")
Back to Journals » Infection and Drug Resistance » Volume 17
Antimicrobial Resistance and Molecular Characterization of Salmonella Rissen Isolated in China During 2008–2019
Authors Wang L, Nie L, Liu Y, Hu L, Zhou A, Wang D, Xu X, Guo J
Received 6 December 2023
Accepted for publication 4 March 2024
Published 12 March 2024 Volume 2024:17 Pages 969—976
DOI https://doi.org/10.2147/IDR.S453611
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Lili Wang,1,2,* Lu Nie,3,* Yue Liu,4,* Liang Hu,2 Aiping Zhou,2 Dongjiang Wang,2 Xuebin Xu,4 Jian Guo2
1Department of Laboratory Medicine, Zhoushan Women and Children Hospital, Zhoushan, People’s Republic of China; 2Department of Laboratory Medicine, Shanghai East Hospital, School of Life Sciences and Technology, Tongji University, Shanghai, People’s Republic of China; 3Department of Laboratory Medicine, The First People’s Hospital of Foshan, Foshan, Guangdong, People’s Republic of China; 4Shanghai Municipal Center for Disease Control and Prevention, Shanghai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jian Guo; Xuebin Xu, Email [email protected]; [email protected]
Background: This study aimed to provide epidemiological features of Salmonella enterica serovar Rissen, determine antimicrobial susceptibility, virulence gene profiles, and describe the potential association of S. Rissen from different sources in China.
Methods: During 2008– 2019, a total of non-repetitive 228 S. Rissen isolates were collected from human, animals and environment in China. The antimicrobial susceptibility test, screening of antimicrobial and virulence genes by PCR, and pulsed-field gel electrophoresis (PFGE) were performed.
Results: Among the 154 isolates from human, the majority of the cases (80.5%) occurred in summer, and S. Rissen was mainly detected in people aged 21– 40 (37.7%) and 41– 60 (28.6%) years old, and 74 non-human source S. Rissen strains were identified, with pork being the most common source. About 93.4% isolates were resistant to at least one of the 12 tested antimicrobial agents, and high frequencies of resistance were observed for tetracyclines (91.2%), trimethoprim-sulfamethoxazole (74.1%) and ampicillin (67.5%). A total of 171 (75%) isolates were resistant to at least three categories of antimicrobials, and the most common resistance profile was Tetracycline(s)-β-Lactams-Sulfonamides. The resistance rates to chloramphenicol, quinolones and sulfafurazole were significantly higher in strains isolated from human compared to non-human source strains. Among these isolates, the β-Lactams resistance was mainly associated with gene blaTEM (54.7%), sulfonamide resistance with sul2 (45.7%) and sul3 (54.3%), tetracycline resistance with tetA (81.3%). All the isolates harbored virulence genes hilA, sopB, sciN, stn and ssrB, and most of them harbored ssaQ (98.7%), mgtC (98.7%) and invA (98.2%). The majority (91.7%) of S. Rissen isolates showed high similarity (> 80%) with each other in PFGE patterns and came from human, animals and environment.
Conclusion: The high frequencies of multidrug resistance and probable clonal dissemination in this serovar call for the necessity of systematic surveillance on S. Rissen in China.
Keywords: Salmonella Rissen, antimicrobial susceptibility test, resistance genes, virulence genes, pulsed-field gel electrophoresis
Introduction
Salmonella, an important foodborne pathogen causing gastroenteritis in the human population, is responsible for about 93.8 million cases and at least 155,000 deaths worldwide each year.1 Over 2600 serovars of Salmonella have been reported, among which several serovars can cause human infections.2 Salmonella enterica serovar Rissen (S. Rissen) is considered one of the most common serovars in pork production systems and can be transmitted to humans through food or water contamination.3,4 Apart from pork, S. Rissen was also isolated from other food associated poultry and seafood.5,6 S. Rissen has been known to cause outbreaks and sporadic cases of human infections.7,8
The emergence and spread of antimicrobial resistance (AMR) in salmonella pose a serious threat to human and animal health. The profiles of antimicrobial resistance and virulence genes played a crucial role in Salmonella surveillance and clinical therapy. The antimicrobial susceptibilities of S. Rissen were diverse in different geographic locations. In Thailand, over 90% of isolates were resistant to ampicillin, sulfisoxazole, tetracycline and streptomycin, while in the USA, about half of the S. Rissen isolates were pan-sensitive.9 In a survey concerning the multidrug resistance patterns of emerging S. Rissen along the Food Chain in China, most of the isolates were resistant to tetracycline, trimethoprim-sulfamethoxazole, chloramphenicol, streptomycin, sulfisoxazole, and ampicillin.10 Limited data on the virulence profiles of S. Rissen are available around the world. Pulsed-field gel electrophoresis (PFGE) typing, was considered to be a “gold standard” tool to investigate outbreaks and perform epidemiological surveys and has already been used to discover the common origin of S. Rissen isolates across a long geographic distance.11
Herein, the objective of this study was to provide epidemiological features of the infections caused by S. Rissen, to determine antimicrobial susceptibility, virulence gene profiles, and to disclose the potential relatedness of S. Rissen isolated from humans, food associated with poultry and animals, and environment in China.
Materials and Methods
Collection of S. rissen Strains
A total of 228 non-repetitive strains of S. Rissen were isolated from six provinces (Guangxi, Guangdong, Fujian, Hubei, Hebei and Shanxi) and one municipality (Shanghai) in China during 2008–2019. The 154 isolates were obtained from anal swabs or feces of human aged from 6 months to 81 years, and the 74 isolates were from foods (animals) and river/lake. The isolation of Salmonella was performed following the Standard ISO-6579 and the procedure recommended by the World Health Organization.12,13 The suspected Salmonella isolates were confirmed using API identification kits (bioMe´rieux, Marcy l’Etoile, France). Serotyping of the S. Rissen was carried out according to the White–Kauffmann–Le Minor scheme by slide agglutination with O and H antigen specific sera (Bio-Rad, Marnes-La Coquette, France; Staten Serum Institute, Copenhagen, Denmark; Sifin, Berlin, Germany) by the Shanghai Municipal Center for Disease Control (Shanghai, China).14
Antimicrobial Susceptibility Testing
Fourteen antimicrobials were used, including gentamicin (GEN), streptomycin (STR), ampicillin (AMP), amoxicillin-clavulanic acid (AMC), cefoxitin (FOX), ceftiofur (EFT), ceftriaxone (CRO), azithromycin (AZM), chloramphenicol (CHL), nalidixic acid (NAL), ciprofloxacin (CIP), sulfafurazole (SIZ), trimethoprim-sulfamethoxazole (SXT) and tetracycline (TET). MICs were determined by broth microdilution using dehydrated panels CMV3AGNF (Thermo Fisher Scientific, Waltham, MA) following the manufacturer’s instructions. Resistance was defined using CLSI 2023 criteria, except for STR and EFT, for which there are no clinical breakpoints. The reference strain, E. coli ATCC 25922 was used as a quality control strain for determining the MIC of the antimicrobial agents.
Detection of Resistance-Associated Genes and Virulence Genes
β-lactamase associated genes (blaTEM, blaOXA, and blaSHV), sulfonamides-resistance associated genes (sul1, sul2, sul3, and dfr), tetracycline resistance-related genes (tetA and tetB), aminoglycosides-resistance associated genes (strA, strB, and aadA), chloramphenicol-resistance associated genes (cat), class 1 integron family (intl1, intl1 VR(cx) and qacED1), and 12 virulence genes (invA, hilA, ssaQ, ssrB, mgtC, spi4D, sopB, sciN, safB, viaB, stn and pltA) were screened by PCR using suitable reported primers.15 PCR products were sequenced using a DNA analyzer (Applied Biosystem, Foster City, CA, USA). The obtained sequences were analyzed by comparison with the National Centre for Biotechnology Information (NCBI) database sequences using the BLAST program. Each run included appropriate positive and negative controls.
Pulsed-Field Gel Electrophoresis (PFGE)
The genomic DNA of Salmonella Rissen isolates and standard marker Salmonella enterica serovar Braenderup strain H9812 were embedded in 1% SeaKem Gold agarose and digested with 50 U of restriction enzyme XbaI (TaKaRa, Dalian, China) for 2 h. The samples of digested genomic DNA were run on CHEEF Mapper system (Bio-Rad, Hercules, CA) from 2.16 to 63.8 seconds for 19 h at 14°C. PFGE images were analyzed with BioNumerics software package 6.5 (Applied Maths, Kortrijk, Belgium) using the unweighted pair group method and an arithmetic average (UPGMA) clustering algorithm.
Results
Epidemiology of S. rissen in Patients
A total of 228 strains of S. Rissen were collected, with the largest number of strains collected in East China (Shanghai, n = 122; Fujian, n = 3), followed by South China (Guangxi, n = 80; Guangdong, n = 15), North China (Shanxi, n = 4; Hebei, n = 2), and Central China (Hubei, n = 2). 154 S. Rissen isolates were collected from humans, of which 51.9% were females (Table 1). The human source S. Rissen isolates were collected from hospitals. The prevalence of S. Rissen has increased in Shanghai from 2012 to 2014. Out of the cases, 47.4% showed symptoms of gastroenteritis infection, including one case that occurred in an outbreak. The majority of the cases (80.5%, 124/154) occurred from July to September. S. Rissen was mainly detected in people aged 21–40 (n = 58, 37.7%) and 41–60 (n = 44, 28.6%) years old. A total of 74 non-human source S. Rissen strains were identified (Table 1). About 74.3% of the isolates come from foods of animal origin, and 24.3% of the isolates come from environmental samples. The most common source was pork (43.2%, 32/74), followed by rivers/lakes (24.3%, 18/74).
 |
Table 1 The Epidemiological Characteristics of Human Source and Non-Human Source Salmonella rissen Isolates |
Antimicrobial Resistance
Of the 228 S. Rissen isolates, 93.4% (213/228) isolates were resistant to at least one of the 12 tested antimicrobial agents. The results in Table 2, showed that S. Rissen isolates revealed high resistance to tetracycline (91.2%, 208/228), trimethoprim-sulfamethoxazole (74.1%, 169/228) and ampicillin (67.5%, 157/228). Likewise, the resistance to sulfafurazole was 41.2%, chloramphenicol (39.5%), ciprofloxacin (24.1%), amoxicillin-clavulanic acid (22.8%), nalidixic acid (11.0%) and 9.2% to cefoxitin. Moreover, almost all the isolates were sensitive to ceftriaxone (99.1%, 226/228) and azithromycin (99.1%, 226/228), followed by gentamicin (89.5%) and nalidixic acid (89.0%). Notably, the resistance rates to chloramphenicol, quinolones and sulfafurazole were significantly higher in strains isolated from human compared to non-human source strains, while the resistance rate to ampicillin was significantly lower in strains isolated from human than in non-human source strains (p<0.05) (Figure 1). The 14 tested antimicrobial agents were divided into seven categories: tetracyclines, aminoglycosides, sulfonamides, chloramphenicols, quinolones, macrolides, and β-lactams. High frequencies of resistance were observed for tetracyclines (91.2%, 208/228), sulfonamides (75.9%, 173/228), and β-lactams (69.7%, 159/228). Almost all the isolates were susceptible to macrolides (99.1%, 226/228).
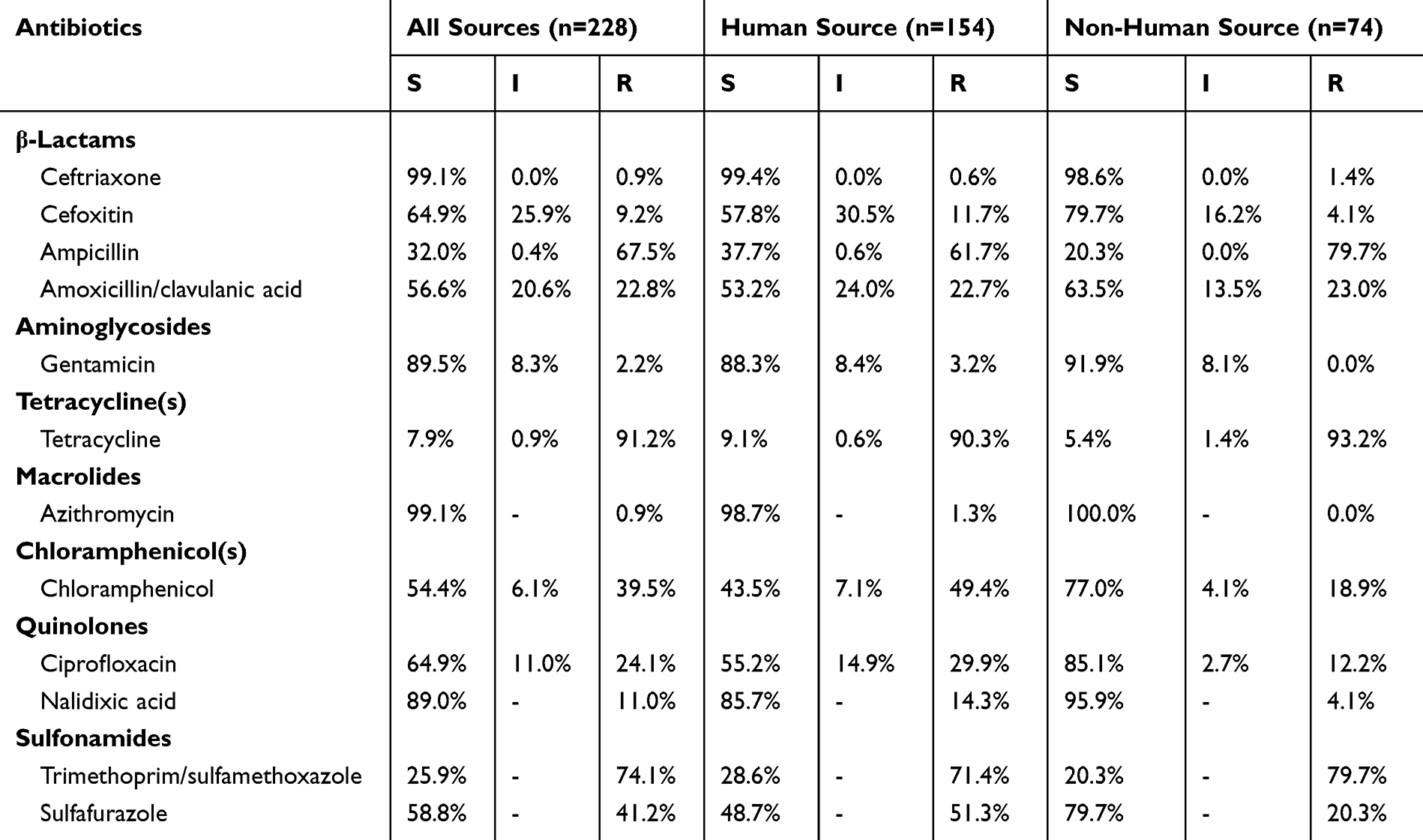 |
Table 2 The Antimicrobial Susceptibility Profiles of Human Source and Non-Human Source Salmonella rissen Isolates |
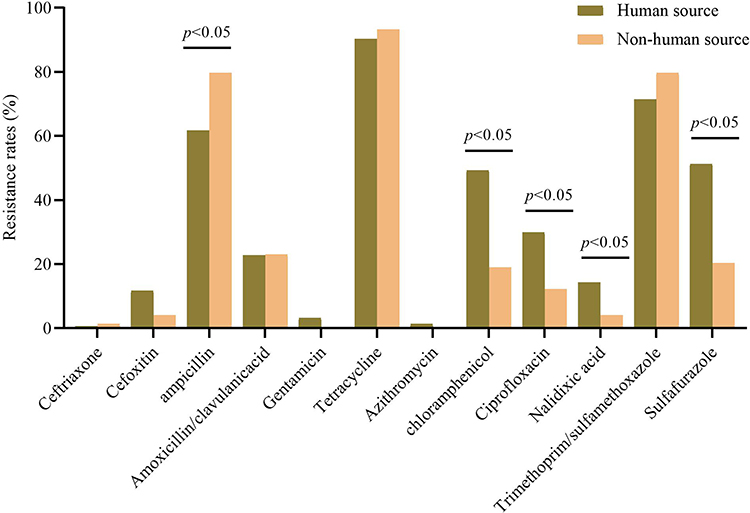 |
Figure 1 The resistance rates of human source and non-human source Salmonella rissen isolates. |
A total of 171 (75%) isolates were resistant to at least three categories of antimicrobials, and the most common resistance profile was Tetracycline(s)-β-Lactams-Sulfonamides, which was represented by 35.1% (80/228). Twelve multidrug resistant (MDR) patterns were detected among these isolates, including three isolates that exhibited resistance to six categories of antimicrobials (Tetracycline(s)-β-Lactams-Aminoglycosides-Chloramphenicol(s)-Quinolones-Sulfonamides/2, Tetracycline(s)-β-Lactams-Macrolides-Chloramphenicol(s)-Quinolones-Sulfonamides/1) (Table 3).
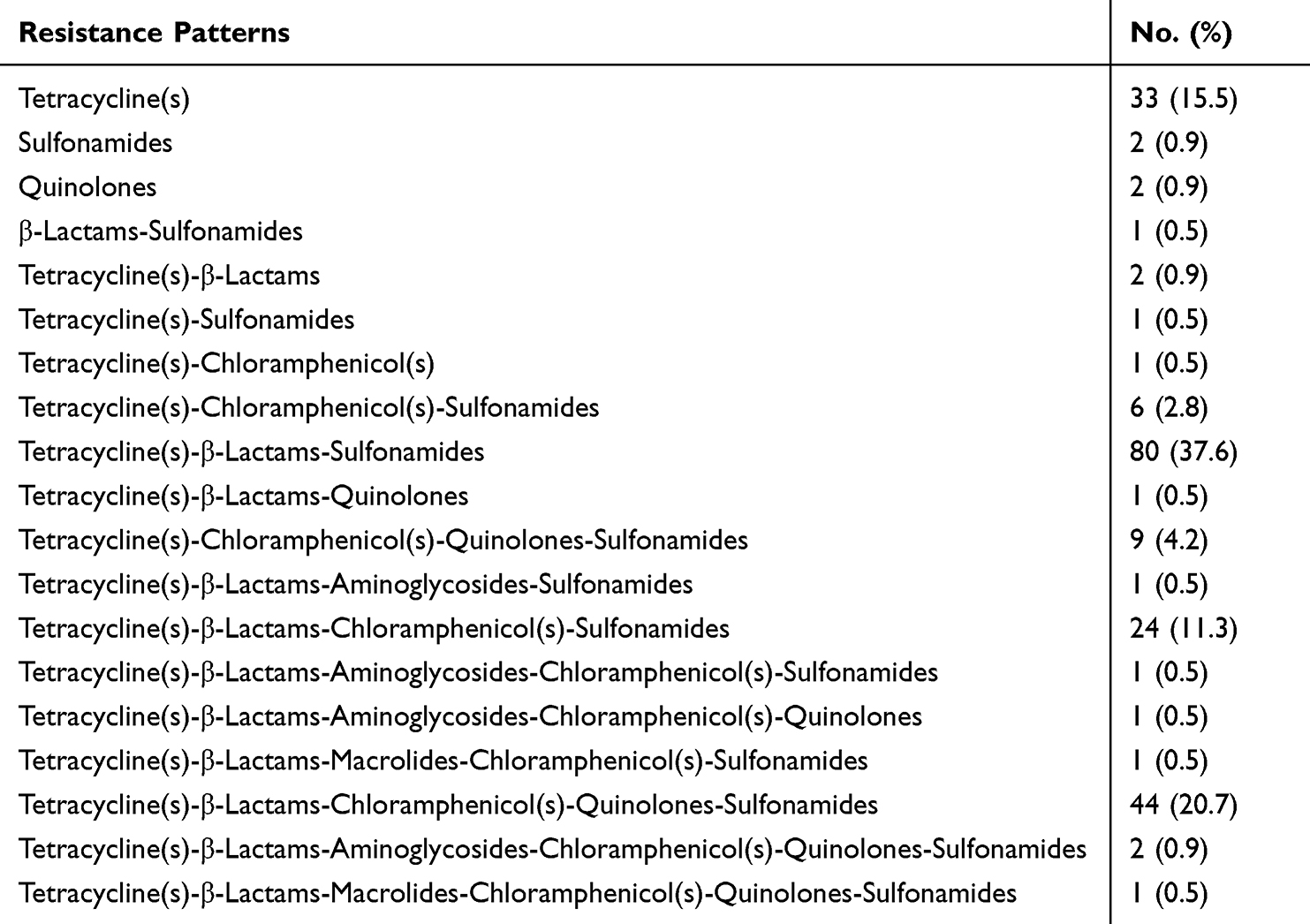 |
Table 3 The Resistance Patterns Among Salmonella rissen Isolates |
Detection of Resistance-Associated Genes
About half of the β-lactamase-resistant isolates obtained produced amplicons specific to blaTEM, but none of the β-lactamase-resistant isolates from all sources produced amplicons specific to blaOXA (Table 4). Most of the TET-resistant isolates (81.3%) produced amplicons specific to tetA, followed by tetB (19.7%). Half of the SMX-resistant isolates obtained produced amplicons specific to sul2/sul3, and only one isolate from a patient produced sul1 and dfr specific amplicons. Among the 5 GEN-resistant isolates from human, one isolate produced strA and strB specific amplicon gene and two isolates produced the aadA gene. None of chloramphenicol-resistant isolates carried the cat gene. We also screened genes associated with class one integron in the 228 isolates, and all strains harbored intl1 VR (cx). The frequencies of carrying intl1 and qacED1 were 71.5% (163/228) and 46.5% (106/228), respectively.
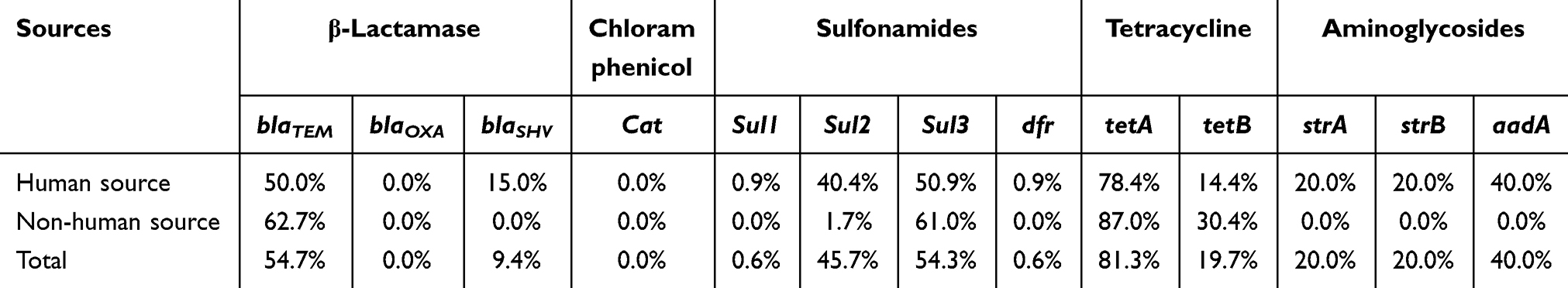 |
Table 4 The Distribution of Antimicrobial Resistance Genes in Salmonella Rissen Isolates from Different Sources |
Detection of Virulence Genes
All of the isolates were found to harbor the virulence genes hilA, sopB, sciN, stn, and ssrB. The genes ssaQ and mgtC were detected in the majority of the isolates (98.7%), followed by invA (98.2%, 224/228). The occurrence of genes pltA (5.3%, 12/228) and safB (2.2%, 5/228) was relatively low. None of the isolates carried the genes spi4D and viaB. There were no significant differences observed in the virulence genes between isolates obtained from human and non-human source.
Molecular Typing by PFGE
The isolates were grouped into four major clusters (A to D) and four individual distinct profiles with 80% pattern similarity (Figure S1). A total of 93 PFGE patterns were generated for the 228 isolates tested, with a similarity index ranging from 59.4% to 100%. Of which, thirty PFGE patterns containing at least two isolates that were identical. Cluster A was the largest and comprised 91.7% (209/228) of the total number of isolates, including 140 human and 69 non-human sources (one from environment and the other from animals). The second most common cluster (cluster B, n = 6) included isolates from human, chicken and river. The cluster C included five isolates, all derived from patient feces; cluster D consisted of four isolates, all derived from patient feces.
Discussion
Foodborne salmonellosis has emerged as a significant global public health concern. S. Rissen, a zoonotic bacterium, has been linked to a growing number of human infections.11,16 The consumption of contaminated food or water facilitates the spread and infection of this pathogen in both humans and animals.17
In this study, the almost equal distribution between male and female patients (51.9% females) indicates that gender does not significantly influence the likelihood of infection. However, the high prevalence in age groups of 21–40 and 41–60 years was observed, which could be related to lifestyle, dietary habits, or higher exposure to risk factors. The majority of the cases (80.5%) occurring in summer aligns with the common understanding that Salmonella infections often peak in the warmer months, potentially due to increased foodborne transmission routes. This might be the reason that Salmonella infections in Thailand are evenly distributed throughout the four seasons.18 The most common source of S. Rissen from non-human source was pork, which was consistent with findings from other studies.6,16 The identification of S. Rissen in rivers and lakes (24.3%) suggests environmental reservoirs may be potential sources of contamination, emphasizing the importance of controlling the spread of S. Rissen in humans, animals and the environment.
The abuse of antimicrobials in human, animal husbandry, and poultry farming has led to frequent drug resistance. The majority (91.2%) of S. Rissen isolates from all sources in this study exhibited resistance to tetracycline and displayed limited resistance to other antimicrobials. Some studies also found 84–93% of isolates were resistant to tetracycline in Thailand, while half (48.6%) of the US isolates were resistant to TET.9,11 Similar to findings in Thailand,9 S. Rissen isolates in China exhibited showed high frequencies of resistance for several routine antimicrobials, such as tetracyclines, sulfonamides and β-lactams. The finding that 75% of isolates were resistant to at least three categories of antimicrobials and the identification of common resistance profiles (eg, Tetracyclin-β-Lactams-Sulfonamides) pointed to significant MDR challenges, which necessitates careful consideration in choosing empirical treatments and highlights the need for routine susceptibility testing in clinical practice. It is worth noting that the resistance rates to chloramphenicol, quinolones and sulfafurazole were significantly higher in strains isolated from human compared to non-human source strains. The observed differences in resistance patterns between human and non-human source strains could be indicative of different selective pressures or exposure to antimicrobial agents in clinical settings compared to environmental or food source.
Compared with the data reported by Spain in 2015, the profiles of resistance gene patterns in these two countries shared something in common and had differences as well, the common lies at for tetracycline resistance, tetA is the predominant genes. The difference is that sul1 and sul3 were the main genes mediated sulfonamide resistance in Spain but in our case were sul2 and sul3.16 Furthermore, the cat gene was not detected in chloramphenicol-resistant strains. It is necessary to expand the screening range of resistance genes and conduct further research and analysis. In our study, resistance genes are widespread in S. Rissen, which was essential for the development of new methods to control these zoonotic diseases. Virulence genes like hilA, ssaQ, invA, sopB, mgtC, sciN and ssrB were present in majority of the study isolates as was reported in other studies.19,20 The high detection rate of virulence genes in these isolates highlighted their pathogenic potential, which could lead to serious salmonellosis and pose a threat to public health.
It was surprising to discover that the majority (91.7%) of S. Rissen isolates in this study showed a high level of similarity (>80%) in PFGE patterns, for S. Rissen was generally considered to be a genetically diverse serovar. Moreover, indistinguishable patterns were discovered to be shared by several isolates from patients, foods and environment, including some isolates from another province. Previous reports have provided proofs that S. Rissen clinical cases were linked to contaminated pork meat, meat handlers or other food sources, even between different countries.11,16,21 We also observed the formation of several sub-clusters, where isolates from patients, foods, and river water shared the same PFGE pattern and antimicrobial resistance profile. This suggested that multiple S. Rissen clones are circulating in China. Therefore, our data suggested clonal dissemination of S. Rissen might exist, probably not only in China. It was necessary to collect more epidemiological information and isolates from other provinces to support that postulation.
A limitation of this study is that we were unable to include isolates from Thailand, which has been reported as the main source of S. Rissen isolates.6,18 Isolates from Denmark and the USA were found to be linked to those from Thailand, and it was suggested that traveling to Thailand and consuming pork products imported from Thailand were potential risk factors for S. Rissen infections.9,11 Therefore, it was meaningful to compare the characteristics of S. Rissen isolates from China and Thailand.
Conclusions
We described several epidemiological, phenotypic and genotypic characterizations of S. Rissen in China. The high frequencies of multidrug resistance and probable clonal dissemination in this serovar highlighted the necessity of systematic surveillance on S. Rissen in China.
Ethical Aspects
All specimens from patients and other sources were collected as part of the surveillance on Salmonella, according to the national guidelines in China. Consequently, informed consent was not sought from patients, and the study was approved by Shanghai Municipal Center for Disease Control and Prevention Ethical Review Committee.
Funding
This study was supported by grants from National Natural Science Foundation of China (Grant No. 81971990).
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Majowicz SE, Musto J, Scallan E, et al. The global burden of nontyphoidal Salmonella gastroenteritis. Clinical Infect Dis. 2010;50(6):882–889. doi:10.1086/650733
2. Guibourdenche M, Roggentin P, Mikoleit M, et al. Supplement 2003–2007 (No. 47) to the White-Kauffmann-Le minor scheme. Res Microbiol. 2010;161(1):26–29. doi:10.1016/j.resmic.2009.10.002
3. Wang Z, Zhang Y, Xu H, et al. Whole-genome sequencing analysis reveals pig as the main reservoir for persistent evolution of Salmonella enterica serovar rissen causing human salmonellosis. Food Res Int. 2022;154:111007. doi:10.1016/j.foodres.2022.111007
4. Ferrari RG, Rosario DKA, Cunha-Neto A, Mano SB, Figueiredo EES, Conte-Junior CA. Worldwide epidemiology of Salmonella serovars in animal-based foods: a meta-analysis. Appl Environ Microbiol. 2019;85(14):1.
5. Kumar R, Surendran PK, Thampuran N. Distribution and genotypic characterization of Salmonella serovars isolated from tropical seafood of Cochin, India. J Appl Microbiol. 2009;106(2):515–524. doi:10.1111/j.1365-2672.2008.04020.x
6. Trongjit S, Angkititrakul S, Tuttle RE, Poungseree J, Padungtod P, Chuanchuen R. Prevalence and antimicrobial resistance in Salmonella enterica isolated from broiler chickens, pigs and meat products in Thailand-Cambodia border provinces. Microbiol Immunol. 2017;61(1):23–33. doi:10.1111/1348-0421.12462
7. Boschi T, Aquilini D, Degl’Innocenti R, et al. Cluster of cases of Salmonella enterica serotype Rissen infection in a general hospital, Italy, 2007. Zoonoses Public Health. 2010;57(7–8):518–522. doi:10.1111/j.1863-2378.2009.01272.x
8. Flynn D. How Salmonella rissen came to America. Available from:. http://www.foodsafetynews.com/2010/08/how-salmonella-rissen-came-to-america/#.WV9PPojyvIV.
9. Pornsukarom S, Patchanee P, Erdman M, et al. Comparative phenotypic and genotypic analyses of Salmonella Rissen that originated from food animals in Thailand and United States. Zoonoses Public Health. 2015;62(2):151–158. doi:10.1111/zph.12144
10. Xu X, Biswas S, Gu G, Elbediwi M, Li Y, Yue M. Characterization of multidrug resistance patterns of emerging Salmonella enterica serovar rissen along the food chain in China. Antibiotics. 2020;9(10):660. doi:10.3390/antibiotics9100660
11. Hendriksen RS, Bangtrakulnonth A, Pulsrikarn C, et al. Antimicrobial resistance and molecular epidemiology of Salmonella rissen from animals, food products, and patients in Thailand and Denmark. Foodborne Pathogens Dis. 2008;5(5):605–619. doi:10.1089/fpd.2007.0075
12. ISO. Microbiology of Food and Animal Feeding Stuffs. Horizontal Method for the Detection of Salmonella Spp. London: British Standard Institute; 2002.
13. World Health Organization. Isolation of Salmonella and Shigella from Faecal Specimens. Geneva, Swizerland: WHO Global Foodborne Infections Network; 2010.
14. Popoff MY, Bockemuhl J, Gheesling LL. Supplement 2002 (no. 46) to the Kauffmann-White scheme. Res Microbiol. 2004;155(7):568–570. doi:10.1016/j.resmic.2004.04.005
15. Dutta S, Das S, Mitra U, et al. Antimicrobial resistance, virulence profiles and molecular subtypes of Salmonella enterica serovars typhi and paratyphi A blood isolates from Kolkata, India during 2009–2013. PLoS One. 2014;9(8):e101347. PONE-D-13-48475 [pii]. doi:10.1371/journal.pone.0101347
16. Garcia-Fierro R, Montero I, Bances M, Gonzalez-Hevia MA, Rodicio MR. Antimicrobial drug resistance and molecular typing of Salmonella enterica serovar rissen from different sources. Microb Drug Resist. 2016;22(3):211–217. doi:10.1089/mdr.2015.0161
17. Senior K. Estimating the global burden of foodborne disease. Lancet Infect Dis. 2009;9(2):80–81. doi:10.1016/S1473-3099(09)70008-8
18. Hendriksen RS, Bangtrakulnonth A, Pulsrikarn C, et al. Risk factors and epidemiology of the ten most common Salmonella serovars from patients in Thailand: 2002–2007. Foodborne Pathogens Dis. 2009;6(8):1009–1019. doi:10.1089/fpd.2008.0245
19. Chowdhury G, Debnath F, Bardhan M, et al. Foodborne outbreak by Salmonella enterica serovar weltevreden in West Bengal, India. Foodborne Pathogens Dis. 2024. doi:10.1089/fpd.2023.0064
20. Parvathi A, Vijayan J, Murali G, Chandran P. Comparative virulence genotyping and antimicrobial susceptibility profiling of environmental and clinical Salmonella enterica from Cochin, India. Curr Microbiol. 2011;62(1):21–26. doi:10.1007/s00284-010-9665-7
21. Gomes-Neves E, Antunes P, Tavares A, et al. Salmonella cross-contamination in swine abattoirs in Portugal: carcasses, meat and meat handlers. Int J Food Microbiol. 2012;157(1):82–87. doi:10.1016/j.ijfoodmicro.2012.04.015
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.