")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 20
Gut Microbiota Dysbiosis and Inflammation Dysfunction in Late-Life Depression: An Observational Cross-Sectional Analysis
Authors Chen Y, Le D, Xu J , Jin P, Zhang Y, Liao Z
Received 24 November 2023
Accepted for publication 17 February 2024
Published 27 February 2024 Volume 2024:20 Pages 399—414
DOI https://doi.org/10.2147/NDT.S449224
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yuping Ning
Yan Chen,1 Dansheng Le,1 Jiaxi Xu,2 Piaopiao Jin,3 Yuhan Zhang,4 Zhengluan Liao1
1Center for Rehabilitation Medicine, Department of Psychiatry, Zhejiang Provincial People’s Hospital (Affiliated People’s Hospital), Hangzhou Medical College, Hangzhou, Zhejiang, People’s Republic of China; 2Department of Psychiatry, Tongde Hospital of Zhejiang Province, Hangzhou, Zhejiang, People’s Republic of China; 3Department of Psychiatry, Yiwu Central Hospital, Jin Hu, Zhejiang, People’s Republic of China; 4The Second Clinical College of Zhejiang Chinese Medical University, Hangzhou, Zhejiang, People’s Republic of China
Correspondence: Zhengluan Liao, Center for Rehabilitation Medicine, Department of Psychiatry, Zhejiang Provincial People’s Hospital (Affiliated People’s Hospital), Hangzhou Medical College, No. 158 Shangtang Road, Hangzhou, Zhejiang, People’s Republic of China, Tel/Fax +86-571-87312143, Email [email protected]
Purpose: There are some challenges to diagnosis in the context of similar diagnostic criteria for late-life depression (LLD) and adult depression due to cognitive impairment and other clinical manifestations. The association between gut microbiota and inflammation remains unclear in LLD. We analyzed gut microbiota characteristics and serum inflammatory cytokines in individuals with LLD to explore the combined role of these two factors in potential biomarkers of LLD.
Methods: This was an observational cross-sectional study. Fecal samples and peripheral blood from 29 patients and 33 sex- and age-matched healthy controls (HCs) were collected to detect gut microbiota and 12 inflammatory factors. We analyzed differences in diversity and composition of gut microbiota and evaluated relations among gut microbiota, inflammatory factors, and neuropsychological scales. We extracted potential biomarkers using receiver-operating characteristic curve analysis to predict LLD utilizing the combination of the microbiota and inflammatory cytokines.
Results: Elevated systemic inflammatory cytokine levels and gut microbiota dysbiosis were found in LLD patients. Relative abundance of Verrucomicrobia at the phylum level and Megamonas, Citrobacter, and Akkermansia at the genus level among LLD patients was lower than HCs. Abundance of Coprococcus, Lachnobacterium, Oscillospira, and Sutterella was higher in LLD patients. Notably, IL6, IFNγ, Verrucomicrobia, and Akkermansia levels were correlated with depression severity. Our study identified IL6, Akkermansia, and Sutterella as predictors of LLD, and their combination achieved an area under the curve of 0.962 in distinguishing LLD patients from HCs.
Conclusion: This research offers evidence of changes within gut microbiota and systemic inflammation in LLD. These findings possibly help elucidate functions of gut microbiota and systemic inflammation in LLD development and offer fresh ideas on biomarkers for clinical practise in the context of LLD.
Keywords: late-life depression, gut microbiota, inflammation, inflammatory cytokines
Introduction
Late-life depression (LLD) is defined as a depressive disorder that occurs during old age (>60 years)1 and includes both the first onset of depression during old age and depression that occurs prior and continues into or recurs during old age. This study focuses specifically on primary depressive disorders with first onset occurring at an older age (>60 years). With the increase in life expectancy worldwide, the prevalence of LLD has increased significantly, and approximately 4% of the world’s elderly are diagnosed with LLD.2 Although the diagnostic criteria for symptomatology of LLD and adult depression are similar, unlike depression occurring in younger adults, LLD is characterized by prominent physical symptoms, while emotional symptoms are inconspicuous and cognitive impairment more severe.3,4 Moreover, research4 suggests that LLD may be a precursor of dementia. Above all, it is suggested that LLD may have different pathological mechanisms from adult depression. As a result, the low rates of treatment and treatment success present a great burden to patients and society. However, the current strategy for diagnosing LLD is mainly based on mental assessment, which is subjective to some extent and subject to misdiagnosis. Therefore, there is an urgent need for a clearer understanding of the pathogenesis of LLD and a better objective biomarker for LLD.
Pertinently, the microbiome–gut–brain axis has been widely studied lately and is considered a critical factor involved in the pathogenesis of neuropsychiatric diseases that involves bidirectional regulation of intestinal microbes and the brain through neuroanatomical, neuroimmune, and neuroendocrine pathways, microbial metabolites, and the blood‒brain and intestinal mucosal barriers.5 Countless studies on gut microbiota and aging have been done. More specifically, Zhuang et al6 found that Lachnospiraceae abundance was decreased among Alzheimer’s disease (AD) patients, with Ruminococcaceae abundance increased. For frail old people, Akkermansia, Parabacteroides, and Klebsiella levels are increased, but Prevotella, Faecalibacterium, Megamonas, Roseburia, and Blautia levels diminished.7 However, changes in gut microbiota in LLD and the mechanisms leading to LLD remain unclear.
Inflammation is another process extensively studied in the context of psychoneurological disease pathogenesis. The interaction of peripheral immunoresponses with the central nervous system can lead to important alterations in central nervous system immunity. Cytokines and chemokines cross the blood‒brain barrier by active transport8 and stimulate microglia in the central nervous system, thereby activating immunoresponses. If immunoimbalance within peripheral and central nervous systems is maintained, immunoresponses will be amplified by immunoproteins and -cytokines, leading to neurodegeneration.9 Studies have also shown that psychoneurotic diseases cause dysregulation of innate and adaptive immunoresponses, leading to changes in proinflammatory and anti-inflammatory cytokines.10 Levels of IL6 and TNFα are tremendously increased among acute patients suffering from schizophrenia and bipolar mania.11
Both gut microbiota and inflammation are possibly vital for the pathogenesis of neuropsychiatric diseases. Some studies indicate that the gut microbiota promotes neuroinflammation through immunoactivation, thus causing disease.12 However, many studies have found that the etiology of neuropsychiatric diseases is complex, and a single inflammatory pathway cannot completely explain the occurrence and development of disease.13,14 Moreover, studies have found that not all gut microbes work through inflammatory pathways. For example, many types of Lactobacillus and Bifidobacterium directly generate γ-aminobutyric acid, a primary inhibitory neurotransmitter in brain.15 Moreover, Candida, Escherichia coli, and Enterococcus produce the neurotransmitter serotonin, while some Bacillus strains produce dopamine.15 Therefore, the gut microbiota can mediate bidirectional microbiome–gut–brain axis interactions to regulate host neurophysiology by producing excessive metabolites in the intestinal cavity and exchanging sensory information with the host. Furthermore, probiotics have been found to have both broad anti-inflammatory and antidepressant effects, further supporting the joint role of gut microbiota and inflammation in the pathophysiology of depression.16 Therefore, combining gut microbiota with inflammation may reveal a more complete picture of LLD. Our aim could be particularly valuable to understand the gut microbial and inflammatory characteristic in LLD and to further identify biomarkers that might differentiate LLD and HC subjects. This study first explored the overall composition of systemic inflammatory cytokines in peripheral blood and the gut microbiota in LLD patients. Then, we analyzed the correlation among changed gut microbiota features, systemic inflammatory markers, and clinical characteristics of LLD patients to identify a relationship between the gut microbiota and inflammation. Additionally, we extracted potential biomarkers to predict LLD utilizing a combination of the microbiota and inflammatory cytokines.
Methods
Subjects
In total, 35 LLD patients and healthy controls (HCs) were enrolled in this observational cross-sectional analysis, eight (six LLD patients and two HCs) of which were not included because their stool samples did not meet the criteria due to the presence of diarrhea on the day of stool sample collection. Finally, data from a recruitment of 29 untreated LLD patients and 33 HCs from the Psychiatry Department of Zhejiang Provincial People’s Hospital from January 2020 to December 2021 were analyzed. All subjects or their caregivers signed informed consent forms and completed questionnaires for collecting demographic information, sex, age, BMI, years of education, medical history, smoking, and alcohol-use history included. This study complied with the guidelines of the Declaration of Helsinki and obtained approval from the Ethics Committee of Zhejiang Provincial People’s Hospital (2019KY184).
Inclusion criteria for the LLD group were age 60–85 years old and patients that had first met the criteria for major depression disorder (MDD) on the basis of the Diagnostic and Statistical Manual of Mental Disorders, fifth edition after the age of 60 years. Exclusion criteria were 1) neurological disease, such as dementia, brain tumor, or stroke; 2) history of substance abuse or dependence; 3) depressive disorders secondary to other mental disorders, such as bipolar depression or schizophrenia; and 4) serious heart, lung, liver, kidney, blood system, or endocrine disease or tumors. The HC group denied neurological or mental disorders and had normal cognitive function, with a clinical dementia rating17 score of 0 and Montreal Cognitive Assessment — Beijing version score ≥26.18
The exclusion criteria for gut microbiota sampling 1) major gastrointestinal surgery (including cholecystectomy and appendectomy) and major intestinal resection within the last 5 years; 2) concurrent inflammatory bowel disease, such as ulcerative colitis, Crohn’s disease or uncertain colitis, irritable bowel syndrome, or other persistent, infectious gastroenteritis, colitis or gastritis; 3) persistent or chronic diarrhea of unknown cause, recurrent Clostridium difficile infection, or Helicobacter pylori infection untreated; 4) previous gastrointestinal ulcers, bleeding, abdominal pain, dyspepsia, diarrhea, constipation, gastrointestinal polyps, dysplasia, or cancer; and 5) use of probiotics, prebiotics, antibiotics, drugs containing bismuth alkaline salicylate, and other components within 3 months prior to sampling.
Assessment of Depressive Symptoms
All patients underwent a medical history assessment and psychiatric examination by two highly trained psychiatrists. On the day of enrollment, depression severity was determined using the Chinese version of the 24-item Hamilton Depression Rating Scale (HDRS-24) and the Chinese version of the 30-item Geriatric Depression Scale (GDS-30). The HDRS-24 is the most widely used scale to assess depression in clinical practice.19 Symptoms are scored on a five-point scale, ranging from asymptomatic to extremely severe symptoms (with a small number of questions ranging from 0 to 2) on seven subscales: anxiety/somatization, weight, cognitive impairment, circadian change, delay, sleep disturbance, and hopelessness. According to Davis JM’s delineation score, the cut-off points for the approval of HDRS-24 are as follows: >35 is considered severe, >20 is considered moderate, <8 indicates no depression.19 The GDS-30 was used to assess depressive symptoms in the elderly.20 Participants answered “yes” or “no” to each question. Scores range from 0 to 30, with 0–10 being normal, 11–20 mildly depressed, and 21–30 moderately to severely depressed.21
Analysis of Serum Inflammatory Factors
Venous blood (5 mL) was collected from every subject on an empty stomach in the morning, then these were centrifuged at 4°C for 15 min at 3000 r/min within 1 hour. The serum was aliquoted and stored at −80 °C. Levels of 12 inflammatory markers in plasma were measured using a Cytometric Bead Array human cytokine kit (BD, San Diego, CA, USA) assays according to the manufacturer’s instructions. Markers comprised IL2, IL4, IL5, IL6, IL8, IL10, IL12p70, IL17, IL1β, IFNγ, IFNα, and TNFα.
Fecal Sample Collection and DNA Extraction
A sample of at least 3 g was collected from each subject, and sampling was repeated three times. DNA extraction was performed with nucleic acid extraction or purification reagent (Hangzhou Guhe: GHFDE100), and DNA concentration and quality were defined by a NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), followed by agarose gel electrophoresis.
16S rRNA Sequencing
The V4 region of the bacterial 16S rRNA gene was amplified by PCR via forward primer 515F (5ʹ-GTGCCAGCMGCCGCGGTAA-3ʹ) and reverse primer 806R (5ʹ-GGACTACHVGGGTWTCTAAT-3ʹ). Barcode uses a specific 7-bp sequence synthesized into the sequence. Phusion high-fidelity PCR master mix with HF buffer was employed for a 50 μL/25 μL PCR reaction system, with F/R primers before and after 3 μL (1 μL) each, a 10 μL DNA sample, and a ddH2O complement to 50 μL. The PCR system was configured to perform PCR amplification with reaction conditions of predenaturation 98°C 30 s, 30 cycles; denaturation 98°C, 15 s; annealing 58°C, 15 s; and extension 72°C, 15 s, and final elongation at 72°C for 1 min. PCR product purification was done using AMPure XP Beads (Beckman Coulter, Indianapolis, IN) and quantification via a Qubit dsDNA HS assay kit. Following quantification, an Illumina Novaseq 6000 pin-END 2×150 bp platform was employed for sequencing (GUHE Info Technology, Hangzhou, China).
Sequence Data Processing and Bioinformatic Analysis
Data of every sample were split from original data based on barcode sequence and primer sequence. After deleting barcode and primer sequences, reads of every sample were spliced via Vsearch v2.4.4 for obtaining raw tag data. Meanwhile, sequence quality was controlled and filtered. Criteria for screening low-quality sequences were sequence <150 bp, mean quality values <20, sequences involving ambiguous base and sequences containing over 8 bp single-nucleotide repeats were discarded, and then the final effective tags were acquired.
OTU analysis uses Vsearch v2.4.4. This includes de-repeating sequences (--derep_fulllength), clustering (--cluster_fast,--id 0.97), and de-chimerism (--uchime_ref).22 By default, sequences were clustered into OTUs with 97% similarity, and representative sequences of OTU were chosen via default parameters. Representative sequences were annotated by species on the basis of the SILVA128 database23 through VSEARCH, and the OTU list was further generated. At each taxonomic level, the community composition of kingdom, phylum, class, order, family, genus, and species was counted. OTU abundance per sample and classification was recorded, and content in all samples < 0.001% of the total sequence of OTUs was removed.
Sequence data analysis primarily adopts QIIME and the R package (v3.2.0). QIIME software was employed for calculating the OTU-level alpha-diversity index, including the Chao1, ACE, Shannon, and Simpson indices. Alpha diversity refers to interarray variance analysis for comparison of OTU abundance and uniformity among samples. Beta-diversity analysis was done via Qiime software for calculating the UniFrac distance metric,24 then a principal coordinate analysis (PCoA) map was used for analyzing the beta diversity of microbial community structure among diverse samples.
Monte Carlo permutation and Student’s t-tests were employed for drawing a box plot for comparing differences in UniFrac distances between groups. Markers assessing intergroup differentiation of microbial community structure were evaluated via permutational analysis of variance and a method derived from “vegan” of the R pack. The R stats package Kruskal method was employed for comparing differences among different taxonomic levels, classes, orders, families, and genera between samples or groups. Linear discriminant analysis (LDA) effect size (LEfSe) analysis uses default LEfSe setting for detecting differences in taxonomic units between groups. The LDA score threshold was set to 2 and the P-value threshold to 0.05.
To gain an in-depth understanding of the differences in gut microbiota functional pathways between the LLD group and HCs, PICRUSt (Phylogenetic Investigation of Communities by Reconstruction of Unobserved States version 1.1.3) analysis performed based on the Kyoto Encyclopedia of Genes and Genomes (KEGG) database was applied to microbial function prediction. Here, we present only the third-level results.
Statistical Analysis
Statistical analysis was done via SPSS 25.0 and data visualization via GraphPad Prism 6. Data are displayed as means ± SD. Two-sided independent Student’s t-tests were employed for data with normal distribution and Mann–Whitney U test for data with abnormal distribution. Statistical data are displayed as percentages, and the χ2 test was used to group comparison. Correlations among neuropsychiatric scale scores, serum inflammatory factor levels, and gut microbe abundance were determined using Spearman correlation analysis. Since fitting of the regression model requires no correlation between the variables used as predictive variables, whether the altered gut microbiota and inflammatory factors can be used as predictive variables could be determined through the correlation results. Features that were not correlated could be included in the regression model. Logistic regression analysis was then applied to identify indicators among inflammatory factors and gut microbiota constituents that could be used to distinguish LLD patients from HCs. The value of inflammatory cytokines and gut microbiota characteristics in the identification of LLD was assessed via receiver-operating characteristic (ROC) curves. Two-sided P<0.05 was regarded as statistically significant.
Results
Clinical Characteristics
In sum, 29 LLD patients and 33 elderly HCs were included. The two groups showed no significant differences in sex, age, BMI, smoking, or alcohol consumption (Table 1). The HDRS-24 and GDS scores of LLD patients are shown in Table 1. Serum levels of the inflammatory cytokines IL2, IL4, IL5, IL6, IL8, IL10, IL12P70, IL17, IL1β, TNFα, IFNα, and IFNγ were tremendously higher among LLD group relative to HCs (P<0.001) (Table 1).
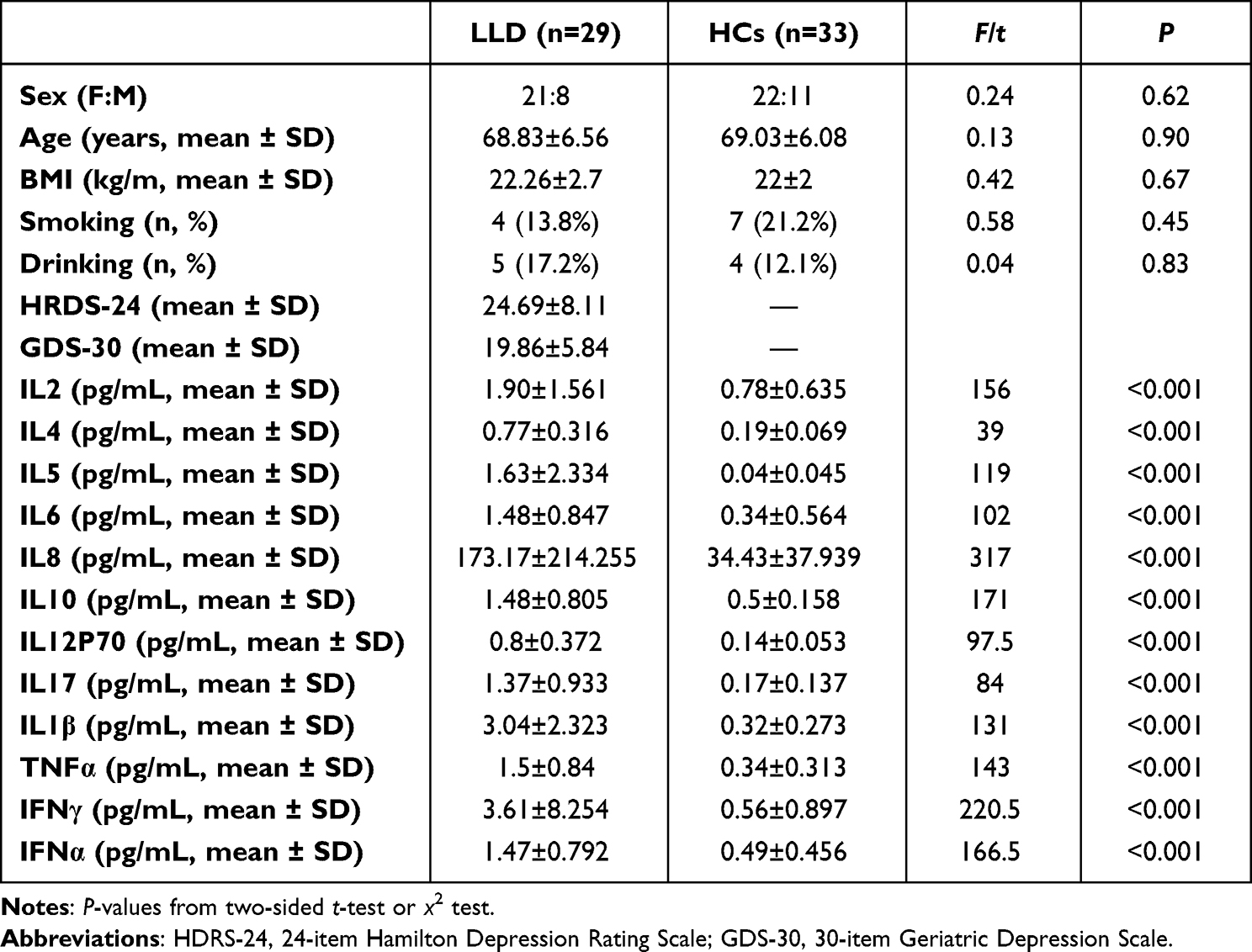 |
Table 1 Clinical data of participants |
Comparative Analysis of Diversity Between Groups
Alpha diversity reflects the abundance and evenness of species within a given ecosystem. Chao1, Shannon, and Simpson indices were employed for measuring the intestinal microbial diversity of individuals in two groups. The Chao1 index in LLD group remained tremendously higher (P=0.0431) (Figure 1A). No significant differences between the groups on other indices were found (Figure 1B and C).
 |
Figure 1 Alpha diversity of gut microbiota in LLD group and HCs. Abbreviations: LLD, late-life depression; HCs, healthy controls. Notes: Gut microbiota alpha-diversity indices in LLD and HCs. Box plots display differences among microbiome-diversity indices between LLD and HCs via (A) Shannon diversity index, (B) Simpson index, and (C) Chao1 index. Each box plot denotes medians, interquartile ranges, and minimum and maximum values. |
Beta-diversity analysis reflects the evolutionary relationship between each sample sequence, ie, abundance information, for calculating distance between samples as an indicator of significant microbial community differences between groups. In this study, we used PCoA (with unweighted UniFrac distance) for describing beta diversity. There existed tremendous structural differences between the LLD and HC groups in the first dimension of spatial distance (Figure 2A). The PCoA results showed the former two main factors explained 20.17% and 6.25% of the entire variation, respectively (P<0.05). As per Figure 2B, there existed tremendous differences in gut microbiota composition between LLD patients and HCs (Bray–Curtis: R=0.129, P=0.001).
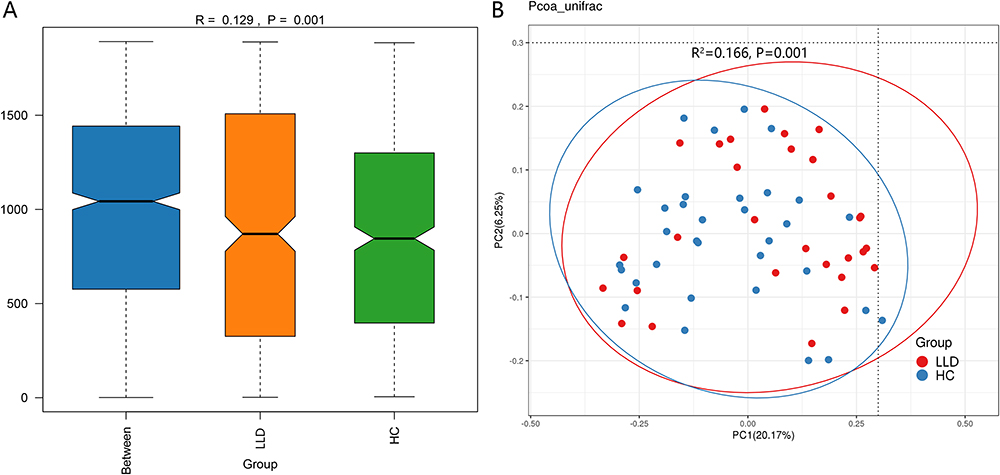 |
Figure 2 Beta diversity of gut microbiota in LLD group and HCs. Abbreviations: LLD, late-life depression; HCs, healthy controls. Notes: Gut microbiota beta-diversity indices in LLD and HCs. Level of similarity between gut microbial communities detected in LLD (orange) and HCs (green) was evaluated through (A) unweighted analysis of similarity (ANOSIMs) and (B) principal coordinate (PC) analysis (according to UniFrac distance matrix). ANOSIM R values display community variations between the compared groups. One symbol denotes one sample. |
Changed Microbiota Composition and Abundance in LLD Patients
Relative abundance of gut microbes was compared between two groups. At the phylum level, Verrucomicrobia abundance remained tremendously lower in the LLD group (P=0.0148, Figure 3A). At the genus level, Megamonas, Citrobacter, and Akkermansia levels were lower (P<0.05) in LLD patients than in HCs (Figure 3B–D). However, Coprococcus, Lachnobacterium, Oscillospira, and Sutterella (P<0.05) abundance remained higher in the LLD group (Figure 3E–H).
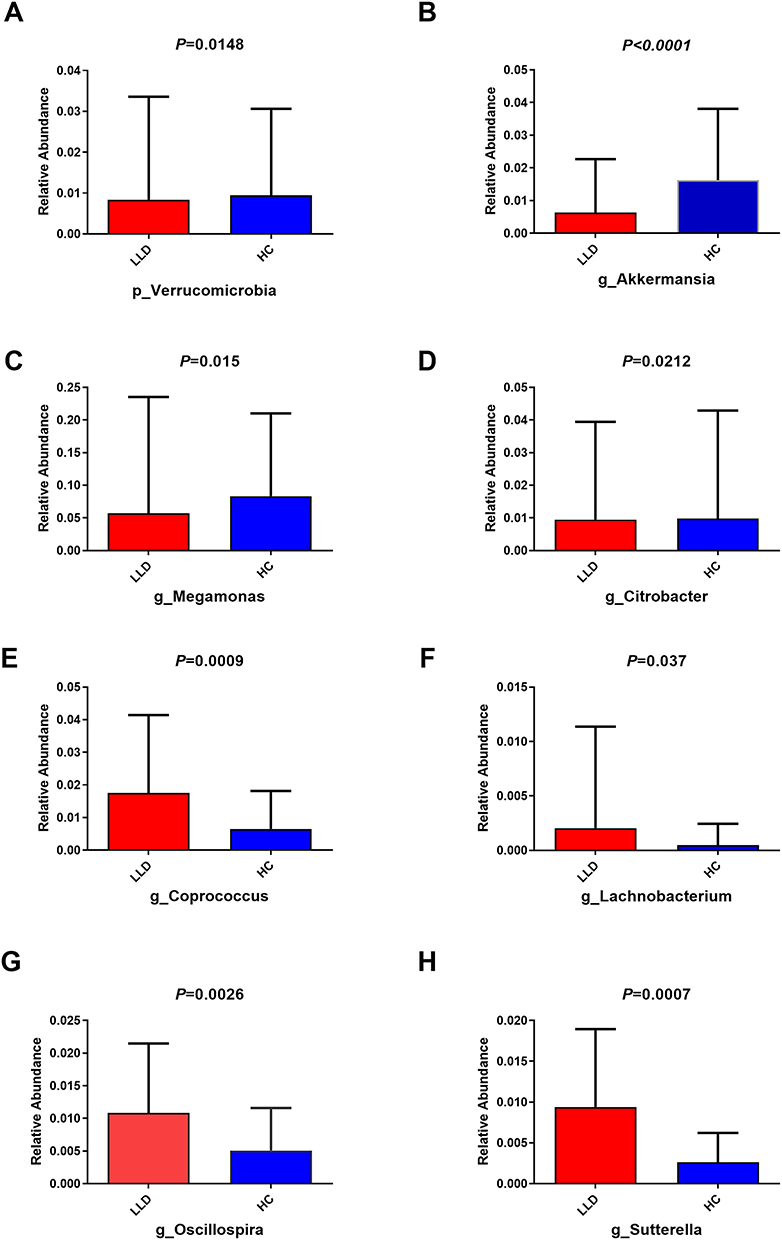 |
Figure 3 Altered microbiota composition and abundance distribution among LLD patients. Abbreviations: LLD, late-life depression; HCs, healthy controls. Notes: Altered microbiota composition and abundance distribution in LLD patients. At phylum level, (A) Verrucomicrobia abundance was greatly lower in LLD relative to HCs. At genus level, (B) Akkermansia, (C) Megamonas and (D) Citrobacter levels were lower in LLD relative to HCs. (E) Coprococcus, (F) Lachnobacterium, (G) Oscillospira and (H) Sutterella abundance remained higher in LLD relative to HCs. |
LEfSe analysis can further identify species or communities (ie, biomarkers) that have significant differences among samples. LEfSe analysis revealed 23 characteristic species with tremendous difference in relative abundance between two groups (P<0.05, LDA >2; Figure 4B). At the phylum level, Verrucomicrobia was enriched in HCs, whereas at the genus level, the biological markers Akkermansia, Megamonas, and Citrobacter were identified in HCs. Biomarkers of the LLD group at the genus level were Lachnobacterium, Phyllobacterium, Dorea, Oscillospira, Coprococcus, Sutterella, and Acinetobacter (P<0.05, LDA >2; Figure 4A and B).
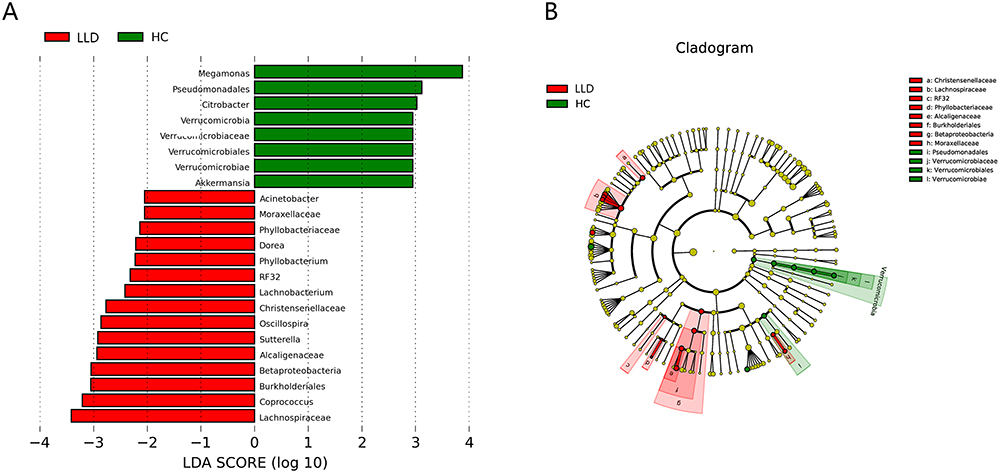 |
Figure 4 LEfSe analysis of LLD patients and HCs. Abbreviations: LLD, late-life depression; HCs, healthy controls. Notes: Taxonomic representation of great difference in relative abundance between LLD and HCs. (A) LDA scores for bacterial taxa with differential abundance between LLD and HCs. Positive and negative LDA scores denotebacterial taxa enriched in LLD and HCs separately. Taxons with P<0.01 (Wilcoxon rank-sum test) and LDA >2 are displayed. (B) Cladogram-generated LEfSe indicating difference in bacterial taxa between subjects with LLD and HCs. Nodes in red denote taxa enriched in LLD relative to HCs, while nodes in green denote taxa enriched in HCs relative to LLD. |
Comparison of Predicted Gut Microbiota Function Between Groups
To reveal the differences in gut microbiota functions between groups, PICRUSt analysis was employed. Through PICRUSt functional analysis of the 16S rRNA gene sequence, three levels of functional gene enrichment in KEGG pathways were observed. At level 3, we found 38 metabolic pathways with tremendous difference between groups (P<0.05, Figure 5). For example, LLD patients showed enrichment of amino acid–related enzymes, aminoacyl-tRNA biosynthesis, and other pathways, but glutathione metabolism and ubiquinone biosynthesis pathways were less enriched.
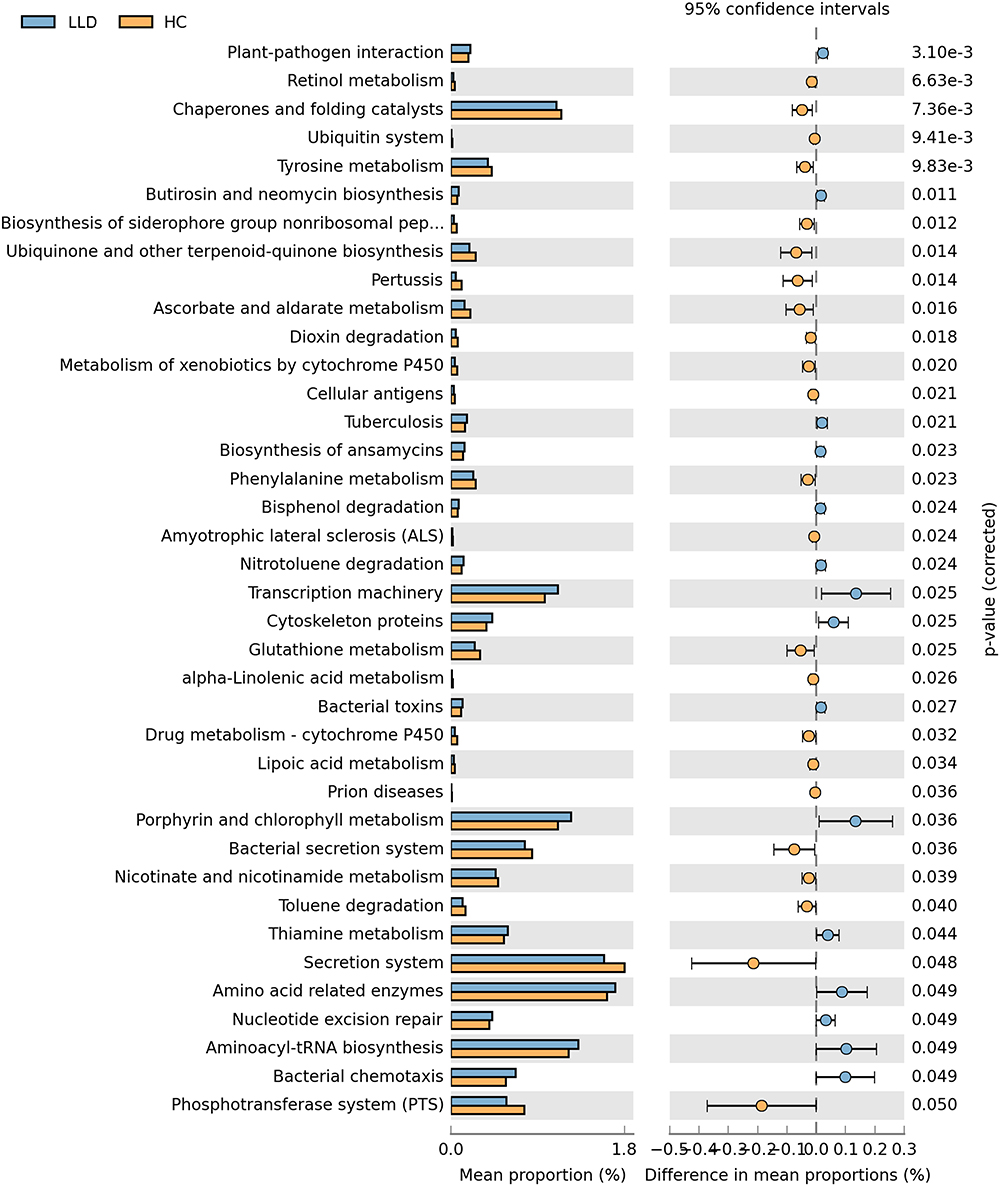 |
Figure 5 PICRUSt-based examination of functions of gut microbiome in LLD patients and HCs. Abbreviations: LLD, late-life depression; HCs, healthy controls. Notes: The left of the figure displays abundance ratios between the groups. The middle section displays proportional variation in functional abundance within 95% CI. P values at the right. |
Correlation Between Gut Microbiota Changes and Clinical Indicators
For determining whether alterations in gut microbiota are completely associated with inflammation, and since fitting of the linear regression model requires that there should be no correlation between variables used as predictors, it can be determined whether intestinal microbes and inflammatory factors can be used as predictors through the correlation findings. Verrucomicrobia and Akkermansia levels were negatively related to HDRS and GDS scores (P<0.05; Table 2, Figure 6). Both IL6 (r=0.4380, P=0.0175) and IFNγ (r=0.4251, P=0.0215) levels had positive correlations with GDS scores (Table 2 and Figure 6). Moreover, Megamonas abundance was negatively correlated with IL1β, IL4, IFNγ, and IL17 levels (P<0.05; Table 2 and Figure 6).
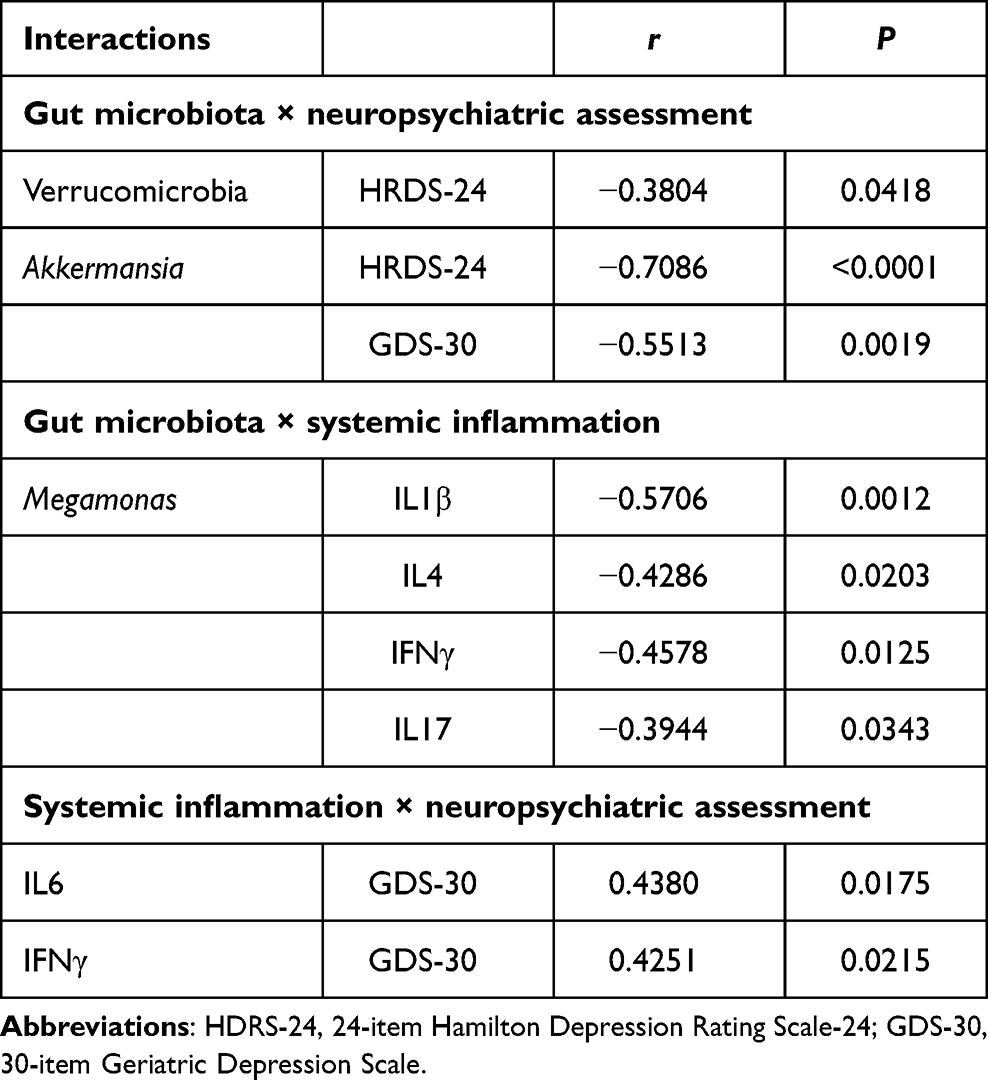 |
Table 2 Correlations among changed gut microbiota, neuropsychiatric assessment and systemic inflammation markers |
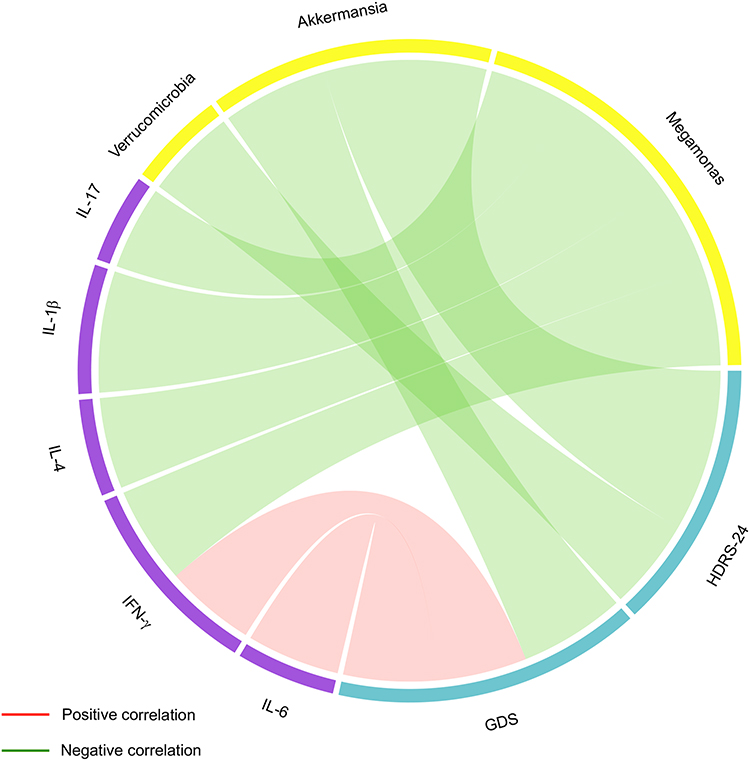 |
Figure 6 Connections among changed gut microbes, clinical variables, and inflammatory factors identified by Spearman correlation. Abbreviations: HDRS-24, 24-item Hamilton Depression Rating Scale; GDS-30, 30-item Geriatric Depression Scale. Notes: Connections among changed gut microbiota, neuropsychiatric assessment, and systemic inflammation identified by Spearman correlation. Correlations with P<0.05 are displayed. |
Combined Analysis of Gut Microbiota Composition and Inflammatory Factor Levels Can Improve the Accuracy of LLD Diagnosis
Logistic regression was first applied to determine gut microbiota constituents and inflammatory factors that could be used to distinguish LLD patients from HCs. The results showed tremendous differences in IL6 (P<0.001), Akkermansia (P=0.018), and Sutterella (P=0.003) levels between LLD and HCs. Then, potential discriminative value of these classifiers was evaluated using ROC curves. We found that when gut microbiota features were used alone as a predictor, the AUC was 0.746 (95% CI 0.621–0.871, Figure 7). When inflammatory factors were used as a predictor, the AUC was 0.893 (95% CI 0.909–0.9777, Figure 7). However, when the two were used in combination as predictors, the AUC was 0.962 (95% CI 0.92–1, Figure 7). Combined analysis of gut microbiota composition and inflammatory factor levels can improve the accuracy of LLD diagnosis.
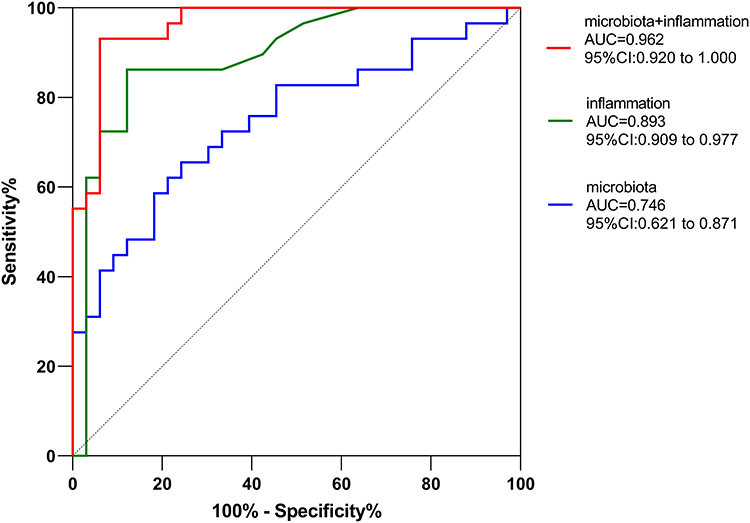 |
Figure 7 Receiver-operating characteristic curve analysis of sensitivity and specificity of differentially abundant genera as diagnostic factors for LLD. Abbreviation: LLD, late-life depression. Notes: The ROC generated using microbiota features is marked in blue, inflammatory factor levels marked in green, and microbiota features combined with inflammatory factor levels marked in red. |
Discussion
This is the initial research for revealing gut microbiome characteristics and systemic inflammation in LLD patients and to conduct a combined analysis of the two to identify biomarkers of LLD. We explored characteristics in gut microbiota and systemic inflammatory factors in patients with LLD and explored their association with depression severity. Elevated systemic inflammatory factor levels, increased taxonomic richness, and changed composition of gut microbiota were found among patients suffering from LLD. IL6 and IFNγ levels and Verrucomicrobia and Akkermansia abundance were related to depression severity. Moreover, Megamonas abundance was negatively correlated with IL1β, IL4, IFNγ, and IL17 levels. More importantly, IL6, Akkermansia and Sutterella levels were predictors of LLD. Notably, the combination of gut microbiota and inflammatory factors had greater diagnostic power as a potential biomarker for LLD.
We found increased levels of both proinflammatory cytokines (eg, IL6, IFNγ, and TNFα) and anti-inflammatory cytokines (eg, IL4 and IL10) in LLD. We speculate that there are various mechanisms regulating cytokine-production changes in the body during LLD and that each cytokine is involved in activation of and participation in multiple downstream signaling pathways, exerting complex synergistic or antagonistic effects that eventually lead to the emergence of clinical manifestations. More importantly, we found that IL6 and IFNγ levels had positive correlations with LLD disease severity, suggesting that these two inflammatory factors participate in LLD occurrence and development. IL6 is a proinflammatory cytokine expressed in neurons and glial cells that is vital for immune and acute-phase responses.25 Some scholars have proposed that high IL6 levels can promote serotonin degradation and reduce serotonin production, which may impair neural plasticity and lead to structural brain abnormalities, such as hippocampal and frontal lobe atrophy,10,26 which have been confirmed to be associated with LLD and LLD-induced cognitive impairment. Consistently with our study, many others have found elevated IL6 levels among patients suffering from LLD that are positively related to disease severity.27–29 In contrast, the results of studies on IFNγ in MDD are inconsistent. Zhang et al30 found that IFNγ levels increased in mice with depression-like features and were associated with cognitive deficits. However, Buspavanich et al31 found a reduction in IFNγ levels among MDD patients. Pertinently, IFNγ is a proinflammatory factor participating in central nervous system inflammation and activates microglia in the brain to induce proinflammatory responses.32 Studies have shown IFNγ-activated microglia alter the hippocampal neurogenic niche, inhibit the proliferation of neural stem and progenitor cells, and facilitate the apoptosis of immature neurons, thereby causing depression symptoms and cognitive deficits in mouse.30 Our study confirms this idea clinically and demonstrates IFNγ levels are related to LLD severity. In brief, this research reveals that LLD is an inflammatory state where proinflammatory and anti-inflammatory cytokines coexist and that IL6 and IFNγ are related to disease severity.
In our study, the Chao1 index in the LLD group remained tremendously higher than the HC group, indicating that richness of gut microbiota among LLD patients remains greater than in HCs. However, the Shannon and Simpson indices did not differ tremendously between the groups, indicating that diversity of gut microbiota did not differ between the two. Our findings are similar to previous studies on MDD,33,34 but other studies on LLD have revealed that richness and diversity in intestinal microbiota showed no difference between patients and controls.35 We hypothesize the augmented gut microbiota richness among LLD patients is possibly associated with the increase in the levels of harmful bacteria rather than in those of the core microbiota in the gut. However, notably, the gut microbiota is a dynamic community influenced by factors like genetics, diet, age, stress, geography, and health status. Therefore, the characteristics of the diversity and richness of gut microbiota in LLD patients need to be further studied.
Beta-diversity analysis revealed tremendous changes in gut microbiota compositions in LLD patients. We discovered microbes with decreased abundance were mainly probiotics, and most of the bacteria with increased abundance were involved in protein and fat metabolism. Verrucomicrobia and Akkermansia are strains of the same family. Verrucomicrobia includes a small group of bacteria found in the human gut, and a recent study showed that Akkermansia are beneficial bacteria that can potentially be used in next-generation therapies.36 Previous research has revealed Verrucomicrobia abundance decreases among patients suffering from anxiety and depression,37 and an increase in Akkermansia abundance can decrease anxiety and enhance cognition in middle-aged mice.38 This research matches preceding research and complements studies among elderly populations.7 Notably, Akkermansia abundance remained tremendously decreased among patients suffering from LLD and related to depression severity, and logistic regression showed that Akkermansia abundance is a risk factor that can be used to predict the probability of LLD. However, there was no correlation between Akkermansia abundance and inflammatory factor levels. This finding reveals Akkermansia may have no function in LLD pathogenesis through inflammatory pathways. Interestingly, Wang et al39 found that through direct interactions between the Akkermansia FF protein Amuc_1100 and TLR2, Akkermansia promoted expression of the 5-HT synthesis rate–limiting enzyme Tph1 in RIN-14B cells and decreased expression of serotonin-reuptake transporters in Caco-2 cells, hence increasing the biosynthesis and extracellular availability of 5-HT, which suggested that Akkermansia can affect LLD by regulating the release of neurotransmitters directly through the intestinal barrier. Further studies are needed in the context of LLD.
We further found that Megamonas abundance was correlated with levels of IL1β, IL4, IL17, and IFNγ, which are mostly proinflammatory factors. Megamonas belongs to the phylum Firmicutes, which comprises important short-chain fatty acids producing bacteria that produce acetic acid, propionic acid, and lactic acid by fermenting various carbohydrates. Of note, the abundance of Megamonas has been shown to differ in various studies of depression.40 Zheng et al41 found that Megamonas abundance decreased in patients with depression, and another study showed that Megamonas abundance decreased in elderly patients with frailty,7 which is consistent with our study. Research has revealed that butyrate is vital for inhibiting intestinal inflammation: it can change intestinal permeability and influence regulatory T-cell activity to exert anti-inflammatory effects.42 Therefore, we speculate that in patients with LLD, Megamonas reduces butyrate output, thus influencing regulatory T-cell activity and inducing proinflammatory cytokine release, which triggers local gut inflammation and drives LLD through the microbe–gut–brain axis. Based on the results regarding the aforementioned inflammatory factors, we hypothesize that Megamonas may mediate the occurrence and development of LLD by upregulating IFNγ production, but further in-depth mechanistic research is still needed. Other studies have shown that Megamonas is involved in the release of inflammatory factors in psoriasis,43 cancer pain,44 and other diseases, but the underlying mechanism remains to be explored.
Furthermore, we found that Coprococcus, Lachnobacterium, Oscillospira, and Sutterella levels were elevated at the genus level in LLD patients. Most of these bacteria with increased levels are related to protein and fat metabolism. For example, studies have shown that elevated Oscillospira levels may be related to increased protein metabolism.40 Coprococcus has a positive correlation with obesity.45 Increased protein metabolism mediated by the microbiota involves fermentation or bacterial protein degradation, a process possibly involving the transfer of essential amino acids from host to microorganism and leading to the production of toxic products like ammonia, putrescine, and phenol, which in turn cause intestinal barrier disruption and neuroinflammation. More remarkably, through logistic analysis, Sutterella is predictive of LLD. Sutterella is an important intestinal commensal bacterium.46 Research has revealed that Sutterella abundance is significantly increased among patients suffering from MDD and generalized anxiety disorder.47,48 Many studies have found that Sutterella is positively correlated with obesity, as well as with weight and fat gain.49–51 The bidirectional relation of obesity and depression is well established, and research has revealed hyperactivation of the hypothalamic–pituitary–adrenal (HPA) axis bringing about maladaptive release of cortisol may be a common mechanism for both.52 Therefore, we speculate that Sutterella might affect cortisol release by affecting the HPA axis, thus triggering LLD.
Through correlation analysis, we found that not all alterations in gut microbiota were related to inflammation, suggesting that some alterations in gut microbiota possibly have pathological functions in LLD through other pathways. PICRUSt analysis revealed activation of multiple signaling pathways related to gut microbiota, including amino acid metabolism, glutathione metabolism, and the HPA axis, which suggests that some gut microbes may also participate in the development of LLD through other noninflammatory pathways. Thus, we conclude that gut microbiota and inflammation interact in the pathogenesis of LLD, but there are also disease-causing processes that are independent of this mechanism.
A single indicator cannot fully explain the pathogenesis of LLD, nor can it be used to predict LLD more accurately. Therefore, we used both gut microbiota and inflammatory index to be biomarkers of LLD. Initially, we excluded the gut microbes associated with inflammatory factors through correlation analysis and then extracted IL6, Akkermansia and Sutterella as predictors through logistic regression. The ROC curve was employed for assessing sample classification accuracy. We found that the combination of inflammatory cytokines and gut microbiota could improve LLD diagnosis accuracy. Our research therefore provides a new approach for the distinction of LLD. Previous studies have demonstrated that gut microbes or inflammatory cytokines can be used as noninvasive biological markers to identify neuropsychiatric disorders, such as bipolar disorder,53 methamphetamine abuse,54 and schizophrenia.55 However, there is little research on the combined analysis of the two factors. Some studies have reported that other biological markers can be used in conjunction with inflammatory markers to more accurately predict neuropsychiatric disorders. For instance, Carniel and da Rocha56 found that BDNF and inflammatory factors can be used as predictors of MDD. In addition, Wei et al57 demonstrated that the combination of the systemic inflammation response index, neutrophil:high-density lipoprotein ratio, and monocyte:high-density lipoprotein ratio can improve the effectiveness of bipolar disorder diagnosis, though preceding research has primarily emphasized functions of gut microbiota and inflammation, with little attention given to gut microbiota functions in causing mental health disorders through other mechanisms. We believe that this component of noninflammatory microbiota should not be ignored in the prediction of the onset of LLD and should be included in the diagnostic model with inflammation indicators to predict LLD more accurately. However, biomarkers based on intestinal microbial markers and inflammatory factors need to be validated in larger samples.
There exist certain shortcomings of this research. Primarily, gut microbiota can be influenced by a variety of living environments, and although the subjects in this research were not allowed to use antibiotics or probiotics, none of them was given a standardized diet. Second, the research only contained small clinical samples of the elderly. Therefore, the sample size needs to be further expanded. Third, this was a cross-sectional study, and longitudinal research may provide stronger evidence.
Conclusion
Together, our findings reveal that alterations in gut microbiota and inflammatory cytokines in LLD contribute to the development of LLD. Most importantly, the combination of gut microbiota and inflammatory markers has improved power as a potential biomarker for LLD. These discoveries thus offer a deeper awareness of gut microbiota functions on LLD pathogenesis and elucidate new strategies for LLD diagnosis and treatment.
Abbreviations
LLD, late-life depression; HCs, healthy controls; ROC, receiver-operating characteristic; MDD, major depressive disorder; AD, Alzheimer’s disease; HDRS-24, 24-item Hamilton Depression Rating Scale; GDS-30, 30-item Geriatric Depression Scale; ELISA, enzyme-linked immunosorbent assay; QIIME, Quantitative Insights Into Microbial Ecology; OTU, operational taxonomic unit; PCoA, principal coordinate analysis; KEGG, Kyoto Encyclopedia of Genes and Genomes; ANOSIM, analysis of similarities; LDA, linear discriminant analysis; LEfSe, LDA effective size; PICRUSt, Phylogenetic Investigation of Communities by Reconstruction of Unobserved States.
Ethics Approval and Consent
This research obtained approval from the Ethics Committee of Zhejiang Provincial People’s Hospital (2019KY184). Consent was informed and signed by all subjects or their caregivers.
Funding
This paper obtained support from the Science Foundation of the Health Commission of Zhejiang Province (2020KY405).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Pomara N, Bruno D, Plaska CR, et al. Plasma amyloid-beta dynamics in late-life major depression: a longitudinal study. Transl Psychiatry. 2022;12(1):301. doi:10.1038/s41398-022-02077-8
2. Chang DD, Eyreeuro HA, Abbott R, et al. Pharmacogenetic guidelines and decision support tools for depression treatment: application to late-life. Pharmacogenomics. 2018;19(16):1269–1284. doi:10.2217/pgs-2018-0099
3. Kok RM, Reynolds CF. Management of depression in older adults: a review. JAMA. 2017;317(20):2114–2122. doi:10.1001/jama.2017.5706
4. Diniz BS, Butters MA, Albert SM, Dew MA, Reynolds CF 3rd. Late-life depression and risk of vascular dementia and Alzheimer’s disease: systematic review and meta-analysis of community-based cohort studies. Br J Psychiatry. 2013;202(5):329–335. doi:10.1192/bjp.bp.112.118307
5. Wang HX, Wang YP. Gut microbiota-brain axis. Chin Med J. 2016;129(19):2373–2380. doi:10.4103/0366-6999.190667
6. Zhuang ZQ, Shen LL, Li WW, et al. Gut microbiota is altered in patients with Alzheimer’s disease. J Alzheimers Dis. 2018;63(4):1337–1346. doi:10.3233/JAD-180176
7. Xu Y, Wang Y, Li H, et al. Altered fecal microbiota composition in older adults with frailty. Front Cell Infect Microbiol. 2021;11:696186. doi:10.3389/fcimb.2021.696186
8. Lee SH, Shin C, Ko YH, et al. Plasminogen activator inhibitor-1: potential inflammatory marker in late-life depression. Clin Psychopharmacol Neurosci. 2023;21(1):147–161. doi:10.9758/cpn.2023.21.1.147
9. Bruunsgaard H, Pedersen M, Pedersen BK. Aging and proinflammatory cytokines. Curr Opin Hematol. 2001;8(3):131–136. doi:10.1097/00062752-200105000-00001
10. Linnemann C, Lang UE. Pathways connecting late-life depression and dementia. Front Pharmacol. 2020;11:279. doi:10.3389/fphar.2020.00279
11. Goldsmith DR, Rapaport MH, Miller BJ. A meta-analysis of blood cytokine network alterations in psychiatric patients: comparisons between schizophrenia, bipolar disorder and depression. Mol Psychiatry. 2016;21(12):1696–1709. doi:10.1038/mp.2016.3
12. Morais LH, Schreiber H, Mazmanian SK. The gut microbiota-brain axis in behaviour and brain disorders. Nat Rev Microbiol. 2021;19(4):241–255. doi:10.1038/s41579-020-00460-0
13. Menard C, Hodes GE, Russo SJ. Pathogenesis of depression: insights from human and rodent studies. Neuroscience. 2016;321:138–162. doi:10.1016/j.neuroscience.2015.05.053
14. Leng F, Edison P. Neuroinflammation and microglial activation in Alzheimer disease: where do we go from here? Nat Rev Neurol. 2021;17(3):157–172. doi:10.1038/s41582-020-00435-y
15. Sorboni SG, Moghaddam HS, Jafarzadeh-Esfehani R, Soleimanpour S. A comprehensive review on the role of the gut microbiome in human neurological disorders. Clin Microbiol Rev. 2022;35(1):e0033820. doi:10.1128/CMR.00338-20
16. Dogaru IA, Puiu MG, Manea M, Dionisie V. Current perspectives on pharmacological and non-pharmacological interventions for the inflammatory mechanism of unipolar depression. Brain Sci. 2022;12(10):1403. doi:10.3390/brainsci12101403
17. Morris JC, Ernesto C, Schafer K, et al. Clinical dementia rating training and reliability in multicenter studies: the Alzheimer’s Disease Cooperative Study experience. Neurology. 1997;48(6):1508–1510. doi:10.1212/wnl.48.6.1508
18. Nasreddine ZS, Phillips NA, Bédirian V, et al. The Montreal Cognitive Assessment, MoCA: a brief screening tool for mild cognitive impairment. J Am Geriatr Soc. 2005;53(4):695–699. doi:10.1111/j.1532-5415.2005.53221.x
19. Yong N, Hu H, Fan X, et al. Prevalence and risk factors for depression and anxiety among outpatient migraineurs in mainland China. J Headache Pain. 2012;13(4):303–310. doi:10.1007/s10194-012-0442-9
20. Yesavage JA, Brink TL, Rose TL, et al. Development and validation of a geriatric depression screening scale: a preliminary report. J Psychiatr Res. 1982;17(1):37–49. doi:10.1016/0022-3956(82)90033-4
21. Liu A, Peng Y, Zhu W, et al. Analysis of factors associated with depression in community-dwelling older adults in Wuhan, China. Front Aging Neurosci. 2021;13:743193. doi:10.3389/fnagi.2021.743193
22. Rognes T, Flouri T, Nichols B, Quince C, Mahe F. VSEARCH: a versatile open source tool for metagenomics. PeerJ. 2016;4:e2584. doi:10.7717/peerj.2584
23. Quast C, Pruesse E, Yilmaz P, et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 2013;41(Database issue):D590–D596. doi:10.1093/nar/gks1219
24. Lozupone C, Knight R. UniFrac: a new phylogenetic method for comparing microbial communities. Appl Environ Microbiol. 2005;71(12):8228–8235. doi:10.1128/AEM.71.12.8228-8235.2005
25. Ryan J, Pilkington L, Neuhaus K, Ritchie K, Ancelin ML, Saffery R. Investigating the epigenetic profile of the inflammatory gene IL-6 in late-life depression. BMC Psychiatry. 2017;17(1):354. doi:10.1186/s12888-017-1515-8
26. Charlton RA, Lamar M, Zhang A, et al. Associations between pro-inflammatory cytokines, learning, and memory in late-life depression and healthy aging. Int J Geriatr Psychiatry. 2018;33(1):104–112. doi:10.1002/gps.4686
27. Kim JM, Stewart R, Kim JW, et al. Changes in pro-inflammatory cytokine levels and late-life depression: a two year population based longitudinal study. Psychoneuroendocrinology. 2018;90:85–91. doi:10.1016/j.psyneuen.2018.02.006
28. Bremmer MA, Beekman AT, Deeg DJ, et al. Inflammatory markers in late-life depression: results from a population-based study. J Affect Disord. 2008;106(3):249–255. doi:10.1016/j.jad.2007.07.002
29. Kuo CY, Lin CH, Lane HY. Molecular basis of late-life depression. Int J Mol Sci. 2021;22(14):7421. doi:10.3390/ijms22147421
30. Zhang J, He H, Qiao Y, et al. Priming of microglia with IFN-gamma impairs adult hippocampal neurogenesis and leads to depression-like behaviors and cognitive defects. Glia. 2020;68(12):2674–2692. doi:10.1002/glia.23878
31. Buspavanich P, Adli M, Himmerich H, et al. Faster speed of onset of the depressive episode is associated with lower cytokine serum levels (IL-2, −4, −6, −10, TNF-alpha and IFN-gamma) in patients with major depression. J Psychiatr Res. 2021;141:287–292. doi:10.1016/j.jpsychires.2021.06.033
32. Papageorgiou IE, Lewen A, Galow LV, et al. TLR4-activated microglia require IFN-gamma to induce severe neuronal dysfunction and death in situ. Proc Natl Acad Sci U S A. 2016;113(1):212–217. doi:10.1073/pnas.1513853113
33. Jiang H, Ling Z, Zhang Y, et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav Immun. 2015;48:186–194. doi:10.1016/j.bbi.2015.03.016
34. Ye X, Wang D, Zhu H, et al. Gut microbiota changes in patients with major depressive disorder treated with vortioxetine. Front Psychiatry. 2021;12:641491. doi:10.3389/fpsyt.2021.641491
35. Lee SM, Dong TS, Krause-Sorio B, et al. The intestinal microbiota as a predictor for antidepressant treatment outcome in geriatric depression: a prospective pilot study. Int Psychogeriatr. 2022;34(1):33–45. doi:10.1017/S1041610221000120
36. Hasani A, Ebrahimzadeh S, Hemmati F, Khabbaz A, Hasani A, Gholizadeh P. The role of akkermansia muciniphila in obesity, diabetes and atherosclerosis. J Med Microbiol. 2021;70(10). doi:10.1099/jmm.0.001435
37. Peirce JM, Alvina K. The role of inflammation and the gut microbiome in depression and anxiety. J Neurosci Res. 2019;97(10):1223–1241. doi:10.1002/jnr.24476
38. Boehme M, van de Wouw M, Bastiaanssen TFS, et al. Mid-life microbiota crises: middle age is associated with pervasive neuroimmune alterations that are reversed by targeting the gut microbiome. Mol Psychiatry. 2020;25(10):2567–2583. doi:10.1038/s41380-019-0425-1
39. Wang J, Xu W, Wang R, Cheng R, Tang Z, Zhang M. The outer membrane protein Amuc_1100 of Akkermansia muciniphila promotes intestinal 5-HT biosynthesis and extracellular availability through TLR2 signalling. Food Funct. 2021;12(8):3597–3610. doi:10.1039/d1fo00115a
40. Cheung SG, Goldenthal AR, Uhlemann AC, Mann JJ, Miller JM, Sublette ME. Systematic review of gut microbiota and major depression. Front Psychiatry. 2019;10:34. doi:10.3389/fpsyt.2019.00034
41. Zheng P, Zeng B, Zhou C, et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol Psychiatry. 2016;21(6):786–796. doi:10.1038/mp.2016.44
42. Guillemin GJ, Brew BJ, Noonan CE, Takikawa O, Cullen KM. Indoleamine 2,3 dioxygenase and quinolinic acid immunoreactivity in Alzheimer’s disease hippocampus. Neuropathol Appl Neurobiol. 2005;31(4):395–404. doi:10.1111/j.1365-2990.2005.00655.x
43. Zhang X, Shi L, Sun T, Guo K, Geng S. Dysbiosis of gut microbiota and its correlation with dysregulation of cytokines in psoriasis patients. BMC Microbiol. 2021;21(1):78. doi:10.1186/s12866-021-02125-1
44. Wang H, Luo J, Chen X, et al. Clinical observation of the effects of oral opioid on inflammatory cytokines and gut microbiota in patients with moderate to severe cancer pain: a Retrospective Cohort Study. Pain Ther. 2022;11(2):667–681. doi:10.1007/s40122-022-00386-w
45. Lin H, An Y, Tang H, Wang Y. Alterations of bile acids and gut microbiota in obesity induced by high fat diet in rat model. J Agric Food Chem. 2019;67(13):3624–3632. doi:10.1021/acs.jafc.9b00249
46. Williams BL, Hornig M, Parekh T, Lipkin WI, Biron C. Application of novel PCR-based methods for detection, quantitation, and phylogenetic characterization of sutterella species in intestinal biopsy samples from children with autism and gastrointestinal disturbances. mBio. 2012;3(1). doi:10.1128/mBio.00261-11
47. Dong Z, Shen X, Hao Y, et al. Gut microbiome: a potential indicator for differential diagnosis of major depressive disorder and general anxiety disorder. Front Psychiatry. 2021;12:651536. doi:10.3389/fpsyt.2021.651536
48. Barandouzi ZA, Starkweather AR, Henderson WA, Gyamfi A, Cong XS. Altered composition of gut microbiota in depression: a systematic review. Front Psychiatry. 2020;11:541. doi:10.3389/fpsyt.2020.00541
49. Pinart M, Dotsch A, Schlicht K, et al. Gut microbiome composition in obese and non-obese persons: a systematic review and meta-analysis. Nutrients. 2021;14(1):12. doi:10.3390/nu14010012
50. Hou YP, He QQ, Ouyang HM, et al. Human gut microbiota associated with obesity in Chinese children and adolescents. Biomed Res Int. 2017;2017:7585989. doi:10.1155/2017/7585989
51. Zacarias MF, Collado MC, Gomez-Gallego C, et al. Pregestational overweight and obesity are associated with differences in gut microbiota composition and systemic inflammation in the third trimester. PLoS One. 2018;13(7):e0200305. doi:10.1371/journal.pone.0200305
52. Milaneschi Y, Simmons WK, van Rossum EFC, Penninx BW. Depression and obesity: evidence of shared biological mechanisms. Mol Psychiatry. 2019;24(1):18–33. doi:10.1038/s41380-018-0017-5
53. Hu S, Li A, Huang T, et al. Gut microbiota changes in patients with bipolar depression. Adv Sci. 2019;6(14):1900752. doi:10.1002/advs.201900752
54. Deng D, Su H, Song Y, et al. Altered fecal microbiota correlated with systemic inflammation in male subjects with methamphetamine use disorder. Front Cell Infect Microbiol. 2021;11:783917. doi:10.3389/fcimb.2021.783917
55. Ling Z, Jin G, Yan X, et al. Fecal dysbiosis and immune dysfunction in Chinese elderly patients with schizophrenia: an Observational Study. Front Cell Infect Microbiol. 2022;12:886872. doi:10.3389/fcimb.2022.886872
56. Carniel BP, da Rocha NS. Brain-derived neurotrophic factor (BDNF) and inflammatory markers: perspectives for the management of depression. Prog Neuropsychopharmacol Biol Psychiatry. 2021;108:110151. doi:10.1016/j.pnpbp.2020.110151
57. Wei Y, Wang T, Li G, et al. Investigation of systemic immune-inflammation index, neutrophil/high-density lipoprotein ratio, lymphocyte/high-density lipoprotein ratio, and monocyte/high-density lipoprotein ratio as indicators of inflammation in patients with schizophrenia and bipolar disorder. Front Psychiatry. 2022;13:941728. doi:10.3389/fpsyt.2022.941728
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.