")
Back to Journals » Infection and Drug Resistance » Volume 17
Roles of Nucleolar Factor RCL1 in Itraconazole Resistance of Clinical Candida albicans Under Different Stress Conditions
Authors Yang J, Ma Y, Li B, Xi Z, Zhang L, Wang Y, Feng W
Received 17 July 2023
Accepted for publication 2 February 2024
Published 27 February 2024 Volume 2024:17 Pages 769—777
DOI https://doi.org/10.2147/IDR.S431024
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Sandip Patil
Jing Yang, Yan Ma, Bo Li, Zhiqin Xi, Li Zhang, Yuxi Wang, Wenli Feng
Department of Dermatovenereology, The Second Hospital of Shanxi Medical University, Taiyuan, Shanxi, 030001, People’s Republic of China
Correspondence: Wenli Feng; Jing Yang, Department of Dermatovenereology, The Second Hospital of Shanxi Medical University, No. 382, Wuyi Road, Taiyuan, Shanxi, 030001, People’s Republic of China, Tel +86-351-3365410, Email [email protected]; [email protected]
Purpose: RNA terminal phosphate cyclase like 1 (RCL1) undergoes overexpression during the immune response of Candida albicans following drug treatment. This study aims to investigate the expression levels of RCL1 in C. albicans under various stress conditions.
Methods: Fifteen itraconazole (ITR)-resistant strains of clinical C. albicans, and one standard strain were employed for RCL1 sequencing, and mutations in RCL1 were analyzed. Subsequently, 14 out of the 15 ITR-resistant clinical strains and 14 clinical strains sensitive to ITR, fluconazole (FCA) as well as voriconazole (VRC) were cultured under diverse conditions. The expression of RCL1 ITR-resistant and sensitive C. albicans was then assessed using real-time quantitative PCR (RT-qPCR) assays.
Results: Compared to the standard strain, three missense mutations (C6A, G10A, and A11T) were identified in the RCL1 gene of ITR-resistant C. albicans through successful forward sequencing. Additionally, using successful reverse sequencing, one synonymous mutation (C1T) and four missense mutations (C1T, A3T, A7G, and T8G) were found in the RCL1 gene of ITR-resistant C. albicans. RCL1 expression was significantly higher in ITR-resistant C. albicans than in sensitive strains under standard conditions (37°C, 0.03% CO2, pH 4.0). Low temperature (25°C) increased RCL1 expression in sensitive C. albicans while decreasing it in ITR-resistant strains. Elevated CO2 concentrations (5% CO2) had a negligible effect on RCL1 expression in sensitive C. albicans, but effectively reduced RCL1 level in ITR-resistant strains. Furthermore, a medium with a pH of 7 decreased the expression of RCL1 in both resistant and sensitive C. albicans.
Conclusion: This study demonstrated that RCL1 mutations in ITR-resistant C. albicans, and variations in culture conditions significantly influence RCL1 expression in both ITR-resistant and sensitive C. albicans, thereby inducing alterations in the dimorphism of C. albicans.
Keywords: Candida albicans, itraconazole resistance, RCL1, culture conditions
Introduction
Invasive Candidiasis (IC) is a prevalent fungal infection associated with a high mortality rate of 70%. The manifestations of IC range from candidemia to fulminant sepsis, exhibiting varying degrees of severity.1,2 Several factors, such as prolonged non-standard and high-dose application of immunosuppressants and glucocorticoids, an increase in acquired human immunodeficiency virus (HIV) infections, and the use of various catheters, stents, and other medical instruments, contribute to the annual rise in IC incidence.3 IC typically involves the invasion of the bloodstream by candida fungi, including species such as Candida albicans, Candida parapsilosis, and Candida tropicalis. C. albicans remains the primary pathogen causing IC.1,2
C. albicans, a conditionally pathogenic fungus, can co-exist in healthy individuals as a part of the normal microbiota without causing disease.4 It commonly resides on mucosal surfaces, like the oral, vaginal, and gastrointestinal mucosa.5 However, under certain conditions, C. albicans can cause a spectrum of diseases, ranging from superficial mucous membranes infections to deep-seated systemic invasions, potentially leading to life-threatening deep fungal infections upon entering the bloodstream.6 Previous studies have indicated that a variety of factors, such as the transformation of yeast to mycelial morphology, phenotypic transformation, biofilm formation, and the expression of cell surface adhesins, enhance the pathogenicity of C. albicans.7–9 Antifungal drugs, including itraconazole (ITR), fluconazole (FCA), and voriconazole (VRC), are effective against C. albicans. However, the abuse and prolonged use of these drugs have led to the emergence of drug-resistant strains, posing a significant challenge to the treatment of C. albicans infections.10 Therefore, it is imperative to elucidate the mechanisms of resistance in C. albicans to improve the prognosis of patients.
Ribosomes are arguably the most critical macromolecular machines, tasked with carrying out protein synthesis in cells. Ribosome biogenesis stands out as one of the most crucial and energy-consuming processes within cells.11 Mature ribosomes consist of chemically modified components, including methylation of the ribose sugars at the C2-position, isomerization of uridines to pseudouridines, and various base modifications in both ribosomal RNA (rRNA) and ribosomal protein (RP).11 In eukaryotes, ribosomes are composed of a small subunit (40Ss) and a large subunit (60S). The process of ribosome biogenesis is complex and subject to strict regulation to ensure the correct production of ribosomes under various environmental and metabolic changes.12 Ribosome biogenesis begins with the transcription of rRNA in the nucleolus, and then the transcribed rRNA undergoes complicated folding, modification, processing, and binding to RP. During this process, precursor rRNA (pre-rRNA) is exported from the nucleus to the cytoplasm, eventually maturing into ribosomes with translational activity.13,14 Over 200 ribosome assembly factors (AFs) and 80 small nucleolar RNAs (snoRNAs) are associated with ribosome biogenesis. Among them, GTPase BMS1 (BMS1) is the sole known GTPase required for the biosynthesis of the 40S ribosome subunit, interacting with RNA terminal phosphate cyclase like 1 (RCL1). RCL1, an RNA 3’-terminal phosphate cyclase, plays a crucial role in the biogenesis of 18S rRNA and pre-rRNA in eukaryotes.15–17 Identified as a stress gene, RCL1 responds more rapidly to drugs compared to other ribosomal genes.18 In our previous studies, we uncovered that RCL1 is overexpressed in C. albicans during the immune response to drug treatment.17 Differential expression analysis and protein–protein interaction (PPI) network analysis of the CAF-21 expression data file (GSE65396) in C. albicans before and after treatment with the macrocyclic compound (RF59) in the GEO database revealed a notable increase in RCL1 and other ribosome-related genes following drug administration.17 However, the physiological significance of RCL1 in Candidal cells, particularly in drug-resistant strains and under different stress conditions, as well as how changes in RCL1 expression may impact the organism and/or the host in an infective situation remain unknown.
The conditions typically employed for the cultivation of C. albicans involve a temperature of 37°C, a CO2 concentration of 0.03% CO2 in the air, and a pH level of 4. It is noteworthy that the female vagina is characterized by weak acidity, with a pH range of 3.8–4.4 under normal circumstances. These specified conditions are considered standard and are henceforth referred to as common or normal conditions in this study. The primary objective of this investigation is to assess the impact of RCL1 on drug resistance in C. albicans through an examination of gene expression patterns. Concurrently, C. albicans was subjected to cultivation under diverse conditions to evaluate RCL1 expression and elucidate the role of RCL1 in the face of varying stressors.
Materials and Methods
Experimental Strains
The 15 strains of clinical ITR-resistant C. albicans and 1 strain of standard C. albicans were utilized for the Sanger sequencing. The quantitative real-time polymerase chain reaction (qRT-PCR) assays were conducted on 14 of the aforementioned 15 strains of clinical ITR-resistant C. albicans (the same strains used in sequencing), 14 strains of clinical C. albicans sensitive to ITR, FCA, and VRC (sensitive C. albicans), and 1 strain of standard C. albicans (the same strains used in sequencing). The standard strain (ATCC11006) was procured from the Center for Mycology and Mycosis Research, Peking University Health Science Center. All the clinical strains were isolated, identified, and preserved at the Fungal Laboratory, Department of Dermatology and Venereology, Second Hospital of Shanxi Medical University, China. This study received approval from the Ethics Committee of the Second Hospital of Shanxi Medical University.
DNA Extraction, PCR Amplification, and RCL1 Sequencing
The strains of C. albicans were cultured in Yeast Extract Peptone Dextrose (YPD) Liquid Medium (AOBOX BIOTECHNOLOGY, Beijing, China) at 37°C with a rotational speed of 200 rpm/min. After 24 hours of culture, total DNA from C. albicans was extracted using a Yeast DNA Extraction kit (Shanghai Sangong Biotechnology Co., LTD, Shanghai, China) following the manufacturer’s protocols. The concentration (OD260) and purity (OD260/280) of the isolated DNA were determined using a microplate reader (BK-9622, BIOBASE, Shandong, China). To obtain the full-length sequence of the RCL1 gene, the upstream and downstream primers for RCL1 (RCL1-Forward-5’-TGGCATCACCTTAGTCGCTG-3’, RCL1-Reverse-5’-GTAAACTCCCAGCCGAACCA-3’) were designed and synthesized by Shanghai Sangong Biotechnology Co., LTD. (Shanghai, China). Subsequently, the RCL1 sequence was amplified on a PCR system (ProFlex, Applied Biosystems, California, USA) using 2×Hieff® Ultra-Rapid HotStart PCR Master Mix (with Dye) (10157ES03, YESEN, Shanghai, China) following the manufacturer’s instructions. The PCR products, with a length of 77 bp, were purified by 1% agarose gel electrophoresis, and sent to Shanghai Sangong Biotechnology Co., Ltd. for sequencing. Mutation sites in the RCL1 sequencing were observed using SnapGene 4.1.8 software (Insightful Science, California, USA).
Culture of C. albicans Under Diverse Stress Conditions
Culture of C. albicans Under Different Temperatures
Sterile inoculation rings were used to inoculate 14 strains of clinical ITR-resistant C. albicans and 14 strains of clinical sensitive C. albicans onto YPD solid medium (pH = 4, YPDA Medium, PM2011, Coolaber, Beijing, China). The strains were then incubated in either a 25°C or 37°C incubator (SPX-250B-Z, Shanghai Boxun Industrial Co., LTD, Shanghai, China) with 0.03% CO2 for 48 hours. Subsequently, a single colony was selected and transferred into YPD liquid medium (pH = 4), shaken, and cultured in a 25°C or 37°C atmosphere with 0.03% CO2 for an additional 24 hours.
Culture of C. albicans Under Different CO2 Concentrations
The sterile inoculation rings were employed to introduce 14 strains of clinical ITR-resistant C. albicans and 14 strains of clinical sensitive C. albicans onto YPD solid medium (adjusted to pH = 4). Subsequently, the strains were incubated in a 37°C incubator, with 5% or 0.03% CO2, for a period of 48 hour. Following incubation, a single colony was selected and transferred into YPD liquid medium (pH = 4), agitated, and cultured in a 37°C environment, under 5% or 0.03% CO2 for an additional 24 hours.
Culture of C. albicans Under Different pH Value
Sterile inoculation rings were employed to inoculate 14 strains each of clinical ITR-resistant C. albicans and clinical sensitive C. albicans onto YPD solid medium at pH = 7 or pH = 4. Subsequently, the strains were incubated in a 37°C incubator with 0.03% CO2 for 48 hours. Following this, a single colony was selected and transferred to YPD liquid medium (at pH = 7 or pH = 4), where it was shaken and cultured under a 37°C atmosphere with 0.03% CO2 for an additional 24 hours.
Real-Time Quantitative PCR (RT-qPCR)
The total RNA from C. albicans cultured under various conditions was extracted using the Spin Column Yeast Total RNA Purification Kit (B518657, Shanghai Sangong Biotechnology Co., LTD, Shanghai, China) following the manufacturer’s protocols. The MightyScript First-Strand cDNA Synthesis Master Mix (B639251, Shanghai Sangong Biotechnology Co., LTD, Shanghai, China) was employed for reverse transcription of the total RNA into complementary DNA (cDNA) in accordance with the instructions of the manufacturer. Subsequently, qRT-PCR analysis was conducted using 2×SYBR Green PCR Master Mix (SR1110, Solarbio, Beijing, China) as outlined previously.19 The transcription level of RCL1 was normalized to ACTB (β-actin), and the relative expression of RCL1 was calculated using the 2−ΔΔCt method. The primers used in the qRT-PCR assay were as follows: RCL1-Forward-5’-GCCAACAGGATGCGAGGTTA-3’, RCL1-Reverse-5’- TCAGCGACTAAGGTGATGCC-3’; ACTB-Forward-5’-TGTTGACCGAAGCTCCAATGA-3’, ACTB-Reverse-5’-ACCGGTGGTTCTACCAGAAG-3’.
Statistical Analysis
All data were analyzed using SPSS software (version 26.0, SPSS, Inc., Chicago, IL, USA) and were presented as mean ± standard deviation (SD). The differences between the two groups were assessed using the Student’s t-test. *P < 0.05 indicated a statistical difference.
Results
RCL1 Sequencing Analysis
The RCL1 gene of 16 strains of C. albicans was subjected to bidirectional sequencing in this study. The forward sequencing revealed successful sequencing of six C. albicans strains, including the standard strain ATCC11006 and the ITR-resistant strains SY13, SY17, SY18, SY32, and SY35. RCL1 gene sequences of the five ITR-resistant strains, which were successfully sequenced in the forward direction, were compared to those of the standard strain ATCC11006. Table 1 illustrates that a total of three missense mutations were identified in ITR-resistant C. albicans, namely C6A, G10A, and A11T.
 |
Table 1 Base Mutation Sites and Amino Acid Mutation for RCL1 in Candida albicans Strains Through Forward Sequencing |
Results from reverse sequencing demonstrated the successful sequencing of all 16 strains of C. albicans. Comparative analysis with the RCL1 gene of the standard strain ATCC11006 revealed one synonymous mutation (C1T) and four missense mutations (C1T, A3T, A7G, and T8G) in the 15 ITR-resistant strains that were successfully sequenced in the reverse direction (Table 2).
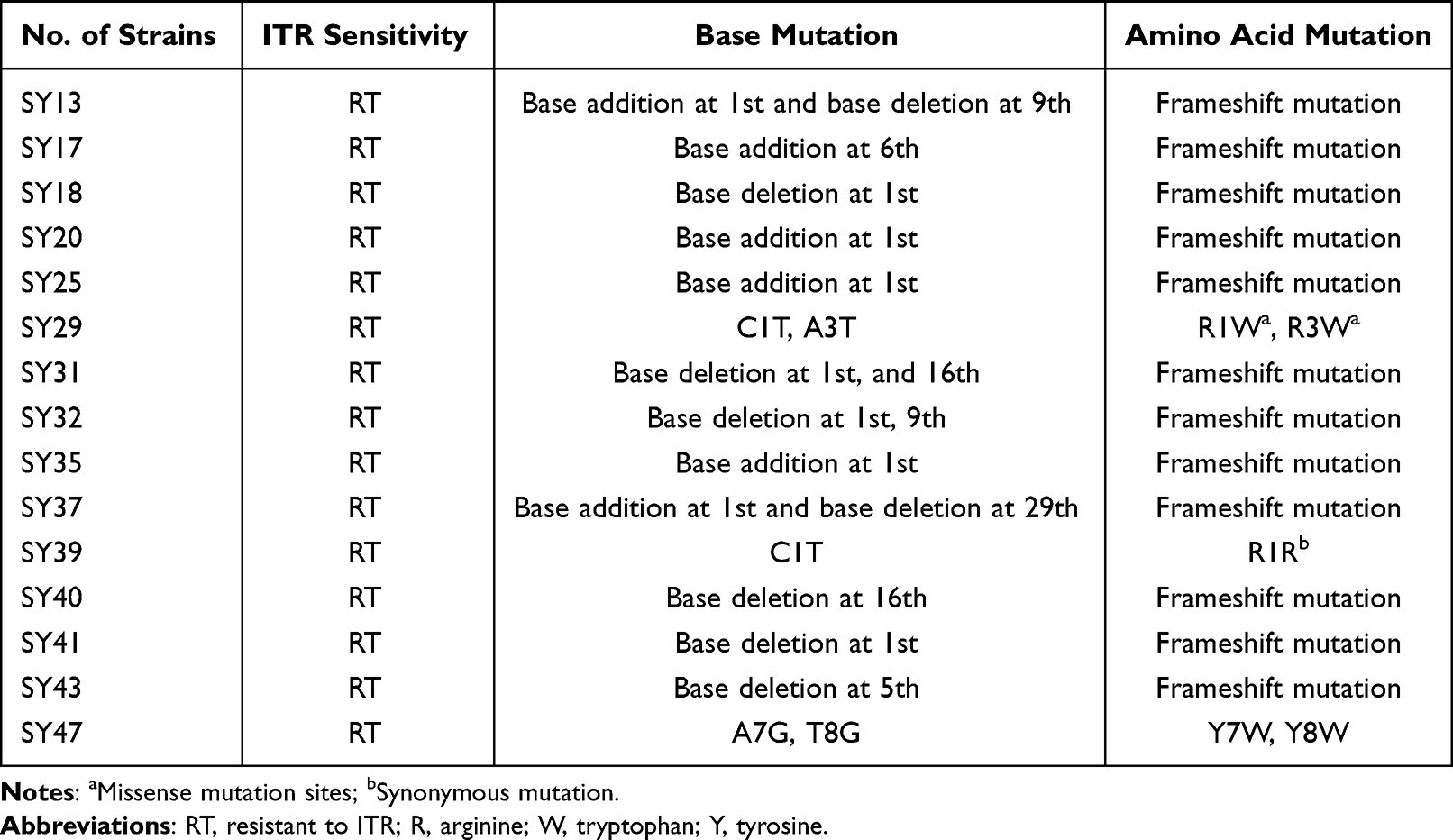 |
Table 2 Base Mutation Sites and Amino Acid Mutation for RCL1 in Candida albicans Strains Through Reverse Sequencing |
Expression of RCL1 in the Sensitive and ITR-Resistant C. albicans
Subsequently, we determined the expression levels of RCL1 in both susceptible and ITR-resistant strains of C. albicans. The data revealed a significant upregulation of RCL1 expression in ITR-resistant C. albicans compared to the susceptible strain under normal conditions (37°C, 0.03% CO2, pH 4.0) (P < 0.05, Figure 1A). At a lower temperature (25°C, 0.03% CO2, pH = 4.0), the expression levels of RCL1 in both sensitive and ITR-resistant C. albicans were comparable, with no statistically significant difference (P > 0.05, Figure 1B). Although the expression of RCL1 in ITR-resistant C. albicans was slightly elevated with increased CO2 concentration (37°C, 5% CO2, pH 4.0), the difference did not reach statistical significance (P > 0.05, Figure 1C). Intriguingly, the expression of RCL1 in ITR-resistant C. albicans was notably higher than in sensitive strains when the pH of the culture conditions was increased to 7.0 (37°C, 0.03% CO2, pH 7.0) (P < 0.05, Figure 1D). These findings suggest that RCL1 expression varies between sensitive and resistant C. albicans strains under different culture conditions.
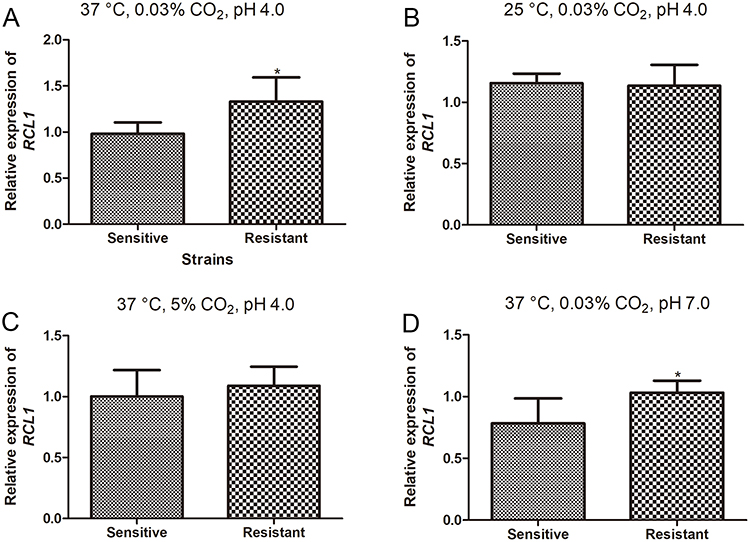 |
Figure 1 Expression of RCL1 in the sensitive and resistant C. albicans. (A) RCL1 expression in sensitive and ITR-resistant C. albicans in normal condition (37°C, 5% CO2, pH = 4.0). N = 14. (B) RCL1 expression in sensitive and ITR-resistant C. albicans at low temperature (25°C, 0.03% CO2, pH = 4.0). N = 14. (C) The level of RCL1 in sensitive and ITR-resistant C. albicans at high CO2 concentration (37°C, 5% CO2, pH = 4.0). N = 14. (D) The expression of RCL1 in sensitive and ITR-sensitive C. albicans in neutral conditions (37°C, 0.03% CO2, pH = 7.0). N = 14. *P < 005, compared with the sensitive strains. |
Expression of RCL1 in C. albicans Under Diverse Temperature Conditions
The expression levels of RCL1 in C. albicans under various temperature conditions were determined using qRT-PCR. In the sensitive strains of C. albicans, RCL1 expression at 25°C was significantly higher than that at 37°C (P < 0.05, Figure 2A). In contrast, the level of RCL1 was remarkably lower at 25°C compared to 37°C in the ITR-resistant strains of C. albicans (P < 0.05, Figure 2B). These data indicate that temperature could influence the expression of RCL1 in both the sensitive and resistant strains of C. albicans.
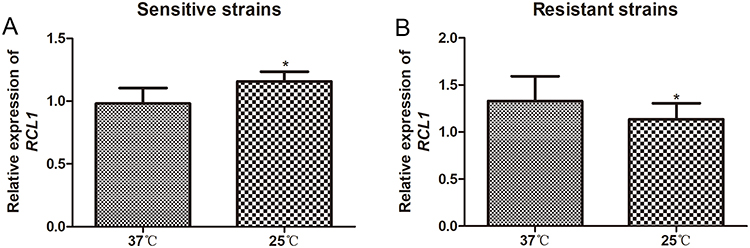 |
Figure 2 Expression of RCL1 in C. albicans under diverse temperature conditions. (A) RCL1 expression in sensitive C. albicans at 37°C or 25°C. N = 14. (B) RCL1 expression in ITR-resistant C. albicans at 37°C or 25°C. N = 14. *P < 005, compared with the strains cultured at 37°C. |
Expression of RCL1 in C. albicans Under Diverse CO2 Conditions
The RCL1 expression in C. albicans was investigated under different CO2 conditions. Figure 3A illustrates that an increased CO2 concentration (5% CO2) had minimal impact on RCL1 expression in the sensitive strains of C. albicans (P > 0.05). However, in ITR-resistant C. albicans, the RCL1 level significantly decreased under 5% CO2 conditions compared to 0.03% CO2 conditions (P < 0.05, Figure 3B). These results demonstrate that CO2 concentration influences the expression of RCL1 in the resistant C. albicans but not in the sensitive C. albicans.
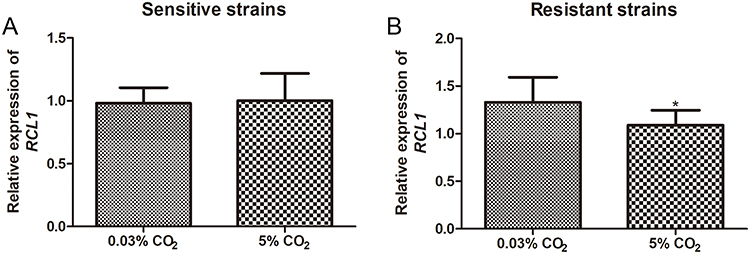 |
Figure 3 Expression of RCL1 in C. albicans under diverse CO2 condition. (A) The RCL1 level in sensitive C. albicans under 0.03% or 5% CO2 concentration. N = 14. (B) The RCL1 expression in ITR-resistant C. albicans under 0.03% or 5% CO2 concentration. N = 14. *P < 005, compared with the strains cultured with 0.03% CO2. |
Expression of RCL1 in C. albicans Under Diverse pH Conditions
Further analyses revealed that an elevated pH value (pH = 7.0) significantly diminished the expression levels of RCL1 in both the sensitive and ITR-resistant strains of C. albicans (P < 0.05, Figure 4A and B). These findings suggest that distinct pH values modulate RCL1 expression in both the sensitive and resistant strains of C. albicans.
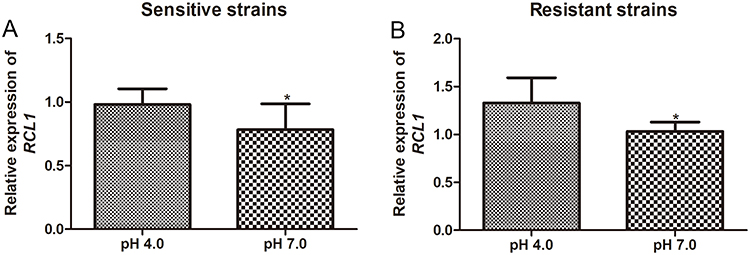 |
Figure 4 Expression of RCL1 in C. albicans under different pH condition. (A) The RCL1 expression in sensitive C. albicans under pH = 4 or pH = 7 condition. N = 14. (B) The RCL1 expression in ITR-resistant C. albicans under pH = 4 or pH = 7 condition. N = 14. *P < 005, compared with the strains cultured at pH = 4. |
Discussion
C. albicans is a common human pathogen, and IC caused by this fungus poses a threat to the lives and health of patients.20 RCL1 is a phosphate cyclase closely associated with the processing of rRNA.21 Growing evidence indicates that RCL1 can bind to BMS1, a typical GTPase involved in the synthesis of 18S rRNA.22 It has been reported that the RCL1–BMS1 complex plays a crucial role in processing yeast pre-rRNAs.23 Moreover, our previous study demonstrated a significant increase in RCL1 expression in C. albicans upon treatment with RF59, a macrocyclic compound.17 These findings suggest that RCL1 contributes to the improved the physiological status of C. albicans. However, the expression 1 changes of RCL in C. albicans have not been thoroughly investigated. In this study, we conducted RCL1 sequencing on 16 C. albicans isolates, revealing mutation sites in the RCL1 gene in ITR-resistant C. albicans compared to the standard strain. Specifically, among the five successfully sequenced ITR-resistant strains, three missense mutations were identified in the RCL1 sequence: C6A, G10A, and A11T. Reverse sequencing of 15 successfully sequenced ITR-resistant strains unveiled one synonymous mutation (C1T) and four missense mutations (C1T, A3T, A7G, and T8G) in the RCL1 sequencing. These results suggest that reverse sequencing detected more mutations than forward sequencing, potentially due to base mutations.24 The full length of the RCL1 gene in our study was 77 bp, which may not be sufficient, increasing the likelihood of base mutations.25 Additionally, mutations were only identified in ITR-resistant C. albicans, indicating a potential association between RCL1 mutations and ITR resistance. However, further studies are needed to validate the correlation between RCL1 mutations and ITR resistance.
Stress responses involve various mechanisms that enable microorganisms to survive in adverse and fluctuating environments.9 Microorganisms typically possess intricate “circuits” capable of responding to diverse stresses, exemplified by the response of Candida to antifungal drugs.26 There is a growing body of evidence indicating that C. albicans exhibits different cell morphologies under varying conditions, including “white, opaque, gray”, “yeast state”, and “mycelium state”. These cell morphologies could be converted to each other, which endows C. albicans with diverse pathogenic abilities.27 Taken together, we speculated that stress may exert an essential role in the drug resistance of C. albicans.
Additionally, stress often leads to the reorganization of the nuclear structure. The nucleolus serves as a stress sensor, coordinating the stress response.28 Recently, accumulating evidence has demonstrated that damage to the integrity of the nucleolar system and disruption of ribosome generation can activate the nucleolar stress signaling pathway, ultimately compromising its role in cell growth, apoptosis, and DNA regulation.29 The viral stress gene RCL1 encoding a nucleolar protein is essential for 18S rRNA in ribosome biosynthesis.15,17 Moreover, Enjalbert et al verified that RCL1 is the main regulatory factor in C. albicans under various stress conditions, including oxidation and heavy metal stress.18 As C. albicans is well known for its characteristic dimorphism, existing as both a yeast and mycelium form, different environmental stimuli, such as a neutral pH value, 25°C and 5% CO2 can induce changes in its dimorphism of C. albicans.30,31 The transition from yeast to mycelia is a crucial virulence factor in C. albicans.32 However, the impact of different culture environments (temperature, CO2 concentration, and pH value) on the expression of RCL1 in C. albicans remains unclear.
In this study, we investigated the expression of RCL1 in C. albicans under normal conditions (37°C, 0.03% CO2, pH 4.0). We observed higher RCL1 expression in ITR-resistant C. albicans compared to sensitive C. albicans. The decrease in temperature to 25°C and the increase in CO2 concentration to 5% CO2 both led to similar RCL1 expression levels in both sensitive and resistant strains of C. albicans. Notably, an up-regulated pH value (pH = 7.0) resulted in increased RCL1 expression specifically in the ITR-resistant C. albicans compared to the sensitive C. albicans. These findings suggest that RCL1 expression is inherently higher in resistant C. albicans under normal conditions. Interestingly, alterations in temperature and CO2 concentration effectively eliminated the differences in RCL1 expression levels between sensitive and resistant C. albicans.
Subsequent experiments revealed that, when compared to 37°C, a temperature of 25°C led to a significant increase in the RCL1 level in sensitive C. albicans, while concurrently decreasing it in ITR-resistant C. albicans. Conversely, exposure to 5% CO2 showed no significant effect on RCL1 content in sensitive C. albicans, in contrast to the notable reduction observed in ITR-resistant strains, when compared to 0.03% CO2. Furthermore, an increase in pH was found to up-regulate RCL1 expression in both sensitive and resistant C. albicans.
This study had certain limitations. A significant constraint is the absence of cross-condition testing, which is essential for establishing the effects of stressors (temperature, CO2, and pH) on the expression of RCL1. The findings indicate a notable increase in RCL1 expression in ITR-resistant C. albicans was notably higher compared to sensitive strains, suggesting a potential involvement of RCL1 in ITR resistance. However, this conclusion warrants further exploration in future studies. Furthermore, additional experiments will be conducted to investigate whether various conditions (temperature, CO2 concentration, and pH) regulate the morphological transformation and pathogenicity of C. albicans by influencing RCL1 expression.
Conclusion
In summary, this study unveils that ITR-resistant C. albicans harbors several mutation sites in the RCL1 gene. Forward sequencing identified three missense mutations (C6A, G10A, and A11T) in the five strains of ITR-resistant C. albicans, while reverse sequencing revealed one synonym mutation (C1T) and four missense mutations (C1T, A3T, A7G, and T8G) among the 15 strains of ITR-resistant C. albicans. Different stress conditions (25°C, 5% CO2, pH = 7.0) exert distinct regulatory effects on RCL1 expression in both sensitive and resistant C. albicans strains. These findings suggest that C. albicans may activate its stress response through the RCL1 protein, providing a novel perspective for understanding the molecular targets of potential antifungal drugs for treating patients with invasive candidiasis.
Data Sharing Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Funding
This study was supported by the General Project of National Natural Science Foundation of China (Project number: 82072262), Scientific and Technological Activities funding program for Overseas Students of Shanxi Province (Project number: 20210030), Research Project Supported by Shanxi Scholarship Council of China (Project number: 2020–190), Natural Science Foundation of Shanxi Province (Project number: 202103021224416), and Shanxi Province colleges and universities excellent achievement cultivation project (Project number: 2019KJ025).
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Pappas PG, Lionakis MS, Arendrup MC, Ostrosky-Zeichner L, Kullberg BJ. Invasive candidiasis. Nat Rev Dis Primers. 2018;4:18026.
2. McCarty TP, White CM, Pappas PG. Candidemia and invasive candidiasis. Infect Dis Clin North Am. 2021;35(2):389–413. doi:10.1016/j.idc.2021.03.007
3. Hartnett KP, Jackson KA, Felsen C, et al. Bacterial and fungal infections in persons who inject drugs - Western New York, 2017. MMWR. 2019;68(26):583–586. doi:10.15585/mmwr.mm6826a2
4. Wu T, Cen L, Kaplan C, et al. Cellular components mediating coadherence of Candida albicans and Fusobacterium nucleatum. J Dent Res. 2015;94(10):1432–1438. doi:10.1177/0022034515593706
5. Lee Y, Puumala E, Robbins N, Cowen LE. Antifungal drug resistance: molecular mechanisms in Candida albicans and beyond. Chem Rev. 2021;121(6):3390–3411. doi:10.1021/acs.chemrev.0c00199
6. Kavanaugh NL, Zhang AQ, Nobile CJ, Johnson AD, Ribbeck K, Berman J. Mucins suppress virulence traits of Candida albicans. mBio. 2014;5(6):e01911. doi:10.1128/mBio.01911-14
7. Jacobsen ID, Wilson D, Wächtler B, Brunke S, Naglik JR, Hube B. Candida albicans dimorphism as a therapeutic target. Exp Rev Anti-Infective Ther. 2012;10(1):85–93. doi:10.1586/eri.11.152
8. Nicholls S, MacCallum DM, Kaffarnik FA, Selway L, Peck SC, Brown AJ. Activation of the heat shock transcription factor Hsf1 is essential for the full virulence of the fungal pathogen Candida albicans. Fungal Genet Biol. 2011;48(3):297–305.
9. Mayer FL, Wilson D, Hube B. Candida albicans pathogenicity mechanisms. Virulence. 2013;4(2):119–128. doi:10.4161/viru.22913
10. Feng W, Yang J, Pan Y, et al. The correlation of virulence, pathogenicity, and itraconazole resistance with SAP activity in Candida albicans strains. Can J Microbiol. 2016;62(2):173–178. doi:10.1139/cjm-2015-0457
11. Ojha S, Malla S, Lyons SM, et al. snoRNPs: functions in ribosome biogenesis. Biomolecules. 2020;10(5):783. doi:10.3390/biom10050783
12. de la Cruzde la Cruz J, Gómez-Herreros F, Rodríguez-Galán O, Begley V, De la Cruz muñoz-centeno M, Chávez S. Feedback regulation of ribosome assembly. Curr Genet. 2018;64(2):393–404. doi:10.1007/s00294-017-0764-x
13. Woolford JL Jr, Baserga SJ. Ribosome biogenesis in the yeast saccharomyces cerevisiae. Genetics. 2013;195(3):643–681. doi:10.1534/genetics.113.153197
14. Baßler J, Hurt E. Eukaryotic ribosome assembly. Annu Rev Biochem. 2019;88(1):281–306. doi:10.1146/annurev-biochem-013118-110817
15. Tanaka N, Smith P, Shuman S. Crystal structure of Rcl1, an essential component of the eukaryal pre-rRNA processosome implicated in 18s rRNA biogenesis. RNA. 2011;17(4):595–602. doi:10.1261/rna.2571811
16. Wang Y, Zhu Q, Huang L, et al. Interaction between Bms1 and Rcl1, two ribosome biogenesis factors, is evolutionally conserved in zebrafish and human. J Genet Genomics. 2016;43(7):467–469. doi:10.1016/j.jgg.2016.05.001
17. Yang J, Zhang W, Sun J, et al. Screening of potential genes contributing to the macrocycle drug resistance of C. albicans via microarray analysis. Mol Med Rep. 2017;16(5):7527–7533. doi:10.3892/mmr.2017.7562
18. Enjalbert B, Smith DA, Cornell MJ, et al. Role of the Hog1 stress-activated protein kinase in the global transcriptional response to stress in the fungal pathogen Candida albicans. Mol Biol Cell. 2006;17(2):1018–1032. doi:10.1091/mbc.e05-06-0501
19. Chen Y, Sun L. Inhibition of NEDD8 NEDDylation induced apoptosis in acute myeloid leukemia cells via p53 signaling pathway. Biosci Rep. 2022;42(8). doi:10.1042/BSR20220994
20. Candel FJ, Pazos Pacheco C, Ruiz-Camps I, et al. Update on management of invasive candidiasis. Rev Esp Quimioter. 2017;30(6):397–406.
21. Genschik P, Billy E, Swianiewicz M, Filipowicz W. The human RNA 3’-terminal phosphate cyclase is a member of a new family of proteins conserved in Eucarya, Bacteria and Archaea. EMBO J. 1997;16(10):2955–2967.
22. Horn DM, Mason SL, Karbstein K. Rcl1 protein, a novel nuclease for 18S ribosomal RNA production. J Biol Chem. 2011;286(39):34082–34087. doi:10.1074/jbc.M111.268649
23. Delprato A, Al Kadri Y, Pérébaskine N, et al. Crucial role of the Rcl1p-Bms1p interaction for yeast pre-ribosomal RNA processing. Nucleic Acids Res. 2014;42(15):10161–10172. doi:10.1093/nar/gku682
24. Feng W, Yang J, Ma Y, et al. The effects of secreted aspartyl proteinase inhibitor ritonavir on azoles-resistant strains of Candida albicans as well as regulatory role of SAP2 and ERG11. Immun Inflamm Dis. 2021;9(3):667–680. doi:10.1002/iid3.415
25. Feng W, Yang J, Ji Y, et al. Mrr2 mutations and upregulation are associated with increased fluconazole resistance in Candida albicans isolates from patients with vulvovaginal candidiasis. Lett Appl Microbiol. 2020;70(2):95–101. doi:10.1111/lam.13248
26. Cowen LE, Steinbach WJ. Stress, drugs, and evolution: the role of cellular signaling in fungal drug resistance. Eukaryotic Cell. 2008;7(5):747–764. doi:10.1128/EC.00041-08
27. Wang TX, Yang DD, Sun X, Zhang M, Su C, Y L. Dimorphism in Candida albicans: from commensal to pathogen (in Chinese). Mycosystema. 2020;39(11):2003–2013.
28. Boulon S, Westman BJ, Hutten S, Boisvert FM, Lamond AI. The nucleolus under stress. Molecular Cell. 2010;40(2):216–227. doi:10.1016/j.molcel.2010.09.024
29. Rubbi CP, Milner J. Disruption of the nucleolus mediates stabilization of p53 in response to DNA damage and other stresses. EMBO J. 2003;22(22):6068–6077. doi:10.1093/emboj/cdg579
30. Honorato L, de Araujo JFD, Ellis CC, et al. Extracellular vesicles regulate biofilm formation and yeast-to-hypha differentiation in Candida albicans. mBio. 2022;13(3):e0030122. doi:10.1128/mbio.00301-22
31. Chen H, Zhou X, Ren B, Cheng L. The regulation of hyphae growth in Candida albicans. Virulence. 2020;11(1):337–348. doi:10.1080/21505594.2020.1748930
32. Su C, Yu J, Lu Y. Hyphal development in Candida albicans from different cell states. Curr Genet. 2018;64(6):1239–1243. doi:10.1007/s00294-018-0845-5
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.