Back to Journals » Clinical Interventions in Aging » Volume 20
Beyond Hormone Replacement: Multifaceted Effects of Phytoestrogens for Optimizing Kinesiological and Physiological Adaptations in Postmenopausal Women
Authors Hu Y, Hu Y, Li T, Shi R
Received 29 May 2025
Accepted for publication 9 September 2025
Published 8 October 2025 Volume 2025:20 Pages 1695—1711
DOI https://doi.org/10.2147/CIA.S539923
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Maddalena Illario
Yingjia Hu, Yi Hu, Tao Li, Rengfei Shi
School of Health and Exercise, Shanghai University of Sport, Shanghai, People’s Republic of China
Correspondence: Rengfei Shi, School of Health and Exercise, Shanghai University of Sport, Shanghai, People’s Republic of China, Tel +86 13661498054, Email [email protected]
Abstract: Phytoestrogens (PEs), a class of naturally occurring plant compounds primarily categorized into isoflavones, lignans, flavonoids, coumarins, and stilbenes, exhibit structural similarity to endogenous estrogens and exert regulatory effects through estrogen receptors. This comprehensive review examines the multifaceted roles of PEs in enhancing exercise performance and promoting health among postmenopausal women. Current evidence demonstrates that PEs not only ameliorate characteristic menopausal symptoms but, more significantly, improve physical function through multiple mechanisms: (1) augmenting muscle protein synthesis while mitigating inflammation and oxidative stress to optimize muscular performance; (2) modulating glucolipid metabolism and cardiovascular function to establish physiological foundations for exercise; and (3) preserving bone mineral density and regulating neurotransmitter activity to maintain motor coordination. Although combined PE-exercise interventions demonstrate synergistic benefits, their efficacy is influenced by dosage variations and interindividual metabolic differences. Future investigations should prioritize the development of precision PE applications to optimize kinesiological outcomes and health parameters in postmenopausal populations.
Keywords: phytoestrogens, elderly, exercise performance, physical activity, menopausal symptoms
Introduction
Menopausal syndrome (MPS), sometimes referred to as menopausal symptoms, refers to a cluster of symptoms arising in women around the time of menopause due to dysregulation of the hypothalamic-pituitary-ovarian axis, leading to a rapid decline in endogenous estrogen levels.1 With increasing global life expectancy, over one-third of postmenopausal women spend their lives in a state of chronic estrogen deficiency. It is projected that by 2030, the female population in China experiencing menopause and postmenopause will reach 210 million, with more than 85% potentially experiencing varying degrees of menopausal symptoms;2 worldwide, this demographic is expected to grow to 1.2 billion, with approximately 47 million new cases annually.3
Phytoestrogens (PEs) are a class of naturally occurring plant-derived compounds exhibiting estrogenic activity. Structurally like 17β-estradiol, they can competitively bind to estrogen receptors, exerting selective tissue modulation effects. Based on chemical structure, PEs are primarily classified into isoflavones (eg, genistein, daidzein), lignans (eg, secoisolariciresinol), flavonoids, coumarins (eg, coumestrol), and stilbenes (eg, resveratrol). Variations exist in metabolism and bioactivity among different subclasses and within the same class. Given concerns regarding the potential risks associated with hormone replacement therapy (HRT), PEs have garnered attention for their estrogen-like effects and relative safety, demonstrating benefits in alleviating vasomotor symptoms, maintaining bone mineral density, and modulating lipid metabolism in postmenopausal women.4–6
Beyond typical menopausal symptoms, estrogen deficiency significantly impacts physical function, exercise performance, and quality of life by accelerating sarcopenia, reducing bone density, impairing endurance and balance, and delaying recovery from physical activity.7,8 However, the effects of PEs on specific physical performance indicators such as strength and endurance, as well as physiological adaptations induced by exercise training—encompassing musculoskeletal, cardiovascular, and nervous system responses—remain insufficiently studied and systematically organized. As a potential alternative or adjunctive strategy, PEs are increasingly recognized for their role in alleviating menopausal symptoms and potentially maintaining or enhancing physical capacity and physiological adaptability. Optimizing exercise interventions to improve quality of life, health management, and reduce the incidence of MPS constitutes a pressing public health challenge.
This review aims to systematically synthesize the current evidence regarding the effects of PEs alone or in combination with exercise training on physical performance and underlying physiological adaptations in postmenopausal women. It critically evaluates the potential influence of PEs on exercise training adaptation and recovery processes, identifies existing knowledge gaps, and provides a scientific basis for developing PE-based exercise intervention strategies to enhance physiological functions and exercise capacity in this population.
Literature Search Methodology
This study employs a narrative review methodology, retrieving literature published between 2001 and 2025 from PubMed, Web of Science, and China National Knowledge Infrastructure (CNKI) databases. The search keywords include “phytoestrogens”, “postmenopausal women”, “menopause”, “lignans”, “isoflavones”, “exercise”, “physical activity”, “muscle strength”, and combinations with terms such as “clinical trial” and “mechanism”. Inclusion criteria encompass original research articles or meta-analyses involving human subjects or relevant animal models, published in both Chinese and English. Exclusion criteria include reviews, conference abstracts, and low-quality studies lacking control groups. A total of 138 articles were selected, with a focus on high-evidence-level studies such as randomized controlled trials and mechanistic investigations for comprehensive analysis.
Phytoestrogens and Menopausal Syndrome
Major Symptoms of Menopausal Syndrome
According to the World Health Organization (WHO), the average age of natural menopause globally ranges from 45 to 55 years.9 Approximately 80% of women experience typical menopausal symptoms, primarily vasomotor disturbances such as hot flashes and night sweats;10 neurocognitive issues including anxiety and depression;11 and cardiovascular-metabolic abnormalities characterized by elevated total cholesterol (TC) and low-density lipoprotein cholesterol (LDL-C).12 These physiological changes significantly impair quality of life and predispose women to multidimensional declines in physical function,13 including neuromuscular weakening,14 reduced maximal oxygen uptake (VO2max),15 and increased incidence of osteoarthritis,16 thereby creating a vicious cycle of “metabolic-physical function” deterioration. These pathological alterations are positively correlated with increased risks of chronic diseases in old age.
PEs as a Therapeutic Modality for Menopausal Symptoms
Hormone replacement therapy (HRT) remains the conventional treatment for menopausal syndrome. Initiation of HRT before or after menopause confers primary prevention benefits against osteoporotic fractures.17 However, prolonged HRT use may adversely affect the nervous system, especially when estrogen is administered alone. Morrison et al conducted a randomized controlled trial (RCT) demonstrating that estrogen therapy was ineffective in treating mild to moderate depression in menopausal women, with progesterone supplementation showing no significant improvement in depressive symptoms.18 From a safety perspective, patients requiring long-term HRT should actively explore alternative strategies that harness estrogenic effects while minimizing potential adverse reactions.
In contrast, PEs, as naturally occurring plant-derived estrogens, exert physiological effects through differential activation of estrogen receptor alpha (ERα) and estrogen receptor beta (ERβ). They can mimic endogenous estrogen, reduce lipid levels, and demonstrate significant potential in lowering the risk of coronary heart disease (CHD).19 The British Menopause Society recommends that low-dose natural PEs be used to treat menopausal symptoms, with minimal risk of venous thromboembolism,20 thus providing a safer management option for menopausal syndrome.
Common Types of PEs and Their Roles in Menopausal Symptom Relief
Isoflavones (ISO)
Isoflavones are a class of PEs predominantly found in leguminous plants, with active forms including daidzein, genistein, and puerarin.21–23 Soybeans and their products—such as tofu, soy milk, and natto—serve as high-quality dietary sources, containing approximately 128 milligrams of isoflavones per 100 grams of soybeans.24 The consumption of legume-based diets is widespread across Asia, with traditional Japanese dietary patterns exemplifying this trend.25
Multiple studies have demonstrated that soybean isoflavones can significantly reduce the frequency and severity of hot flashes and alleviate symptoms such as night sweats.26,27 Their mechanism of action may involve mimicking estrogenic activity and modulating estrogen receptor activity, potentially reducing the risk of hormone-dependent cancers, including cervical, breast, and prostate cancers.28–32 It is noteworthy that the bioavailability of isoflavones is influenced by gut microbiota metabolism,33 and the synergistic effects of isoflavones with physical activity interventions, such as resistance training, warrant further investigation.
Lignans
Lignans are a class of polyphenolic compounds widely distributed in plants, with flaxseed being the richest dietary source, containing up to 53,000 micrograms per 100 grams.34 Gut microbiota metabolize lignans into enterolactone, a compound structurally similar to estrogen, conferring a unique bidirectional regulatory capacity: alleviating menopausal symptoms while acting as antagonists in high-estrogen environments.35,36
In cardiovascular health, specific lignans such as matairesinol have been shown to improve endothelial function,37 while the intake of lariciresinol is associated with a 30% reduction in the risk of hypercholesterolemia.38 Clinical evidence indicates that flaxseed consumption can significantly lower total cholesterol levels in postmenopausal women,39 and lignans in sesame oil exert protective effects through modulation of lipid metabolism.40 Regarding metabolic health, the metabolite enterolactone correlates with a reduced risk of type 2 diabetes, particularly among obese individuals and postmenopausal women.41
However, the health effects of lignans exhibit considerable individual variability. Western populations, which consume higher amounts of flaxseed and sesame, tend to have greater lignan intake compared to Eastern populations.42 Some individuals lack key metabolic enzymes necessary for efficient conversion of lignans into bioactive forms, a process closely linked to gut microbiota composition and metabolic capacity.43 Personalized intervention strategies should account for these differences.
Flavonoids
Flavonoids are a widespread class of polyphenolic compounds in plants, exhibiting notable value in menopausal health management. These compounds possess significant anti-inflammatory and antioxidant properties,44,45 with representative constituents such as apigenin and quercetin demonstrating phytoestrogenic activity.
Apigenin is distinguished by its potent antioxidant capacity, prevalent in temperate fruits and vegetables, effectively scavenging free radicals and reducing oxidative stress-induced damage.46 Quercetin offers broader health benefits, including improvements in glycemic and lipid profiles,47 and exhibits multi-target therapeutic potential in breast cancer prevention and treatment.48 Animal studies have confirmed that quercetin supplementation can markedly improve metabolic parameters and enhance antioxidant enzyme activity in ovariectomized rat models.49
It is important to note that, despite the presence of flavonoids in common foods such as apples and broccoli, their health benefits remain subject to debate. For instance, flavonoid intake has not been consistently associated with a reduced risk of cancer.50 While chocolate, a flavonoid-rich processed food, can improve arterial stiffness, its effects on vascular structure are limited.51 These discrepancies underscore the importance of considering flavonoid types and individual characteristics when evaluating their health impacts.
Coumarins
Coumarin compounds are a class of naturally occurring substances characterized by distinctive aromatic properties, demonstrating multifaceted applications in the maintenance of health among menopausal women. The core structure of these compounds is benzopyranone, which can be subdivided into various types, such as simple coumarins and furanocoumarins, based on different substituents.
In the field of exercise physiology, coumarins exert significant vasodilatory effects that enhance blood circulation,52 which is particularly beneficial for menopausal women engaging in aerobic activities, thereby improving cardiopulmonary function and alleviating exercise-induced fatigue. Regarding oncological prevention and treatment, specific coumarin derivatives act through targeting the HDAC1-Sp1-FOSL2 signaling axis or by inhibiting NUDT5,53,54 thereby exerting anti-breast cancer effects. Additionally, 7,8-dihydroxy-4-methylcoumarin (DHMC) not only reduces fasting blood glucose levels but also exhibits notable anxiolytic properties,55 offering potential solutions for common menopausal issues.
Stilbenes
Stilbene compounds are vital secondary metabolites produced by plants to defend against environmental stressors,56 with high concentrations found in grapes, peanuts, and Polygonum multiflorum.57,58 Among these, resveratrol is the most representative, distinguished by its excellent antioxidant and anti-inflammatory properties, which are prominent in supporting menopausal health.59 In cancer prevention, multiple stilbenes derived from muscat grapes demonstrate synergistic effects against triple-negative breast cancer cells that surpass those of resveratrol alone.60,61 In the context of exercise health, clinical studies confirm that resveratrol supplementation effectively reduces exercise-induced muscle damage and enhances athletic performance, which is significant for improving physical capacity in menopausal women.62 These findings suggest that stilbene compounds may serve as effective adjuncts in the health management of menopausal women.
Absorption and Utilization Mechanisms of PEs in the Human Body
PEs are naturally occurring plant secondary metabolites structurally and functionally like mammalian estrogens, exhibiting diverse biological activities. Compared to endogenous estrogens such as 17β-estradiol, PEs are ingested via the gastrointestinal tract and undergo complex processes including digestion, absorption, metabolism, distribution, binding, and excretion. The absorption and utilization of PEs constitute a dynamic, multi-step regulatory process, with their ultimate biological effects dependent on intricate interactions among the host, microbiota, and diet.
Endogenous estrogens, notably 17β-estradiol, are highly active, characterized by a tetracyclic structure comprising rings A, B, C, and D, with ring A containing a phenolic hydroxyl group. The chemical similarity between PEs and endogenous estrogens enables PEs to bind to estrogen receptors ERα and ERβ, thereby exhibiting estrogen-like biological activity (Figure 1). For instance, the A and C rings of isoflavone-type PEs resemble the A and D rings of 17-β-estradiol, both containing phenolic hydroxyl groups; lignan and coumarin derivatives feature phenyl rings and hydroxyl groups structurally similar to those in endogenous estrogens.
 |
Figure 1 Structural formula of 17-β-estradiol and representative PEs. |
The bioavailability of PEs is jointly regulated by their chemical structure, metabolic transformation, and individual host factors. The absorption and utilization process can be delineated into key stages: initial breakdown and absorption in the gastrointestinal tract; subsequent microbial metabolism that releases active constituents into the bloodstream; and finally, binding to estrogen receptors to exert biological effects.
Absorption, Distribution, and Metabolism
PEs predominantly exist in plants in the form of glycosides. Upon oral ingestion, β-glucosidases present in gastric acid and the intestinal tract hydrolyze glycosidic bonds, releasing bioactive aglycones. For example, in isoflavone-rich PEs, the soy aglycone can be metabolized by gut microbiota into more potent metabolites such as equol (Figure 2). Similarly, lignan-type PEs, including matairesinol and seco-isolariciresinol, are converted by intestinal bacteria into enterodiol and enterolactone (Figure 3), which exhibit enhanced antioxidant activity and biological effects.
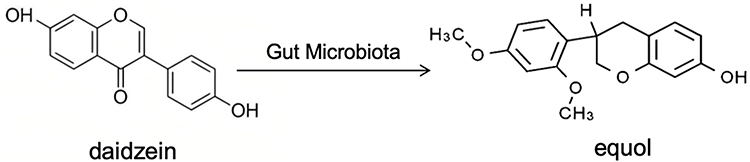 |
Figure 2 Absorption and metabolism of daidzein. |
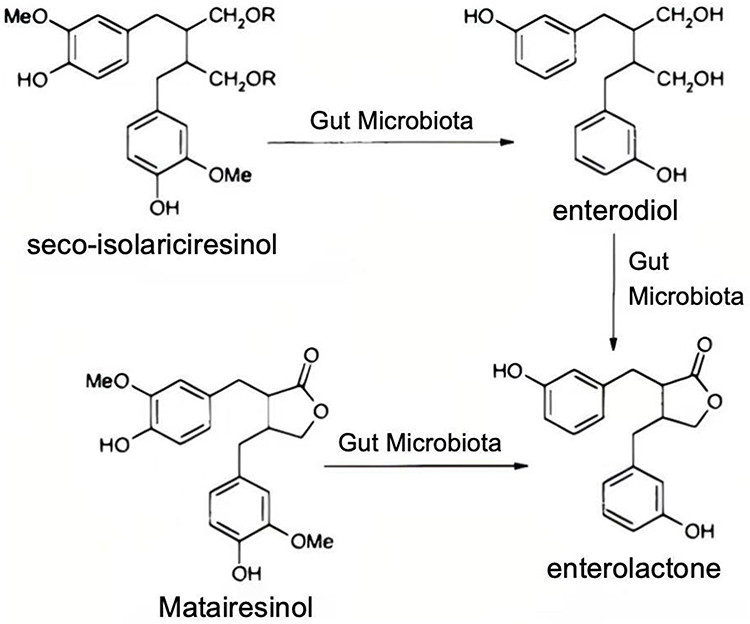 |
Figure 3 Absorption and metabolism of lignans. |
Subsequently, free aglycones traverse the intestinal epithelium via passive diffusion or active transport mechanisms, then enter the portal circulation to reach the liver. They undergo Phase I metabolism (oxidation, reduction, hydrolysis) and Phase II conjugation reactions, transforming into water-soluble metabolites that are distributed via the bloodstream to target tissues.
Receptor Binding
Estrogen receptors (ER) are primarily classified into two subtypes: ERα and ERβ. ERα is predominantly expressed in tissues such as the uterus, mammary glands, and ovaries, which are highly responsive to estrogen. Conversely, ERβ is highly expressed in the brain, lungs, and prostate, serving as the main binding site in these tissues.63
Like endogenous estrogen, PEs interact with these receptors, exhibiting comparable distribution and functional mechanisms. Their metabolites exert biological effects mainly through two pathways (Figure 4).
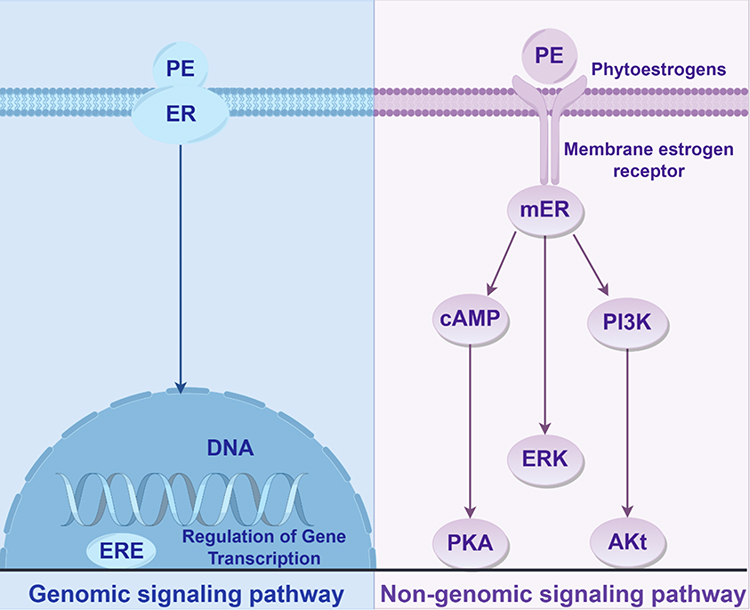 |
Figure 4 Genomic and non-genomic pathways of PEs binding to receptors. |
Classical Estrogen Receptor Pathway
PEs competitively bind to estrogen receptors, activating classical genomic responses. The PE-receptor complex dimerizes and translocates into the nucleus, where it binds to estrogen response elements (EREs), thereby regulating the transcription of target genes.64 This gene expression modulation mimics estrogen’s physiological effects, contributing to the regulation of conditions such as osteoporosis and cardiovascular diseases in postmenopausal women.65
Non-Genomic Signaling Pathways
PE can also bind to membrane-associated estrogen receptors (mER), activating rapid non-genomic signaling cascades such as PI3K/Akt, ERK1/2, and cAMP/PKA pathways. These signals are typically completed within seconds to minutes and enable the swift regulation of cellular functions and metabolic processes.
Excretion and Enterohepatic Circulation
The metabolic fate of PE significantly influences its bioactivity and health benefits. In vivo, PE metabolites are primarily excreted via the kidneys, with approximately 60–70% in the form of glucuronide conjugates in urine, while the remainder is excreted into the intestine via bile. Gut microbiota’s β-glucuronidases hydrolyze these conjugates, allowing free aglycones to be reabsorbed and participate in enterohepatic circulation, thereby markedly prolonging their half-life.66 Studies indicate that the metabolism of soy isoflavones is relatively slow, with the half-life of soy aglycones reaching up to 8 hours.67 This characteristic underpins the potential of soy isoflavones to modulate estrogen-related physiological functions and supports their prospective role in preventing and managing estrogen-associated diseases.
Effects of PEs on Exercise Capacity in Postmenopausal Women and Underlying Mechanisms
The decline in estrogen levels during menopause leads to reduced muscle protein synthesis, increased oxidative stress, and bone loss, collectively impairing physical performance and overall health. PE can mimic estrogen’s effects, effectively improving exercise tolerance, muscle mass, and bone density, thereby enhancing physical capacity.68
The Effects of PEs on Postmenopausal Women’s Muscle Protein Metabolism and Exercise Capacity
Estrogen deficiency impairs muscle protein synthesis, and PEs, due to their estrogen-like activity, are being explored as potential strategies to improve muscle metabolism. This section focuses on the regulatory mechanisms of PEs on muscle proteins in postmenopausal women and evaluates their practical benefits on exercise capacity.
Molecular Mechanisms of PEs in Regulating Muscle Protein Synthesis
Decreased estrogen levels are closely associated with reduced muscle protein synthesis. Although most mechanistic studies originate from animal models or cell experiments, they provide valuable insights into PEs’ role in menopausal women. Research indicates that estrogen activates the mTOR signaling pathway, promoting ribosomal biogenesis and protein translation, thereby underpinning muscle growth and repair. Isoflavone-rich PE (such as dyewood flavonoids) can activate the Akt/mTOR pathway, upregulate the chaperone protein GRP78, and help maintain cellular protein homeostasis and optimize protein folding efficiency.69,70 Flavonoid-based PE (such as apigenin and quercetin) modulate the MAPK/ERK and PI3K/AKT/mTOR pathways to promote myocyte differentiation and inhibit the NF-κB pathway, reducing TNF-α levels, thereby facilitating muscle regeneration and strength recovery, ultimately enhancing exercise capacity.71–73
Potential Effects of PEs in Suppressing Muscle Protein Degradation
The anti-proteolytic effects of PEs are crucial for maintaining muscle homeostasis. PEs reduce protein degradation by regulating proteasome activity. An RCT involving postmenopausal women demonstrated that daily intake of red clover extract (60 mg of isoflavones) combined with resistance training decreased the expression of proteolytic markers FOXO1/FOXO3a and increased protective heat shock proteins (HSP27). However, this intervention did not activate the mTOR pathway or improve muscle strength, suggesting that long-term intervention or combined training may be necessary for functional gains.61 Animal studies show that genistein inhibits the expression of muscle atrophy-related genes Atrogin-1 and MuRF1 via ERβ pathways.74,75 S-Equol, a metabolite of genistein, alleviates musculoskeletal pain, indicating its role in modulating protein metabolism through estrogen receptor pathways, thereby improving muscle health in postmenopausal women.76
Impact of Combined PEs and Exercise Interventions on Muscle Function: Controversies and Limitations
While estrogen supplementation can enhance strength training outcomes in menopausal women, and PEs such as soy isoflavones theoretically improve muscle function by mimicking estrogen pathways, clinical studies on postmenopausal women’s exercise capacity yield conflicting results.
Some studies support a synergistic effect. A double-blind RCT showed that daily supplementation with 60 mg of soy isoflavones combined with 12 weeks of resistance training increased quadriceps cross-sectional area by 3.8% (P<0.05) and maximal isometric strength by 9.2%.77 Another RCT involving women with sarcopenia confirmed that soy protein-enriched milk combined with 16 weeks of resistance training significantly improved bench press and knee extension 1RM strength.78
Conversely, long-term intervention studies present differing conclusions. Fontvieille et al conducted a one-year trial indicating that although exercise training significantly improved lean body mass and physical function in overweight postmenopausal women, short-term metabolic improvements in muscle tissue were not further enhanced by PE supplementation, suggesting limited short-term metabolic and optimization effects during training.79 Further, Choquette et al demonstrated that resistance plus aerobic training significantly increased maximal muscle strength, muscle mass, and relative strength (all P<0.05), whereas soy isoflavone supplementation alone showed no significant effects.80
In summary, exercise training remains the primary intervention for improving muscle function in postmenopausal women. While PEs may support muscle protein synthesis, current evidence for its clinical additive benefits is limited by heterogeneity in intervention protocols (such as duration, PE type, and dosage) and mechanistic understanding. Presently, PEs cannot be recommended as a universal strategy for muscle enhancement; systematic exercise interventions continue to be the preferred approach. Future research should focus on long-term studies to clarify the precise application and efficacy of PEs in this context.
The Potential Impact of PEs’ Anti-Inflammatory and Antioxidant Properties on Exercise Capacity in Postmenopausal Women
Chronic low-grade inflammation and oxidative stress in postmenopausal women may significantly influence their exercise performance and overall physical adaptability. This section evaluates the evidence supporting PEs’ anti-inflammatory and antioxidant mechanisms and explores their potential effects on exercise endurance and recovery in this population.
Preclinical Foundations of Anti-Inflammatory and Antioxidant Mechanisms and Their Limitations in Extrapolation
PEs exert synergistic anti-inflammatory and antioxidant effects through mechanisms such as inhibition of pro-inflammatory pathways, scavenging reactive oxygen species (ROS), and regulation of antioxidant enzymes. In vitro and animal studies indicate that lignan amides possess phenolic hydroxyl groups,81 which may modulate gut microbiota to reduce post-exercise pro-inflammatory cytokine release, thereby alleviating systemic inflammation induced by high-intensity exercise.82 Resveratrol activates the SirT1/PGC-1α pathway to promote mitochondrial biogenesis, extending exercise duration by enhancing muscular endurance, and improves energy utilization during high-intensity activity via the AMPK pathway.83–85 Flavonoids like flavonoids inhibit ROS-mediated macrophage pyroptosis,86 while quercetin may protect the PI3K/Akt/FOXO3 pathway from exercise-induced cardiac stress, and estrogenic compounds such as equol can modulate activities of antioxidant enzymes like SOD and GPx. These findings suggest that PE could mitigate exercise-induced oxidative damage and inflammatory responses, improve energy metabolism, and theoretically enhance endurance and recovery. However, results from animal models cannot be directly extrapolated to human postmenopausal women.
Human Evidence on the Effects of PEs on Exercise Capacity and Recovery in Postmenopausal Women
Given the well-documented anti-inflammatory properties of PEs and considering the prevalent chronic inflammation and oxidative stress in menopausal women that may significantly impair their physical performance, exploring the synergistic effects of PEs supplementation combined with regular exercise holds substantial clinical relevance. Multiple clinical studies provide compelling evidence supporting this approach.
A recent 12-week RCT demonstrated that combined intervention with soybean and hops extracts—rich in isoflavones—significantly alleviated symptoms such as fatigue, arthritis, and myalgia in menopausal women.87 Although this study did not directly assess exercise performance, the amelioration of these symptoms likely contributes to reduced exercise-induced muscle discomfort and inflammatory responses, thereby facilitating faster muscle recovery and potentially enhancing exercise tolerance.
Further, research by Llaneza et al offers more direct evidence.88 Over a 24-month intervention, participants receiving soy isoflavone supplementation alongside exercise and a Mediterranean diet exhibited not only lower BMI and adiposity but also a marked reduction in tumor necrosis factor-alpha (TNF-α) levels. These findings suggest that PEs may indirectly improve exercise endurance and recovery by improving body composition and attenuating systemic inflammation.
Regarding skeletal muscle adaptability, a study investigating the effects of Shatavari—a herbal supplement containing PEs—found that this phytotherapeutic agent significantly upregulated pathways involved in metabolic regulation and cellular repair.89 This intervention appeared to influence postmenopausal women’s body composition and muscle function markers, indicating that Shatavari’s anti-inflammatory and antioxidant properties may modulate muscle function and mitigate post-exercise muscle inflammation. Nonetheless, the specific impact of Shatavari on exercise-induced inflammatory responses warrants further validation through controlled exercise interventions.
Conversely, the findings of Riesco et al are comparatively conservative.90 Their study indicated that soy isoflavone supplementation did not produce a more significant reduction in serum C-reactive protein (CRP) and leptin levels in postmenopausal women than exercise alone, suggesting that physical activity itself exerts substantial anti-inflammatory effects. However, the authors propose that PEs may exert indirect benefits by modulating other adipokines involved in recovery processes.
In summary, current clinical evidence suggests that specific PEs supplementation may influence exercise capacity in menopausal women through multiple mechanisms, including improvements in body composition, reductions in inflammatory biomarkers such as TNF-α, and attenuation of post-exercise inflammatory responses. These effects may collectively support the maintenance of exercise endurance and optimize recovery, although the magnitude and nature of these benefits likely depend on the type of PEs and intervention protocols employed. Future research should aim to more directly evaluate the immediate and long-term impacts of PEs on exercise performance.
The Impact of PEs on Cardiovascular and Metabolic Health and Exercise Capacity in Postmenopausal Women
The decline in estrogen levels following menopause is closely associated with increased cardiovascular disease risk and disturbances in glucose and lipid metabolism. PEs may indirectly influence exercise capacity by improving these metabolic parameters. This section focuses on the underlying regulatory mechanisms and evidence from population studies to explore the potential effects of PEs on cardiovascular metabolic health and physical performance in postmenopausal women.
Potential Regulatory Mechanisms of PEs on Cardiovascular and Metabolic Health
Preclinical studies suggest that PEs may modulate metabolic functions through multiple pathways, providing a biological basis for their potential effects. In glucose metabolism, lignans and isoflavones can activate estrogen receptors or AMP-activated protein kinase (AMPK) pathways to promote glucose uptake in skeletal muscle and adipose tissue,91,92 thereby enhancing insulin sensitivity and regulating hepatic enzymes critical for glucose metabolism, such as glucokinase (GK) and glucose-6-phosphatase (G6Pase).4,93 Regarding energy metabolism, PEs may influence mitochondrial function via ERs or G protein-coupled estrogen receptor (GPER) signaling,94,95 with ERα playing a key role in maintaining exercise endurance and supporting sustained physical performance.96,97 In lipid metabolism, isoflavones can regulate the expression of lipogenic genes in the liver and adipose tissue, such as SREBP-1c, FASN, and SCD1, thereby improving lipid profiles.98,99 Coumarin derivatives of PEs may also affect fat distribution by activating brown adipose tissue or modulating gut microbiota.100–102 Additionally, some PEs promote endothelial nitric oxide (NO) release, improving endothelium-dependent vasodilation, which is crucial for maintaining effective blood flow during exercise.103
Effects of PEs on Postmenopausal Women: Cardiovascular Metabolic Health and Exercise Capacity
From an exercise physiology perspective, the synergistic effects of PEs on exercise-induced metabolic improvements warrant further investigation. Most current studies assess the potential impact of PEs on exercise capacity through changes in metabolic biomarkers. Multiple studies indicate that PEs may provide a physiological basis for enhanced exercise performance by improving glucose and lipid metabolism and cardiovascular function.
In glucose metabolism, a meta-analysis focusing on non-Asian postmenopausal women reported that soy isoflavone supplementation was associated with significant weight loss, reductions in fasting blood glucose, and decreased insulin levels.104 While these metabolic improvements could theoretically enhance exercise efficiency, direct assessments of exercise performance were not conducted. Notably, an intervention study on insulin-resistant postmenopausal women found that, on a basis of 6–24 months of Mediterranean diet and walking exercise, an additional daily intake of 40 mg soy isoflavones significantly lowered HOMA-IR indices (P<0.05), with effects persisting during follow-up, suggesting that PEs may augment exercise-related improvements in insulin sensitivity, offering additional metabolic benefits.105
Regarding lipid metabolism, optimizing lipid profiles may represent another key mechanism. Llaneza et al observed that while diet control (1200 kcal) and exercise improved adipokine levels (leptin and adiponectin) in obese postmenopausal women, only combined supplementation with 80 mg/day soy isoflavones significantly increased serum adiponectin levels.106 This “exercise-phytoestrogen” synergy may improve metabolic environments through multiple pathways, serving as a critical regulatory mechanism for cardiovascular health in menopausal women and supporting exercise capacity.
In terms of cardiovascular function, physical activity and phytoestrogen supplementation appear to be potential strategies for preventing cardiovascular disease in postmenopausal women; however, current findings are somewhat inconsistent. Dechichi et al reported that in normotensive postmenopausal women, 10 weeks of exercise training significantly reduced blood pressure, but additional supplementation with 100 mg/day soy isoflavones did not produce further effects.107 Conversely, a meta-analysis indicated that daily intake of ≥25 g soy protein containing ≥100 mg/d isoflavones was associated with significant blood pressure reductions.108 These discrepancies may reflect dose-dependent effects of PEs and population-specific responses, as well as the inherent capacity of exercise training to modulate blood pressure.
In summary, existing evidence suggests that PEs may indirectly support exercise capacity by improving metabolic parameters. However, most studies lack direct assessments of exercise performance metrics such as VO2max or endurance time, limiting definitive conclusions regarding their practical efficacy. Future research should systematically investigate the direct effects of PEs on exercise capacity.
The Interactive Effects of PEs on Bone Health Preservation and Motor Function in Perimenopausal Women
Bone health constitutes a central challenge in maintaining health post-menopause. The decline in estrogen levels following menopause accelerates bone loss, thereby increasing fracture risk and impairing key components of motor function. PEs may exert osteoprotective effects through mechanisms such as regulation of osteocyte activity, modulation of metabolic signaling pathways, and maintenance of calcium homeostasis, thereby indirectly supporting motor function.
Molecular and Cellular Mechanisms of PEs in Bone Protection
PEs potentially influence bone metabolism via multiple pathways, primarily evidenced by in vitro studies and ovariectomized animal models, which suggest potential routes for PEs in alleviating orthopedic diseases and improving motor function in postmenopausal women. For instance, equol activates ERβ to regulate the OPG/RANKL balance, promoting osteogenesis;109 isobavachalcone enhances osteogenic differentiation of bone marrow stromal cells (BMSCs);110 and phellodendrine reduce osteoarthritis progression by inhibiting ERK/NFATc1 signaling.111 Additionally, PEs can modulate signaling pathways such as Wnt/β-catenin to promote bone formation and activate the Keap1/Nrf2/ARE antioxidant pathway to protect osteocytes.112,113 The combined supplementation of isoflavones and calcium may also maintain bone mineral density by regulating vitamin D activity, thereby improving calcium homeostasis and metabolic processes.114
Population Evidence: The Impact of PEs on Postmenopausal Bone Health and Its Translation to Motor Function
Building upon the aforementioned potential mechanisms, numerous clinical studies have explored the effects of PEs on bone health in postmenopausal women and their functional implications. Notably, soy isoflavones—known for their specific effects on bone—may influence bone density indirectly by affecting bone metabolic hormones such as parathyroid hormone.115
Meta-analyses indicate that soy isoflavones significantly inhibit bone resorption and stimulate bone formation,116 although effects vary markedly depending on dosage and population. An Italian RCT demonstrated that a daily dose of 54 mg, through bidirectional regulation of bone metabolism, improved lumbar spine and hip Bone Mineral Density (BMD) in early postmenopausal women comparably to HRT, without inducing endometrial hyperplasia.117 Conversely, similar doses in American populations showed limited efficacy in preventing hip osteopenia.118–120
In a North American study, Phani et al observed that consuming 90 g of soy protein containing isoflavones prevented significant femoral trochanter bone loss.121 An Asian study by Chen et al found that an 80 mg/d dose effectively increased bone mineral content (BMC) at the hip, especially at the greater trochanter, in women with low bone mass.122 However, a Taiwanese RCT indicated that a higher dose of 300 mg/d yielded no significant benefits.123 A European study administering 110 mg/d of soy isoflavones in fortified foods for one year failed to slow lumbar or overall skeletal BMD decline or improve bone turnover markers.124 The heterogeneity observed across studies may be attributable to genetic backgrounds, baseline dietary intake, and differences in formulations (pure isoflavones versus soy protein).
The synergistic effects of exercise interventions combined with PEs supplementation exhibit complex dose-response relationships. A two-year RCT revealed that resistance combined with aerobic exercise alone, or with 165 mg/d of isoflavones, provided comparable protection of hip BMD; however, their combination unexpectedly accelerated bone loss, suggesting potential dose-dependent negative interactions.125 The most direct clinical evidence comes from a study on flaxseed lignans, which showed that a daily 50 mg dose over six months significantly increased six-minute walk distance and knee extensor strength, thereby confirming PE’s role in modulating bone metabolism and translating into tangible improvements in functional mobility and quality of life.126
Metabolite research indicates that equol, a gut microbiota-derived metabolite of soy isoflavones, exhibits notable osteoprotective effects in preclinical studies.127 Clinical investigations further demonstrate that combined interventions with equol and resveratrol significantly improve bone turnover markers such as osteocalcin;128 however, long-term effects on BMD and motor function in postmenopausal women require further clinical validation.
In summary, PEs may indirectly enhance motor capabilities—such as balance, endurance, and muscle strength—by improving bone health, with some effects mediated through pain reduction and activity limitation alleviation. Future research should focus on elucidating how PE interventions translate into actual improvements in motor function and quality of life.
Regulation of Neurotransmitter-Motor Performance Axis by Physical Exercise: Mechanisms and Translational Implications for Perimenopausal Women
Physical exercise may influence neurofunctional and motor performance in perimenopausal women through modulation of neurotransmitter systems. Key neurotransmitters such as serotonin and dopamine are integral to motor control and coordination, and their homeostasis may be disrupted by endocrine changes during menopause, leading to complex effects on motor capabilities.
Molecular Evidence for Neurotransmitter Regulation
Preclinical studies indicate that flavonoids like quercetin possess neuroprotective properties and the capacity to modulate neurotransmitter activity. Animal experiments demonstrate that quercetin can regulate histone acetylation, thereby reducing endoplasmic reticulum stress protein expression in the hypothalamus and improving mitochondrial function.129 Additionally, research shows that quercetin (40 mg/kg) can modulate nitric oxide levels, alleviating depression-like behaviors induced by social isolation, suggesting promising therapeutic potential for stress-related emotional disturbances.130
Impact of Physical Exercise on Neurocognitive-Motor Integration in Perimenopausal Women
Clinical investigations suggest that specific PEs interventions may indirectly enhance motor performance in perimenopausal women by improving neurocognitive functions. A 40-week randomized controlled trial by Nakamura et al demonstrated that daily supplementation with 110 mg of quercetin glycosides significantly improved cognitive response speed in elderly women, including postmenopausal individuals.131 Given the close association between response speed, fall risk, and motor task performance,132 this finding implies that quercetin glycosides may enhance neuromuscular coordination, thereby improving motor safety.
Research on resveratrol also holds clinical relevance in the context of neurocognitive health among menopausal women. Two RCTs confirmed its efficacy in improving cerebrovascular reactivity and cognitive function during movement in postmenopausal women,133,134 supporting complex motor control and alleviating osteoarthritic pain.135 These combined effects may enhance exercise tolerance and adherence, establishing a foundation for maintaining or improving long-term motor capacity. Furthermore, RCTs from Asian and North American populations indicate that soy isoflavones can improve visual memory in postmenopausal women, although they do not directly influence baseline motor abilities.136,137 Nonetheless, such cognitive benefits may promote more active engagement in physical activity, with broader implications for cognitive health and participation in diverse sports activities.
Collectively, these findings suggest that PEs may support motor performance in perimenopausal women through multiple neurocognitive pathways. However, the precise mechanisms underlying these effects warrant further detailed investigation.
Conclusions and Limitations
The impact of physical exercise on postmenopausal women’s physical capacity is modulated by multiple factors, including the type, dosage, source (dietary versus supplemental), and individual metabolic differences, underscoring the importance of personalized application. Certain specific types of PEs demonstrate potential in supporting postmenopausal women’s physical performance through multiple pathways, potentially exerting beneficial effects on relevant physiological markers and adaptive processes. Current evidence indicates that: 1) PEs can regulate muscle protein metabolism, serving as a personalized adjunct to enhance physical performance, although the central role of structured exercise training remains irreplaceable; 2) their anti-inflammatory and antioxidant properties may indirectly promote improvements in physical capacity by ameliorating postmenopausal inflammatory states and oxidative stress; 3) PEs hold potential regulatory value for cardiovascular and metabolic health in this population; 4) their effects on bone metabolism appear to be influenced by multifactorial interactions, including dosage, demographic characteristics, and intervention modalities; 5) they may indirectly support neuromuscular and cognitive functions through modulation of neurotransmitter systems. These findings suggest potential non-hormonal strategies to address declines in physical capacity associated with estrogen deficiency.
Despite the promising insights, the field faces significant challenges, notably the scarcity of high-quality, large sample RCTs directly assessing the effects of PEs on specific physical performance endpoints in postmenopausal women, such as VO2max, muscle strength, and fatigue recovery. Most existing evidence derives from studies targeting general health indicators, observational research, or animal models, rather than dedicated assessments of exercise performance metrics. Furthermore, the mechanisms underlying PEs’ effects on musculoskeletal and neural tissues, as well as their interactions with exercise interventions, require further elucidation. Variability in bioavailability and metabolic responses among individuals further complicates research efforts.
Future investigations should prioritize large-scale, long-duration RCTs to systematically evaluate the effects of PEs on specific physical performance parameters, including muscle strength, endurance, balance, and post-exercise recovery. Additionally, research should aim to determine optimal dosing, timing, and long-term safety profiles of different PEs modalities, as well as to develop personalized intervention strategies based on individual characteristics. Such studies are essential for scientifically assessing the role of PEs in healthy aging and for optimizing physical performance outcomes in postmenopausal women.
Funding
This work was supported by the National Natural Science Foundation of China (Grant Numbers: 82572947, 32171136) and the Natural Science Foundation of Shanghai (Grant Number: 192R1452900). This work was also supported by the Key Laboratory of Exercise and Health Sciences (Shanghai University of Sport), Ministry of Education.
Disclosure
The author(s) report no conflicts of interest in this work.
References
1. Chen W, Chen M, Tang H, et al. Advances in diagnosis and treatment of perimenopausal syndrome. Open Life Sci. 2023;18(1):20220754. doi:10.1515/biol-2022-0754
2. Jin Z, Huang J, Cai Z. Guidelines on integrated Chinese and western medicine diagnosis and treatment of menopausal syndrome (2023 Edition). Chin J Pract Gynecol Obstet. 2023;39(8):799–808.
3. Hill K. The demography of menopause. Maturitas. 1996;23(2):113–127. doi:10.1016/0378-5122(95)00968-X
4. Bardhi O, Dubey P, Palmer BF, Clegg DJ. Oestrogens, adipose tissues and environmental exposures influence obesity and diabetes across the lifecycle. Proc Nutr Soc. 2024;83(4):263–270. doi:10.1017/S0029665124000119
5. Franco OH, Chowdhury R, Troup J, et al. Use of plant-based therapies and menopausal symptoms: a systematic review and meta-analysis. JAMA. 2016;315(23):2554. doi:10.1001/jama.2016.8012
6. Zhou Y, Su Z, Liu G, Hu S, Chang J. The potential mechanism of soy isoflavones in treating osteoporosis: focusing on bone metabolism and oxidative stress. Phytother Res. 2025;39:1645–1658. doi:10.1002/ptr.8451
7. Hurtado MD, Saadedine M, Kapoor E, Shufelt CL, Faubion SS. Weight gain in midlife women. Curr Obes Rep. 2024;13(2):352–363. doi:10.1007/s13679-024-00555-2
8. Moore IS, Crossley KM, Bo K, et al. Female athlete health domains: a supplement to the international olympic committee consensus statement on methods for recording and reporting epidemiological data on injury and illness in sport. Br J Sports Med. 2023;57(18):1164–1174. doi:10.1136/bjsports-2022-106620
9. Menopause. Available from: https://www.who.int/news-room/fact-sheets/detail/menopause.
10. Forma E, Urbańska K, Bryś M. Menopause hot flashes and molecular mechanisms modulated by food-derived nutrients. Nutrients. 2024;16(5):655. doi:10.3390/nu16050655
11. Grigolon RB, Ceolin G, Deng Y, et al. Effects of nutritional interventions on the severity of depressive and anxiety symptoms of women in the menopausal transition and menopause: a systematic review, meta-analysis, and meta-regression. Menopause. 2023;30(1):95–107. doi:10.1097/GME.0000000000002098
12. Bagheri Lankarani K, Jamalinia M, Zare F, Heydari ST, Ardekani A, Lonardo A. Liver-kidney-metabolic health, sex, and menopause impact total scores and monovessel vs. Multivessel coronary artery calcification. Adv Ther. 2025;42:1729–1744. doi:10.1007/s12325-025-03121-x
13. Lankila H, Kekäläinen T, Hietavala EM, Laakkonen EK. A mediating role of visceral adipose tissue on the association of health behaviours and metabolic inflammation in menopause: a population-based cross-sectional study. Sci Rep. 2025;15(1):1999. doi:10.1038/s41598-025-85134-8
14. Willoughby DS, Florez C, Davis J, et al. Decreased neuromuscular function and muscle quality along with increased systemic inflammation and muscle proteolysis occurring in the presence of decreased estradiol and protein intake in early to intermediate post-menopausal women. Nutrients. 2024;16(2):197. doi:10.3390/nu16020197
15. Nio AQX, Rogers S, Mynors-Wallis R, et al. The menopause alters aerobic adaptations to high-intensity interval training. Med Sci Sports Exerc. 2020;52(10):2096–2106. doi:10.1249/MSS.0000000000002372
16. Pang H, Chen S, Klyne DM, et al. Low back pain and osteoarthritis pain: a perspective of estrogen. Bone Res. 2023;11(1):42. doi:10.1038/s41413-023-00280-x
17. Baber RJ, Panay N, Fenton A; IMS Writing Group. 2016 IMS Recommendations on women’s midlife health and menopause hormone therapy. Climacteric. 2016;19(2):109–150. doi:10.3109/13697137.2015.1129166
18. Morrison MF, Kallan MJ, Ten Have T, Katz I, Tweedy K, Battistini M. Lack of efficacy of estradiol for depression in postmenopausal women: a randomized, controlled trial. Biol Psychiatry. 2004;55(4):406–412. doi:10.1016/j.biopsych.2003.08.011
19. Gulati A, Banker H, Muhammad AA, Anamika F, Jain R. Unlocking the potential: phytoestrogens and cardiovascular health. Curr Cardiol Rev. 2025;21. doi:10.2174/011573403X333952241203050033
20. Hamoda H, Panay N, Pedder H, Arya R, Savvas M. The British menopause society & women’s health concern 2020 recommendations on hormone replacement therapy in menopausal women. Post Reprod Health. 2020;26(4):181–209. doi:10.1177/2053369120957514
21. Zeng X, Li C, Liu Y, et al. HLA-B*35:01-mediated activation of emodin-specific T cells contributes to polygonum multiflorum thunb. -induced liver injury in mice. J Ethnopharmacol. 2024;334:118523. doi:10.1016/j.jep.2024.118523
22. Curiel JA, de la Bastida AR, Langa S, Peirotén Á, Landete JM. Characterization and stabilization of GluLm and its application to deglycosylate dietary flavonoids and lignans. Appl Microbiol Biotechnol. 2024;108(1):80. doi:10.1007/s00253-023-12956-9
23. Bharti R, Chopra BS, Raut S, Khatri N. Pueraria tuberosa: a review on traditional uses, pharmacology, and phytochemistry. Front Pharmacol. 2020;11:582506. doi:10.3389/fphar.2020.582506
24. China Center for Disease Control and Prevention, Institute of Nutrition and Food Safety. China Food Composition Table 2004. Peking University Medical Press; 2005.
25. Ma ZM, Hao XY, Wang DY, Wang XL, Sun YL, Li HY. Evolution and distribution of dietary patterns in China and the research progress of its correlation with health. Sci Technol Food Ind. 2023;44(10):396–405.
26. Taku K, Melby MK, Kronenberg F, Kurzer MS, Messina M. Extracted or synthesized soybean isoflavones reduce menopausal hot flash frequency and severity: systematic review and meta-analysis of randomized controlled trials. Menopause. 2012;19(7):776–790. doi:10.1097/gme.0b013e3182410159
27. Daily JW, Ko BS, Ryuk J, Liu M, Zhang W, Park S. Equol decreases hot flashes in postmenopausal women: a systematic review and meta-analysis of randomized clinical trials. J Med Food. 2019;22(2):127–139. doi:10.1089/jmf.2018.4265
28. Lee YJ, Lee C, Choi D, Lee Y, Lee SH. Effect of soy isoflavone on prostate cancer cell apoptosis through inhibition of STAT3, ERK, and AKT. Curr Issues Mol Biol. 2024;46(11):12512–12526. doi:10.3390/cimb46110743
29. Pathak N, Oliveira M. New oral selective estrogen receptor degraders redefine management of estrogen receptor-positive breast cancer. Annu Rev Med. 2025;76(1):243–255. doi:10.1146/annurev-med-052423-122001
30. Kastora SL, Pantiora E, Hong YH, et al. Safety of topical estrogen therapy during adjuvant endocrine treatment among patients with breast cancer: a meta-analysis based expert panel discussion. Cancer Treat Rev. 2025;133:102880. doi:10.1016/j.ctrv.2025.102880
31. Sukhamwang A, Inthanon S, Dejkriengkraikul P, Semangoen T, Yodkeeree S. Anti-cancer potential of isoflavone-enriched fraction from traditional Thai fermented soybean against hela cervical cancer cells. Int J Mol Sci. 2024;25(17):9277. doi:10.3390/ijms25179277
32. Shpigel J, Luciano EF, Ukandu B, Sauane M, de la Parra C. Soy isoflavone genistein enhances tamoxifen sensitivity in breast cancer via microRNA and glucose metabolism modulation. Int J Mol Sci. 2025;26(2):733. doi:10.3390/ijms26020733
33. Wang Y, Li Z, Wu D, et al. Biotransformation analysis of daidzin in vitro based on fecal bacteria and probiotics. J Pharm Biomed Anal. 2025;255:116623. doi:10.1016/j.jpba.2024.116623
34. Sun C. Nutrition and Food Hygiene.
35. Zhang JM, Sun T, Bing SY, Lin Q. Research progress on feeding value of flaxseed and its processed products and their application in animal production. Chin J Animal Nutr. 2024;36(1):98–106.
36. Li J, Ma X, Luo L, Tang D, Zhang L. The what and who of dietary lignans in human health: special attention to estrogen effects and safety evaluation. J Agric Food Chem. 2023;71(44):16419–16434. doi:10.1021/acs.jafc.3c02680
37. Pellegrini N, Valtueña S, Ardigò D, et al. Intake of the plant lignans matairesinol, secoisolariciresinol, pinoresinol, and lariciresinol in relation to vascular inflammation and endothelial dysfunction in middle age-elderly men and post-menopausal women living in northern Italy. Nutr Metab Cardiovasc Dis. 2010;20(1):64–71. doi:10.1016/j.numecd.2009.02.003
38. Witkowska AM, Waśkiewicz A, Zujko ME, et al. Are total and individual dietary lignans related to cardiovascular disease and its risk factors in postmenopausal women? A nationwide study. Nutrients. 2018;10(7):865. doi:10.3390/nu10070865
39. Błaszczuk A, Barańska A, Kanadys W, et al. Role of phytoestrogen-rich bioactive substances (linum usitatissimum L. glycine max L. trifolium pratense L.) in cardiovascular disease prevention in postmenopausal women: a systematic review and meta-analysis. Nutrients. 2022;14(12):2467. doi:10.3390/nu14122467
40. Oboulbiga EB, Douamba Z, Compaoré-Sérémé D, et al. Physicochemical, potential nutritional, antioxidant and health properties of sesame seed oil: a review. Front Nutr. 2023;10:1127926. doi:10.3389/fnut.2023.1127926
41. Wang S, Hu Y, Liu B, Li Y, Wang M, Sun Q. Lignan intake and type 2 diabetes incidence among US men and women. JAMA Network Open. 2024;7(8):e2426367. doi:10.1001/jamanetworkopen.2024.26367
42. Tetens I, Turrini A, Tapanainen H, et al. Dietary intake and main sources of plant lignans in five european countries. Food Nutr Res. 2013;57(1):19805. doi:10.3402/fnr.v57i0.19805
43. Feng CC, Zhang W, Cai ZZ, Li Y, Wang Y, Zhang N. Research progress in biotransformation of plant lignans. Food Ferment Ind. 2021;47(11):295–302.
44. Hu Q, Ma X, Cai T, Li Y. Flavonoid intake, inflammation, and atherosclerotic cardiovascular disease risk in U.S. adults: a cross-sectional study. Nutr Metab. 2025;22(1):24. doi:10.1186/s12986-025-00913-4
45. Yan X, Liu HQ, Zhou YQ, Ren ZH. Physiological activities and research advance in synthesis of flavonoids. Chin J Org Chem. 2008;28(9):1534–1544.
46. Seukep AJ, Mbuntcha HG, Matieta VY, Kuete V, Guo MQ. HPLC-ESI-QTOF-MS profiling of antibacterial bioactive solvent fractions of senna alata (L.) roxb (fabaceae) leaves, and in silico prediction of pharmacokinetic, drug-likeness, and toxicity of major phyto-components. J Ethnopharmacol. 2025;344:119492. doi:10.1016/j.jep.2025.119492
47. Khajehlandi M, Bolboli L. The role of quercetin and exercise in modulating apoptosis and cardiomyopathy via PI3K/AKT/FOXO3 pathways in diabetic obese rats. Cell J. 2025;26(9):559–568. doi:10.22074/cellj.2024.2042044.1682
48. Wu ZY, Qiu KY, Gai YJ, Wu JH, Zhou BX, Shi QF. Quercetin: a natural ally in combating breast cancer. IJN. 2025;20:9155–9177. doi:10.2147/IJN.S518174
49. Matta L, Breves C, Fonte Boa L, et al. Quercetin improves white adipose tissue redox homeostasis in ovariectomized rats. J Endocrinol. 2023;259(2):e230166. doi:10.1530/JOE-23-0166
50. Wang L, Lee IM, Zhang SM, Blumberg JB, Buring JE, Sesso HD. Dietary intake of selected flavonols, flavones, and flavonoid-rich foods and risk of cancer in middle-aged and older women. Am J Clin Nutr. 2009;89(3):905–912. doi:10.3945/ajcn.2008.26913
51. Curtis PJ, Potter J, Kroon PA, et al. Vascular function and atherosclerosis progression after 1 y of flavonoid intake in statin-treated postmenopausal women with type 2 diabetes: a double-blind randomized controlled trial. Am J Clin Nutr. 2013;97(5):936–942. doi:10.3945/ajcn.112.043745
52. Jesus RLC, Silva ILP, Araújo FA, et al. 7-hydroxycoumarin induces vasorelaxation in animals with essential hypertension: focus on potassium channels and intracellular Ca2+ mobilization. Molecules. 2022;27(21):7324. doi:10.3390/molecules27217324
53. Zhu S, Zhu W, Zhao K, et al. Discovery of a novel hybrid coumarin-hydroxamate conjugate targeting the HDAC1-Sp1-FOSL2 signaling axis for breast cancer therapy. Cell Commun Signal. 2024;22(1):361. doi:10.1186/s12964-024-01733-4
54. Niranjan V, Jayaprasad S, Uttarkar A, Kusanur R, Kumar J. Design of novel coumarin derivatives as NUDT5 antagonists that act by restricting ATP synthesis in breast cancer cells. Molecules. 2022;28(1):89. doi:10.3390/molecules28010089
55. Lopes PKF, Engel DF, Bertolini NO, et al. Behavioral, neuroplasticity and metabolic effects of 7,8-dihydroxy-4-methylcoumarin associated with physical activity in mice. Metab Brain Dis. 2021;36(8):2425–2436. doi:10.1007/s11011-021-00849-7
56. Cottart CH, Nivet-Antoine V, Laguillier-Morizot C, Beaudeux JL. Resveratrol bioavailability and toxicity in humans. Mol Nutr Food Res. 2010;54(1):7–16. doi:10.1002/mnfr.200900437
57. Ngo TH, Lee YJ, Choi H, Song KS, Lee KJ, Nam JW. Evaluating the anticancer potential of polygonum multiflorum root-derived stilbenes against H2452 malignant pleural mesothelioma cells. Fitoterapia. 2024;177:106135. doi:10.1016/j.fitote.2024.106135
58. Iqbal N, Javed M, Khoder RM, et al. Innovative CO2-NBs-assisted ultrasonication for the phytochemical extraction of peanut (arachis hypoga) shells: synthesis and characterization of CO2-nanobubbles. Ultrason Sonochem. 2025;112:107198. doi:10.1016/j.ultsonch.2024.107198
59. Liu D, Li LF, Zhai H, et al. Resveratrol inhibits african swine fever virus replication by exerting antiviral and antioxidative stress activities via the Nrf2 signaling pathway. Emerg Microbes Infect. 2025;14:2469662. doi:10.1080/22221751.2025.2469662
60. Balasubramani SP, Rahman MA, Basha SM. Synergistic action of stilbenes in muscadine grape berry extract shows better cytotoxic potential against cancer cells than resveratrol alone. Biomedicines. 2019;7(4):96. doi:10.3390/biomedicines7040096
61. Oxfeldt M, Dalgaard LB, Risikesan J, Johansen FT, Hansen M. Influence of fermented red clover extract on skeletal muscle in early postmenopausal women: a double-blinded cross-over study. Nutrients. 2020;12(11):3587. doi:10.3390/nu12113587
62. Huang CC, Lee MC, Ho CS, Hsu YJ, Ho CC, Kan NW. Protective and recovery effects of resveratrol supplementation on exercise performance and muscle damage following acute plyometric exercise. Nutrients. 2021;13(9):3217. doi:10.3390/nu13093217
63. Tang HX, Zeng M, Liao YD. Research advances in the estrogen receptors in estrogen non-target organ tumors. Chin J Cancer Prev Treat. 2012;19(3):235–240.
64. Seo H, Seo H, Lee SH, Park Y. Receptor mediated biological activities of phytoestrogens. Int J Biol Macromol. 2024;278(Pt 2):134320. doi:10.1016/j.ijbiomac.2024.134320
65. Besong C, Philippeaux S, Bham A, et al. Managing menopause: the evolving role of estrogens, selective serotonin reuptake inhibitors, and phytoestrogens in balancing hormonal fluctuations. Cureus. 2024;16(9):e70440. doi:10.7759/cureus.70440
66. Kumari N, Kumari R, Dua A, et al. From gut to hormones: unraveling the role of gut microbiota in (phyto)estrogen modulation in health and disease. Mol Nutr Food Res. 2024;68(6):e2300688. doi:10.1002/mnfr.202300688
67. Setchell KD, Brown NM, Desai P, et al. Bioavailability of pure isoflavones in healthy humans and analysis of commercial soy isoflavone supplements. J Nutr. 2001;131(4 Suppl):1362S–75S. doi:10.1093/jn/131.4.1362S
68. Nosal BM, Thornton SN, Melnik AV, et al. Blackcurrant anthocyanins attenuate estrogen -deficiency-induced bone loss through modulating microbial-derived short-chain carboxylic acids and phytoestrogen metabolites in peri- and early postmenopausal women. Metabolites. 2024;14(10):541. doi:10.3390/metabo14100541
69. Zhou Y, Lee AS. Mechanism for the suppression of the mammalian stress response by genistein, an anticancer phytoestrogen from soy. J Natl Cancer Inst. 1998;90(5):381–388. doi:10.1093/jnci/90.5.381
70. Yin L, Chen X, Li N, et al. Puerarin ameliorates skeletal muscle wasting and fiber type transformation in STZ-induced type 1 diabetic rats. Biomed Pharmacother. 2021;133:110977. doi:10.1016/j.biopha.2020.110977
71. Ahmed SA, Parama D, Daimari E, et al. Rationalizing the therapeutic potential of apigenin against cancer. Life Sci. 2021;267:118814. doi:10.1016/j.lfs.2020.118814
72. Daneshvar S, Zamanian MY, Ivraghi MS, et al. A comprehensive view on the apigenin impact on colorectal cancer: focusing on cellular and molecular mechanisms. Food Sci Nutr. 2023;11(11):6789–6801. doi:10.1002/fsn3.3645
73. Choi WH, Son HJ, Jang YJ, Ahn J, Jung CH, Ha TY. Apigenin ameliorates the obesity-induced skeletal muscle atrophy by attenuating mitochondrial dysfunction in the muscle of obese mice. Mol Nutr Food Res. 2017;61(12). doi:10.1002/mnfr.201700218
74. Wang K, Zeng H, Yang H. Daidzein improves muscle atrophy caused by lovastatin by regulating the AMPK/FOXO3a axis. Chin Med. 2024;19(1):180. doi:10.1186/s13020-024-01034-5
75. Hirasaka K, Saito S, Yamaguchi S, et al. Dietary supplementation with isoflavones prevents muscle wasting in tumor-bearing mice. J Nutr Sci Vitaminol. 2016;62(3):178–184. doi:10.3177/jnsv.62.178
76. Jenks BH, Iwashita S, Nakagawa Y, et al. A pilot study on the effects of S -equol compared to soy isoflavones on menopausal hot flash frequency. J Women’s Health. 2012;21(6):674–682. doi:10.1089/jwh.2011.3153
77. Ryan-Borchers TA, Park JS, Chew BP, McGuire MK, Fournier LR, Beerman KA. Soy isoflavones modulate immune function in healthy postmenopausal women. Am J Clin Nutr. 2006;83(5):1118–1125. doi:10.1093/ajcn/83.5.1118
78. Orsatti FL, Maestá N, De Oliveira EP, et al. Adding soy protein to milk enhances the effect of resistance training on muscle strength in postmenopausal women. J Dietary Suppl. 2018;15(2):140–152. doi:10.1080/19390211.2017.1330794
79. Fontvieille A, Dionne IJ, Riesco E. Long-term exercise training and soy isoflavones to improve quality of life and climacteric symptoms. Climacteric. 2017;20(3):233–239. doi:10.1080/13697137.2017.1294153
80. Choquette S, Dion T, Brochu M, Dionne IJ. Soy isoflavones and exercise to improve physical capacity in postmenopausal women. Climacteric. 2012;16(1):70–77. doi:10.3109/13697137.2011.643515
81. Xin Y, Xu J, Li N, Yang LY, Zhu HT, Zhang YJ. New terpenoids and lignans from phyllanthus acidus fruits with antioxidant activity. Foods. 2025;14(3):452. doi:10.3390/foods14030452
82. Graf BL, Zhang L, Corradini MG, et al. Physicochemical differences between malanga (xanthosoma sagittifolium) and potato (solanum tuberosum) tubers are associated with differential effects on the gut microbiome. J Funct Foods. 2018;45:268–276. doi:10.1016/j.jff.2018.04.032
83. Menzies KJ, Singh K, Saleem A, Hood DA. Sirtuin 1-mediated effects of exercise and resveratrol on mitochondrial biogenesis. J Biol Chem. 2013;288(10):6968–6979. doi:10.1074/jbc.M112.431155
84. Cao W, Zheng B, Zeng X, He H, Chen L. Stilbene, as phyto-oestrogens, can construct resistant starch through noncovalent interactions with starch: a structural correlation study. Food Hydrocoll. 2024;148:109438. doi:10.1016/j.foodhyd.2023.109438
85. Su LY, Huang WC, Kan NW, Tung TH, Huynh LBP, Huang SY. Effects of resveratrol on muscle inflammation, energy utilisation, and exercise performance in an eccentric contraction exercise mouse model. Nutrients. 2023;15(1):249. doi:10.3390/nu15010249
86. Yang M, Zhang T. Genistein inhibits the release of pro-inflammatory substances from macrophages by suppressing potassium loss- and ROS-mediated caspase-1/gasdermin D pathway activation and pyroptotic cell lysis. Iran J Basic Med Sci. 2024;27(12):1506–1514. doi:10.22038/ijbms.2024.77887.16854
87. Kim HI, Kim MK, Lee I, Yun J, Kim EH, Seo SK. Efficacy and safety of a standardized soy and hop extract on menopausal symptoms: a 12-week, multicenter, randomized, double-blind, placebo-controlled clinical trial. J Altern Complement Med. 2021;27(11):959–967. doi:10.1089/acm.2021.0027
88. Llaneza P, González C, Fernández-Iñarrea J, Alonso A, Díaz F, Pérez-López FR. Soy isoflavones improve insulin sensitivity without changing serum leptin among postmenopausal women. Climacteric. 2012;15(6):611–620. doi:10.3109/13697137.2011.631062
89. O’Leary MF, Jackman SR, Bowtell JL. Shatavari supplementation in postmenopausal women alters the skeletal muscle proteome and pathways involved in training adaptation. Eur J Nutr. 2024;63(3):869–879. doi:10.1007/s00394-023-03310-w
90. Riesco E, Choquette S, Audet M, Lebon J, Tessier D, Dionne IJ. Effect of exercise training combined with phytoestrogens on adipokines and C-reactive protein in postmenopausal women: a randomized trial. Metabolism. 2012;61(2):273–280. doi:10.1016/j.metabol.2011.06.025
91. Lee D, Kim YM, Kim HW, et al. Schisandrin C affects glucose-stimulated insulin secretion in pancreatic β-cells and glucose uptake in skeletal muscle cells. Molecules. 2021;26(21):6509. doi:10.3390/molecules26216509
92. Das D, Sarkar S, Dihingia A, et al. A popular fermented soybean food of northeast India exerted promising antihyperglycemic potential via stimulating PI3K/AKT/AMPK/GLUT4 signaling pathways and regulating muscle glucose metabolism in type 2 diabetes. J Food Biochem. 2022;46(12):e14385. doi:10.1111/jfbc.14385
93. Gan M, Chen X, Chen Z, et al. Genistein alleviates high-fat diet-induced obesity by inhibiting the process of gluconeogenesis in mice. Nutrients. 2022;14(8):1551. doi:10.3390/nu14081551
94. Tse MCL, Pang BPS, Bi X, et al. Estrogen regulates mitochondrial activity through inducing brain-derived neurotrophic factor expression in skeletal muscle. J Cell Physiol. 2025;240(1):e31483. doi:10.1002/jcp.31483
95. Sharma G, Hu C, Staquicini DI, et al. Preclinical efficacy of the GPER-selective agonist G-1 in mouse models of obesity and diabetes. Sci Transl Med. 2020;12(528):eaau5956. doi:10.1126/scitranslmed.aau5956
96. Heine PA, Taylor JA, Iwamoto GA, Lubahn DB, Cooke PS. Increased adipose tissue in male and female estrogen receptor-alpha knockout mice. Proc Natl Acad Sci U S A. 2000;97(23):12729–12734. doi:10.1073/pnas.97.23.12729
97. Gallagher CJ, Langefeld CD, Gordon CJ, et al. Association of the estrogen receptor-alpha gene with the metabolic syndrome and its component traits in african-American families: the insulin resistance atherosclerosis family study. Diabetes. 2007;56(8):2135–2141. doi:10.2337/db06-1017
98. Qin H, Song Z, Shaukat H, Zheng W. Genistein regulates lipid metabolism via estrogen receptor β and its downstream signal akt/mTOR in HepG2 cells. Nutrients. 2021;13(11):4015. doi:10.3390/nu13114015
99. Kim M, Im S, Cho YK, et al. Anti-obesity effects of soybean embryo extract and enzymatically-modified isoquercitrin. Biomolecules. 2020;10(10):1394. doi:10.3390/biom10101394
100. Chu Z, Hu Z, Luo Y, Zhou Y, Yang F, Luo F. Targeting gut-liver axis by dietary lignans ameliorate obesity: evidences and mechanisms. Crit Rev Food Sci Nutr. 2025;65(2):243–264. doi:10.1080/10408398.2023.2272269
101. Osuna-Prieto FJ, Martinez-Tellez B, Segura-Carretero A, Ruiz JR. Activation of brown adipose tissue and promotion of white adipose tissue browning by plant-based dietary components in rodents: a systematic review. Adv Nutr. 2021;12(6):2147–2156. doi:10.1093/advances/nmab084
102. Kim SN, Ahn SY, Song HD, et al. Antiobesity effects of coumestrol through expansion and activation of brown adipose tissue metabolism. J Nutr Biochem. 2020;76:108300. doi:10.1016/j.jnutbio.2019.108300
103. Yang S, Zeng Q, Huang X, Liang Z, Hu H. Effect of isoflavones on blood lipid alterations in postmenopausal females: a systematic review and meta-analysis of randomized trials. Adv Nutr. 2023;14(6):1633–1643. doi:10.1016/j.advnut.2023.09.008
104. Zhang YB, Chen WH, Guo JJ, et al. Soy isoflavone supplementation could reduce body weight and improve glucose metabolism in non-asian postmenopausal women—a meta-analysis. Nutrition. 2013;29(1):8–14. doi:10.1016/j.nut.2012.03.019
105. Llaneza P, Gonzalez C, Fernandez-Iñarrea J, et al. Soy isoflavones, Mediterranean diet, and physical exercise in postmenopausal women with insulin resistance. Menopause. 2010;17(2):372–378. doi:10.1097/gme.0b013e3181ba56fa
106. Llaneza P, González C, Fernandez-Iñarrea J, et al. Soy isoflavones, diet and physical exercise modify serum cytokines in healthy obese postmenopausal women. Phytomedicine. 2011;18(4):245–250. doi:10.1016/j.phymed.2010.07.011
107. Dechichi JGC, Mariano IM, Giolo JS, et al. Isoflavone supplementation does not potentiate the effect of combined exercise training on resting and ambulatory blood pressure in non-obese postmenopausal women: a randomized double-blind controlled trial-a pilot study. Nutrients. 2020;12(11):3495. doi:10.3390/nu12113495
108. Kou T, Wang Q, Cai J, et al. Effect of soybean protein on blood pressure in postmenopausal women: a meta-analysis of randomized controlled trials. Food Funct. 2017;8(8):2663–2671. doi:10.1039/C6FO01845A
109. Ni X, Wu B, Li S, et al. Equol exerts a protective effect on postmenopausal osteoporosis by upregulating OPG/RANKL pathway. Phytomedicine. 2023;108:154509. doi:10.1016/j.phymed.2022.154509
110. Li H, Wang C, Jin Y, et al. Anti-postmenopausal osteoporosis effects of isopsoralen: a bioinformatics-integrated experimental study. Phytother Res. 2023;37(1):231–251. doi:10.1002/ptr.7609
111. Ban Y, Wang Y, Qiao L, et al. Total lignans from Vitex negundo seeds attenuate osteoarthritis and their main component vitedoin A alleviates osteoclast differentiation by suppressing ERK/NFATc1 signaling. Phytother Res. 2023;37(4):1422–1434. doi:10.1002/ptr.7750
112. Mannino F, Imbesi C, Irrera N, Pallio G, Squadrito F, Bitto A. Insights into the antiosteoporotic mechanism of the soy-derived isoflavone genistein: modulation of the wnt/beta-catenin signaling. Biofactors. 2024;50(2):347–359. doi:10.1002/biof.2008
113. Fan Z, Gao J, Chen Y, et al. Psoralen remodels the articular cartilage microenvironment by pharmacologically regulating the Nrf2 pathway in osteoarthritis treatment. Int Immunopharmacol. 2025;149:114221. doi:10.1016/j.intimp.2025.114221
114. Zhang X, Liu Y, Xu Q, et al. The effect of soy isoflavone combined with calcium on bone mineral density in perimenopausal Chinese women: a 6-month randomised double-blind placebo-controlled study. Int J Food Sci Nutr. 2020;71(4):473–481. doi:10.1080/09637486.2019.1673703
115. Sharma K, Awasthi P, Prakash R, et al. Maintenance of increased bone mass after PTH withdrawal by sequential medicarpin treatment via augmentation of cAMP-PKA pathway. J Cell Biochem. 2022;123(11):1762–1779. doi:10.1002/jcb.30313
116. Ma DF, Qin LQ, Wang PY, Katoh R. Soy isoflavone intake inhibits bone resorption and stimulates bone formation in menopausal women: meta-analysis of randomized controlled trials. Eur J Clin Nutr. 2008;62(2):155–161. doi:10.1038/sj.ejcn.1602748
117. Morabito N, Crisafulli A, Vergara C, et al. Effects of genistein and hormone-replacement therapy on bone loss in early postmenopausal women: a randomized double-blind placebo-controlled study. J Bone Miner Res. 2002;17(10):1904–1912. doi:10.1359/jbmr.2002.17.10.1904
118. Levis S. Soy isoflavones in the prevention of menopausal bone loss and menopausal symptoms: a randomized, double-blind trial. Arch Intern Med. 2011;171(15):1363. doi:10.1001/archinternmed.2011.330
119. Wong WW, Lewis RD, Steinberg FM, et al. Soy isoflavone supplementation and bone mineral density in menopausal women: a 2-y multicenter clinical trial. Am J Clin Nutr. 2009;90(5):1433–1439. doi:10.3945/ajcn.2009.28001
120. Alekel DL, Van Loan MD, Koehler KJ, et al. The soy isoflavones for reducing bone loss (SIRBL) study: a 3-y randomized controlled trial in postmenopausal women. Am J Clin Nutr. 2010;91(1):218–230. doi:10.3945/ajcn.2009.28306
121. Vupadhyayula PM, Gallagher JC, Templin T, Logsdon SM, Smith LM. Effects of soy protein isolate on bone mineral density and physical performance indices in postmenopausal women-a 2-year randomized, double-blind, placebo-controlled trial. Menopause. 2009;16(2):320–328. doi:10.1097/gme.0b013e3181844893
122. Chen YM, Ho SC, Lam SSH, Ho SSS, Woo JLF. Soy isoflavones have a favorable effect on bone loss in Chinese postmenopausal women with lower bone mass: a double-blind, randomized, controlled trial. J Clin Endocrinol Metab. 2003;88(10):4740–4747. doi:10.1210/jc.2003-030290
123. Tai TY, Tsai KS, Tu ST, et al. The effect of soy isoflavone on bone mineral density in postmenopausal taiwanese women with bone loss: a 2-year randomized double-blind placebo-controlled study. Osteoporos Int. 2012;23(5):1571–1580. doi:10.1007/s00198-011-1750-7
124. Brink E, Coxam V, Robins S, Wahala K, Cassidy A, Branca F. Long-term consumption of isoflavone-enriched foods does not affect bone mineral density, bone metabolism, or hormonal status in early postmenopausal women: a randomized, double-blind, placebo controlled study. Am J Clin Nutr. 2008;87(3):761–770. doi:10.1093/ajcn/87.3.761
125. Chilibeck PD, Vatanparast H, Pierson R, et al. Effect of exercise training combined with isoflavone supplementation on bone and lipids in postmenopausal women: a randomized clinical trial. J Bone Miner Res. 2013;28(4):780–793. doi:10.1002/jbmr.1815
126. Cornish SM, Chilibeck PD, Paus-Jennsen L, et al. A randomized controlled trial of the effects of flaxseed lignan complex on metabolic syndrome composite score and bone mineral in older adults. Appl Physiol Nutr Metab. 2009;34(2):89–98. doi:10.1139/H08-142
127. Tousen Y, Matsumoto Y, Matsumoto C, et al. The combined effects of soya isoflavones and resistant starch on equol production and trabecular bone loss in ovariectomised mice. Br J Nutr. 2016;116(2):247–257. doi:10.1017/S0007114516001537
128. Corbi G, Nobile V, Conti V, et al. Equol and resveratrol improve bone turnover biomarkers in postmenopausal women: a clinical trial. Int J Mol Sci. 2023;24(15):12063. doi:10.3390/ijms241512063
129. Wang D, Yu Z, Yao R, et al. Quercetin alleviates depressive-like behavior by modulating acetyl-H3K9 mediated ferroptosis pathway in hypothalamus of perimenopausal depression rat model. Biomed Pharmacother. 2024;179:117369. doi:10.1016/j.biopha.2024.117369
130. Tavakol F, Amini-Khoei H, Sureda A, Zarean E, Lorigooini Z. Exploring the anti-depressant effects and nitric oxide modulation of quercetin: a preclinical study in socially isolated mice. World J Biol Psychiatry. 2024;25(10):592–603. doi:10.1080/15622975.2024.2424162
131. Nakamura Y, Watanabe H, Tanaka A, Nishihira J, Murayama N. Effect of quercetin glycosides on cognitive functions and cerebral blood flow: a randomized, double-blind, and placebo-controlled study. Eur Rev Med Pharmacol Sci. 2022;26(23):8700–8712. doi:10.26355/eurrev_202212_30541
132. Nunes J, Armada M, Pereira JL, Ribeiro NF, Carvalho Ó, Santos CP. Biomechanical strategies for mitigating unexpected slips: a review. J Biomech. 2024;173:112235. doi:10.1016/j.jbiomech.2024.112235
133. Zaw JJT, Howe PRC, Wong RHX. Long-term effects of resveratrol on cognition, cerebrovascular function and cardio-metabolic markers in postmenopausal women: a 24-month randomised, double-blind, placebo-controlled, crossover study. Clin Nutr. 2021;40(3):820–829. doi:10.1016/j.clnu.2020.08.025
134. Evans H, Howe P, Wong R. Effects of resveratrol on cognitive performance, mood and cerebrovascular function in post-menopausal women; a 14-week randomised placebo-controlled intervention trial. Nutrients. 2017;9(1):27. doi:10.3390/nu9010027
135. Thaung Zaw JJ, Howe PRC, Wong RHX. Long-term resveratrol supplementation improves pain perception, menopausal symptoms, and overall well-being in postmenopausal women: findings from a 24-month randomized, controlled, crossover trial. Menopause. 2020;28(1):40–49. doi:10.1097/GME.0000000000001643
136. Ho SC, Chan ASY, Ho YP, et al. Effects of soy isoflavone supplementation on cognitive function in Chinese postmenopausal women: a double-blind, randomized, controlled trial. Menopause. 2007;14(3):489–499. doi:10.1097/gme.0b013e31802c4f4f
137. Henderson VW, St. John JA, Hodis HN, et al. Long-term soy isoflavone supplementation and cognition in women: a randomized, controlled trial. Neurology. 2012;78(23):1841–1848. doi:10.1212/WNL.0b013e318258f822
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.