SMCs and renin-negative SMCs significant differences were found, where angiotensin II AT1 receptors in renin-positive cells were downregulated compared to renin-negative SMCs.6 The same stereological estimation of angiotensin II AT1-A and AT1-B receptors in arterioles clarified that the trans-differentiation between the renin-positive SMCs and renin-negative SMCs is regulated by angiotensin II binding angiotensin II AT1-A receptors (Figure 1).15
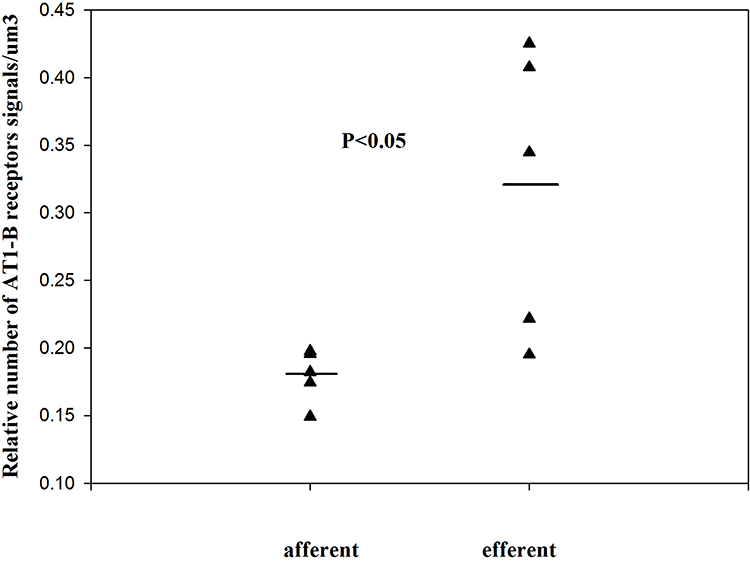 |
Figure 1 The relative number of angiotensin II AT1-B receptors on the SMCs of normal kidney arterioles (single values + mean, Mann–Whitney test). Reproduced with permission from Razga Z, Nyengaard JR. Up- and down-regulation of angiotensin II AT(1)-A and AT(1)-B receptors in afferent and efferent rat kidney arterioles. Journal of the Renin-Angiotensin-Aldosterone System 2008; 9: 196–201. Copyright © 2008, © SAGE Publications.15 The significant differences of AT1 receptors between the afferent end efferent arterioles are expressed the different contractile character of arterioles related with AT1-B receptors. |
Between the afferent and efferent arterioles a 27.5% difference in angiotensin II AT1 receptors was found, but it is not significant.6 When estimating the angiotensin II AT1 receptor subtypes separately, significant differences were found in the relative number of angiotensin II AT1-B receptors between the afferent and efferent arterioles (Figure 2).15 The differences in the contractile effects of angiotensin II between the afferent and efferent arterioles are expressed in the number of angiotensin II AT1-B receptors.
 |
Figure 2 The relative number of angiotensin II AT1-A receptors on renin-positive and renin-negative SMCs of normal afferent arterioles (single values + mean, Mann–Whitney test). Reproduced with permission from Razga Z, Nyengaard JR. Up- and down-regulation of angiotensin II AT(1)-A and AT(1)-B receptors in afferent and efferent rat kidney arterioles. Journal of the Renin-Angiotensin-Aldosterone System 2008; 9: 196–201. Copyright © 2008, © SAGE Publications.15 This significant differences of AT1 receptors show the trans-differentiation between the renin-positive and renin-negative SMCs related to angiotensin II AT1-A receptors subtype. |
Distribution of Angiotensin II AT1 Receptors on Kidney Arterioles in STZ-Induced Diabetic Rats
Insulin upregulates AT1 receptor gene expression in cell cultures of vascular SMCs. Higher levels of AT1 receptor result in enhanced responses of the vascular SMCs to angiotensin II.16,17 In STZ-induced diabetes, AT1-A and AT1-B are downregulated in the afferent and efferent arterioles. These downregulations are extreme and heterogeneous.9 In this experiment, diabetes is shown with elevated blood sugar levels at ten weeks, with normal angiotensin II tissue levels in the kidney. These parameters are evidence that angiotensin II receptor downregulation is independent from angiotensin II levels.9
In diabetic rats, the angiotensin II level in the kidney is elevated compared to normal rats,7,18,19 however the level of renin granulation in the afferent arterioles is also higher than normal.8 In diabetic rats, angiotensin II AT1-A and AT1-B receptors are downregulated on both the renin-positive and renin-negative SMCs surfaces (Figure 3A and B).9 The difference in the number of AT1-A receptors between the renin-positive and renin-negative SMCs is same as in normal rats, the downregulation of AT1-B are enhanced in the renin-positive than in renin-negative SMCs (Figure 3A and B). The number of AT1 (AT1-A + AT1-B) receptors is lower in diabetic rats. A possible reason for high-level trans-differentiation of renin-positive SMCs from renin-negative SMCs with high levels of angiotensin II might be the attached downregulation of the AT1-B receptors of SMCs.9
 |
Figure 3 In STZ-induced diabetes, the angiotensin II AT1 receptors are downregulated in both renin-positive and renin-negative SMCs of kidney arterioles. Adapted from Razga Z, Kovacs G, Bodi N, Talapka P, Nyengaard JR. Heterogeneous downregulation of angiotensin II AT1-A and AT1-B receptors in arterioles in STZ-induced diabetic rat kidneys. Biomed Res Int. 2014;2014:1–6. the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/).9 In diabetes the main morphological changes the high renin granulation with the high angiotensin II level in kidney tissue. This apparent discrepancy was resolved by finding the downregulation of angiotensin II AT1-A receptors (A). The downregulation of angiotensin II AT1-B receptors showed the contractile behavior of both arterioles were changed to abnormal in diabetes (B). |
Distribution of (Pro)renin Receptors in Arteriolar SMCs of Normal Rats
The (pro)renin receptors are localized on cell surfaces and in parts of the cytoplasm. In the kidney, the (pro)renin receptors are shown by immunohistochemistry in glomerular mesangial and epithelial cells, vascular SMCs of arteries, and in distal and collecting tubular cells.20–22 In neonates, the (pro)renin receptors have enhanced expression, supporting the theory that the (pro)renin receptors play a role in kidney development.23 Stereological estimation of (pro)renin receptors suggested that the relative number (pro)renin receptors were upregulated on the surface of renin-positive SMCs compared to renin-negative SMCs and downregulated in the cytoplasm of renin-positive SMCs compared to renin-negative SMCs (Figures 4 and 5).24 The distribution of (pro)renin receptors is related to trans-differentiation between renin-positive SMCs and renin-negative SMCs of the afferent arterioles.24
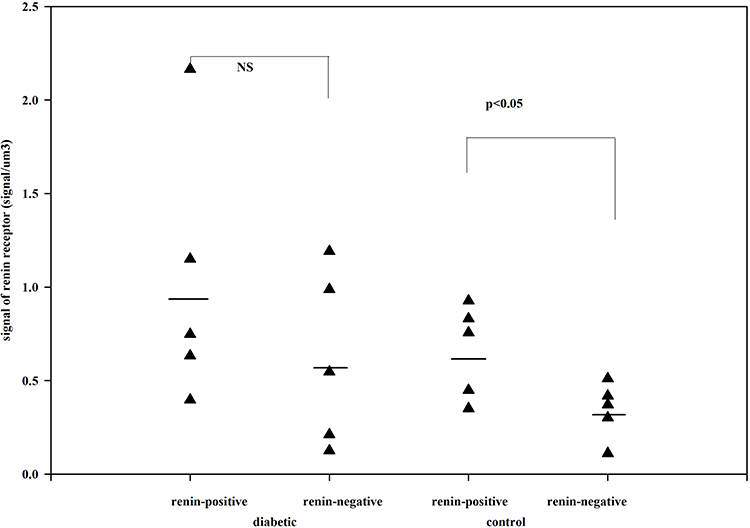 |
Figure 4 The number of renin receptors on the surface of afferent arteriolar SMCs. Adapted from Razga Z, Kovacs G, Bódi N, Talapka P, Bagyánszki M. Regulation of (Pro)Renin receptor in renin-positive smooth muscle cells of kidney arterioles in rats with STZ-induced diabetes. Int J Nephrol. 2019;2019:6. Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/).24 Significant difference was found in relative number of renin receptors between the renin-negative and renin-positive SMCs in normal, but not in diabetes. This difference expressed the renin receptor related to renin granulation that is the trans-differentiation between the renin-positive and renin-negative SMCs in normal. But in diabetes this difference is not presented which was showed the pathological state was processed from normal. |
 |
Figure 5 The number of renin receptors in the cytoplasm of afferent arteriolar SMCs. Adapted from Razga Z, Kovacs G, Bódi N, Talapka P, Bagyánszki M. Regulation of (Pro)Renin receptor in renin-positive smooth muscle cells of kidney arterioles in rats with STZ-induced diabetes. Int J Nephrol. 2019;2019:6. Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/).24 The same distribution is presented as in surface of SMCs. The renin receptors in cytoplasm of SMCs are related to trans-differentiation of renin-granulated cells from SMCs. |
Distribution of (Pro)renin Receptors in Arteriolar SMCs of STZ-Induced Diabetic Rats
The relative number of (pro)renin receptors in kidney arterioles was estimated in STZ-induced diabetic rats, where the angiotensin II kidney level does not show any changes compared to normal. No pathomorphological changes were found in the glomerulus nor in increments of renin granulation of arterioles. However (pro)renin receptors were distributed differently in afferent arterioles compared to normal. The redistribution of (pro)renin receptors resulted in the significant differences between the renin-positive SMCs and renin-negative SMCs were equalized in diabetic kidneys. The redistributions were not related to angiotensin II plasma or kidney levels.24
Distribution of L-Type Calcium Channels in Arteriolar SMCs of Normal and STZ-Induced Diabetic Rats
Calcium channel blockers, such as nifedipine, diltiazem, and nitrendipine, cause vasodilatation mainly on the afferent arterioles. This effect on the arteriolar SMCs includes significant increases in glomerular filtration rate and renal blood flow.25 The Ca2+ level of the intracellular and extracellular spaces of afferent arterioles are affiliated to the release of renin.26 Renin secretion is inhibited by T-type Ca2+ channel blockers, whereas L-type blockers have the opposite effect.27 In the normal kidney, estimation of relative numbers of L-type Ca2+ channels does not show any significant difference in arteriolar renin-positive and renin-negative SMCs.13 This result does not support any relationship between the trans-differentiation of renin-positive and renin-negative SMCs and L-type Ca2+ channels in normal kidneys. But in diabetic kidneys, the relative number of L-type Ca2+ channels are upregulated in renin-positive SMCs, causing a significant difference between the renin-positive and renin-negative SMCs.13 The upregulation of L-type Ca2+ channels in renin-positive SMCs in diabetic kidneys could be related to a higher state of renin granulation in arterioles of diabetic kidneys as a result of the trans-differentiation between the renin-positive SMCs and renin-negative SMCs.13
Trans-Differentiation of Macula Densa from Neighboring Tubular Cells
The MD is modified epithelial cells of the distal tubule facing the vascular pole of glomerulus.28 The morphology of MD cells differs in several ways from neighboring tubular cells. They have a prismatic shape, poorly-developed lateral cytoplasm and protrude into the tubular lumen with palisade nuclei.28 There is significant correlation between the basal area of the MD and the contact area between the Goormagtigh field and the MD.29,30 Considerable enlargement of the MD was found in Bartter’s syndrome. The different morphology of MD from neighboring tubular cells signals the different, specialized function, where the size of MD may be changed.31
Total Volume of Macula Densa
By estimating the volume of macula densa in normal kidneys and after the treatment with angiotensin II AT1 receptor antagonist candesartan, it was revealed that the volume of MD increased significantly.1 The volume of MD cells did not change,1 rather the increase in MD volume was caused by an increase in MD cell number. The proliferation of MD cells was excluded from the immunohistochemistry by using bromodeoxyuridine (BrdU) and Ki67. The increase of MD volume was caused by trans-differentiation of MD cells from neighboring tubular cells. This trans-differentiation was inhibited by angiotensin II via the AT1 receptors (Figures 6–8).1
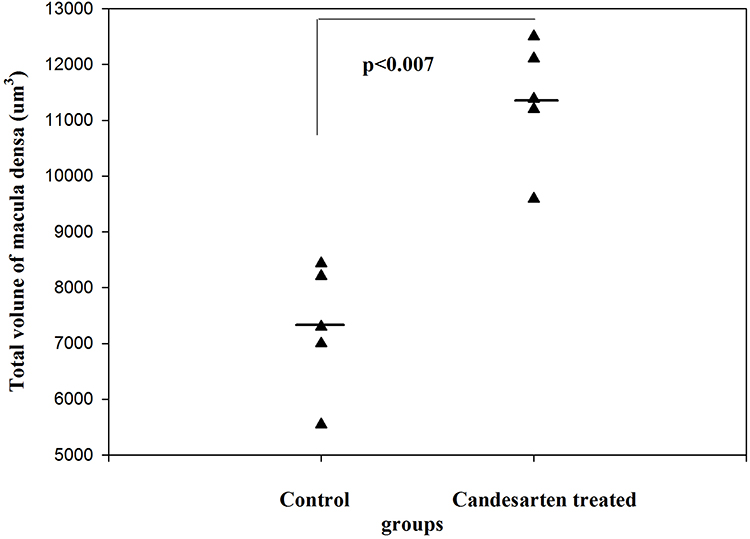 |
Figure 6 The total volume of MD in control and candesartan-treated groups. After treatment with candesartan the volume of MD increased (Paired T-test; p<0.007) by increasing the number of MD cells. Reproduced with permission from Razga Z, Nyengaard JR. The effect of angiotensin II on the number of macula densa cells through the AT1 receptor. Nephron Physiology 2009; 112: 37–43. Copyright © 2009, © 2009 S. Karger AG, Basel.1 The total volume of MD is could be changed in the life related to the activity of RAS. |
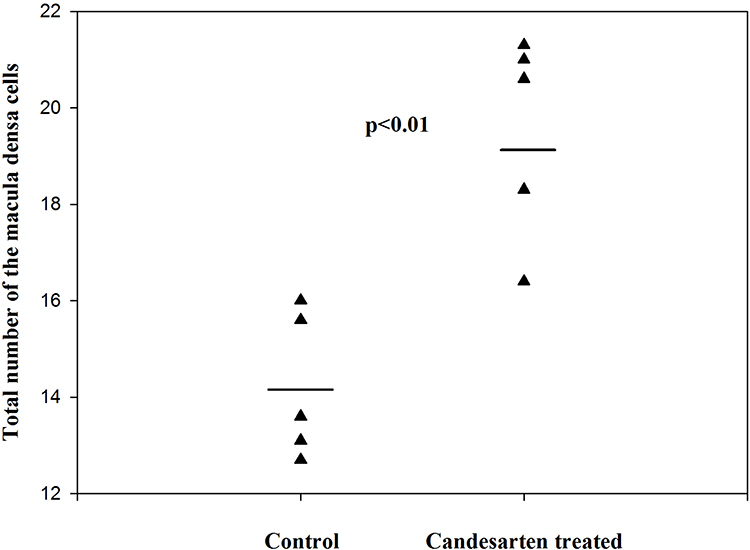 |
Figure 7 The total number of MD cells in control and candesartan-treated groups. There is a significant increase in the candesartan-treated group vs the control (Paired T-test; p<0.011). Reproduced with permission from Razga Z, Nyengaard JR. The effect of angiotensin II on the number of macula densa cells through the AT1 receptor. Nephron Physiology. 2009; 112: 37–43. Copyright © 2009, © 2009 S. Karger AG, Basel 0.1 The changes of total volume of MD are a changes in number of MD cells related to activity of RAS. The proliferation of MD cells was excluded by immunohistochemistry of Ki67 and BrdU test,1 so the changes in number of MD cells are the trans-differentiation related process. |
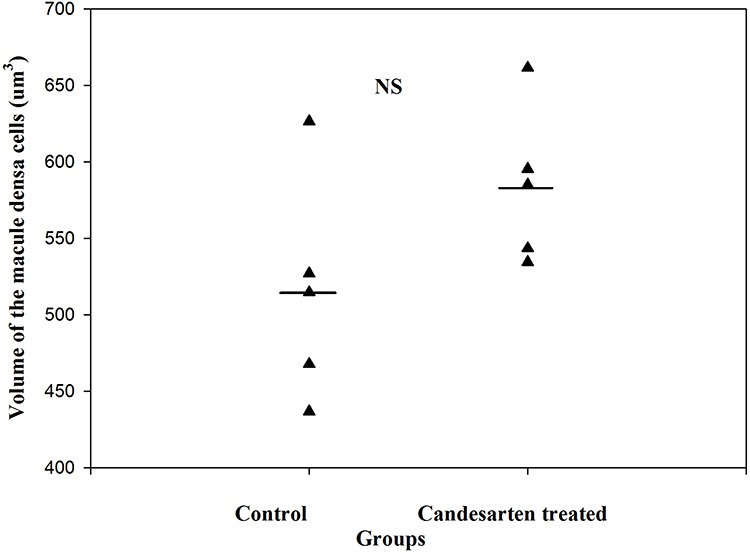 |
Figure 8 The volume of MD cells in the control and candesartan-treated groups. There are no significant differences between the two group. Reproduced with permission from Razga Z, Nyengaard JR. The effect of angiotensin II on the number of macula densa cells through the AT1 receptor. Nephron Physiology. 2009; 112: 37–43. Copyright © 2009, © 2009 S. Karger AG, Basel.1 The size of MD cells were not changed, while the volume of MD increased by treatment of candesartan. This is evidence for increment of MD caused by increment of number of MD cells related to activity of RAS. |
Distribution of Angiotensin II AT1receptros on Macula Densa
The MD releases some molecules, like cyclooxygenase-2 (COX2), prostaglandin-E2 (PgE2), Ca2+, and neuronal nitric oxide (nNOS), which are involved in tubuloglomerular feedback. There are interspecies differences in the COX2 positivity of MD. In humans, COX2 is present in SMCs of afferent arterioles instead of MD.32 In rats, COX2 appears in the MD.33,34 Since angiotensin II inhibits COX2 positivity via the AT1 receptor,35 the activity of angiotensin II should be reduced in MD.1 If MD cells trans-differentiated from neighboring COX2 negative tubular cells,28 according to our hypothesis there should be a significant difference in the number of angiotensin II AT1 receptors. This theory was supported by the results of our estimation of the number of angiotensin II AT1 receptors on surface of macula densa and normal tubular cells facing the macula densa. Where it was found that the number of AT1 receptors was significantly reduced on MD cell surfaces compared to neighboring normal tubular cells (Table 1). This state of AT1 receptor distribution is similar to renin-positive SMCs and renin-negative SMCs of afferent arterioles. Both the trans-differentiation of MD cells and renin-granulated cells are under RAS control.1
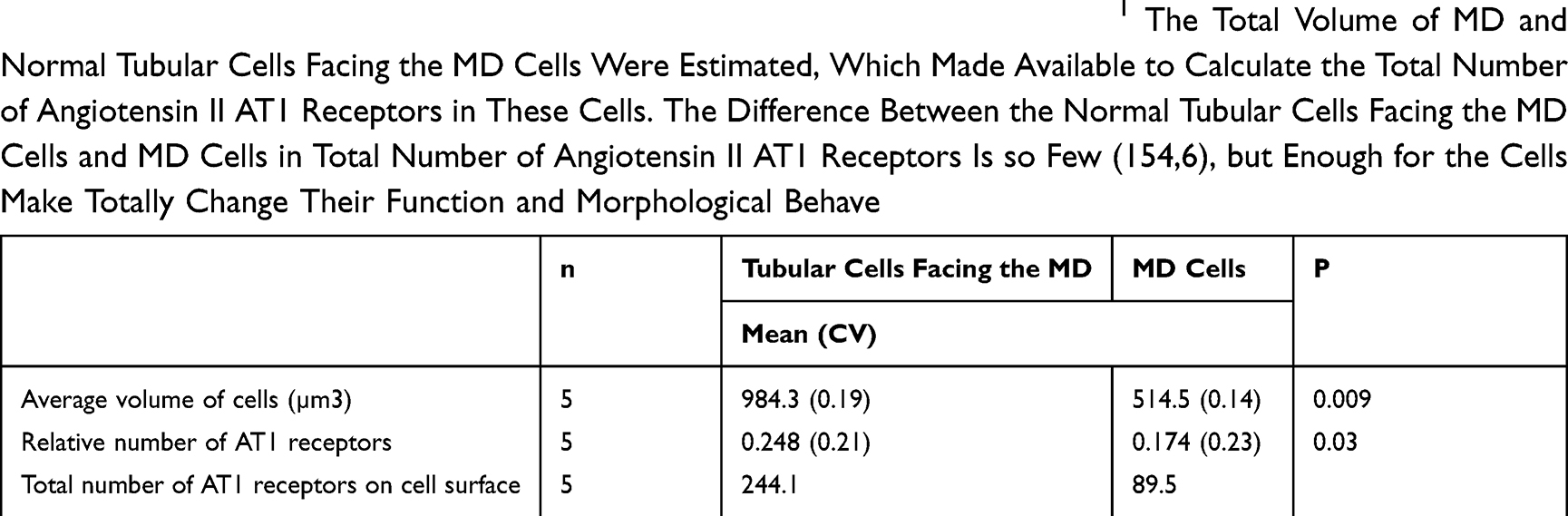 |
Table 1 The Number of Angiotensin II AT1 Receptors on the Macula Densa.1 The Total Volume of MD and Normal Tubular Cells Facing the MD Cells Were Estimated, Which Made Available to Calculate the Total Number of Angiotensin II AT1 Receptors in These Cells. The Difference Between the Normal Tubular Cells Facing the MD Cells and MD Cells in Total Number of Angiotensin II AT1 Receptors Is so Few (154,6), but Enough for the Cells Make Totally Change Their Function and Morphological Behave |
Angiotensin II, TGFβ and Trans-Differentiation
Angiotensin II attenuates the TGFβ induced trans-differentiation via the angiotensin II AT1 receptors.36,37 Any physiological or pathophysiological pathway makes the changes in the salt and water balance like using the mineral corticoids, diuretics and water deplete, etc., induce the trans-differentiation indirect manner by the changing the activity of RAS. The pharmacological effect on activity of RAS with angiotensin II AT1 receptor blocker, ACE blocker or renin antagonist induce the trans-differentiation on juxtaglomerular apparatus indirect manner too. The TGFβ pathway attenuated by angiotensin II through the AT1 receptors are the primary mechanism of trans-differentiation in juxtaglomerular apparatus. Any physiological or pharmacological effect on activity of RAS could generate the trans-differentiation of element of juxtaglomerular apparatus indirectly.
Conclusion
The trans-differentiation of endothelial cells, renin-granulated cells and MD cells are an important mechanism in the functionality of the JGA in normal and in experimental diabetes. The three molecules, angiotensin II AT1 (AT1-A and At1-B) receptors, L-type calcium channels and (pro) renin receptors play important roles in the trans-differentiation of the different cells types of the JGA element in the tubules or arterioles of the kidney. The trans-differentiation of all the cell types are under RAS control, but independent of the angiotensin II plasma or kidney levels in normal and diabetic rats.
Disclosure
The author reports no conflicts of interest in this work.
References
1. Razga Z, Nyengaard JR. The effect of angiotensin II on the number of macula densa cells through the AT1 receptor. Nephron Physiol. 2009;112(2):37–43. doi:10.1159/000213507
2. Ren Y, Garvin JL, Carretero OA. Efferent arteriole tubuloglomerular feedback in the renal nephron. Kidney Int. 2001;59(1):222–229. doi:10.1046/j.1523-1755.2001.00482.x
3. Arendshorst WJ, Brannstrom K, Ruan X. Actions of angiotensin II on the renal microvasculature. J Am Soc Nephrol. 1999;10(Suppl 11):149–161.
4. Rosivall L, Razga Z, Ormos J. Morphological characterization of human juxtaglomerular apparatus. Kidney Int Suppl. 1991;32:9–12.
5. Rosivall L, Razga Z, Mirzahosseini S, Tornoci L. Endothelial permeability of the afferent arteriole and its changes as the result of alteration in the activity of the renin-angiotensin system. J Am Soc Nephrol. 1999;10(Suppl 11):172–177.
6. Razga Z, Nyengaard JR. Estimation of the number of angiotensin II AT1 receptors in rat kidney afferent and efferent arterioles. Anal Quant Cytol Histol. 2007;29(4):208–216.
7. Ichihara A, Hayashi M, Kaneshiro Y, et al. Inhibition of diabetic nephropathy by a decoy peptide corresponding to the “handle” region for nonproteolytic activation of prorenin. J Clin Invest. 2004;114(8):1128–1135. doi:10.1172/JCI21398
8. Ribeiro-Oliveira A
9. Razga Z, Kovacs G, Bodi N, Talapka P, Nyengaard JR. Heterogeneous downregulation of angiotensin II AT1-A and AT1-B receptors in arterioles in STZ-induced diabetic rat kidneys. Biomed Res Int. 2014;2014:1–6. doi:10.1155/2014/947506
10. Osterby R, Bangstad HJ, Nyberg G, Walker JD, Viberti G. A quantitative ultrastructural study of juxtaglomerular arterioles in IDDM patients with micro- and normoalbuminuria. Diabetologia. 1995;38(11):1320–1327. doi:10.1007/BF00401765
11. Razga Z, Nyengaard JR. A stereologic approach to estimate the number of immunogold-labeled molecules in cells of tubules. Anal Quant Cytol Histol. 2006;28(1):54–60.
12. Nilius B, Droogmans G. Ion channels and their functional role in vascular endothelium. Physiol Rev. 2001;81(4):1415–1459. doi:10.1152/physrev.2001.81.4.1415
13. Razga Z, Kovacs G, Bodi N, Talapka P. Upregulation of the L-type calcium channel in renin-positive smooth muscle cells of arterioles in the kidneys of rats with streptozotocin-induced diabetes. Anal Quant Cytol Histol. 2015;37:214–220.
14. Hackenthal E, Metz R, Bührle CP, Taugner R. Intrarenal and intracellular distribution of renin and angiotensin. Kidney Int. 1987;31:S4–S17.
15. Razga Z, Nyengaard JR. Up- and down-regulation of angiotensin II AT1A and AT1B receptors in afferent and efferent rat kidney arterioles. J Renin Angiotensin Aldosterone Syst. 2008;9(4):196–201. doi:10.1177/1470320308098592
16. Nickenig G, Roling J, Strehlow K, Schnabel P, Bohm M. Insulin induces upregulation of vascular AT1 receptor gene expression by posttranscriptional mechanisms. Circulation. 1998;98(22):2453–2460. doi:10.1161/01.CIR.98.22.2453
17. Nickenig G, Bohm M. Vascular AT(1) receptors in hypertension and metabolic syndrome. Herz Kreislauf. 1998;30:329–334.
18. Kang JJ, Toma I, Sipos A, Meer EJ, Vargas SL, Peti-Peterdi J. The collecting duct is the major source of prorenin in diabetes. Hypertension. 2008;51(6):1597–1604. doi:10.1161/HYPERTENSIONAHA.107.107268
19. Peti-Peterdi J, Kang JJ, Toma I. Activation of the renal renin-angiotensin system in diabetes–new concepts. Nephrol Dial Transplant. 2008;23(10):3047–3049. doi:10.1093/ndt/gfn377
20. Nguyen G, Contrepas A. Physiology and pharmacology of the (pro)renin receptor. Curr Opin Pharmacol. 2008;8(2):127–132. doi:10.1016/j.coph.2007.12.009
21. Nguyen G, Muller DN. The biology of the (pro)renin receptor. J Am Soc Nephrol. 2010;21(1):18–23. doi:10.1681/ASN.2009030300
22. Nguyen G. Renin, (pro)renin and receptor: an update. Clin Sci (Lond). 2011;120(5):169–178. doi:10.1042/CS20100432
23. Terada T, Urushihara M, Saijo T, Nakagawa R, Kagami S. (Pro)renin and (pro)renin receptor expression during kidney development in neonates. Eur J Pediatr. 2017;176(2):183–189. doi:10.1007/s00431-016-2820-9
24. Razga Z, Kovacs G, Bódi N, Talapka P, Bagyánszki M. Regulation of (Pro)Renin receptor in renin-positive smooth muscle cells of kidney arterioles in rats with STZ-induced diabetes. Int J Nephrol. 2019;2019:6. doi:10.1155/2019/6971928
25. Hayashi K, Wakino S, Sugano N, Ozawa Y, Homma K, Saruta T. Ca 2+ channel subtypes and pharmacology in the kidney. Circ Res. 2007;100(3):342–353. doi:10.1161/01.RES.0000256155.31133.49
26. Beierwaltes WH. The role of calcium in the regulation of renin secretion. Am J Physiol Renal Physiol. 2010;298(1):F1–F11. doi:10.1152/ajprenal.00143.2009
27. Wagner C, Kramer BK, Hinder M, Kieninger M, Kurtz A. T-type and L-type calcium channel blockers exert opposite effects on renin secretion and renin gene expression in conscious rats. Br J Pharmacol. 1998;124(3):579–585. doi:10.1038/sj.bjp.0701861
28. Taugner R, HACKENTHAL E, Densa M. The Juxtaglomerular Apparatus. Berlin: Springer-Verlag; 1989:25–43.
29. Christensen JA, Meyer DS, Bohle A. Structure of human juxtaglomerular apparatus - morphometric, light-microscopic study on serial sections. Virchows Arch a Pathol Anat Histopathol. 1975;367(2):83–92. doi:10.1007/BF00430947
30. Christensen JA, Bohle A. Juxtaglomerular apparatus in normal rat-kidney. Virchows Arch a Pathol Anat Histopathol. 1978;379(2):143–150. doi:10.1007/BF00432483
31. Christensen JA, Bader H, Bohle A, Meyer DS. Structure of juxtaglomerular apparatus in Addison's-disease, Bartter’s-syndrome, and in Conn’s syndrome - comparative, morphometric, light microscopic study on serial sections. Virchows Arch a Pathol Anat Histopathol. 1976;370(2):103–112. doi:10.1007/BF00430807
32. Komhoff M, Grone HJ, Klein T, Seyberth HW, Nusing RM. Localization of cyclooxygenase-1 and -2 in adult and fetal human kidney: implication for renal function. Am J Physiol. 1997;272:460–468.
33. Castrop H, Schweda F, Schumacher K, Wolf K, Kurtz A. Role of renocortical cyclooxygenase-2 for renal vascular resistance and macula densa control of renin secretion. J Am Soc Nephrol. 2001;12(5):867–874.
34. Castrop H, Klar J, Wagner C, Hocherl K, Kurtz A. General inhibition of renocortical cyclooxygenase-2 expression by the renin-angiotensin system. Am J Physiol Renal Physiol. 2003;284(3):F518–F524. doi:10.1152/ajprenal.00338.2002
35. Harris RC, Zhang MZ, Cheng HF. Cyclooxygenase-2 and the renal renin-angiotensin system. Acta Physiol Scand. 2004;181(4):543–547. doi:10.1111/j.1365-201X.2004.01329.x
36. Ehanire T, Ren L, Bond J, et al. Angiotensin II stimulates canonical TGF-+- signaling pathway through angiotensin type 1 receptor to induce granulation tissue contraction. J Mol Med. 2015;93:289–302.
37. Piera-Velazquez S, Jimenez SA. Endothelial to mesenchymal transition: role in physiology and in the pathogenesis of human diseases. Physiol Rev. 2019;99:1281–1324.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.