")
Back to Journals » ImmunoTargets and Therapy » Volume 13
CD69 is a Promising Immunotherapy and Prognosis Prediction Target in Cancer
Authors Li Y , Gu Y , Yang P , Wang Y , Yu X , Li Y , Jin Z, Xu L
Received 12 September 2023
Accepted for publication 22 December 2023
Published 9 January 2024 Volume 2024:13 Pages 1—14
DOI https://doi.org/10.2147/ITT.S439969
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Flavio Salazar-Onfray
Yuchen Li,1,* Yinfeng Gu,1,* Pengyue Yang,1 Yan Wang,1 Xibao Yu,1 Yangqiu Li,1 Zhenyi Jin,1,2 Ling Xu1,3
1Institute of Hematology, School of Medicine, Key Laboratory for Regenerative Medicine of Ministry of Education, Department of Hematology, First Affiliated Hospital, Jinan University, Guangzhou, 510632, People’s Republic of China; 2Department of Pathology, School of Medicine, Jinan University, Guangzhou, 510632, People’s Republic of China; 3Guangdong Provincial Key Laboratory of Virology, Institute of Medical Microbiology, Jinan University, Guangzhou, 510632, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ling Xu; Zhenyi Jin, Email [email protected]; [email protected]
Abstract: Immunotherapy utilizing T cells that attack tumors is a promising strategy for treatment, but immune suppressive T cell subsets, such as regulatory T cell (Treg), and immune checkpoint molecules, including programmed death-1 (PD-1), can suppress the intensity of a T cell immune reaction and thereby impair tumor clearance. Cluster of differentiation 69 (CD69), known as an early leukocyte activation marker, can be used as a measure or early marker of T cell activation. In recent years, the functions of CD69 in the regulation of Treg/Th17 (T helper cell 17) differentiation and in the tissue retention of T cells have attracted considerable interest. These functions are related to the role of CD69 in immune suppression in tumor environments (TME). In this review, we first summarized current perspectives in the biological function of CD69 and demonstrated that CD69 acts as a regulator of T cell activation, differentiation, retention, and exhaustion. Then, we discussed recent advances in understanding of CD69 deficiency and anti-CD69 antibody administration and shed light on the value of targeting on CD69 for cancer immunotherapy and prognosis prediction.
Keywords: CD69, T cell, immunotherapy
Introduction
In the past two decades, monoclonal antibody (mAb) immunotherapy, immune checkpoint blockade therapy, and chimeric antigen receptor T cell (CAR-T) therapy have changed the therapeutic landscape of tumor patients. Numerous molecules with immunoregulatory functions have been discovered, which can be applied to cancer immunotherapy.
CD69 is a recognized classical marker of white blood cell activation.1 It appears faster on the cytoplasmic membranes of activated cells than CD252 and is thus regarded as an early marker of lymphocyte activation. CD69 can be expressed in hematopoietic or immune cells and plays a regulatory role. It can promote tissue residency, regulate Th17/Treg cell differentiation, and contribute to the exhaustion of resident memory T cells, especially in the tumor microenvironment.
Based on the various roles of CD69 in immune function regulation, targeting CD69 in the treatment of immune diseases and tumors seems to be a theoretically feasible strategy. The practical outcome and effectiveness of this approach have been considered to be of equal importance. Previous studies have tended to focus on the roles of CD69 in inducing Treg differentiation. Given that numerous studies have revealed its involvement in tissue residency and tumor immunity, the immunoregulatory roles of CD69 need to be reassessed. In this review, we summarized the basic expression, transcription regulation, and ligands of CD69, further introduced the immunomodulatory function of CD69 in regulating the balance of Treg/Th17, the retention of tissue-resident memory T (TRM) cells, and the exhaustion of T cells. In addition, we further reviewed the abnormal expression of CD69 in cancer and discussed the prognostic value and the possibility of targeting CD69 for cancer treatment, especially for cancer immunotherapy.
Expression of CD69 in Hematopoietic/Immune Cells
CD69 is a member of the differentiation antigen cluster previously called activation-inducing molecule (AIM),3 activation antigen (EA-1),4 leukocyte surface antigen (Leu-23),5 or MLR-3 antigen.6 Discovered in 1981,7 CD69 is a disulfide-linked homodimer protein with two differentially glycosylated subunits (28–32 kDa). Each subunit consists of an extracellular C-type lectin domain connected to a single-spanning transmembrane region followed by a short cytoplasmic tail. CD69 is broadly expressed on the surfaces of most hematopoietic lineages, including activated T and B lymphocytes, natural killer (NK) cells, murine macrophages, neutrophils, and eosinophils, and constitutively expressed on human monocytes, platelets, epidermal Langerhans cells, and bone-marrow myeloid precursors.1,8 However, the expression of CD69 cannot be detected in unstimulated peripheral blood leukocytes and activated monocytes.9 Recently, CD69 was found to be constitutively expressed in resident CD8+, CD4+, Treg, γδ T cells, innate lymphocytes, natural killer T cells in all the tissues explored, and skin dendritic cells (DCs).10,11 The expression of CD69 can lead to the internalization and degradation of sphingosine 1-phosphate receptor 1 (S1P1), which plays a role in cell egress from lymphoid tissues and enters the blood by binding with sphingosine-1-phosphate (S1P).10–12
Transcriptional Regulation of CD69 Expression
The CD69 gene is located on mouse chromosome 6 and human chromosome 12. This complex includes a variety of genes encoding C-type lectins, which play important roles in the immune system.13 The CD69 proximal promoter contains the canonical TATA box and binding sites for early-response inducible transcription factors, such as NF-κB, AP1, Erg-1, Erg-3, and ATF-3/CREB.14–17 Relative to the promoter, the 5′ region of CD69 contains four conserved noncoding sequences 1 to 4 (CNS1–4). These elements display DNase hypersensitivity and bear regulatory cell-type-specific histone marks that undergo changes during development and activation.13 CNS2 is the most intensively studied element. It is located 4 kb upstream of the human CD69 transcription start site, which is occupied by dense clusters of transcription factors, such as Oct1 and RUNX1,18,19 and it displays promoter and enhancer functions. As demonstrated by Jennifer et al, CNS2 is necessary not only to activate CD69 expression upon induction but also to sustain basal activity.16 This kind of basal activity may contribute to the sustained expression of CD69 in tissue-resident memory T cells (TRMs). As more immune regulation function of CD69 were found, the basic regulation mechanism of CD69 gene expression in different kind of immune cells needs further investigation.
Ligands of CD69
The functions of CD69 can be better understood by studying its ligands. To date, CD69 has four potential ligands: galactolectin-1 (Gal-1), calprotectin (S100A8/S100A9 complex), myosin light chains 9, 12a, and 12b (Myl9/12), and oxidized low-density lipoprotein (ox-LDL). Gal-1 is a β-galactoside-binding protein that can be expressed in macrophages and DCs10 and plays an important role in the regulation of adaptive immune responses by altering the fate and phenotypes of Th cells. IL-10 is the main component of Gal-1 immunomodulatory characteristics, and Gal-1-induced IL-10+ T cells can effectively inhibit T cell proliferation and the T cell-mediated inflammatory response.20 The expression of CD69 by activated T cells triggers an anti-inflammatory mechanism mediated by Gal-1 that can promote the production of cytokines, such as transforming growth factor-β (TGF-β) and IL-35, regulating the immune response and preventing the pathogenic Th17 response.21 Thus, CD69 regulates the key functions of murine and human Th17 effector cells by interacting with Gal-1.22 The S100A8/S100A9 complex belongs to the S100 protein family and is a natural ligand for CD69 in human peripheral blood mononuclear cells. It is expressed and secreted by bone marrow (BM) cells and has proinflammatory and anti-inflammatory effects. Glycosylation-dependent interactions between CD69 and S100A8/S100A9 complexes are required for the secretion of TGF-β and IL-4 and the transformation of naïve CD4+ T cells to Treg cells.23 Therefore, Gal-1 and S100A8/S100A9 can regulate T cell differentiation by upregulating Treg differentiation and suppressing Th17 differentiation. Myl9 and Myl12 belong to the myosin light chain molecular family. Myl9 can be produced by platelets and form Myl9 nets during inflammation, and CD69-expressing antigen-primed T cells attached to Myl9 nets can be recruited to inflamed tissues and cause antigen-specific T cells to remain in inflamed tissues.24 Increased secretion of IL-33 and TNF-α leads to the deterioration of inflammation. This finding suggests that the binding of Myl9 to CD69 can be responsible for the recruitment of lymphocytes and the regulation of inflammation. In addition, ox-LDL is a new ligand of CD69. CD69 can reduce the expression of IL-8 and IFN-γ in T cells by binding to oxLDL and internalizing the receptor. The CD69–oxLDL complex can promote the transcription of nuclear receptor subfamily 4 group A (NR4A) and further regulate the differentiation between Th17 and Treg, promoting affinity to Treg cells.25,26 In addition, the CD69–oxLDL complex can promote the activation of the nuclear factor of activated T cells (NFAT) in human T cells and further induce the expression of PD-1 and NR4A3, thereby negatively regulating the inflammatory response.27
The roles of CD69 can be mediated by different ligands. The binding of Gal-1, S100A8/S100A9 complexes, and ox-LDL to CD69 may be involved in the negative regulation of immune response, whereas the binding of Myl9/12 can lead to the positive development of inflammation. Overall, CD69 may play multiple roles in T cell-mediated immunoregulation by interacting with different ligands. The effects of the interaction of CD69 on T cells with the four ligands are depicted in Figure 1.
 |
Figure 1 T cell immune responses mediated by CD69 binding to ligands. (A). Binding of CD69 to its ligand causes negative immune regulation. CD69 expressed on T cells binds to Gal1/12 expressed by APC or a tumor cell, triggering GAL1/12-mediated anti-inflammatory responses and promoting the production of anti-inflammatory cytokines, such as IL-10, IL-35, and TGF-β, which inhibit T cell proliferation (Left). The S100AB/S100A9 complex expressed by bone marrow cells binds to CD69 on T cells and promote Treg differentiation (Middle). oxLDL, which is modified in vascular endothelial cells, binds to CD69 on T cells, increasing the production of proinflammatory cytokines such as IL-8 and IFN-γ and promoting the transcription of NR4A and Treg differentiation (Right). (B). CD69 promotes inflammation after binding to its ligand. Platelets produce Myl9/12 to form the Myl9/12 network on blood vessels. The network then serves as a platform for the migration of CD69 antigen-specific T cells into inflamed tissues and thereby promotes inflammatory responses. |
CD69 Plays a Role in the Regulation of Treg/Th17 Differentiation
T cell differentiation is primarily dependent on cytokines that stimulate the JAK/STAT signaling pathway, resulting in the expression of lineage-specific transcription factors and effector cytokines.10 Th17 and Treg cells represent two CD4+ T cell subsets that require TGF-β during their development but eventually differentiate into two different phenotypes with opposite activities. In contrast, TGF-β in the absence of IL-6 can activate STAT5, which can push cell differentiation away from Th17 and toward Treg cells. Conversely, TGF-β and IL-6 are required for Th17 differentiation, which is associated with the activation of STAT3 and retinoic acid-associated orphan receptor γt (RORγt).28 Therefore, Treg/Th17 differentiation is based on the STAT3/STAT5 balance.
CD69 can participate in Treg/Th17 differentiation by controlling the STAT3/STAT5 balance (Figure 2A). The cytoplasmic tail of CD69 is associated with the JAK3/STAT5 signaling pathway, and CD69 can inhibit JAK2/STAT3 signaling and thereby negatively regulate the transcription of RORγt and inhibit the differentiation of cells into the Th17 lineage. Additionally, CD69 can enhance the phosphorylation of the JAK3/STAT5 signaling pathway and promote Treg differentiation, and the addition of exogenous IL-2 to CD69-deficient cells restores STAT5 phosphorylation and inhibits Th17 differentiation.29 CD69 deficiency significantly enhances the inflammatory response in the asthmatic models of antigen-induced airway allergy and increases the Th17 response.30 Thus, CD69 can promote the differentiation of T cells to Tregs while inhibiting the differentiation toward Th17 cells and regulating T cell-mediated immunity.
 |
Figure 2 CD69 participates in the balance in Treg/Th17 differentiation. (A). oxLDL binding to CD69 promotes Treg differentiation by enhancing NRA4 (NR4A1 and NR4A3) expression (Left). Binding of S1P to its receptor S1P1R promotes the activation of the mTOR/HIF-1α and JAK2/pSTAT3 pathways, thereby promoting Th17 differentiation. CD69 can promote the internalization and degradation of S1P1R and activate the JAK/pSTAT5 pathway, thereby blocking S1P1R signaling and promoting Treg cell differentiation (Right). (B). The combination of CD69 and CD98-LAT1 complex can increase Trp/Leu uptake, amino acid uptake can promote the activation of the mTORC signaling pathway and thereby promote Th17 differentiation and inhibit Treg differentiation. Endogenous tryptophan photo-product, FICZ, which combined with AHR and promoted its nuclear entry, attenuated the inhibition of HIF-α by intracellular AHR, further promoted Th17 differentiation, and inhibited Treg differentiation. |
In addition to STAT3/STAT5, numerous transcription factors are involved in Treg/Th17 differentiation. Heat shock transcription factor 1 (HSF1) deficiency or inhibition impairs CD69+ Treg generation and colitis repression, highlighting that HSF1 can promote CD69 transcription and CD69+ Treg differentiation.31 NR4A is an important factor controlling Th17/Treg differentiation, and ox-LDL-CD69 enhances the expression of the transcription factor NR4A, especially NR4A3 and NR4A1, thereby promoting Treg cell differentiation26 (Figure 2A). Hypoxia-inducible factor-1α (HIF-1α) promotes Th17 differentiation while inhibiting Treg32 and is related to CD69 regulation in T cell differentiation (Figure 2A). The S1P1–mammalian target of rapamycin (mTOR)–HIF-1α axes is required for Th17/Th1 differentiation.33,34 Therefore, Treg generation induced by a pathway involving HIF-1α may could be promoted by CD69, which according to suppresses S1P1 activity. Notably, CD69 establishes lateral interactions with LAT1–CD98, which could activate the HIF-1α in the downstream mTOR signaling pathway thus leading to the Th17/Th1 development and prevents Treg differentiation10 (Figure 2B). Thus, CD69 can inhibit S1P1 activity while increasing LAT1–CD98, leading to different results in Treg/Th17 differentiation. Therefore, it can be speculated that CD69 plays distinct roles under different conditions. Overall, CD69 plays a role in the regulation of Treg/Th17 differentiation apart from acting as an activation marker. The crucial role of CD69 in immunosuppression is promoting CD69+ Treg differentiation. Undoubtedly, CD69 has become an important part of ongoing basic and clinical research into various kinds of inflammatory diseases. The mechanisms of CD69 in regulating Treg/Th17 balance are shown in Figure 2.
Expression of CD69 in TRM Cells
TRM cells constitute the heterogeneous populations of T cells with effector and memory T cell functions and express resident gene signatures that allow them to transport, reside, and patrol peripheral tissues and respond quickly to red flags for long-term effective immune protective effects. CD69 expression can be detected in most identified TRM cells, regardless of cellular origin (embryonic or BM circulating cells) and the presence or absence of antigens, suggesting that tissue-specific environmental cues may promote the retention or survival of tissue-resident immune cells by controlling CD69 expression.10 Notably, CD69− cells can be functionally present in the vascular compartments of the kidneys and liver, indicating that some T cells without CD69 also show tissue residency. Numerous phenotypes can be detected on TRM cells, including CD103, CD49a, and CD44, and can vary among tissue-derived TRM cells because of variations in tissue compartments.35,36 Inhibitory markers, such as PD-1 and CD101, are expressed on TRM surfaces, but whether their expression correlates with weak cytotoxicity and T cell exhaustion remains unclear.37,38 Reduced expression of receptors controlling the T cell egress,38 including CCR7, CD62L, and S1P1, contributes to the tissue residence of TRM cells. Thomas et al showed that CD69 expression by mucosal memory T cells is a feature of localization in tissues and is not age dependent; even at birth, memory T cells in all tissues express this core phenotypic marker that distinguishes them from memory T cells in blood.39
The S1P–S1P1 system controls the entry of T cells into lymphatic vessels. The S1P level is higher in blood than in intracellular or interstitial fluids, creating a steep S1P gradient that can facilitate the egress of lymphocytes from lymphoid organs, and the blockage of the S1P–S1P1 system results in a decrease in circulating lymphocytes.40 Similarly, a decreased S1P–S1P1 system is required for the tissue-resident ability of TRM cells. The transcriptional downregulation of S1P1 is required for the establishment of TRM cells, and this effect is induced by low Kruppel-like factor 2 (KLF2) expression.41 CD69 is associated with inhibition of S1P1 activity. This effect can be achieved by CD69, which enhances downstream IFN-α/β and decreases S1P1 activity.42 CD69 may form a complex with S1P1, leading to a conformational change in S1P1 and thereby facilitating its internalization and degradation.43 Low KLF2 transcription levels seem to correlate with CD69 expression in specific tissues and induce the tissue residency of T cells.44,45 These findings indicate the pivotal role of CD69 in TRM formation. CD69 negatively modulates S1P1 activity, thus leading to tissue residency. Given that KLF2 controls the transcription of S1P1 and CD62L, whether CD69 controls the expression of CD62L or other molecules related to T cell egress and the detailed mechanism by which CD69 induces tissue residency require further elucidation.46
HIF-1α has also been proven to promote TRM differentiation. The number of CD69+CD103+ cells showed a dose-dependent increase after treatment with the prolyl hydroxylase inhibitor FG-4592 (Roxadustat), given that HIF-1α can be hydroxylated by this enzyme.47 CD69 is a direct HIF-1α target gene responsible for hypoxia in tumor-infiltrating T lymphocytes (TILs).48 Thus, HIF-1α may participate in TRM formation by upregulating the CD69 expression, which may lead to TRM accumulation in the TME.
CD69 is a core marker of memory T cells in multiple tissue compartments. The role of CD69 in TRM formation remains unclear but may partially correlate with the transcription factors KLF2 and HIF-1α.
CD69 Expression on TILs is Associated with T Cell Exhaustion
In chronic infections with persistent antigen stimulation and in TMEs, CD8+ T cells are often in a special state: T cell exhaustion or dysfunction. The stratification loss of effector function and proliferation and significant transcription and metabolic changes are the distinguishing features of T cells in an exhausted state. The main marker of T cell exhaustion is the upregulation of PD-1, and other inhibitory receptors, such as cytotoxic T lymphocyte antigen-4 and T cell immunoglobulin and ITIM domains (TIGIT).49 The PD-1/PD-L1 axis is important for T cell dysfunction because it blocks PD-1 or its ligand, PD-L1, which restores T cell function.50 The transient expression of PD-1 is a characteristic of normal T cell activation, and sustained exposure to antigens induces the sustained expression of PD-1, which may drive T cell exhaustion.51
Research has demonstrated that CD69 expressed on TILs correlates with T cell exhaustion. In 2021, a study performed a transcriptomic analysis and found enhanced CD69 gene expression in pre-exhausted and exhausted CD8+ T cells isolated from colorectal cancer, hepatocellular carcinoma (HCC), and non-small cell lung cancer.52 Another study explored the process of T cell exhaustion in an animal chronic lymphocytic choriomeningitis virus infection model and found that Ly108 (Slamf6) and CD69 define a four-stage developmental trajectory of exhausted T cell subsets. One subset of terminally exhausted T cells hallmarked by Ly108−CD69+ exhibited poor division, limited persistence, and restrained developmental plasticity, and had high PD-1 and Thymocyte Selection Associated High Mobility Group Box (TOX) expression levels and apoptotic rates, while with low TCF1 expression levels. Another subset hallmarked by Ly108+CD69+ represents the progenitor of the other three exhaustion subsets including the terminal exhausted Ly108−CD69+ subset, which shows quiescent and resident features. Moreover, TILs with such phenotypes were also detected in human melanoma.53 In our previous work, we found an increased number of CD8+CD69+ TEM cells in the BM of patients with de novo chronic myeloid leukemia, and a higher percentage of those cells express PD-1 and TIGIT when compared with healthy individuals.54 Recently, similar results were also detected in our acute myeloid leukemia cohorts (Unpublished data). These pieces of evidence indicate that CD69 expression on TILs is associated with T cell exhaustion. Furthermore, a study found that tumor progression is attenuated in CD69-deficient mice bearing 4T1-luc2 tumor cells, and the antitumor cytokine production of TILs was enhanced with decreased levels of exhaustion55. The cytotoxicity of TILs is enhanced after targeting CD69, further revealing the possibility of T cell exhaustion induced by CD69.55,56
Despite the unclear underlying mechanism by which CD69 expression contributes to T cell exhaustion, possible CD69+ T cell exhaustion may be explained by the CD69–S1P1 complex formation. The negative regulation of S1P1 by CD69 is required for the tissue residence of T cells, which may expose the T cells to a persistent antigen encounter environment, which is one of the main causative agents in T cell dysfunction.57–59 Besides, there was evidence showing that CD69 could promote PD-1 expression in CD4+ T cells through the activation of NFAT1 when engaged with oxLDL, which plays a role in the modulation of inflammation and vascular remodeling in cardiovascular diseases. Whether this mechanism also contributes to the exhaustion of TILs remains to be elucidated in the future.
Another notable problem is that CD69 positive TRM cells are usually featured by expressing higher levels of a range of molecules commonly associated with terminally exhausted T cells (PD-1, CD39, and Tox) than other memory subsets, inversely, exhausted T cells could acquire tissue resident features of TRM cells in order to adapt the tissue microenvironment, especially in the TME. Thus, in situations where the antigen burden is both high and unwavering, the extensive overlap between exhausted T cell and TRM cell phenotypes may conflate the classification of either subset.60,61 In summary, CD69 plays an important role in the formation of exhausted T cells, but how CD69 regulates the dynamic formation of exhausted T cells and the transformation of TRM cells into exhausted cells or vice versa under different disease scenarios still needs to be further elucidated.
Expression of CD69 in Various Cancers as a Prognostic Marker
As CD69 has been recognized as one of the makers of TRM cells, based on the gene expression of CD69 and CD4/CD8A, Shota Ida et al, identified TRM cells correlate with the inflammatory tumor microenvironment and improved prognosis in head and neck squamous cell carcinoma.62 Bruni et al reported that the higher frequencies of CD69+ Vδ1 T cells in tissue sites and in peripheral blood are associated with slow liver metastatic progression and long overall survival in patients with colorectal cancer.63 Similarly, combined CD69+CD103+ and γδ-TCR gene signatures showed a favorable prognostic association with overall survival in HCC.64 Besides, a high abundance of stem like CD8 TRM cells CD3+CD8+CD103lowCD69+ has been associated with better patient outcomes in high-grade serous ovarian carcinomas.65 In melanoma, the increased numbers of CD69+CD103+ CD8+ TRM cells were associated with improved survival in immunotherapy-naïve patients.66 However, our recent work shows that the abundance of CD8+ TRM-like cells (CD69+ CD8+ TEM) in the BM of AML patients is associated with inferior outcomes (Unpublished data). Coincidentally, Bryant et al found that accumulation of atypical tissue resident CD69+ terminal effector cells (CD8+CD57+) in myeloma infiltrated BM could prevent differentiation and expansion of clonal myeloma specific CD8+ terminal effector cells and ultimately contribute to myeloma immune escape.67 These results indicated that the infiltrating of CD69+ TRM cell in solid tumors and hematological malignancies may have opposite prognostic values. Mechanisms under these different phenotypes remained to be investigated.
Except to express on tumor infiltrating TRM cells, the expression of CD69 on Tregs may also impact the survival of cancer patients. In the tumor tissue of HCC patients, an accumulation of the CD4+CD69+ Treg subpopulation has been identified. This particular subpopulation is characterized by the absence of foxp3 or CD25 expression but elevated levels of immune inhibitory molecules, including PD-1 and CTLA-4. The frequency of these cells exhibits a significant correlation with tumor progression, with a more pronounced impact observed in late-stage cancer patients. This observation suggests that this specific CD69+Treg subpopulation may serve as a valuable indicator of adverse prognosis in HCC.68 Even there was no other evidence shows that CD69+ Treg play an immunosuppression role in other tumor model or human cancer at present, based on its vital role in regulating Treg/Th17 balance and T cell residency, the importance of CD69+ Treg in human cancer tissues should be respected.
Except for expression on Treg and TRM which indirectly correlate the cancer prognosis, CD69 also has been found to express on malignant tumor and leukemia cells which have been found with potential value as a prognostic marker. A recent study has unveiled the expression of CD69 within the cells of ovarian clear cell carcinoma. The CD69 expressed in cancer cells facilitated the progression of carcinoma cell through its interaction with fibronectin, consequently contributing to an adverse prognosis for patients.69 Another study found a subset of leukemia stem cells with high CD69 expression levels corresponding to self-renewal and enhanced colony-forming capacity, which is responsible for the relapse of acute myeloid leukemia70,71 and poor and poor clinical outcomes.70 Moreover, CD69 expression was also detected on tumor cells from most other hematological malignancies and was associated with poor clinical outcome. For example, CD69 expression in B-cell non-Hodgkin’s lymphoma is associated with poor treatment response and prognosis, which might be related to increased sensitivity to external stimuli and the risk of DNA damage in the oncogenesis of CD69-positive cells.72 CD69 expression on chronic lymphocytic leukemia (CLL) is considered an independent prognostic marker because patients with CD69-positive CLL cells present a weaker response to chemotherapy and shorter overall survival than those with low CD69 expression levels.73 In addition, it was reported that CLL cases with high CD69 expression levels in lymph nodes and BM developed higher resistance to bendamustine than to peripheral blood tumor cells.74 All the above cancer progression or prognostic relevant studies about CD69 are summarized in Table 1.
 |
Table 1 CD69 Expression in Predicting Cancer Progression and Prognosis |
Although the underlying mechanism of CD69 in hematological malignancy development remains unclear, the negative prognostic outcome of patients with CD69-expressing tumor cells indicates the potential of CD69 as a therapeutic target, especially against hematological malignancies.
CD69 Target Therapy in Cancer Treatment
Cumulative evidence suggests that the utilization of anti-CD69 mAb is effective against several autoimmune diseases although CD69 is correlated with the amelioration of some kinds of autoimmune diseases and hypersensitivity.30,75,76 Anti-CD69 mAb effectively ameliorates diseases in animal experiments, including collagen-induced arthritis, airway inflammation, and diarrhea induced by food allergy.76–79 GFC-101, an anti-CD69 fully human mAb, is under preclinical study for the treatment of bowel irritable syndrome, rheumatoid arthritis, and respiration disorders.
Targeting CD69 in malignancies is being studied. As early as 2003, a study found that CD69−/− mice challenged with mutant lymphoma cells (RMA-S) and prostate cancer (RM-1) showed reduced tumor growth and metastases and prolonged survival time compared with wild-type mice. Decreased TGF-β production was also observed in CD69−/− mice, and the study verified the upregulation of TGF-β production mediated by CD69.80 In 2012, Zhao et al found that upon encountering autologous CD69+ T cells, tumor macrophages acquired the ability to produce large amounts of IDO protein in cancer nests; they proposed that such an active induction of immune tolerance should be considered for the rational design of effective immune-based anticancer therapies.81 Recently, Using the CD69-deficient mice, Ryo et al verified that CD69 deficiency attenuates TOX expression in tumor-specific CD8+ T cells in tumor-draining lymph nodes likely through an NFAT-dependent pathway, and thereby promotes the initiation of CD8+ T-cell differentiation, leading to enhanced anti-tumor immune responses. In addition, they also identified that combined treatment with anti-CD69 and anti-PD-1 had an enhanced therapeutic effect on the melanoma mice model when compared with administration anti-CD69 or anti-PD-1 alone.82
Other Studies using anti-CD69 antibodies against tumors also supported the possible therapeutic value of targeting CD69 (summarized in Table 2). A study found that the tumor volume of renal cell carcinoma-bearing mice was significantly decreased after injection with a DC-based vaccine and anti-CD69 antibody. This observed phenomenon may be attributed to the upregulation of CD69 expression on T cells following a DC-based vaccine. Subsequent stimulation of T cell activation and proliferation takes place through the cross-linking of CD69 on T cells by anti-CD69.55 In another study, the application of an anti-CD69 antibody can activate resting NK cells against RMA-S lymphoma and RM-1 prostatic carcinoma lung metastases through increased NK-cell cytolytic activity and IFN-γ production.83 Besides, in vivo treatment with anti-CD69 also showed a significant reduction in tumor growth and metastasis, a significant increase in the number of CD4+ and CD8+ TILs, and a reduction in the frequency of profoundly exhausted CD8+ T cells (PD-1hiTim3+), which indirectly suggested the therapeutic effect of anti-CD69.53 The above studies show that CD69 can act as a target against specific tumors by enhancing the anti-tumor activities of immune cells. Given the potential value of targeting CD69 in cancer immunotherapy, studies on anti-CD69 antibodies against different kinds of tumors in humanized animal models should be launched. While, CD69 is expressed on not only T cells but also almost every leukocyte, side effects from anti-CD69 administration may be unavoidable. Thus, the safety of treatment with a CD69 mAb needs to be further investigated. Evidence from Ryo’s CD69 deficiency mice showed that there was no severe defects occurred, thus they predict that anti-CD69 associated adverse events will be tolerable.82 However, strong evidence showed that CD69 plays a role in the balance of Treg/Th17, possible clinical trials targeting the inhibition of CD69 should caution of the risk of inducing autoimmune diseases like asthma, myocarditis, colitis, contact dermatitis, and rheumatoid arthritis and even other side effects.84 Moreover, the use of mAb itself has a certain risk, which may produce cytokine storm or other adverse reactions.85–87 Therefore, more preclinical experiments are needed to verify the safety of CD69 mAb for clinical treatment.
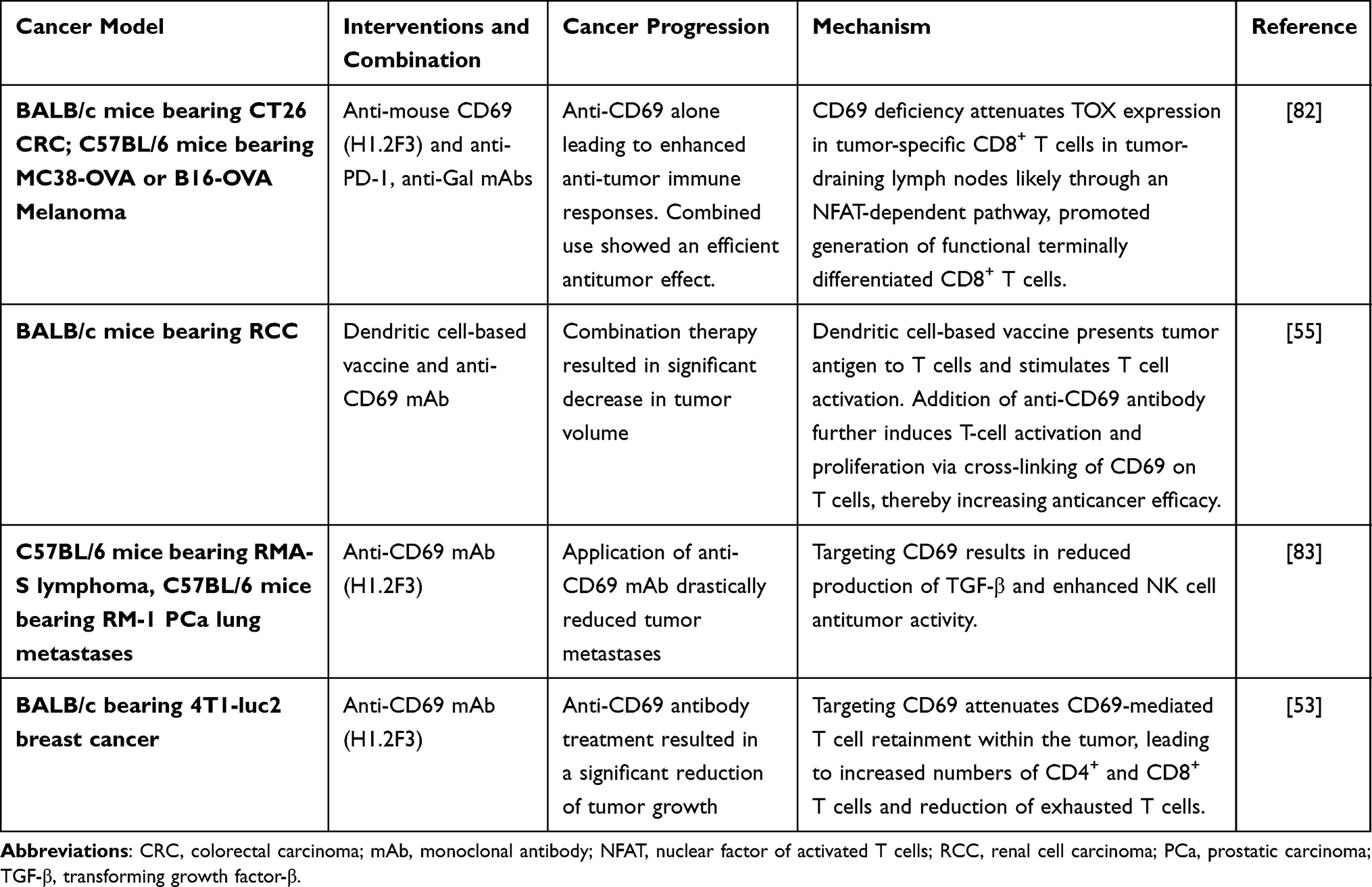 |
Table 2 Researches Based on Anti-CD69 Antibody for Cancer Treatment |
The mechanism of various immune regulatory functions involved in CD69 is inseparable from the relationship between its ligands, so it may be a new therapeutic idea to target the CD69 ligands alone or at the same time for cancer immunotherapy. Gal-1 in particular, is known to be deeply involved in the initiation, amplification, and resolution of inflammatory responses. A growing body of evidence indicates that Gal-1 can suppress the T-cell response through apoptotic and non-apoptotic mechanisms by binding to the glycoproteins (including CD2, CD3, CD7, CD43, CD45 as well as CD69) on T cells. In addition, the expression or overexpression of Gal-1 in various tumors or surrounding tissues is considered to be a sign of malignant tumor progression and, consequently, of a poor prognosis for patients.88 It has been reported that the blockade of the biological activity of Gal-1 in melanoma tissue results in a reduced tumor mass and stimulates the generation of a tumor-specific T-cell.89,90 Taken together, all observations support that Gal-1 may contribute to the immune escape of tumors by modulating the survival or polarization of effector T cells. Given the therapeutic benefits associated with Gal-1 blockade, several Gal-1 inhibitors have been designed and evaluated in pre-clinical tumor models.88,91 Exception for Gal-1, other ligands of CD69 including Myl9, S100A8, and S100A9 also have been found with the immune microenvironment modulation and tumor promotion ability in different cancers.92–95 For example, it has been reported that Myl9 expression is increased in human esophageal squamous cell carcinoma and that Myl9 expression is associated with immune infiltration.96,97 Thus, targeting these targets for cancer therapy also should be considered. However, the interaction between CD69 and the above ligands are mostly explored in inflammatory settings, therefore, it is still necessary to further explore how CD69 and its ligands crosstalk in anti-cancer immunotherapy settings in the future studies.
Conclusion and Prospects
In earlier studies, CD69 was defined as an activation marker of different leukocyte subsets, especially for T and NK cells, while it was found to be involved in the pathogenesis of several chronic inflammatory diseases. Recent studies have shown that CD69 can regulate the differentiation of Th17/Treg cells toward Treg cells, and the expression of CD69 by TRM cells may contribute to their tumor tissue retention and further exhaustion. Besides, anti-CD69 antibody application enhances antitumor effects by activating innate and innate and adaptive T cells, as shown in pre-clinical studies.12 These evidences indicate that CD69 may play an immune inhibitory role in antitumor immunity. Thus, CD69 could be considered a target molecule for cancer immunotherapy. However, opposing evidence indicates that CD69 promotes the balance of Th17/Treg cells into Th17 cells, and some patients with CD69+ immune cell infiltration within some kinds of tumors (bladder cancer, gastric cancer, lung cancer, and melanoma) also have a positive prognosis.38 Therefore, the feasibility of targeting CD69 for cancer immunotherapy should be based on the following points: (i) to establish the correlation between CD69+ immune cells and the prognosis in different kinds of tumor; (ii) to ensure the functions of CD69+ cells in TMEs, especially for TRM cells; (iii) to confirm whether anti-CD69 mAb produces effective cancer suppression effect in humanized animal models; and (iv) to verify the therapeutic value in clinical trials. Further studies will allow people to establish whether targeting CD69 is effective against tumors and produce a possible value of targeting this molecule in clinical cancer treatment.
In addition, CD69 expression on TILs and tumor cells associated with clinical prognosis establishes the independent prognostic value of CD69 in cancer. Studies need to further demonstrate whether CD69 relates to clinical outcomes in different cancers, providing information about the development of specific cancers and CD69 function. Further research into CD69 will shed light on the unique prognostic and therapeutic value of CD69 and its benefits to the clinical treatment of cancer.
Abbreviation
BM, bone marrow; CD69, cluster of differentiation 69; CLL, chronic lymphocytic leukemia; CNS, conserved noncoding sequences; DCs, dendritic cells; Gal-1, galactolectin-1; HIF-1α, hypoxia-inducible factor-1α; HCC, hepatocellular carcinoma; KLF2, kruppel-like factor 2; mAb, monoclonal antibody; Myl12, myosin light chain 12; Myl9, myosin light chain 9; Myl9/12, myosin light chain 9, 12a, 12b; NFAT, nuclear factor of activated T cell; NK, natural killer cell; NR4A, nuclear receptor subfamily 4 group A; ox-LDL, oxidized low-density lipoprotein; PD-1, programmed cell death-1; S1P, sphingosine-1-phosphate; S1P1, sphingosine1-phosphate receptor 1; TGF-β, transforming growth factor-β; Th17, T helper cell 17; TILs, tumor-infiltrating T cell; TME, tumor microenvironment; TOX, thymocyte selection associated high mobility group box; Treg, regulatory T cell; TRM, tissue resident memory T cell.
Acknowledgment
Yuchen Li and Yinfeng Gu contributed equally to this work and should be considered joint first authors.
Funding
This work is supported by grants from the National Natural Science Foundation of China (Nos. 82000108, 82293630, 82293632, 82070152); Guangdong Basic and Applied Basic Research Foundation (2020A1515110310); The Science and Technology Program of Guangzhou City (No. 202201010164); the Natural Science Foundation of Guangdong Province (No. 2023A1515030271) and the National Innovation and Entrepreneurship Training Program For Undergraduate (No. CX23386).
Disclosure
The authors declare that they have no competing interests.
References
1. Testi R, D’Ambrosio D, De Maria R, Santoni A. The CD69 receptor: a multipurpose cell-surface trigger for hematopoietic cells. Immunol Today. 1994;15(10):479–483.
2. González-Amaro R, Cortés JR, Sánchez-Madrid F, Martín P. Is CD69 an effective brake to control inflammatory diseases? Trends Mol Med. 2013;19(10):625–632.
3. Cebrián M, Yagüe E, Rincón M, López-Botet M, de Landázuri MO, Sánchez-Madrid F. Triggering of T cell proliferation through AIM, an activation inducer molecule expressed on activated human lymphocytes. J Exp Med. 1988;168(5):1621–1637. doi:10.1084/jem.168.5.1621
4. Hara T, Jung LK, Bjorndahl JM, Fu SM. Human T cell activation. III. Rapid induction of a phosphorylated 28 kD/32 kD disulfide-linked early activation antigen (EA 1) by 12-o-tetradecanoyl phorbol-13-acetate, mitogens, and antigens. J Exp Med. 1986;164(6):1988–2005. doi:10.1084/jem.164.6.1988
5. Lanier LL, Buck DW, Rhodes L, et al. Interleukin 2 activation of natural killer cells rapidly induces the expression and phosphorylation of the Leu-23 activation antigen. J Exp Med. 1988;167(5):1572–1585. doi:10.1084/jem.167.5.1572
6. Cosulich ME, Rubartelli A, Risso A, Cozzolino F, Bargellesi A. Functional characterization of an antigen involved in an early step of T-cell activation. Proc Natl Acad Sci U S A. 1987;84(12):4205–4209. doi:10.1073/pnas.84.12.4205
7. Qinghua L, Li W, Keqiang W. Advances in the application of CD69 molecules in γδ τT cells. Chin J Biomed Eng. 2016;22:337–339.
8. Marzio R, Mauël J, Betz-Corradin S. CD69 and regulation of the immune function. Immunopharmacol Immunotoxicol. 1999;21(3):565–582. doi:10.3109/08923979909007126
9. Bieber T, Rieger A, Stingl G, Sander E, Wanek P, Strobel I. CD69, an early activation antigen on lymphocytes, is constitutively expressed by human epidermal Langerhans cells. J Invest Dermatol. 1992;98(5):771–776. doi:10.1111/1523-1747.ep12499948
10. Cibrián D, Sánchez-Madrid F. CD69: from activation marker to metabolic gatekeeper. Eur J Immunol. 2017;47(6):946–953. doi:10.1002/eji.201646837
11. Mackay LK, Kallies A. Transcriptional Regulation of Tissue-Resident Lymphocytes. Trends Immunol. 2017;38(2):94–103.
12. Koyama-Nasu R, Wang Y, Hasegawa I, Endo Y, Nakayama T, Kimura MY. The cellular and molecular basis of CD69 function in anti-tumor immunity. Int Immunol. 2022;34(11):555–561. doi:10.1093/intimm/dxac024
13. Vazquez BN, Laguna T, Carabana J, Krangel MS, Lauzurica P. CD69 gene is differentially regulated in T and B cells by evolutionarily conserved promoter-distal elements. J Immunol. 2009;183(10):6513–6521.
14. López-Cabrera M, Muñoz E, Blázquez MV, Ursa MA, Santis AG, Sánchez-Madrid F. Transcriptional regulation of the gene encoding the human C-type lectin leukocyte receptor AIM/CD69 and functional characterization of its tumor necrosis factor-alpha-responsive elements. J Biol Chem. 1995;270(37):21545–21551. doi:10.1074/jbc.270.37.21545
15. Santis AG, López-Cabrera M, Hamann J, Strauss M, Sánchez-Madrid F. Structure of the gene coding for the human early lymphocyte activation antigen CD69: a C-type lectin receptor evolutionarily related with the gene families of natural killer cell-specific receptors. Eur J Immunol. 1994;24(7):1692–1697. doi:10.1002/eji.1830240735
16. Redondo-Antón J, Fontela MG, Notario L, et al. Functional characterization of a dual enhancer/promoter regulatory element leading human CD69 expression. Front Genet. 2020;11:552949. doi:10.3389/fgene.2020.552949
17. Castellanos Mdel C, López-Giral S, López-Cabrera M, de Landázuri MO. Multiple cis-acting elements regulate the expression of the early T cell activation antigen CD69. Eur J Immunol. 2002;32(11):3108–3117.
18. Laguna T, Notario L, Pippa R, et al. New insights on the transcriptional regulation of CD69 gene through a potent enhancer located in the conserved non-coding sequence 2. Mol Immunol. 2015;66(2):171–179. doi:10.1016/j.molimm.2015.02.031
19. Fontela MG, Notario L, Alari-Pahissa E, Lorente E, Lauzurica P. The Conserved Non-Coding Sequence 2 (CNS2) Enhances CD69 Transcription through Cooperation between the Transcription Factors Oct1 and RUNX1. Genes. 2019;10(9):651. doi:10.3390/genes10090651
20. Cedeno-Laurent F, Opperman M, Barthel SR, Kuchroo VK, Dimitroff CJ. Galectin-1 triggers an immunoregulatory signature in Th cells functionally defined by IL-10 expression. J Immunol. 2012;188(7):3127–3137. doi:10.4049/jimmunol.1103433
21. Adamson AS, Collins K, Laurence A, O’Shea JJ. The Current STATus of lymphocyte signaling: new roles for old players. Curr Opin Immunol. 2009;21(2):161–166. doi:10.1016/j.coi.2009.03.013
22. de la Fuente H, Cruz-Adalia A, Martinez Del Hoyo G, et al. The leukocyte activation receptor CD69 controls T cell differentiation through its interaction with galectin-1. Mol Cell Biol. 2014;34(13):2479–2487. doi:10.1128/MCB.00348-14
23. Lin CR, Wei TY, Tsai HY, Wu YT, Wu PY, Chen ST. Glycosylation-dependent interaction between CD69 and S100A8/S100A9 complex is required for regulatory T-cell differentiation. FASEB j. 2015;29(12):5006–5017. doi:10.1096/fj.15-273987
24. Kimura MY, Koyama-Nasu R, Yagi R, Nakayama T. A new therapeutic target: the CD69-Myl9 system in immune responses. Semin Immunopathol. 2019;41(3):349–358. doi:10.1007/s00281-019-00734-7
25. Jiménez-Fernández M, de la Fuente H, Martín P, Cibrián D, Sánchez-Madrid F. Unraveling CD69 signaling pathways, ligands and laterally associated molecules. Excli j. 2023;22:334–351. doi:10.17179/excli2022-5751
26. Tsilingiri K, de la Fuente H, Relaño M, et al. Oxidized low-density lipoprotein receptor in lymphocytes prevents atherosclerosis and predicts subclinical disease. Circulation. 2019;139(2):243–255. doi:10.1161/CIRCULATIONAHA.118.034326
27. Jiménez-Fernández M, Rodríguez-Sinovas C, Cañes L, et al. CD69-oxLDL ligand engagement induces Programmed Cell Death 1 (PD-1) expression in human CD4 + T lymphocytes. Cell Mol Life Sci. 2022;79(8):468. doi:10.1007/s00018-022-04481-1
28. Barbi J, Pardoll D, Pan F. Metabolic control of the Treg/Th17 axis. Immunol Rev. 2013;252(1):52–77. doi:10.1111/imr.12029
29. Martín P, Gómez M, Lamana A, et al. CD69 association with Jak3/Stat5 proteins regulates Th17 cell differentiation. Mol Cell Biol. 2010;30(20):4877–4889. doi:10.1128/MCB.00456-10
30. Martín P, Gómez M, Lamana A, et al. The leukocyte activation antigen CD69 limits allergic asthma and skin contact hypersensitivity. J Allergy Clin Immunol. 2010;126(2):355–365.e351–353. doi:10.1016/j.jaci.2010.05.010
31. Yu L, Zhou B, Zhu Y, et al. HSF1 promotes CD69(+) Treg differentiation to inhibit colitis progression. Theranostics. 2023;13(6):1892–1905. doi:10.7150/thno.78078
32. Pollizzi KN, Powell JD. Integrating canonical and metabolic signalling programmes in the regulation of T cell responses. Nat Rev Immunol. 2014;14(7):435–446. doi:10.1038/nri3701
33. Liu G, Yang K, Burns S, Shrestha S, Chi H. The S1P(1)-mTOR axis directs the reciprocal differentiation of T(H)1 and T(reg) cells. Nat Immunol. 2010;11(11):1047–1056. doi:10.1038/ni.1939
34. Dang EV, Barbi J, Yang HY, et al. Control of T(H)17/T(reg) balance by hypoxia-inducible factor 1. Cell. 2011;146(5):772–784. doi:10.1016/j.cell.2011.07.033
35. Cheuk S, Schlums H, Gallais Sérézal I, et al. CD49a expression defines tissue-resident CD8(+) T Cells poised for cytotoxic function in human skin. Immunity. 2017;46(2):287–300. doi:10.1016/j.immuni.2017.01.009
36. Mueller SN, Mackay LK. Tissue-resident memory T cells: local specialists in immune defence. Nat Rev Immunol. 2016;16(2):79–89. doi:10.1038/nri.2015.3
37. Kumar BV, Ma W, Miron M, et al. Human tissue-resident memory T cells are defined by core transcriptional and functional signatures in lymphoid and mucosal sites. Cell Rep. 2017;20(12):2921–2934. doi:10.1016/j.celrep.2017.08.078
38. Okła K, Farber DL, Zou W. Tissue-resident memory T cells in tumor immunity and immunotherapy. J Exp Med. 2021;218(4). doi:10.1084/jem.20201605
39. Connors TJ, Matsumoto R, Verma S, et al. Site-specific development and progressive maturation of human tissue-resident memory T cells over infancy and childhood. Immunity. 2023;56(8):1894–1909.e5. doi:10.1016/j.immuni.2023.06.008
40. Obinata H, Hla T. Sphingosine 1-phosphate and inflammation. Int Immunol. 2019;31(9):617–625. doi:10.1093/intimm/dxz037
41. Skon CN, Lee JY, Anderson KG, Masopust D, Hogquist KA, Jameson SC. Transcriptional downregulation of S1pr1 is required for the establishment of resident memory CD8+ T cells. Nat Immunol. 2013;14(12):1285–1293. doi:10.1038/ni.2745
42. Shiow LR, Rosen DB, Brdicková N, et al. CD69 acts downstream of interferon-alpha/beta to inhibit S1P1 and lymphocyte egress from lymphoid organs. Nature. 2006;440(7083):540–544. doi:10.1038/nature04606
43. Bankovich AJ, Shiow LR, Cyster JG. CD69 suppresses sphingosine 1-phosophate receptor-1 (S1P1) function through interaction with membrane helix 4. J Biol Chem. 2010;285(29):22328–22337. doi:10.1074/jbc.M110.123299
44. Mackay LK, Braun A, Macleod BL, et al. Cutting edge: CD69 interference with sphingosine-1-phosphate receptor function regulates peripheral T cell retention. J Immunol. 2015;194(5):2059–2063. doi:10.4049/jimmunol.1402256
45. Walsh DA, Borges da Silva H, Beura LK, et al. The functional requirement for CD69 in establishment of resident memory CD8(+) T cells varies with tissue location. J Immunol. 2019;203(4):946–955. doi:10.4049/jimmunol.1900052
46. Cowman SJ, Koh MY. Revisiting the HIF switch in the tumor and its immune microenvironment. Trends Cancer. 2022;8(1):28–42. doi:10.1016/j.trecan.2021.10.004
47. Hasan F, Chiu Y, Shaw RM, Wang J, Yee C. Hypoxia acts as an environmental cue for the human tissue-resident memory T cell differentiation program. JCI Insight. 2021;6(10). doi:10.1172/jci.insight.138970
48. Thommen DS, Schumacher TN. T cell dysfunction in cancer. Cancer Cell. 2018;33(4):547–562. doi:10.1016/j.ccell.2018.03.012
49. Day CL, Kaufmann DE, Kiepiela P, et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature. 2006;443(7109):350–354. doi:10.1038/nature05115
50. Ahmadzadeh M, Johnson LA, Heemskerk B, et al. Tumor antigen-specific CD8 T cells infiltrating the tumor express high levels of PD-1 and are functionally impaired. Blood. 2009;114(8):1537–1544. doi:10.1182/blood-2008-12-195792
51. Bai Y, Hu M, Chen Z, Wei J, Du H. Single-Cell transcriptome analysis reveals RGS1 as a new marker and promoting factor for T-cell exhaustion in multiple cancers. Front Immunol. 2021;12:767070. doi:10.3389/fimmu.2021.767070
52. Beltra JC, Manne S, Abdel-Hakeem MS, et al. Developmental relationships of four exhausted CD8(+) T cell subsets reveals underlying transcriptional and epigenetic landscape control mechanisms. Immunity. 2020;52(5):825–841.e828. doi:10.1016/j.immuni.2020.04.014
53. Mita Y, Kimura MY, Hayashizaki K, et al. Crucial role of CD69 in anti-tumor immunity through regulating the exhaustion of tumor-infiltrating T cells. Int Immunol. 2018;30(12):559–567. doi:10.1093/intimm/dxy050
54. Yao D, Lai J, Lu Y, et al. Comprehensive analysis of the immune pattern of T cell subsets in chronic myeloid leukemia before and after TKI treatment. Front Immunol. 2023;14:1078118. doi:10.3389/fimmu.2023.1078118
55. Wei SM, Pan HL, Wang L, et al. Combination therapy with dendritic cell-based vaccine and anti-CD69 antibody enhances antitumor efficacy in renal cell carcinoma-bearing mice. Turk J Med Sci. 2017;47(2):658–667. doi:10.3906/sag-1601-198
56. Schietinger A, Philip M, Krisnawan VE, et al. Tumor-specific T cell dysfunction is a dynamic antigen-driven differentiation program initiated early during tumorigenesis. Immunity. 2016;45(2):389–401. doi:10.1016/j.immuni.2016.07.011
57. Freeman CM, Han MK, Martinez FJ, et al. Cytotoxic potential of lung CD8(+) T cells increases with chronic obstructive pulmonary disease severity and with in vitro stimulation by IL-18 or IL-15. J Immunol. 2010;184(11):6504–6513. doi:10.4049/jimmunol.1000006
58. Gamradt P, Laoubi L, Nosbaum A, et al. Inhibitory checkpoint receptors control CD8(+) resident memory T cells to prevent skin allergy. J Allergy Clin Immunol. 2019;143(6):2147–2157.e2149. doi:10.1016/j.jaci.2018.11.048
59. Wang Z, Wang S, Goplen NP, et al. PD-1(hi) CD8(+) resident memory T cells balance immunity and fibrotic sequelae. Sci Immunol. 2019;4(36). doi:10.1126/sciimmunol.aaw1217
60. Gebhardt T, Park SL, Parish IA. Stem-like exhausted and memory CD8(+) T cells in cancer. Nat Rev Cancer. 2023;23(11):780–798. doi:10.1038/s41568-023-00615-0
61. Schenkel JM, Pauken KE. Localization, tissue biology and T cell state - implications for cancer immunotherapy. Nat Rev Immunol. 2023;23(12):807–823. doi:10.1038/s41577-023-00884-8
62. Ida S, Takahashi H, Kawabata-Iwakawa R, Mito I, Tada H, Chikamatsu K. Tissue-resident memory T cells correlate with the inflammatory tumor microenvironment and improved prognosis in head and neck squamous cell carcinoma. Oral Oncol. 2021;122:105508. doi:10.1016/j.oraloncology.2021.105508
63. Bruni E, Cimino MM, Donadon M, et al. Intrahepatic CD69(+)Vδ1 T cells re-circulate in the blood of patients with metastatic colorectal cancer and limit tumor progression. J Immunother Cancer. 2022;10(7). doi:10.1136/jitc-2022-004579
64. Zakeri N, Hall A, Swadling L, et al. Characterisation and induction of tissue-resident gamma delta T-cells to target hepatocellular carcinoma. Nat Commun. 2022;13(1):1372. doi:10.1038/s41467-022-29012-1
65. Anadon CM, Yu X, Hänggi K, et al. Ovarian cancer immunogenicity is governed by a narrow subset of progenitor tissue-resident memory T cells. Cancer Cell. 2022;40(5):545–557.e513. doi:10.1016/j.ccell.2022.03.008
66. Edwards J, Wilmott JS, Madore J, et al. CD103(+) tumor-resident CD8(+) T cells are associated with improved survival in immunotherapy-naïve melanoma patients and expand significantly during anti-PD-1 treatment. Clin Cancer Res. 2018;24(13):3036–3045. doi:10.1158/1078-0432.CCR-17-2257
67. Bryant C, Lau KHA, Vuckovic S, et al. Accumulation of CD69+ Terminal Effector CD8+ T cells occurs in the bone marrow of newly diagnosed Myeloma patients who lack protective clonal Vb expanded cytotoxic T cells. Clin Lymphoma Myeloma Leukemia. 2019;19(10):e29. doi:10.1016/j.clml.2019.09.043
68. Han Y, Yang Y, Chen Z, et al. Human hepatocellular carcinoma-infiltrating CD4+CD69+Foxp3⁻ regulatory T cell suppresses T cell response via membrane-bound TGF-β1. J Mol Med. 2014;92(5):539–550. doi:10.1007/s00109-014-1143-4
69. Koizume S, Kanayama T, Kimura Y, et al. Cancer cell-derived CD69 induced under lipid and oxygen starvation promotes ovarian cancer progression through fibronectin. Cancer Sci. 2023;114(6):2485–2498. doi:10.1111/cas.15774
70. Zhang Y, Jiang S, He F, et al. Single-cell transcriptomics reveals multiple chemoresistant properties in leukemic stem and progenitor cells in pediatric AML. Genome Biol. 2023;24(1):199. doi:10.1186/s13059-023-03031-7
71. Antony ML, Chang D, Noble-Orcutt KE, et al. CD69 marks a subpopulation of acute myeloid leukemia with enhanced colony forming capacity and a unique signaling activation state. Leuk Lymphoma. 2023;64(7):1262–1274. doi:10.1080/10428194.2023.2207698
72. Erlanson M, Grönlund E, Löfvenberg E, Roos G, Lindh J. Expression of activation markers CD23 and CD69 in B-cell non-Hodgkin’s lymphoma. Eur J Haematol. 1998;60(2):125–132. doi:10.1111/j.1600-0609.1998.tb01009.x
73. Del Poeta G, Del Principe MI, Zucchetto A, et al. CD69 is independently prognostic in chronic lymphocytic leukemia: a comprehensive clinical and biological profiling study. Haematologica. 2012;97(2):279–287. doi:10.3324/haematol.2011.052829
74. Montraveta A, Lee-Vergés E, Roldán J, et al. CD69 expression potentially predicts response to bendamustine and its modulation by ibrutinib or idelalisib enhances cytotoxic effect in chronic lymphocytic leukemia. Oncotarget. 2016;7(5):5507–5520. doi:10.18632/oncotarget.6685
75. Radulovic K, Manta C, Rossini V, et al. CD69 regulates type I IFN-induced tolerogenic signals to mucosal CD4 T cells that attenuate their colitogenic potential. J Immunol. 2012;188(4):2001–2013. doi:10.4049/jimmunol.1100765
76. Sancho D, Gómez M, Martinez Del Hoyo G, et al. CD69 targeting differentially affects the course of collagen-induced arthritis. J Leukoc Biol. 2006;80(6):1233–1241. doi:10.1189/jlb.1205749
77. Miki-Hosokawa T, Hasegawa A, Iwamura C, et al. CD69 controls the pathogenesis of allergic airway inflammation. J Immunol. 2009;183(12):8203–8215. doi:10.4049/jimmunol.0900646
78. Wang HY, Dai Y, Wang JL, Yang XY, Jiang XG. Anti-CD69 monoclonal antibody treatment inhibits airway inflammation in a mouse model of asthma. J Zhejiang Univ Sci B. 2015;16(7):622–631. doi:10.1631/jzus.B1400285
79. Zaim E, Ashino S, Osaka T, Yanagisawa N, Yagi J. Effect of cholecalciferol in food allergy mouse model is associated with decrease of CD69(+) CD4(+) T cells. J Nutr Sci Vitaminol. 2019;65(2):113–122. doi:10.3177/jnsv.65.113
80. Esplugues E, Sancho D, Vega-Ramos J, et al. Enhanced antitumor immunity in mice deficient in CD69. J Exp Med. 2003;197(9):1093–1106. doi:10.1084/jem.20021337
81. Zhao Q, Kuang DM, Wu Y, et al. Activated CD69+ T cells foster immune privilege by regulating IDO expression in tumor-associated macrophages. J Immunol. 2012;188(3):1117–1124. doi:10.4049/jimmunol.1100164
82. Koyama-Nasu R, Kimura MY, Kiuchi M, et al. CD69 imposes tumor-specific CD8+ T-cell Fate in tumor-draining lymph nodes. Cancer Immunol Res. 2023;11(8):1085–1099. doi:10.1158/2326-6066.CIR-22-0406
83. Esplugues E, Vega-Ramos J, Cartoixà D, et al. Induction of tumor NK-cell immunity by anti-CD69 antibody therapy. Blood. 2005;105(11):4399–4406. doi:10.1182/blood-2004-10-3854
84. Martín P, Blanco-Domínguez R, Sánchez-Díaz R. Novel human immunomodulatory T cell receptors and their double-edged potential in autoimmunity, cardiovascular disease and cancer. Cell Mol Immunol. 2021;18(4):919–935. doi:10.1038/s41423-020-00586-4
85. Suntharalingam G, Perry MR, Ward S, et al. Cytokine storm in a Phase 1 trial of the anti-CD28 monoclonal antibody TGN1412. N Engl J Med. 2006;355(10):1018–1028. doi:10.1056/NEJMoa063842
86. Hansel TT, Kropshofer H, Singer T, Mitchell JA, George AJ. The safety and side effects of monoclonal antibodies. Nat Rev Drug Discov. 2010;9(4):325–338. doi:10.1038/nrd3003
87. Pallardy M. Toxic effects and use of therapeutic monoclonal antibodies. Med Sci. 2009;25(12):1130–1134.
88. Camby I, Le Mercier M, Lefranc F, Kiss R. Galectin-1: a small protein with major functions. Glycobiology. 2006;16(11):137R–157R. doi:10.1093/glycob/cwl025
89. Rubinstein N, Alvarez M, Zwirner NW, et al. Targeted inhibition of galectin-1 gene expression in tumor cells results in heightened T cell-mediated rejection; A potential mechanism of tumor-immune privilege. Cancer Cell. 2004;5(3):241–251. doi:10.1016/S1535-6108(04)00024-8
90. Chou FC, Chen HY, Kuo CC, Sytwu HK. Role of Galectins in Tumors and in Clinical Immunotherapy. Int J Mol Sci. 2018;19(2):430. doi:10.3390/ijms19020430
91. Pérez Sáez JM, Hockl PF, Cagnoni AJ, et al. Characterization of a neutralizing anti-human galectin-1 monoclonal antibody with angioregulatory and immunomodulatory activities. Angiogenesis. 2021;24(1):1–5. doi:10.1007/s10456-020-09749-3
92. Wagner NB, Weide B, Gries M, et al. Tumor microenvironment-derived S100A8/A9 is a novel prognostic biomarker for advanced melanoma patients and during immunotherapy with anti-PD-1 antibodies. J Immunother Cancer. 2019;7(1):343. doi:10.1186/s40425-019-0828-1
93. Chen Y, Ouyang Y, Li Z, Wang X, Ma J. S100A8 and S100A9 in Cancer. Biochim Biophys Acta Rev Cancer. 2023;1878(3):188891. doi:10.1016/j.bbcan.2023.188891
94. Wu Z, Jiang D, Huang X, Cai M, Yuan K, Huang P. S100A8 as a promising biomarker and oncogenic immune protein in the tumor microenvironment: an integrative pancancer analysis. J Oncol. 2022;2022:6947652.
95. Deng S, Cheng D, Wang J, et al. MYL9 expressed in cancer-associated fibroblasts regulate the immune microenvironment of colorectal cancer and promotes tumor progression in an autocrine manner. J Exp Clin Cancer Res. 2023;42(1):294.
96. Wang JH, Zhang L, Huang ST, et al. Expression and prognostic significance of MYL9 in esophageal squamous cell carcinoma. PLoS One. 2017;12:4.
97. Lv M, Luo L, Chen X. The landscape of prognostic and immunological role of myosin light chain 9 (MYL9) in human tumors. Immun Inflamm Dis. 2022;10(2):241–254.
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.