")
Back to Journals » Breast Cancer: Targets and Therapy » Volume 15
The Emerging Roles of Exosomal miRNAs in Breast Cancer Progression and Potential Clinical Applications
Authors Li J, He D , Bi Y , Liu S
Received 13 September 2023
Accepted for publication 8 November 2023
Published 16 November 2023 Volume 2023:15 Pages 825—840
DOI https://doi.org/10.2147/BCTT.S432750
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Harikrishna Nakshatri
Jie Li,1,* Dejiao He,2,* Yajun Bi,3,* Shengxuan Liu4
1Department of Thyroid and Breast Surgery, Shenzhen Qianhai Shekou Free Trade Zone Hospital, Shenzhen, 518067, People’s Republic of China; 2Department of Nephrology, Renmin Hospital of Wuhan University, Wuhan, 430060, People’s Republic of China; 3Department of Pediatrics, Dalian Municipal Women and Children’s Medical Center (Group), Dalian Medical University, Dalian, Liaoning Province, 116011, People’s Republic of China; 4Department of Pediatrics, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei Province, 430030, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Shengxuan Liu, Department of Pediatrics, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei Province, 430030, People’s Republic of China, Tel +86 133 8756 1016, Email [email protected]
Abstract: Breast cancer remains the leading malignancy in terms of morbidity and mortality today. The tumor microenvironment of breast cancer includes multiple cell types, secreted proteins, and signaling components such as exosomes. Among these, exosomes have a lipid bilayer structure. Exosomes can reflect the biological traits of the parent cell and carry a variety of biologically active components, including proteins, lipids, small molecules, and non-coding RNAs, which include miRNA, lncRNA, and circRNA. MiRNAs are a group of non-coding RNAs of approximately 20– 23 nucleotides in length encoded by the genome, triggering silencing and functional repression of target genes. MiRNAs have been shown to play a significant role in the development of cancer owing to their role in the prognosis, pathogenesis, diagnosis, and treatment of cancer. MiRNAs in exosomes can serve as effective mediators of information transfer from parental cells to recipient cells and trigger changes in biological traits such as proliferation, invasion, migration, and drug resistance. These changes can profoundly alter the progression of breast cancer. Therefore, here, we systematically summarize the association of exosomal miRNAs on breast cancer progression, diagnosis, and treatment in the hope of providing novel strategies and directions for subsequent breast cancer treatment.
Keywords: breast cancer, tumor microenvironment, exosome, miRNAs, detection, therapy
Introduction
Breast cancer (BC) is a highly prevalent female cancer type with high heterogeneity.1 Over the course of 19 years, 30,000 fresh instances of cancer have been identified globally, resulting in almost 100,000 fatalities.2 In addition, BC cancer in women has surpassed lung cancer as the most common cancer worldwide. Approximately 36,850 women lost their lives due to this disease, which made up 3.11% of all cancer occurrences. In addition, BC deaths are expected to rise by more than 50% from 685,000 in 2020 to over 1 million in 2040.3 The diagnosis of BC primarily relies on radiological, hematological, and clinical evaluation, supplemented by classical biopsy confirmation. The advent of new treatments also holds great promise, such as platinum, poly(ADP-ribose) polymerase (PARP) inhibitors, and antibody-drug conjugates, even cell-based therapy.4 Despite huge advancements in diagnosis and treatment, BC patients still face core problems such as high incidence, younger age at diagnosis, high metastasis, and high recurrence. The continued exploration of molecular mechanisms will lay the foundation for precise diagnosis and therapy of BC.
Exosomes are naturally occurring extracellular vesicles of phospholipid bilayers that are secreted by almost all cells and are approximately 40–160 nm in diameter.5 Exosomes are mainly formed through early endosomes, late endosomes, multiple vesicular bodies (MVBs), fusion with lysosomes, and release. Exosomes have a large number of characteristic membrane proteins, such as CD63, CD9, CD81, and major histocompatibility complex (MHC) molecules, and exosomes contain profusely biologically active components, including proteins, lipids, small molecule metabolites, mRNA, non-coding RNAs (ncRNAs) and other nucleic acid molecules.6 Exosomes themselves are highly heterogeneous, therefore the active components, species, and abundance, with spatiotemporal specificity, will have a profound impact on the functional impact of recipient cells. In addition, in the tumor microenvironment (TME), exosomes are important mediators of various cellular interactions7,8 (Figure 1). Exosomes can act as messengers between tumor cells and non-tumor cells, such as stromal cells and immune cells, thus influencing tumor evolution.
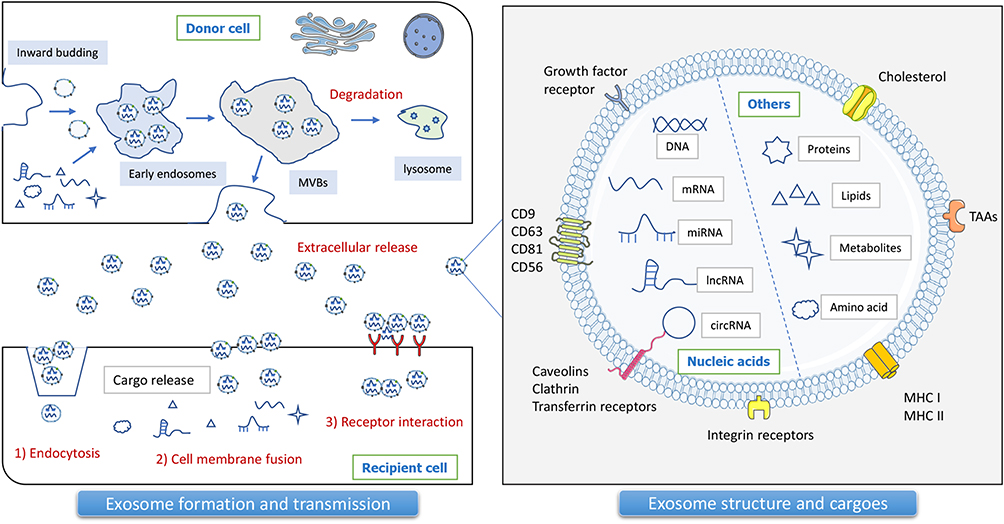 |
Figure 1 Exosome biogenesis, structure, and cargoes. Exosomes are naturally occurring membrane molecules through a complex sequence of biosynthetic processes, including early endosomes, late endosomes, multivesicular bodies (MVBs), and extracellular space release. Specific exosome markers are closely related to the parent cell and the sorting process. Tumor exosome biomarkers mainly include tetraspanins (CD9, CD63, CD81), MVB biogenesis proteins (Alix), antigen-presenting molecules (MHC-I, HSP70), tumor-associated antigens (TAAs), and others. Exosomes encapsulate multiple bioactive substances, including proteins, lipids, metabolites, DNA, and mRNA, as well as ncRNAs, especially miRNAs, lncRNAs, and circRNAs. Exosomes can transfer cargoes from donor cells to recipient cells by means of endocytosis, cell membrane fusion, and receptor interaction. |
NcRNAs, represented by miRNAs, lncRNAs, and circRNAs, are important players in molecular mechanisms of physiology and pathology by regulating gene expressions at transcriptional and post-transcriptional levels.9 Multi-omics-based sequencing tools and bioinformatics analysis are currently effective tools for mining potential tumor markers.10 Multi-omics can adequately screen for exosome-loaded ncRNA species and abundance, which are required for cellular interactions in the tumor microenvironment and during tumor evolution. Notably, miRNAs, consisting of 20–25 nucleotides, are ubiquitous ncRNAs in eukaryotes and are located in the genome. MiRNAs are very popular epigenetic factors that drive gene expression.11 Studies have shown that miRNAs are involved in almost the entire process of BC, including tumor initiation, invasion and metastasis, and immune system regulation.
Exosomal miRNA is a mapping of parental cells, and can profoundly reflect the biological status of parental cells. Mechanistically, exosome-encapsulated miRNAs have been verified to be delivered from donor cells to recipient cells through receptor-mediated endocytosis, and exert the potentially silencing function of miRNAs on downstream genes, thereby exerting anti-tumor effects.12 In addition, exosomal miRNAs can also affect the extracellular matrix (ECM), angiogenesis, and immune cell recruitment and polarization through delivery to other cell types in the TME. At present, there are also many breakthroughs in exosome-related BC biology. Exosomes from BC patient cells and serum drive non-tumorigenic epithelial cells to encourage tumor formation in a Dicer-dependent manner.13 Dysregulation of miRNA expression profile levels is a characteristic of BC cells and is reflected in the miRNA cargo of tumor-derived exosomes. For example, Scognamiglio et al reported that TNBC-derived exosomes, harboring miR-185-5p, miR-652-5p, and miR-1246, synergistically promoted fibroblasts to change to cancer-associated fibroblasts (CAFs) subtypes and enhanced their migratory function for promoting BC.14 Exosome-carried miR-181b-5p endowed senescence-mediated doxorubicin resistance by inhibiting BCLAF1 in BC progression.15 BC-exosome miR-155 could inhibit adipogenesis in preadipocytes and promote browning of white adipose tissue by targeting UBQLN1 in adipocytes, featuring cancer cachexia-related fat loss.16 In addition, Kim et al showed that RAS activation in BC cells triggered osteolytic bone metastasis by exciting exosome-mediated osteoclast production of miRNAs, including miR-494-3p, to osteoblasts.17 This feature makes exosomal miRNAs derived from tumor tissue or other body fluids high-value indicators for tumor differential diagnosis, treatment monitoring, and prognosis evaluation.18
Given the increasingly prominent roles of exosomal miRNAs in BC, here we highlight the novel roles and mechanisms of exosomal miRNAs, including BC progression regulation, as well as their roles in clinical BC diagnosis and treatment. This field will provide novel exosomal miRNA-based strategies for combating BC.
Exosome-Derived miRNAs in Remodeling BC Progression
Exosome-Derived miRNAs from BC Cells
Interaction Between BC Cells
BC-derived exosomes can function as paracrine and autocrine components to trigger the functional changes in BC cells. MiR-155, a common oncogenic signal in tumor exosomes, can lead to BC-associated cachexia by reprogramming systemic energy metabolism, regulating transcription of key proteins, and mediating chemoresistance.19,20 MiR-9 and miR-155 are overexpressed in highly metastatic triple-negative breast cancer (TNBC) cells as well as their exosomes.21 Bioinformatics studies and luciferase assays showed that miR-9 and miR-155 could specifically inhibit PTEN and DUSP14 respectively, and that their exosome-encapsulated forms could be transferred into MCF-7 cells, thus enhancing the invasive properties of the recipient cells.21
Exosomal miR-760, released from CCL18-stimulated metastatic BC cells, enhanced the malignant behaviors of BC cells, including proliferation, invasion, and chemoresistance, through activating the ARF6-mediated Src/PI3K/Akt signaling pathway.22 BC exosomal miR-134-5p could suppress BC growth by inhibiting ARHGAP1/PI3K/AKT pathway, indicating a therapeutic role for miR-134-5p.23 MiR-10b from BC exosome also promoted cell invasion ability in vitro by targeting HOXD10 and KLF4.24 In addition, Ding et al showed that exosomal miR-222 was not only highly expressed in the serum of BC patients with lymphatic metastases, but was also tightly linked to the high invasiveness of BC cell lines.25 Exosomal miR-222 facilitated migration and invasion of BC cells in vitro, and fostered tumorigenic properties by directly targeting the tumor-suppressive PDLIM2 and consequently activating the NF-κB signal pathway.25 BC exosomes shuttled miR-1910-3p to breast epithelial cells and BC cells, and activated NF-κB and WNT/β-catenin pathways by suppressing MTMR3 expression, ultimately promoting BC cell propagation, metastasis, and autophagy.26 Exosomal miR-363-5p was validated to regulate platelet-derived growth factor (PDGF) activity by directly targeting PDGFB to constrain BC cell migration properties.27 MiR-7641 was an important component of BC cell exosomes and significantly promoted BC tumor growth.28 Exosomal miR-7-5p downregulation facilitated BC migration and invasion by targeting RYK and engaging the atypical WNT pathway.29
It is worth noting that the treatments represented by radiotherapy and chemotherapy can also affect the BC process by triggering changes in exosomal miRNAs. Radiation-induced bystander effect (RIBE), mediated by 2 Gy X-ray, could promote greater invasiveness of MCF-7 cells by enhancing their exosomal secretion.30 Among them, changes to critical miRNAs in these tumor exosomes, such as up-regulated miR-30a, miR-9a, and down-regulated miR-200b, were responsible for this phenomenon.
Interaction Between BC Cell-Stroma Cell
BC cell-derived exosomes can stimulate the conversion of normal fibroblasts (NFs) to CAFs. BC-originated exosomes boosted the activation of CAFs via the miR-146a/TXNIP axis, which initiated the Wnt pathway, thereby enhancing the aggressiveness and metastasis capability of BC cells.31 Additionally, miR-9 of BC exosomes enhanced CAF malignant behaviors to cause tumor growth.32 CAF-domesticated monocytes secrete exosomal miR-181a to activate AKT signaling in breast cancer cells and exert pro-tumor effects.33 Exosomes derived from tumor-adipocyte interactions, containing miR-144 and miR-126, could activate beige/brown differentiation and reshape the metabolism profile in resident stromal adipocytes to elicit BC progression.34 Hashimoto et al proposed a novel pathway by which tumor-derived hsa-miR-940 was transferred to mesenchymal stem cells (MSCs) via exosomes and promoted the phenotype of bone metastasis.35
Roles in Angiogenesis and Vascular Remodeling
Tumor vascular system disorder is not only an important tumor characteristic, but also a key factor impacting tumor hypoxia, metastasis invasion, and treatment resistance.36 Feng et al showed that BC cell-released exosomal miR-22-3p, was able to inhibit TAGLN-sustained tumor vascular abnormalities, and enhanced EMT transformation and BC stemness, thereby leading to tumor budding and distant metastasis. Aggressive BC cell lines secreted a specific type of miR-105, a regulator that targeted the tight junction protein ZO-1 to mediate cell migration movement.37 And in endothelial monolayers, BC-released miR-105 via exosome transfer essentially disrupted the completeness of these naturally formed barricades for blocking metastasis. Further in vivo assay confirmed the inhibition of miR-105 acted to restrain the BC metastasis capability with enhanced vascular integrity.
Endothelial barrier penetration is a key mechanism for proximal and distal tumor metastasis, contributing to nutrient exchange and migration of tumor cells and lymphocytes. Di Modica’s study showed that miR-939 expression in the TNBC cohort significantly correlated with lymph node (LN) status and predicted worse disease-free survival.38 Moreover, overexpressed miR-939 in exosomes released from MDA-MB-231 cells, could be internalized in HUVEC cells, thereby decreasing VE-cadherin and promoting transendothelial migration of MDA-MB-231 cells by disrupting the endothelial barrier.38
Roles in Organotropic Metastasis
Let-7a is a tumor suppressor-like miRNA and is significantly downregulated in various cancer type, including BC. Let-7a, transferred by exosomes, suppressed the proliferation, migration, and invasion of MDA-MB-231 cells by binding to the 3’-UTR region of c-Myc gene with decreased c-Myc expression.39 Furthermore, the pluripotency factor Lin28B facilitated BC lung metastasis by establishing immunosuppressive pre-metastatic ecological niches with neutrophil recruitment and N2 neutrophil conversion.40 The release of exosomes with low let-7 characteristics by BC cells was a prerequisite for this process. Both high Lin28B and low let-7s in tumors were indicators of poor prognosis and lung metastasis in BC patients.40
Abnormalities in glycolysis and glucose metabolism are one of the main features of cancer. BC tumors reprogram the TME by reprogramming changes in metabolic levels, leading to reprogramming of energy metabolism. High abundance of miR-122 uploaded in BC exosomes enhanced the nutrient supply in the pre-metastatic niche to promote tumor metastasis, through the downregulation of glycolytic enzyme pyruvate kinase.41 Meanwhile, in vivo inhibition of miR-122 restored the glucose uptake capacity in distant organs, thereby significantly reducing the extent of breast cancer brain metastasis (BCBM) and lung metastases. Moreover, BC-originated exosomal miR-105 enhanced BC growth via MYC-dependent metabolic reprogramming in tumorous stromal CAFs, accompanied by elevated glucose and glutamine metabolism to fuel adjacent BC cells.42 Wang et al reported an important molecular biological mechanism of tumor exosome-mediated liver metastasis. Specifically, the cancer-secreted exosomal miR-4443 up-regulated matrix metalloproteinase (MMP) in the TME at the primary and distal metastatic sites via downregulation of TIMP2, allowing BC cells to segregate from the primary tumor and metastasize into the neoplastic milieu.43
Patients with estrogen receptor (ER)+ have a higher bone metastatic property than negative patients, and this bone-oriented nature of ER tumors is still intriguing. Wu et al found that bone-tropic BC cells secreted miR-19a and integrin-binding sialoprotein (IBSP), leading to an increase in circulating levels.44 Furthermore, IBSP could recruit osteoclasts to establish an osteoblastic microenvironment in bone, which helped to transport exosomal miR-19a to osteoclasts and induce osteoclast formation. Within this mechanism, ER+ BC cells constructed a distinct TME conducive to bone colonization. Guo et al demonstrated that miR-20a-5p was highly expressed in tumor tissues and MDA-MB-231 cell exosomes.45 Their further in vitro studies highlighted that miR-20a-5p from BC exosomes could promote tumor cell migration and invasion, and enhanced osteoblast proliferation and differentiation by targeting SRCIN1. In another bone metastasis study, SCP28, a BC cell line with bone metastasis capacity, was able to cause osteoclast differentiation and activation and reduced bone density through secreted exosomes.46 Exosomal miR-21 derived from SCP28 cells displayed the ability to regulate PDCD4 protein levels to promote osteoclastogenesis, being an important regulator of bone-metastatic niche formation.
Exosome-Derived miRNAs from Other Cells
CAF-derived exosomes play an integral role in tumorigenesis, epithelial-mesenchymal transition (EMT), metastasis, and immune responsiveness. CAF-exosome miRNAs delivered to BC cells can induce a series of target protein silencing effects and corresponding cascade responses to reshape the malignant behavior of tumors. Focal adhesion kinase (FAK) was an essential kinase for CAFs to promote BC migration and metastasis.47 Importantly, FAK-null CAFs had correspondingly altered exosomal miRNA expression profiles compared to normal CAFs, such as miR-16 and miR-148a in exosomes contributing to reduced tumor cell activity and metastasis. Liu et al revealed that oncogenic miR-3613-3p was up-regulated in exosomes from TGF-β1 cultured fibroblasts and CAFs from BC tissues.48 Mechanistically, downregulation of miR-3613-3p in exosomes of CAFs inhibited cell proliferation and metastasis of BC by targeting SOCS2. Exosomal miR-18b of CAFs stimulated nuclear Snail allotropy by targeting TCEAL7 to induce NF-κB pathway activation, leading to enhanced EMT, invasion, and lung metastasis in BC tumors.49 Chen et al verified that exosomal miR-500a-5p from patient-derived CAFs was internalized and resulted in BC cell proliferation and enhanced pulmonary surface nodules in the xenograft model, by targeting USP28.50 This suggested that inhibition of CAF-derived miR-500a-5p was a potentially effective intervention for treating BC. CAF exosomal miR-4516 was markedly reduced in TNBC tissue, and its deficiency promoted tumor progression in a FOSL1-dependent pathway.51
MSC-derived exosomal miRNAs can act as anti-angiogenic molecules that inhibit blood vessels and affect tumor progression. Pakravan et al deciphered a paracrine mechanism of MSC-derived exosomal miR-100 that restrained angiogenesis by inhibiting mTOR/HIF-1α/VEGF signaling axis in BC cells.52 In addition, MSC exosomal miR-16 could target and inhibit VEGF expression, possessing an anti-angiogenic function.53 In a TNBC study, miR-106a-5p was more abundant in TNBC tissues and BC cells and was positively associated with tumor grade.54 Functional assays showed that miR-106a-5p derived from bone marrow-mesenchymal stem cells (BMSCs) was a facilitator in enhancing TNBC progression. The incubation with miR-106a-5p-overexpressed exosomes of BMSCs with BC cells endowed more powerful carcinogenic properties in mice. Notably, exosomal miRNAs in patient BMSCs, especially miR-23b, conferred BC cell dormancy in the metastatic niche by inhibiting downstream MARCKS genes.55 BC cells promoted the release of miR-222/223-rich exosomes from MSCs, which in turn promoted BC cell dormancy and resistance to bone metastases.56
Preadipocytes and mature adipocytes are important components in BC TME, and exosomes derived from preadipocytes also possess important functions in modulating the cancer stem cells (CSCs) niches. MiR-140 is lowly expressed in several tumors and is involved in regulating the biological behavior of tumor cells. Gernapudi et al showed that miR-140 expression in preadipocyte-derived exosomes effectively repressed the renewal of CSCs, thus affecting the growth and metastatic properties of BC tumors.57 Indeed, adipocytes adjacent to BC cells, represented by cancer-related adipocytes (CAAs), are the frontier adipocytes of tumor infiltration, and can interact with BC cells in the manner of carriers such as ncRNA and exosomes, and affect their respective behaviors.
Exosome-Derived miRNAs in Immune Regulation
Exosome-Derived miRNAs from BC Cells
Macrophages are the most abundant tumor-infiltrating immune cell type with multiple functions and close interaction with tumors.58 Macrophages have extremely high tumor heterogeneity, but are usually divided into M1 and M2 types, with M2 being the predominant type of infiltrating tumor-associated macrophages (TAM) that can support tumor metastasis and invasion.35 Macrophage exosomes are the most studied form of interaction between macrophages and BC tumors.
Blocking the production of M2 macrophage exosomes and the interactions mediated by them may reverse the tumor-suppressive features. 4T1 cell-derived exosomes delivered miR-33 to IL-4-induced M2 macrophages, and thus polarized M2 polarizes to an M1 phenotype, as manifested by increased secretion of TNF-α/IL-1β, and decreased IL-10/TGF-β secretion.59 These macrophages internalized with miR-33 could significantly inhibit the growth and invasion of 4T1 cells. Coincidentally, Moradi-Chaleshtori et al utilized a similar approach to deliver tumor-derived exosomal miR-130 to macrophages, resulting in phenotypic and functional up-regulation of M1 macrophages.60,61 The effect of repolarization of M2 to M1 made macrophages more potent in killing tumor cells. 4T1 cell exosome-mediated miR-183-5p transfer could enhance NF-κB pathway and secretion of anti-inflammatory IL-1β and IL-6 factors in macrophages by targeting PPP2CA.62 Exosomal miR-222 released by adriamycin-resistant BC cells conferred M2 macrophage polarization through PTEN/Akt pathway, leading to BC progression.63 Tumor-derived exogenous miR-148b-3p contributes to BC migration and invasion through TSC2/mTORC1-mediated M2 macrophage polarization.64 These studies further reinforce the non-negligible roles of specific BC-exosomal miRNAs in TME interactions.
The effect of exosomes secreted by BC cells on macrophages facilitates the establishment of ecological niches associated with BCBM and lung metastasis. The tumor-derived exosomal miR-138-5p was transferred to activate the M2 phenotype and suppressed M1 polarization, which led to an upregulation of the number of niches formed in mouse lung metastases.65 In addition, the circulating miR-138-5p levels were positively correlated with BC progression. Xing et al demonstrated that lncRNA XIST-mediated elevated expression of BC exosomal miR-503 promoted M1-M2 polarization and M2 functional activation in microglia, consequently leading to T cell proliferation suppression.66 Thus, exosomal miR-503 is an important substance of the immune microenvironment affecting the interaction between tumor cells and T cells, and the correlated pathway triggered BCBM.
Exosome-Derived miRNAs from Immune Cells
Macrophages also have the nefarious function of delivering exosomal oncogenic miRNAs to BC cells. An earlier study by Yang et al showed that exosomes released by IL-4-activated M2 macrophages conducted miR-223 into BC cells, mediating increased BC cell invasion by inhibiting Mef2c-b-catenin pathway.67 Yue et al proposed a novel targeting mechanism whereby exosome-derived miR-5100 from PGRN-/- TAMs could attenuate the metastatic ability of BC cells by inhibiting the CXCL12/CXCR4 axis.68 This regulatory axis might be one of the main reasons for the inhibition of BC lung metastasis in PGRN-/- mice. MiR-503-3p was highly expressed in BC tissues and cell lines. Meanwhile, the macrophage-derived exosome miR-503-3p facilitated glycolysis and reduced mitochondrial oxidative phosphorylation by targeting DACT2 and activating the Wnt/β-catenin pathway in BC cells.69
Myeloid-derived suppressor cells (MDSCs), a heterogeneous and plastic group of myeloid precursors with immunosuppressive capacity, are important negative regulators of inflammation and the immune response within TME.70 Exosomes shuttled by MDSCs are also equipped with the immunosuppressive ability of parental cells. In vivo, miR-9 and miR-181a in BC exosomes activated the JAK/STAT signaling by binding to SOCS3 and PIAS3 accordingly, thereby promoting the expansion of early-stage MDSCs and corresponding immunosuppressive state.71 DOX-mediated MDSC exosomal miR-126a reduced DOX-caused MDSC death and promoted tumor angiogenesis in a S100A8/A9-dependent manner, and promoted chemoresistance and lung metastasis.72 Exosomal miRNAs influence immune effects and tumor proliferation, vascular and malignant behavior through their transmission between tumor cells and immune cells (Table 1).
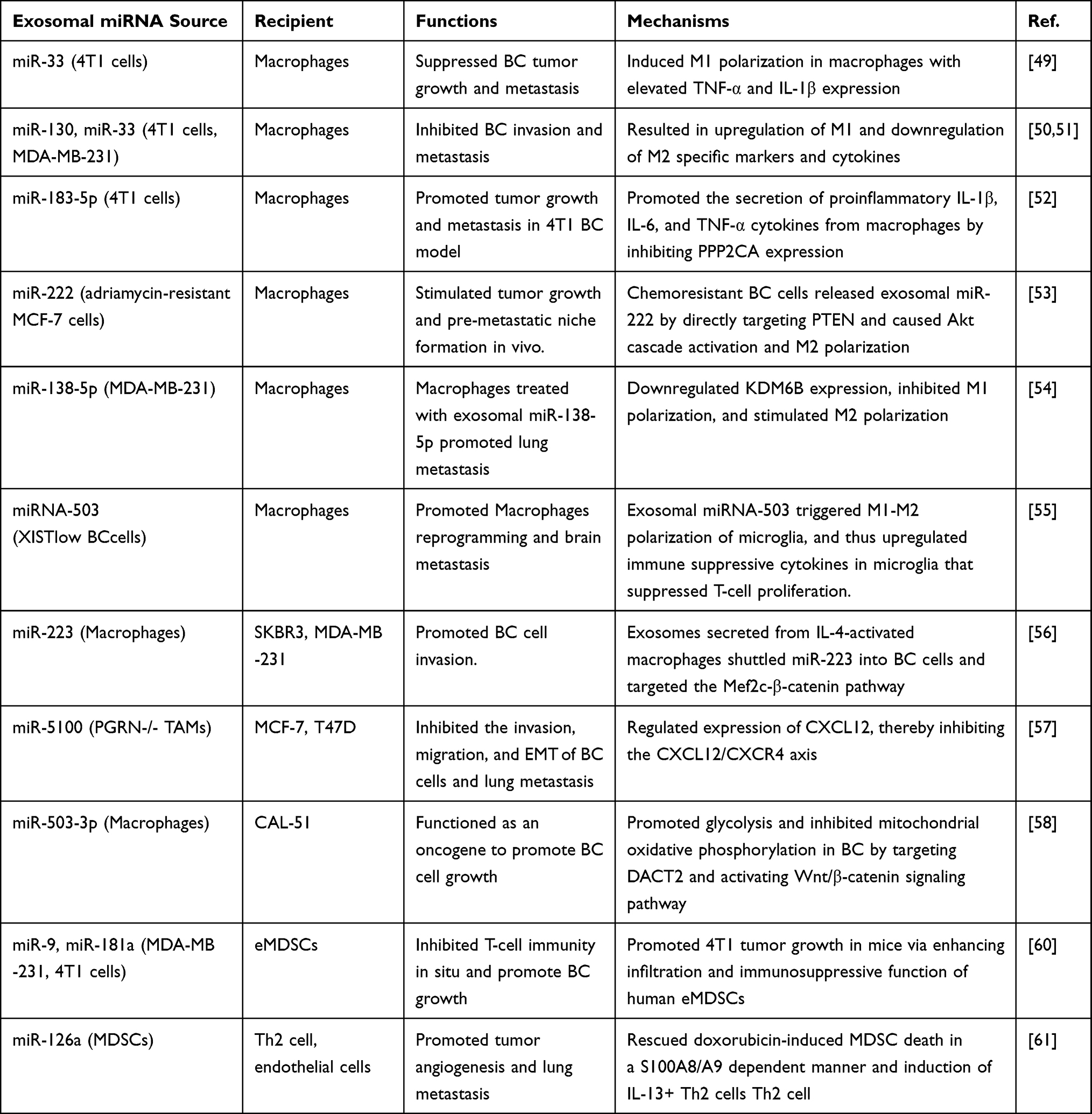 |
Table 1 The Roles of Exosomal miRNA in Immune Regulation in BC |
Exosome-Derived miRNAs in BC Diagnosis
Individualized diagnosis and early screening for BC are essential. Existing conventional diagnostic modalities such as magnetic resonance imaging (MRI), ultrasound (US), mammography, and case-based diagnostic modalities have achieved considerable accomplishments, but there is still room for improvement in terms of non-invasiveness, timeliness, and accuracy. Accurate, efficient, and sensitive exosomal miRNA diagnostic strategies will provide novel and effective tools for BC risk prediction, early screening, and treatment surveillance. Exosomes and exosomal miRNAs isolated from saliva, blood, or body fluid samples of BC patients with in situ tumors or metastases to the brain, lung, bone, or viscera, are potentially valuable, non-invasive diagnostic or prognostic biomarkers for accurate BC diagnosis, including differential diagnosis and treatment monitoring.
Differential Diagnosis
The decreased miR-17-5p, and up-regulated hsa-miR-423-5p and miR-202 in serum exosomes, were fascinating indicators to differentiate BC patients from healthy control.73–75 Through bioinformatics and experimental validation, Xin et al found that miR-455-5p and miR-1255a were involved in CDKN1B-mediated cell cycle process and SMAD4-mediated TGF-β pathway, respectively.76 Whereas high expression of miR-455-5p (basal-like) and miR-1255a (overall) was correlated with poorer overall survival, and high expression of their target genes was linked to superior overall, non-recurrence or non-distant metastasis survival. Circulating exosome miR-155 could be used to indicate the prognosis of patients with trastuzumab resistance, showing that higher miR-155 predicted poorer progression free survival (PFS) and event-free survival (EFS).77 MiR-1246 also has a similar indicator function.77,78 Exosome-loaded miR-223-3p was associated with BC malignancy and could serve as a predictor for the early detection of invasive BC.79 In addition, the miR-1910-3p expression alteration in serum exosomes of BC patients might also be regarded as a diagnostic indicator. The detection of serum exosomal miR-1910-3p in combination with tumor marker CA153 in routine blood tests, could significantly improve the sensitivity and specificity of serum diagnosis of BC.26 Wang et al successfully identified a significantly down-regulated miR-363-5p in the plasma exosomes of BC patients with LN metastasis, which was also confirmed by independent dataset validation.27 Exosomal miR-363-5p in BC plasma was related to better overall survival (OS) rate, and possessed the potential to differentiate LN+ and LN- BC status. The expression levels of miRNA-21-5p and miR-10b-5p in serum exosomes also could be used for the evaluation of BC diagnosis and prognosis.80
High expression of hsa-miR-576-3p and low expression of hsa-miR-130a-3p in serum exosomes of BC patients were significantly correlated with the prognosis of BCBM patients.81 In addition, hsa-miR-340-5p levels were closely related to Ki67+ tumor cells, while hsa-miR-342-3p levels were negatively relevant to the tumor stage. Li et al used qRT-PCR to confirm that serum exosomal miR-148a was differentially distributed in patients with benign and malignant breast tumors and healthy controls.82 Specifically, there was a significant association between serum exosomal miR-148a enrichment and LN metastasis, TNM staging, and differentiation, and BC patients with low serum exosomal miR-148a expression levels had poorer overall and disease-free survival. Therefore, serum exosomal miR-148a was a high-value target for differentiating BC patients from healthy volunteers and for determining tumor prognosis. The up-regulated exosomal hsa-miR-21-5p in plasma can be used as an effective biomarker to differentiate healthy individuals from BC patients, with a sensitivity and specificity of 86.7% and 93.3%, respectively.83 MiR-7641 was significantly associated with survival prognosis of BC patients, with significantly higher plasma expression levels in patients with metastasis.28
In addition to serum and plasma indicators, exosomal miRNAs present in other body fluids, such as tears, urine, and saliva, can also serve as potential indicators for BC differential diagnosis. MiR-21 and MMP-1 in urinary exosomes were used as combined diagnostic indicators by Ando et al and were able to achieve a sensitivity and specificity of 95% and 79% for metastatic BC.84 This indicated that urinary exosomal miRNAs in tumor patients still had a diagnostic value that was not weaker than blood specimens. Inubushi et al proposed a novel perspective that miR-21 and miR-200c were highly expressed in tear exosomes from patients with metastatic BC, which might be biomarkers for metastatic BC diagnosis.85 However, the number of case analyses included in this study was relatively small and this novel concept deserved further exploration.
Single exosomal miRNA indicator has shown prognostic efficacy, while the model constructed by multiple indicators can provide a more comprehensive and clear understanding of the disease. The combinational predictive model based on miR-421, miR-128-1, and miR-128-2 associated with genomic instability (GI) in plasma exosomes, was regarded as a signature for BC early diagnosis and prognosis.86 Notably, the combination of plasma exosomal miR-1246 and miR-21 is also an ideal method to indicate BC status compared to their separated levels.87 There are few relevant studies at present, but it is very valuable.
Treatment Monitoring
During treatment, the contents of exosomes undergo dynamic and specific changes in tumor tissue and body fluids represented by blood. Corresponding changes in exosomal miRNA expression abundance are effective indicators of disease treatment effects. Li et al found that the expression level of specific miRNA in serum exosomes was significantly correlated with LN metastasis and the clinical stage of BC.88 Among them, miR-3662, miR-146a, and miR-1290 were up-regulated in serum exosomes, and they were ponderable biomarkers for monitoring BC patient status during surgery and chemotherapy.
In another related study, the exosomal miR-21 and miR-105 expression levels were higher in metastatic patients before neoadjuvant therapy.89 During neoadjuvant therapy, exosomal miRNA-21 expression levels were directly connected with tumor size and negatively linked to Ki67 expression. Finally, elevated exosomal miR-21, miR-222, and miR-155 levels were remarkably related to the existence of circulating tumor cells (CTCs). Exosomal expression levels of miR-3662, miR-146a, and miR-1290 in serum have attracted much attention, specifically for monitoring patient status during surgery and chemotherapy.88 These studies collectively suggest that exosomal miRNA-based liquid biopsy and related diagnostic methods can serve as a useful complementary tool to evaluate the diagnosis, treatment, and prognostic monitoring for BC management.
Exosome-Derived miRNAs in BC Treatment
Drug Resistance
Exosomes of multiple cell sources, including BC cells, and stem cells, are involved in BC drug resistance and therapeutic progression. Exosomes from drug-resistant BC cells propagate chemoresistance by a horizontal transfer of miRNAs. Aberrant miRNA and exosomal miRNA expression levels in the serum of BC patients are intensively implicated in resistance to multiple therapeutic agents for BC.90,91 In immunotherapy, anti-PD-1 treatment promoted the expression of lymphocyte-derived exosomal miRNA-4315 and suppressed BIM-mediated tumor apoptosis, endowing the potential of exosomal miRNA-4315 in blood as patient stratification for anti-PD-1 treatment.92 Exosome-shuttled miR-567 restrained the trastuzumab resistance, and could be internalized by recipient BC cells to enhance the curative effect of chemotherapy by inhibiting ATG5 in BC.93
The expression profiles of miRNAs in TNBC chemotherapy-sensitive and drug-resistant tissues are quite heterogeneous. Among them, miR-770 was closely related to chemotherapy resistance, and its high expression implied a better BC prognosis.94 It was found that exosomal miR-770 could be transferred to TNBC cells, resulting in enhanced DOX resistance and metastasis both in vitro and in xenograft model. Exosome-mediated transfer of miR-3613-5p and miR-181b-5p also promoted doxorubicin resistance in BC cells, by targeting PTEN and BCLAF1 respectively.15,95 Andreeva et al identified miR-181a-2 as a key miRNA mediating tamoxifen drug resistance.96 This action was consistent with a continuous blocking of oestrogen receptors and promotion of the PI3K/Akt pathway. Exosomal miRNA-205 derived from tamoxifen-resistant MCF-7 promoted BC chemoresistance and tumorigenesis, but was triggered by binding to E2F1.92 Furthermore, exosomal miR-221/222 and miR-9, were critical supporters of tamoxifen-related resistance in ER+ BC.97,98 Exosomal miR-423-5p also mediated the cisplatin resistance in TNBC cells.78
Emerging findings have revealed that horizontally transferred exosomal miRNAs are key mechanisms of chemotherapy resistance through which BC cells acquire CSC-like characteristics and drug resistance transmission. For instance, miR-155 regulated the stemness and chemoresistance of BC cells and CSC cells.20 Chemotherapy-induced miR-378a-3p and miR-378d in BC exosomes, which enhanced BC stemness characterized by WNT and NOTCH activation, leading to DOX and PTX chemoresistance via activation of EZH2/STAT3 signaling in xenograft models.93 Consistent with this, chemotherapeutic medicine plus EZH2 inhibitor tazemetostat significantly relieved exosome-mediated resistance in a nude mouse tumor xenograft model.
Drug Delivery Based on Stem Cell-Derived Exosomes
Stem cell-derived exosomes have exhibited prospective advances in treating various cancer types, due to their encapsulated lethal effector molecule. Exosomes derived from human umbilical cord-derived MSCs (HUCMSCs), BMSCs, and adipose-derived stem cells (ADSCs), are supposed to be the most common stem cell types in BC treatment. MiR-342-3p is lowly expressed in patients with metastatic tumors. MiR-342-3p uploaded in MSC exosomes not only inhibited metastasis and chemoresistance of BC cells by targeting ID4,99 but also by inhibiting INHBA/IL13Rα2 axis.100 Exosomal miRNA-551b-3p from BMSCs suppressed BC progression by modulating TRIM31/Akt signaling.101 MiR-1236 carried by ADSC exosomes promoted the resistance response of BC cells to cisplatin, specifically involving two distinct mechanisms related to SLC9A1 downregulation and Wnt/β-linked protein inactivation.102 Similarly, Du et al showed that HUCMSC exosome had anticancer effects in ER- MCF-7 cells.103 Specifically, HUCMSC exosome-derived miR-21-5p inhibited the expression level of ZNF367 by binding to the ZNF367 3’UTR, leading to reduced MCF-7 cell invasiveness. MiR-3182 from HUCMSC-exosomes internalized by TNBC cells, caused diminished invasiveness of recipient cells and induced apoptosis by targeting the mTOR-S6KB1 pathway.104 MiR-148b-3p derived from HUCMSC exosomes could target TRIM59 to inhibit BC progression.105 These results suggest that exosomal miRNAs harboring tumor-killing effects might be credible therapeutic candidates in BC therapy. Sheykhhasan et al exploited the function of ADSC-derived exosomes on T-47D tumor cells.106 Their results showed that ADSC exosome-mediated miR-145 delivery effectively inhibited tumor cell apoptosis and metastasis.
Discussion
Exosomes are involved in multiple cellular interactions in BC TME and consequently have a fundamental impact on BC processes. However, because of the wide range of sources of exosomes, the direction and results of exosome propagation deserve to be explored in depth. Moreover, ncRNA substances in exosomes can influence the expression of receptor cells in an epigenetic manner. In terms of tumor immune effects, since most studies have focused on the interaction between macrophages and tumor cells, the exosomal interaction properties and the influence of other humoral and cellular immune cells are also a direction worth investigating. It is worth mentioning that exosomes secreted by the same cell at different stages or conditions are different. The exosome-rich component of early and advanced tumors differs greatly in their ability to direct and domesticate the metastatic niche.107 This highlights that maternal exosomes have different effects on recipient cells in different stages of the same cell.
The main separation methods for exosomes include differential centrifugation, density gradient centrifugation, sedimentation, filtration, size exclusion chromatography, kit extraction, immunoaffinity, and microfluidics. However, realizing high-purity, high-efficiency and non-invasive separation of exosomes is still the main challenge for exosome separation at present. This is because various separation methods possess their advantages and disadvantages, such as expensive equipment, insufficient purity, and protein contamination. A combination of multiple separation methods can be used to obtain a higher quality yield of exosomes than a single method.
For detection approaches, the identification of exosomes is mainly dependent on morphological features, particle size, and signature proteins, which are detected by electron microscopy, dynamic light scattering, nanoparticle tracer analysis, Western blot method, and flow cytometry. Of these methods, the main advantage of exosome size-based assays is their simplicity, rapidity, and suitability for high-throughput analyses, but their accuracy is relatively low. In contrast, assays that characterize biofunctional molecules in exosomes are usually more accurate, but the sample pre-processing steps are cumbersome and time-consuming. The miRNAs in exosomes are often still analyzed by conventional qPCR. The combination of multiple methods is the current mainstream of exosome detection.
For the diagnostic application, exosome-related diagnostics have shown huge prospects and are accompanied by the potential to revolutionize the strategy of clinical diagnosis and management of BC. This is essential because exosomal miRNA levels and contents are an accurate reflection of parental BC cells. However, there are currently non-homogeneous processes for the isolation and purification of exosomes. This causes non-uniformity in exosome purity, recovery, and particle size, which in turn affects the evaluation of BC diagnosis, metastasis, and treatment monitoring. In addition, the lack of standardized methods among different studies has resulted in significant differences in key metrics such as threshold, sensitivity, specificity, and detection limit for exosomal miRNA diagnosis.
Finally, in the aspect of therapeutic application, the genetic substances in exosomes can be used as targets for tumor therapy, and on the other hand, exosomes can also be used as vectors for targeted therapies. Highly expressed substances in BC exosomes can be intervened with specific inhibitors or antibodies, thus cutting off the malignant propagation pathway of tumors. In addition, blocking exosome secretion from BC or evil cells and their interaction also produces effective tumor eradication. Tumor-derived exosomes possess tumor-targeted homing recognition, high biosafety, and secretion, whereas non-tumor exosomes, represented by MSCs, can be genetically programmed or uploaded with drugs that carry tumor-killing factors to curb BC growth.
Conclusion
Collectively, exosome-derived miRNAs possess the ability to promote tumor growth, proliferation, metastasis, and invasion, as well as the colonization of specific organs, demonstrating their potential values in clinical BC diagnosis and treatment (Figure 2). This field will provide novel exosomal miRNA-based strategies for combating BC.
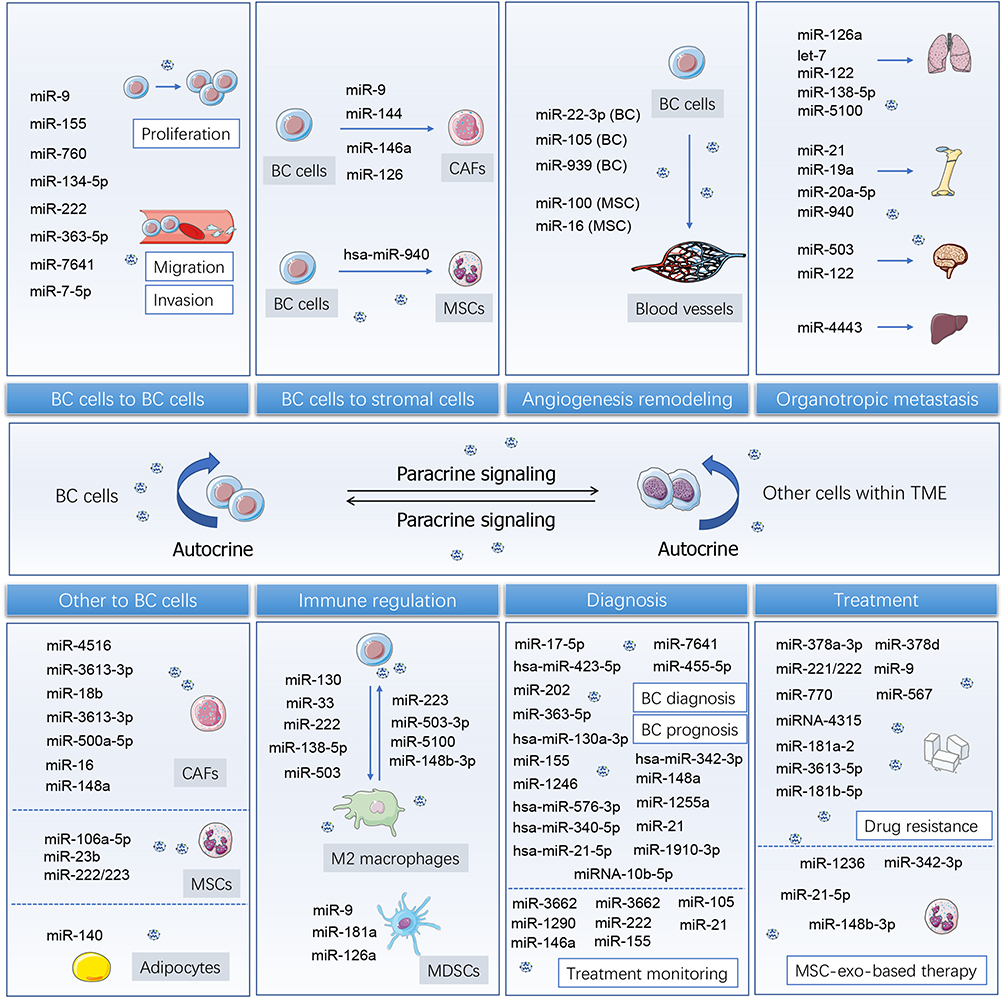 |
Figure 2 Exosome-derived miRNAs are emerging supporters in breast cancer progression, diagnosis, and treatment. Exosomes are important carriers for close interaction between BC cells and adjacent BC cells, stromal cells, vascular cells, and immune cells. Exosomal miRNAs shuttling between receptors and ligands can cause malignant remodeling of receptor cells, and then promote tumor growth, proliferation, metastasis, and invasion, as well as the colonization of specific organs. Tumor-derived exosomal miRNAs or circulating exosomal miRNAs show characteristic expression in different stages of specific diseases or under treatment, thus being endowed as tumor diagnostic markers. |
Exosome-encapsulated miRNAs and other biomolecules have the potential to indicate tumor subtype, metastasis, and prognosis due to their abundance and expression characteristics. At present, exosomal miRNAs have not demonstrated a confirmatory diagnosis of different subtypes. Therefore, the subsequent explorations of prognosis, estrogen receptor status, and potential metastasis by exosomal miRNAs for different breast tumor subtypes, are an urgent need for future advancement. Besides, targeted blockade or activation strategies based on exosomal miRNAs have the potential to provide novel therapeutic strategies for inhibiting breast cancer progression, suppressing chemoresistance, and adjuvant augmentation of conventional therapy. Exosomes can also be used as drug delivery carriers for RNAs, proteins, small molecules, and other drugs for combating BC. Overall, exosome-based therapeutic modalities are emerging and fascinating strategies for BC tumor treatment. Up to the present, most of the studies on exosomes are still in the preclinical stage, and the subsequent continuous and in-depth research is promising.
Abbreviations
BMSCs, bone marrow mesenchymal stem cells; BC, breast cancer; BCBM, breast cancer brain metastasis; CSCs, cancer stem cells; CAFs, cancer-associated fibroblasts; CTCs, circulating tumor cells; EMT, epithelial-mesenchymal transition; ER, estrogen receptor; EFS, event-free survival; ECM, extracellular matrix; FAK, focal adhesion kinase; HUCMSCs, human umbilical cord-derived MSCs; IBSP, integrin-binding sialoprotein; LN, lymph node; MRI, magnetic resonance imaging; MHC, major histocompatibility complex; MMP, matrix metalloproteinase; MSCs, mesenchymal stem cells; MVBs, multiple vesicular bodies; MDSC, myeloid-derived suppressor cells; ncRNAs, non-coding RNAs; NFs, normal fibroblasts; PDGF, platelet-derived growth factor;PARP, poly(ADP-ribose) polymerase; PFS, progression-free survival; RIBE, radiation-induced bystander effect; TNBC, triple-negative breast cancer; TME, tumor microenvironment; TAMs, tumor-associated macrophages; US, ultrasound.
Acknowledgments
Jie Li, Dejiao He and Yajun Bi are co-first authors for this study. We thank the institute for providing the funding and the individuals for offering help during the manuscript process.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis, and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Nature Science Foundation of Hubei Province (2020CFB685).
Disclosure
All the authors declare that there is no conflict of interest in this work.
References
1. Loibl S, Poortmans P, Morrow M, Denkert C, Curigliano G. Breast cancer. Lancet. 2021;397(10286):1750–1769. doi:10.1016/S0140-6736(20)32381-3
2. Cao W, Chen HD, Yu YW, Li N, Chen WQ. Changing profiles of cancer burden worldwide and in China: a secondary analysis of the global cancer statistics 2020. Chin Med J. 2021;134(7):783–791. doi:10.1097/CM9.0000000000001474
3. Arnold M, Morgan E, Rumgay H, et al. Current and future burden of breast cancer: global statistics for 2020 and 2040. The Breast. 2022;66:15–23. doi:10.1016/j.breast.2022.08.010
4. von Arx C, De Placido P, Caltavituro A, et al. The evolving therapeutic landscape of trastuzumab-drug conjugates: future perspectives beyond HER2-positive breast cancer. Cancer Treat Rev. 2023;113:102500. doi:10.1016/j.ctrv.2022.102500
5. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478):eaau6977. doi:10.1126/science.aau6977
6. Paskeh MDA, Entezari M, Mirzaei S, et al. Emerging role of exosomes in cancer progression and tumor microenvironment remodeling. J Hematol Oncol. 2022;15(1):83. doi:10.1186/s13045-022-01305-4
7. Yang E, Wang X, Gong Z, Yu M, Wu H, Zhang D. Exosome-mediated metabolic reprogramming: the emerging role in tumor microenvironment remodeling and its influence on cancer progression. Signal Transduct Target Ther. 2020;5(1):242. doi:10.1038/s41392-020-00359-5
8. Fang Z, Ding Y, Xue Z, Li P, Li J, Li F. Roles of exosomes as drug delivery systems in cancer immunotherapy: a mini-review. Discov Oncol. 2022;13(1):74. doi:10.1007/s12672-022-00539-5
9. Xu YX, Pu SD, Li X, et al. Exosomal ncRNAs: novel therapeutic target and biomarker for diabetic complications. Pharmacol Res. 2022;178:106135. doi:10.1016/j.phrs.2022.106135
10. Sammut SJ, Crispin-Ortuzar M, Chin SF, et al. Multi-omic machine learning predictor of breast cancer therapy response. Nature. 2022;601(7894):623–629.
11. DeVeale B, Swindlehurst-Chan J, Blelloch R. The roles of microRNAs in mouse development. Nat Rev Genet. 2021;22(5):307–323. doi:10.1038/s41576-020-00309-5
12. Mori MA, Ludwig RG, Garcia-Martin R, Brandão BB, Kahn CR. Extracellular miRNAs: from biomarkers to mediators of physiology and disease. Cell Metab. 2019;30(4):656–673. doi:10.1016/j.cmet.2019.07.011
13. Melo SA, Sugimoto H, O’Connell JT, et al. Cancer exosomes perform cell-independent microrna biogenesis and promote tumorigenesis. Cancer Cell. 2014;26(5):707–721. doi:10.1016/j.ccell.2014.09.005
14. Scognamiglio I, Cocca L, Puoti I, et al. Exosomal microRNAs synergistically trigger stromal fibroblasts in breast cancer. Mol Ther Nucleic Acid. 2022;28:17–31. doi:10.1016/j.omtn.2022.02.013
15. Zhao S, Pan T, Deng J, et al. Exosomal transfer of miR-181b-5p confers senescence-mediated doxorubicin resistance via modulating BCLAF1 in breast cancer. Br J Cancer. 2023;128(4):665–677. doi:10.1038/s41416-022-02077-x
16. Sun S, Wang Z, Yao F, et al. Breast cancer cell-derived exosome-delivered microRNA-155 targets UBQLN1 in adipocytes and facilitates cancer cachexia-related fat loss. Hum Mol Genet. 2023;32(13):2219–2228. doi:10.1093/hmg/ddad055
17. Kim O, Tran P, Gal M, et al. RAS‑stimulated release of exosomal miR‑494‑3p promotes the osteolytic bone metastasis of breast cancer cells. Int J Mol Med. 2023;52(3):84. doi:10.3892/ijmm.2023.5287
18. Liu C, Li B, Lin H, et al. Multiplexed analysis of small extracellular vesicle-derived mRNAs by droplet digital PCR and machine learning improves breast cancer diagnosis. Biosens Bioelectron. 2021;194:113615. doi:10.1016/j.bios.2021.113615
19. Li X, Wang S, Mu W, et al. Reactive oxygen species reprogram macrophages to suppress antitumor immune response through the exosomal miR-155-5p/PD-L1 pathway. J Exp Clin Cancer Res. 2022;41(1):41. doi:10.1186/s13046-022-02244-1
20. Santos JC, Sarian LO, Matheu A, Ribeiro ML, Derchain SFM, Derchain SFM. Exosome-mediated breast cancer chemoresistance via miR-155 transfer. Sci Rep. 2018;8(1):829. doi:10.1038/s41598-018-19339-5
21. Kia V, Paryan M, Mortazavi Y, Biglari A, Mohammadi-Yeganeh S. Evaluation of exosomal miR-9 and miR-155 targeting PTEN and DUSP14 in highly metastatic breast cancer and their effect on low metastatic cells. J Cell Biochem. 2019;120(4):5666–5676. doi:10.1002/jcb.27850
22. Huang X, Lai S, Qu F, et al. CCL18 promotes breast cancer progression by exosomal miR-760 activation of ARF6/Src/PI3K/Akt pathway. Mol Ther Oncol. 2022;25(26):1–15. doi:10.1016/j.omto.2022.03.004
23. Yang C, Zhang G, Zhang Y, Zhang S, Li J, Liu Y. Exosome miR-134-5p restrains breast cancer progression via regulating PI3K/AKT pathway by targeting ARHGAP1. J Obstet Gynaecol Res. 2021;47(11):4037–4048. doi:10.1111/jog.14983
24. Singh R, Pochampally R, Watabe K, Lu Z, Mo YY. Exosome-mediated transfer of miR-10b promotes cell invasion in breast cancer. Mol Cancer. 2014;13(1):256. doi:10.1186/1476-4598-13-256
25. Ding J, Xu Z, Zhang Y, et al. Exosome-mediated miR-222 transferring: an insight into NF-κB-mediated breast cancer metastasis. Exp Cell Res. 2018;369(1):129–138. doi:10.1016/j.yexcr.2018.05.014
26. Wang B, Mao J, Wang B, et al. Exosomal miR-1910-3p promotes proliferation, metastasis, and autophagy of breast cancer cells by targeting MTMR3 and activating the NF-κB signaling pathway. Cancer Lett. 2020;489:87–99. doi:10.1016/j.canlet.2020.05.038
27. Wang X, Qian T, Bao S, et al. Circulating exosomal miR-363-5p inhibits lymph node metastasis by downregulating PDGFB and serves as a potential noninvasive biomarker for breast cancer. Mol Oncol. 2021;15(9):2466–2479. doi:10.1002/1878-0261.13029
28. Shen S, Song Y, Zhao B, et al. Cancer-derived exosomal miR-7641 promotes breast cancer progression and metastasis. Cell Commun Signal. 2021;19(1):20. doi:10.1186/s12964-020-00700-z
29. Liang Z, Liu L, Gao R, Che C, Yang G. Downregulation of exosomal miR-7-5p promotes breast cancer migration and invasion by targeting RYK and participating in the atypical WNT signalling pathway. Cell Mol Biol Lett. 2022;27(1):88. doi:10.1186/s11658-022-00393-x
30. AL-Abedi R, Tuncay Cagatay S, Mayah A, Brooks SA, Kadhim M. Ionising radiation promotes invasive potential of breast cancer cells: the role of exosomes in the process. Int J Mol Sci. 2021;22(21):11570. doi:10.3390/ijms222111570
31. Yang SS, Ma S, Dou H, et al. Breast cancer-derived exosomes regulate cell invasion and metastasis in breast cancer via miR-146a to activate cancer associated fibroblasts in tumor microenvironment. Exp Cell Res. 2020;391(2):111983. doi:10.1016/j.yexcr.2020.111983
32. Baroni S, Romero-Cordoba S, Plantamura I, et al. Exosome-mediated delivery of miR-9 induces cancer-associated fibroblast-like properties in human breast fibroblasts. Cell Death Dis. 2016;7(7):e2312–e2312. doi:10.1038/cddis.2016.224
33. Pakravan K, Mossahebi-Mohammadi M, Ghazimoradi MH, Cho WC, Sadeghizadeh M, Babashah S. Monocytes educated by cancer-associated fibroblasts secrete exosomal miR-181a to activate AKT signaling in breast cancer cells. J Transl Med. 2022;20(1):559. doi:10.1186/s12967-022-03780-2
34. Wu Q, Li J, Li Z, et al. Exosomes from the tumour-adipocyte interplay stimulate beige/brown differentiation and reprogram metabolism in stromal adipocytes to promote tumour progression. J Exp Clin Cancer Res. 2019;38(1):223. doi:10.1186/s13046-019-1210-3
35. Hashimoto K, Ochi H, Sunamura S, et al. Cancer-secreted hsa-miR-940 induces an osteoblastic phenotype in the bone metastatic microenvironment via targeting ARHGAP1 and FAM134A. Proc Natl Acad Sci. 2018;115(9):2204–2209. doi:10.1073/pnas.1717363115
36. Feng Y, Wang L, Wang T, et al. Tumor cell-secreted exosomal miR-22-3p inhibits transgelin and induces vascular abnormalization to promote tumor budding. Mol Ther. 2021;29(6):2151–2166. doi:10.1016/j.ymthe.2021.02.009
37. Zhou W, Fong MY, Min Y, et al. Cancer-Secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell. 2014;25(4):501–515. doi:10.1016/j.ccr.2014.03.007
38. Di Modica M, Regondi V, Sandri M, et al. Breast cancer-secreted miR-939 downregulates VE-cadherin and destroys the barrier function of endothelial monolayers. Cancer Lett. 2017;384:94–100. doi:10.1016/j.canlet.2016.09.013
39. Du J, Fan JJ, Dong C, Li HT, Ma BL. Inhibition effect of exosomes-mediated Let-7a on the development and metastasis of Triple negative breast cancer by down-regulating the expression of c-Myc. Eur Rev Med Pharmacol Sci. 2019;23(12):5301–5314. doi:10.26355/eurrev_201906_18197
40. Qi M, Xia Y, Wu Y, et al. Lin28B-high breast cancer cells promote immune suppression in the lung pre-metastatic niche via exosomes and support cancer progression. Nat Commun. 2022;13(1):897. doi:10.1038/s41467-022-28438-x
41. Fong MY, Zhou W, Liu L, et al. Breast-cancer-secreted miR-122 reprograms glucose metabolism in premetastatic niche to promote metastasis. Nat Cell Biol. 2015;17(2):183–194. doi:10.1038/ncb3094
42. Yan W, Wu X, Zhou W, et al. Cancer-cell-secreted exosomal miR-105 promotes tumour growth through the MYC-dependent metabolic reprogramming of stromal cells. Nat Cell Biol. 2018;20(5):597–609. doi:10.1038/s41556-018-0083-6
43. Wang J, Zhang Q, Wang D, et al. Microenvironment-induced TIMP2 loss by cancer-secreted exosomal miR-4443 promotes liver metastasis of breast cancer. J Cell Physiol. 2020;235(7––8):5722–5735. doi:10.1002/jcp.29507
44. Wu K, Feng J, Lyu F, et al. Exosomal miR-19a and IBSP cooperate to induce osteolytic bone metastasis of estrogen receptor-positive breast cancer. Nat Commun. 2021;12(1):5196. doi:10.1038/s41467-021-25473-y
45. Guo L, Zhu Y, Li L, et al. Breast cancer cell-derived exosomal miR-20a-5p promotes the proliferation and differentiation of osteoclasts by targeting SRCIN1. Cancer Med. 2019;8(12):5687–5701. doi:10.1002/cam4.2454
46. Yuan X, Qian N, Ling S, et al. Breast cancer exosomes contribute to pre-metastatic niche formation and promote bone metastasis of tumor cells. Theranostics. 2021;11(3):1429–1445. doi:10.7150/thno.45351
47. Wu HJ, Hao M, Yeo SK, Guan JL. FAK signaling in cancer-associated fibroblasts promotes breast cancer cell migration and metastasis by exosomal miRNAs-mediated intercellular communication. Oncogene. 2020;39(12):2539–2549. doi:10.1038/s41388-020-1162-2
48. Liu Y, Yang Y, Du J, Lin D, Li F. MiR-3613-3p from carcinoma-associated fibroblasts exosomes promoted breast cancer cell proliferation and metastasis by regulating SOCS2 expression. IUBMB Life. 2020;72(8):1705–1714. doi:10.1002/iub.2292
49. Yan Z, Sheng Z, Zheng Y, et al. Cancer-associated fibroblast-derived exosomal miR-18b promotes breast cancer invasion and metastasis by regulating TCEAL7. Cell Death Dis. 2021;12(12):1120. doi:10.1038/s41419-021-04409-w
50. Chen B, Sang Y, Song X, et al. Exosomal miR-500a-5p derived from cancer-associated fibroblasts promotes breast cancer cell proliferation and metastasis through targeting USP28. Theranostics. 2021;11(8):3932–3947. doi:10.7150/thno.53412
51. Kim JE, Kim BG, Jang Y, Kang S, Lee JH, Cho NH. The stromal loss of miR-4516 promotes the FOSL1-dependent proliferation and malignancy of triple negative breast cancer. Cancer Lett. 2020;469:256–265. doi:10.1016/j.canlet.2019.10.039
52. Pakravan K, Babashah S, Sadeghizadeh M, et al. MicroRNA-100 shuttled by mesenchymal stem cell-derived exosomes suppresses in vitro angiogenesis through modulating the mTOR/HIF-1α/VEGF signaling axis in breast cancer cells. Cell Oncol. 2017;40(5):457–470. doi:10.1007/s13402-017-0335-7
53. Lee JK, Park SR, Jung BK, et al. Exosomes derived from mesenchymal stem cells suppress angiogenesis by down-regulating VEGF expression in breast cancer cells. PLoS One. 2013;8(12):e84256. doi:10.1371/journal.pone.0084256
54. Xing L, Tang X, Wu K, Huang X, Yi Y, Huan J. LncRNA HAND2-AS1 suppressed the growth of triple negative breast cancer via reducing secretion of MSCs derived exosomal miR-106a-5p. Aging. 2021;13(1):424–436. doi:10.18632/aging.202148
55. Ono M, Kosaka N, Tominaga N, et al. Exosomes from bone marrow mesenchymal stem cells contain a microRNA that promotes dormancy in metastatic breast cancer cells. Sci Signal. 2014;7(332):ra63. doi:10.1126/scisignal.2005231
56. Bliss SA, Sinha G, Sandiford OA, et al. Mesenchymal stem cell-derived exosomes stimulate cycling quiescence and early breast cancer dormancy in bone marrow. Cancer Res. 2016;76(19):5832–5844. doi:10.1158/0008-5472.CAN-16-1092
57. Gernapudi R, Yao Y, Zhang Y, et al. Targeting exosomes from preadipocytes inhibits preadipocyte to cancer stem cell signaling in early-stage breast cancer. Breast Cancer Res Treat. 2015;150(3):685–695. doi:10.1007/s10549-015-3326-2
58. Chow A, Schad S, Green MD, et al. Tim-4+ cavity-resident macrophages impair anti-tumor CD8+ T cell immunity. Cancer Cell. 2021;39(7):973–988.e9. doi:10.1016/j.ccell.2021.05.006
59. Moradi-Chaleshtori M, Bandehpour M, Heidari N, Mohammadi-Yeganeh S, Mahmoud Hashemi S. Exosome-mediated miR-33 transfer induces M1 polarization in mouse macrophages and exerts antitumor effect in 4T1 breast cancer cell line. Int Immunopharmacol. 2021;90:107198. doi:10.1016/j.intimp.2020.107198
60. Moradi-Chaleshtori M, Shojaei S, Mohammadi-Yeganeh S, Hashemi SM. Transfer of miRNA in tumor-derived exosomes suppresses breast tumor cell invasion and migration by inducing M1 polarization in macrophages. Life Sci. 2021;282:119800. doi:10.1016/j.lfs.2021.119800
61. Moradi-Chaleshtori M, Bandehpour M, Soudi S, Mohammadi-Yeganeh S, Hashemi SM. In vitro and in vivo evaluation of anti-tumoral effect of M1 phenotype induction in macrophages by miR-130 and miR-33 containing exosomes. Cancer Immunol Immunother. 2021;70(5):1323–1339. doi:10.1007/s00262-020-02762-x
62. Guo J, Duan Z, Zhang C, et al. Mouse 4T1 breast cancer cell–derived exosomes induce proinflammatory cytokine production in macrophages via miR-183. J Immunol. 2020;205(10):2916–2925. doi:10.4049/jimmunol.1901104
63. Chen WX, Wang DD, Zhu B, et al. Exosomal miR-222 from Adriamycin-resistant MCF-7 breast cancer cells promote macrophages M2 polarization via PTEN/Akt to induce tumor progression. Aging. 2021;13(7):10415–10430. doi:10.18632/aging.202802
64. Hao C, Sheng Z, Wang W, et al. Tumor-derived exosomal miR-148b-3p mediates M2 macrophage polarization via TSC2/mTORC1 to promote breast cancer migration and invasion. Thorac Cancer. 2023;14(16):1477–1491. doi:10.1111/1759-7714.14891
65. Xun J, Du L, Gao R, et al. Cancer-derived exosomal miR-138-5p modulates polarization of tumor-Associated macrophages through inhibition of KDM6B. Theranostics. 2021;11(14):6847–6859. doi:10.7150/thno.51864
66. Xing F, Liu Y, Wu SY, et al. Loss of XIST in breast cancer activates MSN-c-Met and reprograms microglia via exosomal miRNA to promote brain metastasis. Cancer Res. 2018;78(15):4316–4330. doi:10.1158/0008-5472.CAN-18-1102
67. Yang M, Chen J, Su F, et al. Microvesicles secreted by macrophages shuttle invasion-potentiating microRNAs into breast cancer cells. Mol Cancer. 2011;10(1):117. doi:10.1186/1476-4598-10-117
68. Yue S, Ye X, Zhou T, et al. PGRN TAMs-derived exosomes inhibit breast cancer cell invasion and migration and its mechanism exploration. Life Sci. 2021;264:118687. doi:10.1016/j.lfs.2020.118687
69. Huang S, Fan P, Zhang C, et al. Exosomal microRNA-503-3p derived from macrophages represses glycolysis and promotes mitochondrial oxidative phosphorylation in breast cancer cells by elevating DACT2. Cell Death Discov. 2021;7(1):119. doi:10.1038/s41420-021-00492-2
70. Grover A, Sanseviero E, Timosenko E, Gabrilovich DI. Myeloid-derived suppressor cells: a propitious road to clinic. Cancer Discov. 2021;11(11):2693–2706. doi:10.1158/2159-8290.CD-21-0764
71. Jiang M, Zhang W, Zhang R, et al. Cancer exosome-derived miR-9 and miR-181a promote the development of early-stage MDSCs via interfering with SOCS3 and PIAS3 respectively in breast cancer. Oncogene. 2020;39(24):4681–4694. doi:10.1038/s41388-020-1322-4
72. Deng Z, Rong Y, Teng Y, et al. Exosomes miR-126a released from MDSC induced by DOX treatment promotes lung metastasis. Oncogene. 2017;36(5):639–651. doi:10.1038/onc.2016.229
73. Lv S, Wang Y, Xu W, Dong X. Serum exosomal miR-17-5p as a promising biomarker diagnostic biomarker for breast cancer. Clin Lab. 2020;66(9):1823–1834. doi:10.7754/Clin.Lab.2020.200127
74. Liu D, Li B, Shi X, et al. Cross-platform genomic identification and clinical validation of breast cancer diagnostic biomarkers. Aging. 2021;13(3):4258–4273. doi:10.18632/aging.202388
75. Kim J, Park S, Hwang D, Kim S, Lee H. Diagnostic value of circulating miR-202 in early-stage breast cancer in South Korea. Medicina. 2020;56(7):340. doi:10.3390/medicina56070340
76. Xin Y, Wang X, Meng K, Ni C, Lv Z, Guan D. Identification of exosomal miR-455-5p and miR-1255a as therapeutic targets for breast cancer. Biosci Rep. 2020;40(1):BSR20190303. doi:10.1042/BSR20190303
77. Zhang Z, Zhang L, Yu G, et al. Exosomal miR-1246 and miR-155 as predictive and prognostic biomarkers for trastuzumab-based therapy resistance in HER2-positive breast cancer. Cancer Chemother Pharmacol. 2020;86(6):761–772. doi:10.1007/s00280-020-04168-z
78. Li XJ, Ren ZJ, Tang JH, Yu Q. Exosomal MicroRNA MiR-1246 promotes cell proliferation, invasion and drug resistance by targeting CCNG2 in breast cancer. Cell Physiol Biochem. 2018;44(5):1741–1748. doi:10.1159/000485780
79. Yoshikawa M, Iinuma H, Umemoto Y, Yanagisawa T, Matsumoto A, Jinno H. Exosome-encapsulated microRNA-223-3p as a minimally invasive biomarker for the early detection of invasive breast cancer. Oncol Lett. 2018;15(6):9584–9592. doi:10.3892/ol.2018.8457
80. Asadirad A, Khodadadi A, Talaiezadeh A, Shohan M, Rashno M, Joudaki N. Evaluation of miRNA-21-5p and miRNA-10b-5p levels in serum-derived exosomes of breast cancer patients in different grades. Mol Cell Probes. 2022;64:101831. doi:10.1016/j.mcp.2022.101831
81. Curtaz CJ, Reifschläger L, Strähle L, et al. Analysis of microRNAs in exosomes of breast cancer patients in search of molecular prognostic factors in brain metastases. Int J Mol Sci. 2022;23(7):3683. doi:10.3390/ijms23073683
82. Li D, Wang J, Ma LJ, et al. Identification of serum exosomal miR-148a as a novel prognostic biomarker for breast cancer. Eur Rev Med Pharmacol Sci. 2020;24(13):7303–7309. doi:10.26355/eurrev_202007_21889
83. Liu M, Mo F, Song X, et al.. Exosomal hsa-miR-21-5p is a biomarker for breast cancer diagnosis. Peer J. 2021;9:e12147.
84. Ando W, Kikuchi K, Uematsu T, et al. Novel breast cancer screening: combined expression of miR-21 and MMP-1 in urinary exosomes detects 95% of breast cancer without metastasis. Sci Rep. 2019;9(1):13595. doi:10.1038/s41598-019-50084-5
85. Inubushi S, Kawaguchi H, Mizumoto S, et al. Oncogenic miRNAs identified in tear exosomes from metastatic breast cancer patients. Anticancer Res. 2020;40(6):3091–3096. doi:10.21873/anticanres.14290
86. Bao S, Hu T, Liu J, et al. Genomic instability-derived plasma extracellular vesicle-microRNA signature as a minimally invasive predictor of risk and unfavorable prognosis in breast cancer. J Nanobiotechnology. 2021;19(1):22. doi:10.1186/s12951-020-00767-3
87. Hannafon BN, Trigoso YD, Calloway CL, et al. Plasma exosome microRNAs are indicative of breast cancer. Breast Cancer Res. 2016;18(1):90. doi:10.1186/s13058-016-0753-x
88. Li S, Zhang M, Xu F, Wang Y, Leng D. Detection significance of miR-3662, miR-146a, and miR-1290 in serum exosomes of breast cancer patients. J Cancer Res Ther. 2021;17(3):749–755. doi:10.4103/jcrt.jcrt_280_21
89. Rodríguez-Martínez A, de Miguel-Pérez D, Ortega FG, et al. Exosomal miRNA profile as complementary tool in the diagnostic and prediction of treatment response in localized breast cancer under neoadjuvant chemotherapy. Breast Cancer Res. 2019;21(1):21. doi:10.1186/s13058-019-1109-0
90. Muluhngwi P, Klinge CM. Identification of miRNAs as biomarkers for acquired endocrine resistance in breast cancer. Mol Cell Endocrinol. 2017;456:76–86. doi:10.1016/j.mce.2017.02.004
91. Ryspayeva D, Halytskiy V, Kobyliak N, et al. Response to neoadjuvant chemotherapy in breast cancer: do microRNAs matter? Discov Oncol. 2022;13(1):43. doi:10.1007/s12672-022-00507-z
92. Guyon N, Garnier D, Briand J, et al. Anti-PD1 therapy induces lymphocyte-derived exosomal miRNA-4315 release inhibiting Bim-mediated apoptosis of tumor cells. Cell Death Dis. 2020;11(12):1048. doi:10.1038/s41419-020-03224-z
93. Yang Q, Zhao S, Shi Z, et al. Chemotherapy-elicited exosomal miR-378a-3p and miR-378d promote breast cancer stemness and chemoresistance via the activation of EZH2/STAT3 signaling. J Exp Clin Cancer Res. 2021;40(1):120. doi:10.1186/s13046-021-01901-1
94. Li Y, Liang Y, Sang Y, et al. MiR-770 suppresses the chemo-resistance and metastasis of triple negative breast cancer via direct targeting of STMN1. Cell Death Dis. 2018;9(1):14. doi:10.1038/s41419-017-0030-7
95. Luo L, Zhang X, Rousuli Y, Aini A, Liman R. Exosome-mediated transfer of miR-3613-5Pp enhances doxorubicin resistance by suppression of pten expression in breast cancer cells. J Oncol. 2022;2022:9494910. doi:10.1155/2022/9494910
96. Andreeva OE, Sorokin DV, Mikhaevich EI, et al. Towards unravelling the role of ERα-targeting miRNAs in the exosome-mediated transferring of the hormone resistance. Molecules. 2021;26(21):6661. doi:10.3390/molecules26216661
97. Wei Y, Lai X, Yu S, et al. Exosomal miR-221/222 enhances tamoxifen resistance in recipient ER-positive breast cancer cells. Breast Cancer Res Treat. 2014;147(2):423–431. doi:10.1007/s10549-014-3037-0
98. Liu J, Zhu S, Tang W, Huang Q, Mei Y, Yang H. Exosomes from tamoxifen-resistant breast cancer cells transmit drug resistance partly by delivering miR-9-5p. Cancer Cell Int. 2021;21(1):55. doi:10.1186/s12935-020-01659-0
99. Yu S, Zhou Y, Niu L, Qiao Y, Yan Y. Mesenchymal stem cell-derived exosome mir-342-3p inhibits metastasis and chemo-resistance of breast cancer through regulating ID4. Genes Genomics. 2022;44(5):539–550. doi:10.1007/s13258-021-01200-1
100. Liu Q, Zhang J, Liu Y, Peng H, Wu Y. Extracellular vesicles extracted from bone marrow mesenchymal stem cells carrying MicroRNA-342-3p inhibit the INHBA/IL13Rα2 axis to suppress the growth and metastasis of breast cancer. Transl Oncol. 2022;18:101333. doi:10.1016/j.tranon.2021.101333
101. Yang Z, Xu B, Wu S, et al. Exosomal microRNA-551b-3p from bone marrow-derived mesenchymal stromal cells inhibits breast cancer progression via regulating TRIM31/Akt signaling. Hum Cell. 2022;35(6):1797–1812. doi:10.1007/s13577-022-00753-x
102. Jia Z, Zhu H, Sun H, et al. Adipose mesenchymal stem cell-derived exosomal microrna-1236 reduces resistance of breast cancer cells to cisplatin by suppressing slc9a1 and the wnt/β-catenin signaling. Cancer Manag Res. 2020;12:8733–8744. doi:10.2147/CMAR.S270200
103. Du L, Tao X, Shen X. Human umbilical cord mesenchymal stem cell-derived exosomes inhibit migration and invasion of breast cancer cells via miR-21-5p/ZNF367 pathway. Breast Cancer. 2021;28(4):829–837. doi:10.1007/s12282-021-01218-z
104. Khazaei-Poul Y, Shojaei S, Koochaki A, Ghanbarian H, Mohammadi-Yeganeh S. Evaluating the influence of human umbilical cord mesenchymal stem cells-derived exosomes loaded with miR-3182 on metastatic performance of triple negative breast cancer cells. Life Sci. 2021;286:120015. doi:10.1016/j.lfs.2021.120015
105. Yuan L, Liu Y, Qu Y, Liu L, Li H. Exosomes derived from MicroRNA-148b-3p-overexpressing human umbilical cord mesenchymal stem cells restrain breast cancer progression. Front Oncol. 2019;9:1076. doi:10.3389/fonc.2019.01076
106. Sheykhhasan M, Kalhor N, Sheikholeslami A, Dolati M, Amini E, Fazaeli H. Exosomes of mesenchymal stem cells as a proper vehicle for transfecting miR-145 into the breast cancer cell line and its effect on metastasis. Biomed Res Int. 2021;2021:5516078. doi:10.1155/2021/5516078
107. Zhou Y, Chen F, Xie X, et al. Tumor-derived exosome promotes metastasis via altering its phenotype and inclusions. J Cancer. 2021;12(14):4240–4246. doi:10.7150/jca.48043
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.