")
Back to Journals » Drug Design, Development and Therapy » Volume 9
Bu-Shen-Ning-Xin decoction: inhibition of osteoclastogenesis by abrogation of the RANKL-induced NFATc1 and NF-κB signaling pathways via selective estrogen receptor α
Authors Wang L , Qiu X, Gui Y, Xu Y, Gober H, Li D
Received 13 May 2015
Accepted for publication 5 June 2015
Published 21 July 2015 Volume 2015:9 Pages 3755—3766
DOI https://doi.org/10.2147/DDDT.S88512
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Wei Duan
Ling Wang,1,2,* Xue-Min Qiu,1,2,* Yu-Yan Gui,1,2 Ying-Ping Xu,1,2 Hans-Jürgen Gober,3 Da-Jin Li1
1Laboratory for Reproductive Immunology, Hospital and Institute of Obstetrics and Gynecology, IBS, Fudan University Shanghai Medical College, 2Shanghai Key Laboratory of Female Reproductive Endocrine Related Diseases, Shanghai, People’s Republic of China; 3Department of Pharmacy, Wagner Jauregg Hospital and Children’s Hospital, Wagner Jauregg Weg, Linz, Austria
*These authors contributed equally to this work
Introduction: Bu-Shen-Ning-Xin decoction (BSNXD) is a traditional Chinese medicinal composition that has been used as a remedy for postmenopausal osteoporosis, but the mechanisms affecting bone metabolism are not fully understood.
Purpose: We investigated the molecular mechanism and signaling pathway underlying the effect of BSNXD on osteoclastogenesis.
Materials and methods: A postmenopausal osteoporosis animal model generated by ovariectomy was administered BSNXD and drug-derived serum was prepared. An enzyme immunoassay was conducted to measure the 17-β-estradiol (E2) concentration in the drug-derived serum. Bone marrow-derived monocyte/macrophage precursor cells were treated with drug-derived serum, and tartrate-resistance acid phosphatase staining was conducted to observe osteoclastogenesis. A bone resorption assay was performed to analyze the effect on osteoclastic resorptive function. Real-time PCR, flow cytometry, Western blotting, transfection, and luciferase assays were conducted to explore the related mechanism.
Results: E2 was not elevated in BSNXD-derived serum. BSNXD-derived serum suppressed receptor activation of nuclear factor κB ligand (RANKL)-activated osteoclastogenesis in a dose-dependent manner; this effect could be reversed by estrogen receptor α antagonist methyl-piperidino-pyrazole. The serum suppressed RANKL-induced NF-κB transcription and inhibited the accumulation of nuclear factor of activated T-cells, cytoplasmic 1 in osteoclast precursor cells; the inhibitory effect was abolished by methyl-piperidino-pyrazole but not the estrogen receptor β antagonist or androgen receptor antagonist.
Conclusion: These results collectively suggest that administration of BSNXD presents inhibitory effects on osteoclast differentiation by abrogating the RANKL-induced nuclear factor of activated T-cells, cytoplasmic 1 and NF-κB signaling pathways downstream of estrogen receptor α, thereby contributing to the inhibitory effect on bone resorption.
Keywords: herbal formula, osteoclastogenesis, estrogen receptor α, NF-κB, NFATc1
Introduction
Postmenopausal osteoporosis (PMO) is a major health concern in countries with an aging population (ie, People’s Republic of China) and occurs due to estrogen deprivation postmenopause.1 The balance between osteoblast-mediated bone formation and osteoclast-mediated bone resorption is disrupted by estrogen deficiency in favor of increased osteoclast-mediated bone resorption, thereby leading to bone loss.2
Mature osteoclasts are multinucleated giant cells that play a pivotal role as bone-resorbing cells during bone remodeling by maintaining a balance with bone-forming osteoblasts. Osteoclasts differentiate from hematopoietic stem cells following stimulation by two key factors produced by osteoblasts: macrophage-colony-stimulating factor (M-CSF) and receptor activation of nuclear factor κB ligand (RANKL).3 Several regulators of osteoclast fate determination and differentiation have been identified, and a variety of transcription factors (ie, nuclear factor of activated T-cells, calcineurin-dependent 1 or NFATc1) seem to be pivotal for osteoclast differentiation.4
Increasing estrogen levels in postmenopausal women via estrogen-replacement therapy could prevent postmenopausal bone loss. However, the safety of this practice is controversial. According to the Women’s Health Initiative Study, long-term estrogen replacement causes an unacceptable increase in the risk of various diseases, such as heart attack, stroke, and breast and uterine cancer.5 Moreover, the recently proposed “critical time window” claimed that use of estrogen after menopause would lead to increased health risks, while using the same during the early menopause transition might be beneficial.6 Consequently, other potential, effective, and safe therapies need to be examined, including traditional Chinese medicines (TCMs).
Bu-Shen-Ning-Xin decoction (BSNXD) is a type of TCM remedy that has been used for hundreds of years to treat and prevent menopause-related disorders and aging diseases,7 including PMO. BSNXD consists of drying rehmannia root, common anemarrhena rhizome, bark of the Chinese corktree, barbary wolfberry fruit, Chinese dodder seed, shorthorned epimedium herb, spina date seed, and oriental waterplantain rhizome (Table 1). Although BSNXD has a beneficial action on skeletal mass,8 the cellular and molecular mechanism mediating this action is not fully understood.
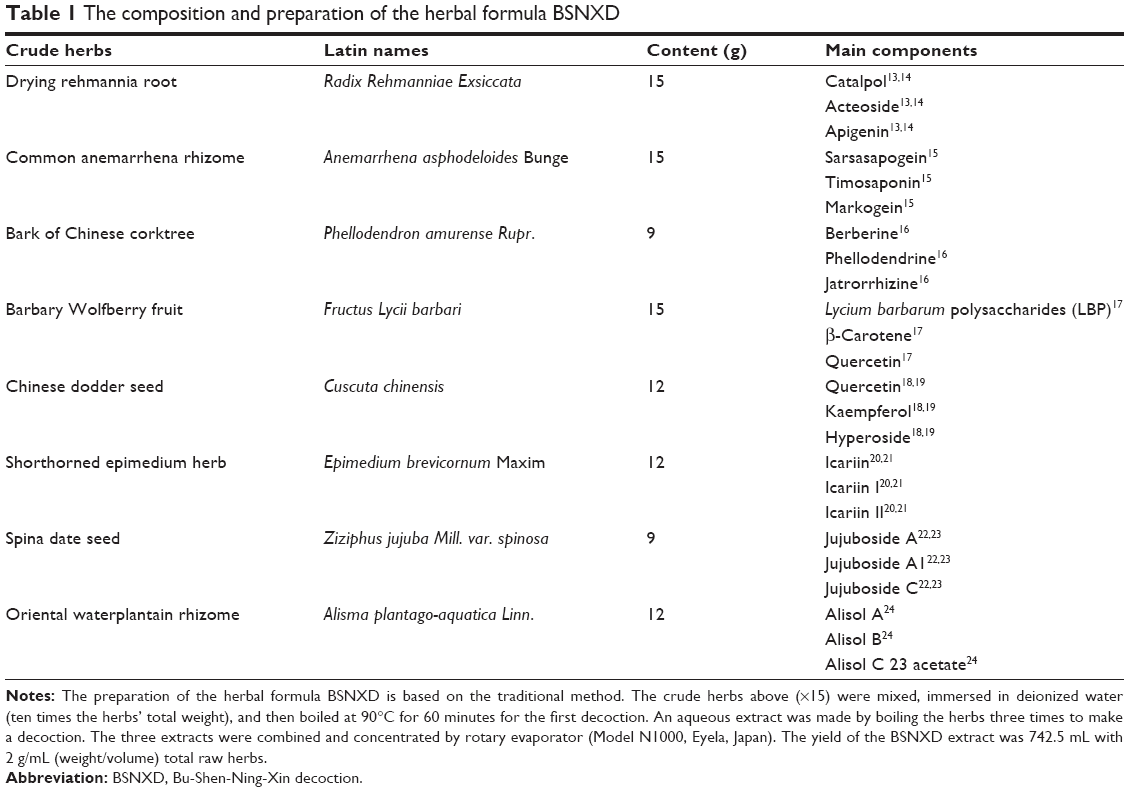 | Table 1 The composition and preparation of the herbal formula BSNXD |
Unlike modern medicine that comprises target-oriented therapeutic interventions, TCMs aim to restore the Yin-Yang balance of body energy using a holistic and synergistic approach so that the body’s normal function and homeostasis can be restored.9 The hypothesis underlying TCMs is that multiple active phytochemical components in the formulae may simultaneously target multiple molecules/pathways and thereby potentially achieve superior effects compared with single compounds alone.10 However, the specific mechanisms are always vague.
To investigate the effects of BSNXD on PMO, we used a female mouse ovariectomy (OVX) model to deplete ovarian hormones.11,12 Previously, we prepared pharmacological serum from ovariectomized mice treated with BSNXD and found that BSNXD pharmacological serum promoted proliferation and inhibited apoptosis of mouse osteoblasts through the activation of the MAPK signaling pathways via ERK1/2 phosphorylation.8 The present study investigated the effects of mid-dose BSNXD-derived serum on murine osteoclastogenesis in vitro from osteoclast precursor cells stimulated with RANKL and M-CSF and explored the involvement of RANKL-induced NFATc1 expression regulation on the effect of BSNXD on osteoclastogenesis.
Materials and methods
Preparation of BSNXD extracts
The herbal formula of BSNXD is composed of eight crude herbs that are prepared as shown in Table 1.13–24 The rule of compositions comes from traditional Chinese medicinal theory, and the compositions are derived from our clinical experience. The BSNXD was obtained from the pharmacy of the Hospital of Obstetrics and Gynecology, Fudan University, Shanghai, People’s Republic of China. These products were manufactured under good manufacturing practice conditions at the Institute of Obstetrics and Gynecology, Fudan University Shanghai Medical College, according to the protocol described in Chinese Pharmacopoeia 200525 with modifications. Crude water extracts were prepared from powdered BSNXD.7,8
Experimental drug administration
The animal experiments were approved by the experimental animal ethics committee of Fudan University. We used 90 eight-week-old female BALB/c mice with a body mass between 20 g and 30 g purchased from the Laboratory Animal Facility of Chinese Academy of Sciences (Shanghai, People’s Republic of China). All of the mice underwent bilateral oophorectomy. After OVX, the mice were randomly divided into three groups (OVX, OVX + BSNXD mid-dose, and OVX + E2 groups; n=30 mice per group).
The OVX control group received a saline treatment (n=30). The OVX + BSNXD mid-dose group mice were treated with 0.5 mL of evaporated BSNXD extract twice daily (with 1 g/mL, w/v, of the total raw herbs) by oral administration (Table 1); this dosage was ninefold higher than the human adult dose based on an established formula for human–mouse drug conversions (n=30). The OVX + E2 group mice received estrogen (17-β-estradiol, E2) treatment (100 μg/kg per day orally, n=30).26–29 After 4 weeks, all mice were weighed and sacrificed after the last treatment to harvest blood samples. Successful OVX was confirmed by observation of a lack of ovarian tissue and atrophied uterine horns.
Drug-derived serum preparation and E2 measurement by enzyme immunoassay
All of the mice were sacrificed after 1 month of treatment, and blood samples were quickly obtained from the mice by cardiac puncture. Blood volumes up to 1 mL have frequently been obtained from mice. Serum samples were prepared by centrifugation. The sera were inactivated at 56°C for 30 minutes, filtered with a 0.2 μm filter, and used at different concentrations. The sera were stored at -20°C for the determination of E2 concentrations or the following experiments. The E2 concentrations in serum samples were measured using the E2 enzyme immunoassay kit following the manufacturer’s protocol (BioCheck Inc., Burlingame, CA, USA).
Bone marrow isolation and primary osteoclast culture
Mice in the OVX group were sacrificed by CO2 asphyxiation. For each mouse, the right femurs and tibias were isolated and cleaned by removing any associated muscles or connective tissues with sterile gauze. Bone marrow was flushed from the right femur and tibia using a 23 G needle with basal culture medium (Modified Eagle Medium [MEM] without phenol red supplemented with 50 U/mL penicillin and 50 μg/mL streptomycin; Thermo Fisher Scientific, Waltham, MA, USA). Total bone marrow from the right femur and tibia of each mouse was pooled and passed through a 40 μm nylon mesh filter to remove any bone particulates. Bone marrow cells were cultured with 10 ng/mL M-CSF (R&D Systems, Minneapolis, MN, USA) for 2 days and used as bone marrow-derived monocyte/macrophage precursor cells (BMMs). BMMs were cultured with 50 ng/mL RANKL (Peprotech, Rocky Hill, NJ, USA) and 10 ng/mL M-CSF for 3 days.30,31 The OVX + BSNXD mid-dose group-derived sera were added to the culture medium (without phenol red) at 10%, 15%, and 20% concentrations. To estimate the effect of OVX + BSNXD group-derived serum on osteoclastogenesis, OVX group-derived BMMs were exposed to serial concentrations of OVX + BSNXD mid-dose group-derived serum or OVX + E2 group-derived serum in the presence of RANKL stimulation for 72 hours.
OVX group-derived BMMs were treated with 10−6 M methyl-piperidino-pyrazole (MPP, estrogen receptor [ER]α antagonist; Tocris Cookson Inc., Ellisville, MO, USA), 10 nM R,R-THC (ERβ antagonist; Tocris Cookson Inc.), or 10 μM flutamide (FLUT, androgen receptor [AR] antagonist; Sigma-Aldrich Co., St Louis, MO, USA) in MEM without phenol red for 1 hour prior to exposure to 20% OVX group-derived serum (control serum) or 20% OVX + BSNXD mid-dose group-derived serum for 72 hours.27,32,33 Osteoclastogenesis was evaluated by tartrate-resistance acid phosphatase (TRAP) staining using the Leukocyte Acid Phosphatase Kit (Sigma-Aldrich Co.). TRAP-positive multinucleated cells (TRAP+ MNCs; more than five nuclei) were counted microscopically. The results from at least six independent experiments are shown.
Bone resorption assay
BMMs were pretreated with the control or 20% mid-dose BSNXD-derived serum in the presence of RANKL for 48–72 hours to induce osteoclastogenesis on collagen-coated dishes. Then, the mature osteoclasts in each group were harvested by treatment with trypsin-EDTA (Thermo Fisher Scientific, Waltham, MA, USA). The collected cells were seeded and cultured for 2 days on dentin slices treated with 1 M NH4OH with sonication and stained with 0.5% toluidine blue.34,35
Quantitative real-time PCR
Total RNA was extracted from the control and the various time points of mid-dose BSNXD drug serum-exposed BMMs according to the manufacturer’s instructions (SV Total RNA Isolation System; Promega Corporation, Fitchburg, WI, USA). Quantitative real-time polymerase chain reaction (PCR) amplifications were performed with gene-specific primers using a 7500 real-time PCR System (Thermo Fisher Scientific). The primers for Csf1R, RANK, and β-actin36,37 are listed in Table 2. The relative standard curve assay was used to calculate the amplification differences for each primer set. Relative Csf1R or RANK expression levels were obtained from relative standard curves run in triplicate after dividing each value by the β-actin value.
 | Table 2 Sequences of the primers for Csf1R, RANK, and β-actin |
Flow cytometry
Staining for BMM cell surface expression of Csf1R and RANK was performed using monoclonal phycoerythrin-conjugated anti-mouse CD115 (CSF1R), phycoerythrin anti-mouse RANK (CD265), and isotype controls (eBioscience Inc., San Diego, CA, USA) on a FACScan flow cytometer with CellQuest software (BD Biosciences, San Jose, CA, USA).
Transfection and luciferase assay
Transient transfections with NF-κB-luciferase were accomplished using Lipofectamine (Gibco-BRL). The luciferase transfection protocol and luciferase activity assay using the dual luciferase kit (Promega Corporation) and TD-20/20 luminometer (TurnerDesigns, Sunnyvale, CA, USA) have been extensively described.38–40 Briefly, OVX group-derived BMMs were transfected with 2 μg of NF-κB-luciferase. A significant RANKL-induced increase in NF-κB-luciferase activity was first detected at 6 hours; similar results were obtained at both the 6 hours and 24 hours time points.40 To determine the effects of OVX + BSNXD mid-dose group-derived serum on RANKL-induced NF-κB-luciferase activity, the cells were treated with the indicated agents for 24 hours after transfection. Concentrations of 10−6 M MPP, 10 nM R,R-THC, or 10 μM FLUT in MEM without phenol red were administered for 1 hour prior to stimulation with 20% OVX group-derived serum (control serum) or 20% OVX + BSNXD mid-dose group-derived serum in the presence of 50 ng/mL RANKL. Cells were harvested 12 hours after treatment; then, cell extracts were prepared and luciferase activities were measured.
Western blot analysis
BMMs were harvested after stimulation with RANKL and M-CSF. The cultures were washed with PBS and lysed with RIPA buffer (20 mM Tris–HCl, pH 7.5, 150 mM NaCl, 1 mM EDTA, 50 mM β-glycerophosphate, 1% NP-40, 1 mM Na3VO4, and 1× protease inhibitor cocktail). Total protein content was measured using the Quant-iT™ protein assay kit (Thermo Fisher Scientific; Q32860). The extracted proteins were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis and electrotransferred onto PVDF membranes. The membranes were blocked for 1 hour at room temperature in 5% fat-free milk in Tris-buffered saline and Tween 20 (TBST) buffer (10 mM Tris–HCl, pH 7.5, 150 mM NaCl, and 0.1% Tween 20) with gentle agitation, washed once, and then incubated overnight at 4°C with primary antibodies against phospho-PLCγ2, PLCγ2 (Cell Signaling Technology), NFATc1 (Santa Cruz Biotechnology Inc., Dallas, TX, USA), and β-actin (Santa Cruz Biotechnology Inc.). The blots were washed twice with TBST and incubated for 1 hour at room temperature with horseradish peroxidase-conjugated secondary antibodies (Bio-Rad Laboratories Inc., Hercules, CA, USA; anti-mouse, 170-5047), followed by another round of triple washing. Immunocomplexes were observed by a chemiluminescence reaction with enhanced chemiluminescence reagents (Thermo Fisher Scientific; 34080) and photographed. Then, the membranes were stripped by incubation with stripping solution (Pierce; Thermo Fisher Scientific) for 15–20 minutes at room temperature and reprobed to detect other proteins. The actin antibody was used as the loading control.
Statistical analysis
All values were expressed as the mean ± SEM. Differences between experimental groups were analyzed using ANOVA and the Kruskal–Wallis tests, with P<0.05 considered significant. Data were analyzed using the SPSS software.
Results
The effect of mid-dose BSNXD-derived serum on osteoclastogenesis in vitro
We added the BSNXD drug directly to the culture of murine BMMs in our in vitro experiments but observed no effect on RANKL-induced osteoclastogenesis (data not shown). Therefore, we analyzed the modulation of RANKL-induced osteoclastogenesis by murine mid-dose BSNXD-derived serum. Murine BMMs were incubated with serial concentration of mid-dose BSNXD-derived serum or E2-derived serum in the presence of RANKL and M-CSF and allowed to grow and differentiate into osteoclasts. Our results showed that the mid-dose BSNXD-derived serum decreased the numbers of TRAP-positive multinucleated osteoclasts in a dose-dependent manner (P<0.05 or P<0.01), with maximal effects observed in the 20% mid-dose BSNXD-derived serum; notably, the 20% mid-dose BSNXD-derived serum showed decrease in osteoclastogenesis compared with the 20% E2-derived serum (Figure 1A and B). Neither the 10%–20% mid-dose BSNXD-derived serum nor the E2-derived serum affected the cell viability of BMCs, as determined by the MTT assay (data not shown).
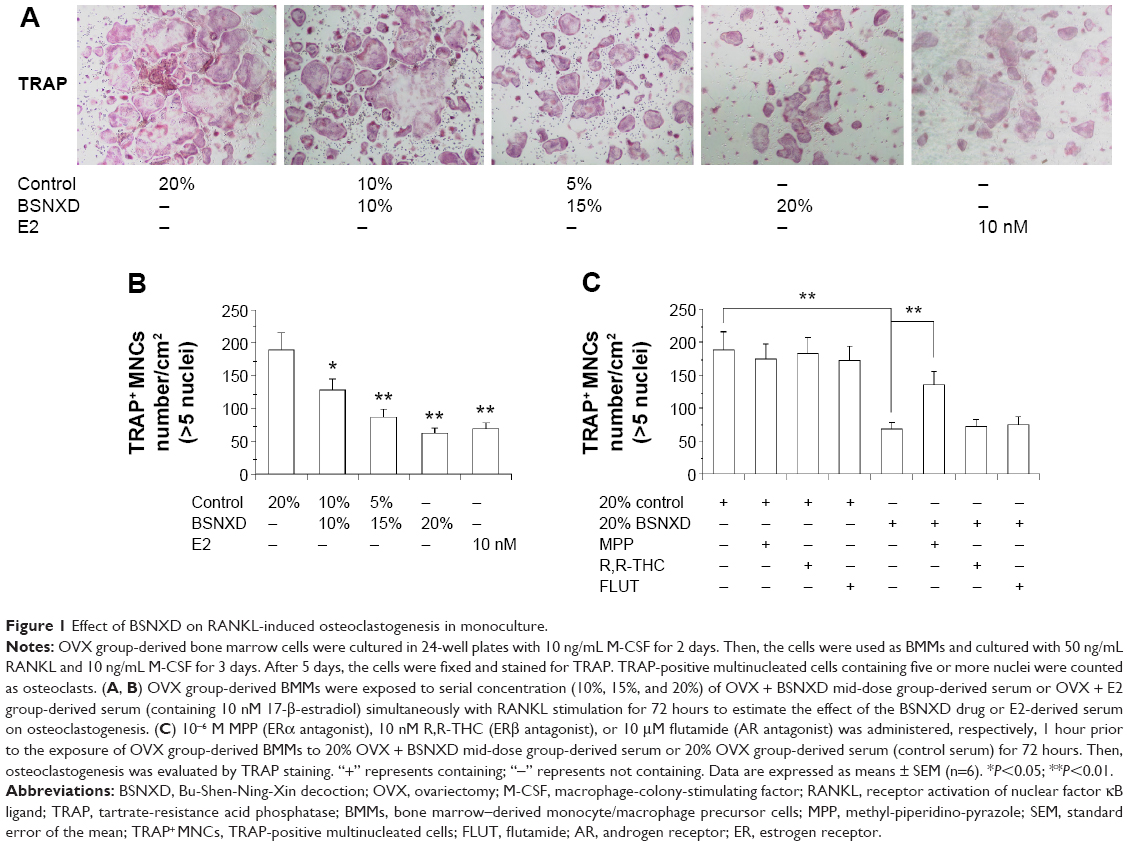 | Figure 1 Effect of BSNXD on RANKL-induced osteoclastogenesis in monoculture. |
Accordingly, we used the 20% mid-dose BSNXD-derived serum to explore the downstream signaling pathway and molecular mechanisms. To determine whether the ER subtype or AR was involved in the effects of the BSNXD-derived serum, selective ERα, ERβ, and AR antagonists were used in an additional set of experiments.
Pretreatment of BMMs with the selective ERα antagonist (10−6 M MPP), ERβ antagonist (10 nM R,R-THC), or AR antagonist FLUT (10 μM FLUT) for 1 hour prior to treatment with 20% OVX-derived serum (control serum) for 72 hours did not affect osteoclastogenesis. In contrast, pretreatment of BMMs with 10−6 M MPP for 1 hour prior to treatment with 20% mid-dose BSNXD-derived serum for 72 hours significantly increased osteoclastogenesis (P<0.01); however, treatment with 10 nM R,R-THC or 10 μM FLUT did not have an effect (Figure 1C). This result suggested that the mid-dose BSNXD-derived serum inhibited osteoclastogenesis via the ERα pathway rather than the ERβ or AR pathways.
BSNXD did not elevate estradiol levels in pharmacological serum
The serum estradiol concentration of OVX mice was significantly lower than the Sham mice (P<0.05, data not shown). Administration of BSNXD to the OVX mice did not significantly change the estradiol concentration in the serum (P>0.05, data not shown).
BSNXD-derived serum suppresses RANKL-induced NF-κB transcription via ERα in osteoclasts
To determine the intracellular signal pathway and transcription factor involved in the action of BSNXD-derived serum, BMMs transfected with NF-κB-luciferase were pretreated with 10−6 M MPP, 10 nM R,R-THC, or 10 μM FLUT 1 hour prior to the treatment with 20% OVX-derived serum (control serum) or 20% OVX + BSNXD mid-dose group-derived serum in the presence of RANKL. The NF-κB-luciferase activity decreased significantly after treatment with 20% BSNXD-derived serum compared with the 20% control serum (P<0.01); the effect of the BSNXD-derived serum was significantly blocked by the ERα antagonist (MPP) (P<0.01) but not by the ERβ antagonist (R,R-THC) or AR antagonist FLUT. Pretreatment with MPP, R,R-THC, or FLUT prior to treatment with the 20% OVX-derived serum did not affect NF-κB-luciferase activity (Figure 2A). This result suggests that ERα is essential for the suppression of the effect of BSNXD-derived serum on RANKL-induced NF-κB transcription in osteoclasts.
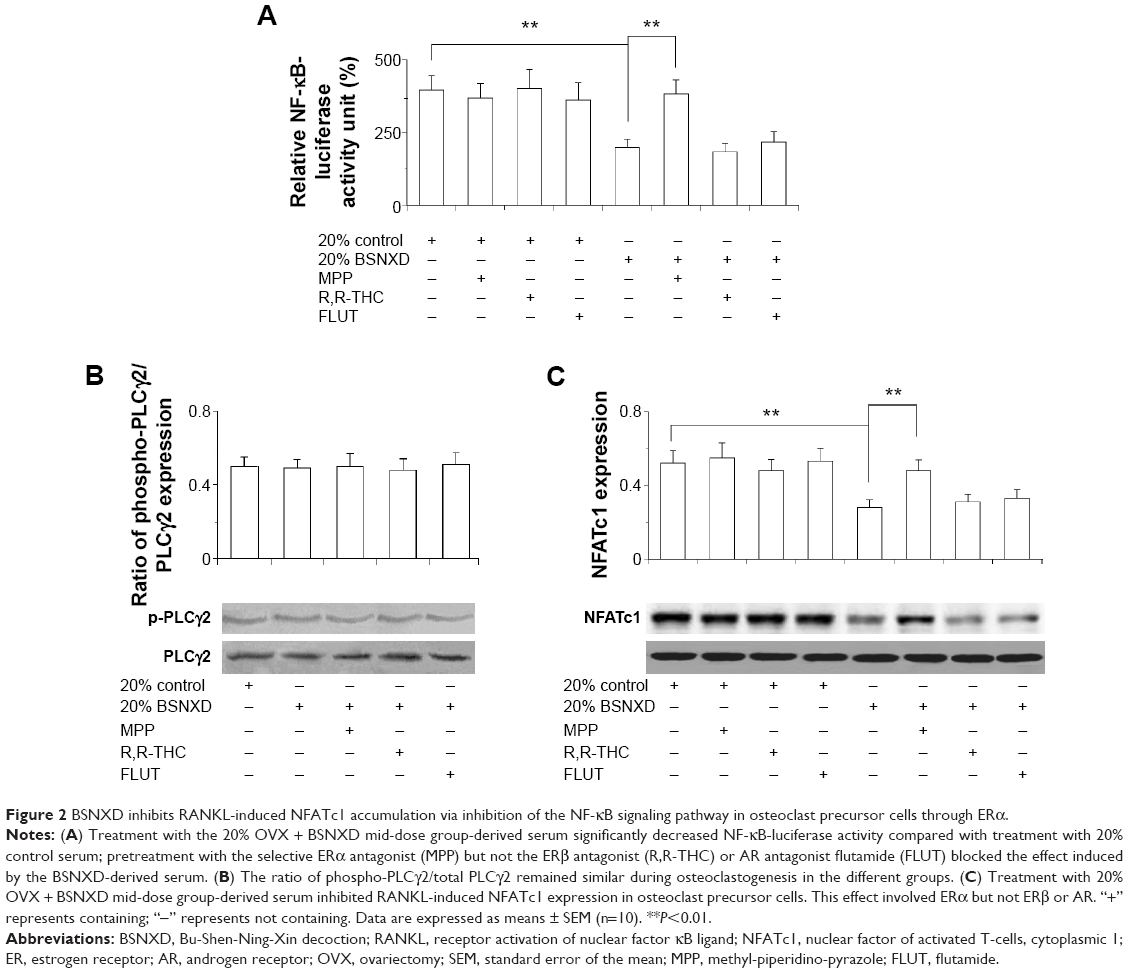 | Figure 2 BSNXD inhibits RANKL-induced NFATc1 accumulation via inhibition of the NF-κB signaling pathway in osteoclast precursor cells through ERα. |
BSNXD-derived serum inhibits RANKL-induced NFATc1 accumulation via the ERα-mediated pathway in osteoclast precursor cells
We investigated the effect of BSNXD-derived serum on RANKL-induced NFATc1 expression and PLC-γ2 phosphorylation during osteoclastogenesis. Exposure to 20% mid-dose BSNXD-derived serum with RANKL treatment did not affect PLC-γ2 phosphorylation during osteoclastogenesis (Figure 2B). Pretreatment with MPP, R,R-THC, or FLUT prior to treatment with the 20% OVX-derived serum did not affect RANKL-induced NFATc1 expression in BMMs. Interestingly, the 20% mid-dose BSNXD-derived serum significantly inhibited RANKL-induced NFATc1 expression in BMMs compared with the control serum (P<0.01). This effect could be abolished by the ERα antagonist (MPP), suggesting that BSNXD downregulated the expression of NFATc1 via ERα signaling pathway (P<0.01). In contrast, the ERβ antagonist (R,R-THC) and AR antagonist FLUT had no significant effect on NFATc1 expression (Figure 2C). These results suggest that this effect involves ERα rather than ERβ or AR.
BSNXD-derived serum does not affect Csf1R and RANK expression during osteoclastogenesis
To investigate whether the BSNXD-induced decrease in osteoclast numbers is due to the downregulation of CSF1R (M-CSF Receptor/CD115) or RANK (RANKL receptor/CD265) expression in osteoclast precursors (OCPs), we examined CSF1R and RANK mRNA and protein levels in OCPs by real-time PCR and flow cytometry, respectively. Bone marrow cells were treated with 20% mid-dose BSNXD-derived serum or control serum, and CSF1R and RANK mRNA and protein expression levels were compared between pre- and post-treatment in the presence of RANKL. There were no significant changes in the CSF1R mRNA and protein levels in bone marrow cells after 48 hours of treatment compared with the control (Figure 3A); there were also no differences in the RANK mRNA or protein levels (Figure 3B). These results suggest that the BSNXD-induced osteoclastogenesis inhibition occurs independently of Csf1R or RANK in OCPs.
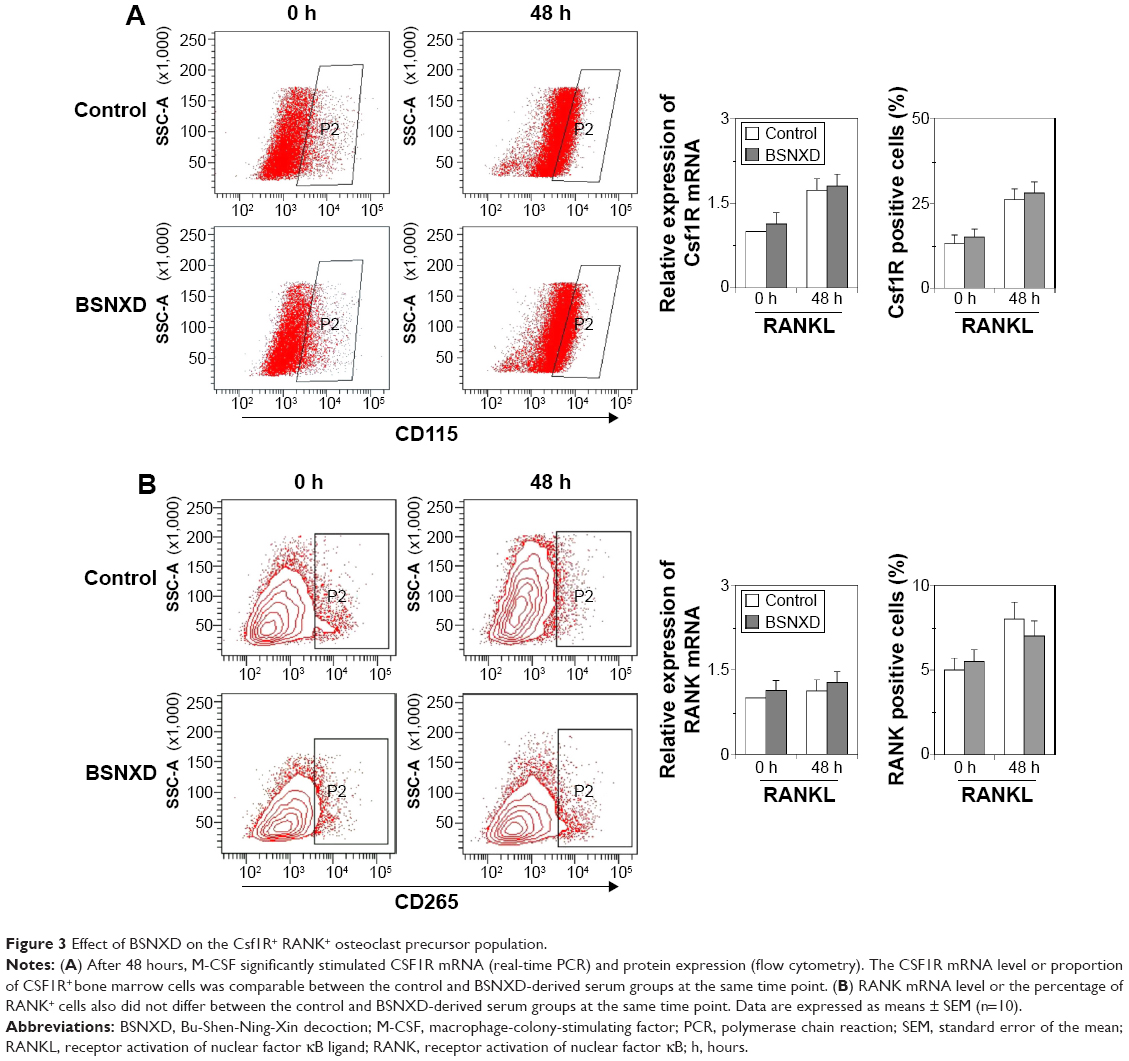 | Figure 3 Effect of BSNXD on the Csf1R+ RANK+ osteoclast precursor population. |
BSNXD-derived serum has no effect on the osteoclast bone-resorptive function
Next, we examined whether the BSNXD-derived serum influenced osteoclast function in vitro. We pretreated BMMs with the control or 20% mid-dose BSNXD-derived serum in the presence of RANKL to induce osteoclastogenesis in vitro. Then, the same number of mature osteoclasts were harvested from each group and seeded onto dentin slices. Our results showed that the BSNXD serum did not affect the bone-resorbing activity (ie, the resorption area and osteoclast pits) compared with the control (Figure 4A and B). Thus, BSNXD did not present an effect on the osteoclast bone-resorptive function.
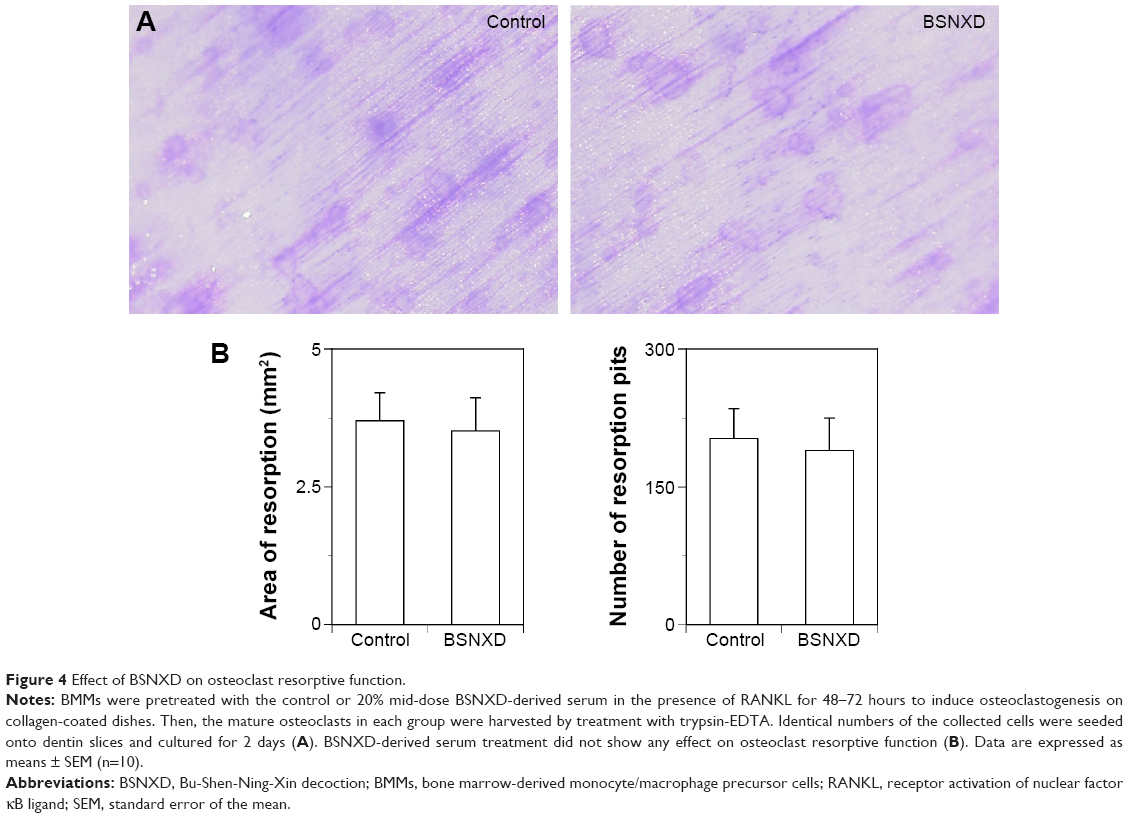 | Figure 4 Effect of BSNXD on osteoclast resorptive function. |
Discussion
Owing to the arrest of ovarian function after OVX, there is a sudden and dramatic decrease in estrogen production that leads to accelerated bone resorption by increasing osteoclast formation and lifespans. Osteoclast differentiation is primarily regulated by two signaling pathway: RANKL and M-CSF.41–43 M-CSF promotes the proliferation and survival of BMMs.44 RANKL is an osteoblast-derived factor that is required for osteoclasts formation; RANKL activates the differentiation process by inducing NFATc1, the master transcription factor for osteoclastogenesis.4
After oral administration of BSNXD, the compounds are absorbed into the bloodstream and become active compounds. The compounds may become functional by oxidation, hydrolysis, or reduction; primary metabolites may also undergo conjugation reactions to form secondary metabolites.45,46 The serum effect on osteoclasts was dose-dependent and suppressed osteoclast differentiation with no effect on osteoclast viability. Our data also showed that the estrogen level in the BSNXD-derived serum was comparable with the control serum. However, the effect of the BSNXD-derived serum was comparable with the E2-derived serum, suggesting that the suppression of osteoclastogenesis by the BSNXD-derived serum was not due to toxic effects on osteoclasts or the elevation of serum E2 levels.
Analysis of the bone phenotype in ERα−/−, ERβ−/−, and ERαβ−/− mice revealed that both receptor subtypes participated in the control of bone turnover in females.47 In male mice, estradiol regulates trabecular bone remodeling exclusively through ERα, while in females, both ER subtypes influence bone turnover and trabecular structure. OVX of ERβ−/− and wild-type mice led to the loss of trabecular bone mineral density, implying that ERα played an important role in maintaining bone integrity and homeostasis.48 Therefore, ERα is the main mediator of the protective effects of estradiol in bone. Indeed, ERα−/− mice have a low trabecular bone mass.49 Estradiol prevented ovariectomy-induced bone loss in ERβ−/− mice to levels similar to those observed in wild-type males and females, indicating that ERα was the major mediator of estradiol effects in bone. Furthermore, the evidence that ERα activation due to a rare genetic mutation leads to severe osteoporosis in humans has confirmed that ERα plays an important role in sustaining bone homeostasis.50,51
MPP is a nonsteroidal, pyrazole compound that contains a basic side-chain addition that is reported to convert the pyrazole from an ERα agonist to an ERα antagonist.52 Co-transfection and in vitro binding assays demonstrated that MPP displayed a 200-fold preferential binding affinity for ERα compared with ERβ.53 Moreover, an additional in vitro study showed that MPP functioned as an ERα-selective antagonist for estrogen-regulated genes.54 R,R-THC is an agonist for ERα but a full antagonist for ERβ. The relative potency of R,R-THC in ERα and ERβ transcription assays correlates rather well with its relative affinity in ligand binding assays: it has a higher affinity for ERβ, which is consistent with its greater potency as an ERβ antagonist than an ERα agonist. An ERβ-selective antagonist (R,R-THC) could be used to identify ERβ-mediated activity in animals based on their selective suppression by this ligand.55 Flutamide, a silent antagonist of AR, competes with testosterone and its powerful metabolite dihydrotestosterone for binding to ARs and is primarily used to treat prostate cancer.56 FLUT could be used to identify AR-mediated activity in cells based on its suppression by this ligand.13
Our data showed that the suppressive effect of BSNXD-derived serum on RANKL-induced osteoclastogenesis occurred via the ERα but not the ERβ or AR pathways. In the present study, we found no differences in bone-resorbing activity of mature osteoclasts after treatment with BSNXD-derived serum or control serum. Therefore, BSNXD does not have an effect on the resorptive function of mature osteoclasts.
CSF1R (the receptor for M-CSF) is an essential survival factor for OCPs. M-CSF/CSF1R signaling drives the differentiation of monocytes into macrophages or osteoclasts. CSF1R also acts as a potent stimulator of RANK expression.57 Because CSF1R and RANK are both expressed on OCPs, one may expect that BSNXD inhibited osteoclastogenesis via downregulating RANK and CSF1R expression in the OCPs, and thereby decreasing the sensitivity of precursor cells to RANK and M-CSF. However, we did not observe any changes in the expression of RANK (RANKL receptor/CD265) and CSF1R (M-CSF Receptor/CD115) in the OCPs. Thus, the inhibition of osteoclastogenesis by BSNXD did not result from the decrease of the Csf1R+ RANK + OCP. Osteoclast formation and functions are mediated by RANKL-induced NF-κB activation.58 NF-kB is considered to be one of the essential transcription factors for osteoclastogenesis because inhibition of this transcription factor or other proteins in the NF-kB signaling pathway results in the absence or significant reduction of osteoclast formation.59 Therefore, the inhibition of RANKL-induced NF-κB activity might be an effective approach to target osteoclastogenesis and treat osteoporosis. The results from the present study revealed that BSNXD-derived serum inhibited RANKL-induced NF-κB activity during osteoclastogenesis in an ERα-dependent manner.
Binding of RANKL–RANK also activates transcription factor NFATc1, which is a master regulator of osteoclastogenesis,60 and then directs the expression of osteoclast-specific genes. Our data revealed that the suppression of RANKL-induced activation of NFATc1 via the ERα pathway by BSNXD was correlated with its inhibition of osteoclastogenesis. In our study, the attenuation of the NF-κB signaling pathway downstream of ERα may correspond to a mechanism that contributes to the suppression of NFATc1 expression by BSNXD. Thus, the mechanism that mediates the anti-osteoclastogenic roles of BSNXD may be the abrogation of RANKL-induced NFATc1 expression via inhibition of the NF-κB signaling pathway downstream of ERα (Figure 5).
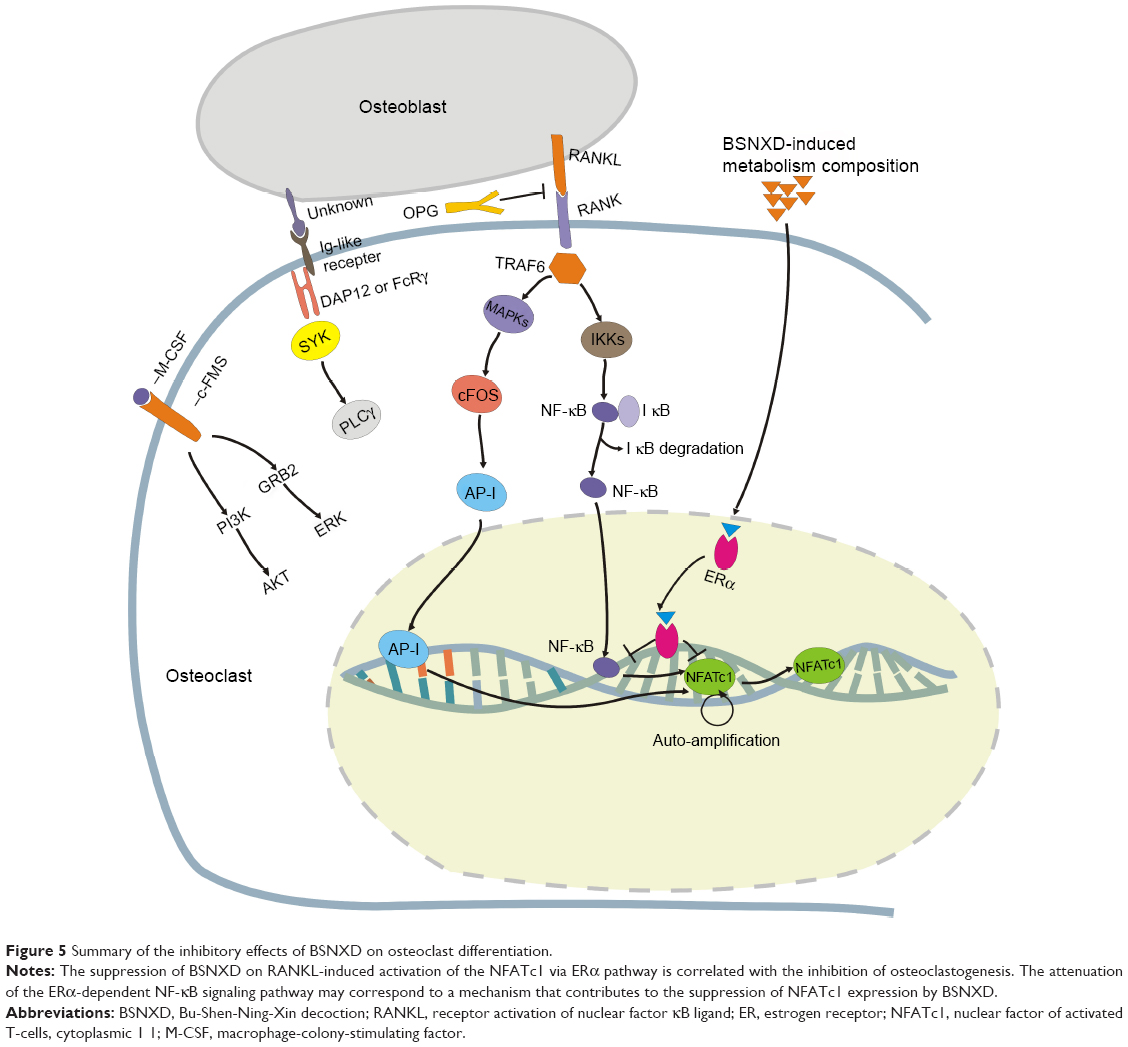 | Figure 5 Summary of the inhibitory effects of BSNXD on osteoclast differentiation. |
Conclusion
In conclusion, this study revealed that the inhibitory effects of BSNXD on osteoclast differentiation were related to the abrogation of RANKL-induced NFATc1 expression via inhibition of the ERα-dependent NF-κB signaling pathway in a holistic manner. This finding suggests that this TCM remedy is a good approach to treat PMO.
Acknowledgments
This work was supported by the Science and Technology Commission of Shanghai Municipality 2015 YIXUEYINGDAO project, No 15401932200 (to L Wang), the FY2008 JSPS Postdoctoral Fellowship Researchers P08471 (to L Wang), National Natural Science Foundation of China No 30801502 (to L Wang), Shanghai Pujiang Program No 11PJ1401900 (to L Wang), National Natural Science Foundation of China No 81401171 (to X-M Qiu) and Program for Outstanding Medical Academic Leader (to D-J Li).
Disclosure
The authors report no conflicts of interest in this work.
References
Watts NB; GLOW Investigators. Insights from the Global Longitudinal Study of Osteoporosis in Women (GLOW). Nat Rev Endocrinol. 2014;10(7):412–422. | ||
Cauley JA. Estrogen and bone health in men and women. Steroids. 2015;99(pt A):11–15. | ||
Novack DV, Teitelbaum SL. The osteoclast: friend or foe? Annu Rev Pathol. 2008;3:457–484. | ||
Takayanagi H, Kim S, Koga T, et al. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev Cell. 2002;3(6):889–901. | ||
Rossouw JE, Anderson GL, Prentice RL, et al; Writing Group for the Women’s Health Initiative Investigators. Risks and benefits of estrogen plus progestin in healthy postmenopausal women: principal results from the Women’s Health Initiative randomized controlled trial. JAMA. 2002;288(3):321–333. | ||
Pinkerton JV, Stovall DW. Reproductive aging, menopause, and health outcomes. Ann N Y Acad Sci. 2010;1204:169–178. | ||
Wang L, Qiu XM, Hao Q, Li DJ. Anti-inflammatory effects of a Chinese herbal medicine in atherosclerosis via estrogen receptor beta mediating nitric oxide production and NF-kappaB suppression in endothelial cells. Cell Death Dis. 2013;4:e551. | ||
Wang Y, Cui K, Zhao H, Li D, Wang W, Zhu Y. Bushen Ningxin decoction pharmacological serum promotes the proliferation and suppresses the apoptosis of murine osteoblasts through MAPK pathway. J Ethnopharmacol. 2009;122(2):221–226. | ||
Efferth T, Li PC, Konkimalla VS, Kaina B. From traditional Chinese medicine to rational cancer therapy. Trends Mol Med. 2007;13(8):353–361. | ||
Chow MS, Huang Y. Utilizing Chinese medicines to improve cancer therapy – fiction or reality? Curr Drug Discov Technol. 2010;7(1):1. | ||
Zhu S, Chen K, Lan Y, Zhang N, Jiang R, Hu J. Alendronate protects against articular cartilage erosion by inhibiting subchondral bone loss in ovariectomized rats. Bone. 2013;53(2):340–349. | ||
Hsiao HB, Lin H, Wu JB, Lin WC. Kinsenoside prevents ovariectomy-induced bone loss and suppresses osteoclastogenesis by regulating classical NF-kappaB pathways. Osteoporos Int. 2013;24(5):1663–1676. | ||
Liu YF, Liang D, Luo H, et al. Hepatoprotective iridoid glycosides from the roots of Rehmannia glutinosa. J Nat Prod. 2012;75(9): 1625–1631. | ||
Liu YF, Liang D, Luo H, et al. Ionone glycosides from the roots of Rehmannia glutinosa. J Asian Nat Prod Res. 2014;16(1):11–19. | ||
Ji X, Feng YF. Advances in studies on saponins in Anemarrhena asphodeloides. Zhong Cao Yao. 2010;4:12–15. | ||
Hu YM, Su GH, Sze SC, Ye W, Tong Y. Quality assessment of Cortex Phellodendri by high-performance liquid chromatography coupled with electrospray ionization mass spectrometry. Biomed Chromatogr. 2010;24(4):438–453. | ||
Rukeya J, Sun YJ, Zhong LZ, Shen Y, Ye XQ. A review of phytochemical composition and bio-active of Lycium Barbarum Fruit (Goji). J Chin Inst Food Sci Technol. 2013;13(8):161–172. | ||
Li JP, Wang J, Zhang YW, Hu SP, Wang LL. Advancement on the research of China dodder. China Med Herald. 2009;6(23):5–6. | ||
Cai XG, Zhao SX. Effects of flavonoids from Cuscuta chinensis on skeleton of ovariectomized female rats. Pharmacol Clin Chin Mater Med. 2007;23(6):27–29. | ||
Yuan H, Cao SP, Chen SY, Guo LN, Zheng J, Lin RC. Research progress on chemical constituents and quality control of Epimedii Folium. Zhong Cao Yao. 2014;45(24):3630–3640. | ||
Islam MN, Kim U, Kim DH, Dong MS, Yoo HH. High-performance liquid chromatography-based multivariate analysis to predict the estrogenic activity of an Epimedium koreanum extract. Biosci Biotechnol Biochem. 2012;76(5):923–927. | ||
Zhang JW, Zhao Q. Research progress of biology characteristics and chemical constituents of semen ziziphi spinosae. China J Chin Med. 2013;28(4):550–552. | ||
Gao QH, Wu CS, Wang M. The jujube (Ziziphus jujuba Mill.) fruit: a review of current knowledge of fruit composition and health benefits. J Agric Food Chem. 2013;61(14):3351–3363. | ||
Zhu YL, Peng GP. Progress in the study on chemical constituents of Alisma orientalis. Nat Prod Res Dev. 2006;18:348–351. | ||
China Tapco PR, editor. Pharmacopoeia of the People’s Republic of China. Beijing: Chemical Industry Press; 2005. | ||
Wang L, Wang YD, Wang WJ, Li DJ. Differential regulation of dehydroepiandrosterone and estrogen on bone and uterus in ovariectomized mice. Osteoporos Int. 2009;20(1):79–92. | ||
Wang L, Wang YD, Wang WJ, Zhu Y, Li DJ. Dehydroepiandrosterone improves murine osteoblast growth and bone tissue morphometry via mitogen-activated protein kinase signaling pathway independent of either androgen receptor or estrogen receptor. J Mol Endocrinol. 2007; 38(4):467–479. | ||
Kohama SG, Anderson CP, Osterburg HH, May PC, Finch CE. Oral administration of estradiol to young C57BL/6J mice induces age-like neuroendocrine dysfunctions in the regulation of estrous cycles. Biol Reprod. 1989;41(2):227–232. | ||
Tyagi AM, Srivastava K, Kureel J, et al. Premature T cell senescence in Ovx mice is inhibited by repletion of estrogen and medicarpin: a possible mechanism for alleviating bone loss. Osteoporos Int. 2012;23(3): 1151–1161. | ||
Shinohara M, Nakamura M, Masuda H, et al. Class IA phosphatidylinositol 3-kinase regulates osteoclastic bone resorption through protein kinase B-mediated vesicle transport. J Bone Miner Res. 2012; 27(12):2464–2475. | ||
Hayashi M, Nakashima T, Taniguchi M, Kodama T, Kumanogoh A, Takayanagi H. Osteoprotection by semaphorin 3A. Nature. 2012;485(7396):69–74. | ||
Chen YJ, Lee MT, Yao HC, Hsiao PW, Ke FC, Hwang JJ. Crucial role of estrogen receptor-alpha interaction with transcription coregulators in follicle-stimulating hormone and transforming growth factor beta1 up-regulation of steroidogenesis in rat ovarian granulosa cells. Endocrinology. 2008;149(9):4658–4668. | ||
Somponpun S, Sladek CD. Role of estrogen receptor-beta in regulation of vasopressin and oxytocin release in vitro. Endocrinology. 2002;143(8):2899–2904. | ||
Kawahara I, Koide M, Tadokoro O, et al. The relationship between calcium accumulation in osteoclast mitochondrial granules and bone resorption. Bone. 2009;45(5):980–986. | ||
Keller J, Brink S, Busse B, et al. Divergent resorbability and effects on osteoclast formation of commonly used bone substitutes in a human in vitro-assay. PLoS One. 2012;7(10):e46757. | ||
Yao Z, Li P, Zhang Q, et al. Tumor necrosis factor-alpha increases circulating osteoclast precursor numbers by promoting their proliferation and differentiation in the bone marrow through up-regulation of c-Fms expression. J Biol Chem. 2006;281(17):11846–11855. | ||
Liang QQ, Li XF, Zhou Q, et al. The expression of osteoprotegerin is required for maintaining the intervertebral disc endplate of aged mice. Bone. 2011;48(6):1362–1369. | ||
Wang L, Hao Q, Wang YD, Wang WJ, Li DJ. Protective effects of dehydroepiandrosterone on atherosclerosis in ovariectomized rabbits via alleviating inflammatory injury in endothelial cells. Atherosclerosis. 2011;214(1):47–57. | ||
Tang CH, Chang CS, Tan TW, Liu SC, Liu JF. The novel isoflavone derivatives inhibit RANKL-induced osteoclast formation. Eur J Pharmacol. 2010;648(1–3):59–66. | ||
Voronov I, Li K, Tenenbaum HC, Manolson MF. Benzo[a]pyrene inhibits osteoclastogenesis by affecting RANKL-induced activation of NF-kappaB. Biochem Pharmacol. 2008;75(10):2034–2044. | ||
Yang CR, Wang JH, Hsieh SL, Wang SM, Hsu TL, Lin WW. Decoy receptor 3 (DcR3) induces osteoclast formation from monocyte/macrophage lineage precursor cells. Cell Death Differ. 2004;11(suppl 1):S97–S107. | ||
Hofbauer LC, Kuhne CA, Viereck V. The OPG/RANKL/RANK system in metabolic bone diseases. J Musculoskelet Neuronal Interact. 2004;4(3):268–275. | ||
D’Amelio P, Roato I, D’Amico L, et al. Bone and bone marrow pro-osteoclastogenic cytokines are up-regulated in osteoporosis fragility fractures. Osteoporos Int. 2011;22(11):2869–2877. | ||
Ross FP, Teitelbaum SL. alphavbeta3 and macrophage colony-stimulating factor: partners in osteoclast biology. Immunol Rev. 2005;208:88–105. | ||
Su S, Guo J, Duan JA, et al. Ultra-performance liquid chromatography-tandem mass spectrometry analysis of the bioactive components and their metabolites of Shaofu Zhuyu decoction active extract in rat plasma. J Chromatogr B Analyt Technol Biomed Life Sci. 2010;878(3–4):355–362. | ||
Dong W, Wang P, Meng X, et al. Ultra-performance liquid chromatography-high-definition mass spectrometry analysis of constituents in the root of Radix Stemonae and those absorbed in blood after oral administration of the extract of the crude drug. Phytochem Anal. 2012;23(6):657–667. | ||
Sims NA, Dupont S, Krust A, et al. Deletion of estrogen receptors reveals a regulatory role for estrogen receptors-beta in bone remodeling in females but not in males. Bone. 2002;30(1):18–25. | ||
Nilsson S, Gustafsson JA. Estrogen receptors: therapies targeted to receptor subtypes. Clin Pharmacol Ther. 2011;89(1):44–55. | ||
Windahl SH, Börjesson AE, Farman HH, et al. Estrogen receptor-alpha in osteocytes is important for trabecular bone formation in male mice. Proc Natl Acad Sci U S A. 2013;110(6):2294–2299. | ||
Grumbach MM, Auchus RJ. Estrogen: consequences and implications of human mutations in synthesis and action. J Clin Endocrinol Metab. 1999;84(12):4677–4694. | ||
Leitman DC, Paruthiyil S, Vivar OI, et al. Regulation of specific target genes and biological responses by estrogen receptor subtype agonists. Curr Opin Pharmacol. 2010;10(6):629–636. | ||
Zhou HB, Carlson KE, Stossi F, Katzenellenbogen BS, Katzenellenbogen JA. Analogs of methyl-piperidinopyrazole (MPP): antiestrogens with estrogen receptor alpha selective activity. Bioorg Med Chem Lett. 2009;19(1):108–110. | ||
Sun J, Huang YR, Harrington WR, Sheng S, Katzenellenbogen JA, Katzenellenbogen BS. Antagonists selective for estrogen receptor alpha. Endocrinology. 2002;143(3):941–947. | ||
Harrington WR, Sheng S, Barnett DH, Petz LN, Katzenellenbogen JA, Katzenellenbogen BS. Activities of estrogen receptor alpha- and beta-selective ligands at diverse estrogen responsive gene sites mediating transactivation or transrepression. Mol Cell Endocrinol. 2003;206(1–2):13–22. | ||
Sun J, Meyers MJ, Fink BE, Rajendran R, Katzenellenbogen JA, Katzenellenbogen BS. Novel ligands that function as selective estrogens or antiestrogens for estrogen receptor-alpha or estrogen receptor-beta. Endocrinology. 1999;140(2):800–804. | ||
Koch DC, Jang HS, O’Donnell EF, et al. Anti-androgen flutamide suppresses hepatocellular carcinoma cell proliferation via the aryl hydrocarbon receptor mediated induction of transforming growth factor-beta1. Oncogene. Epub 2015. | ||
Arai F, Miyamoto T, Ohneda O, et al. Commitment and differentiation of osteoclast precursor cells by the sequential expression of c-Fms and receptor activator of nuclear factor kappaB (RANK) receptors. J Exp Med. 1999;190(12):1741–1754. | ||
Soysa NS, Alles N. NF-kappaB functions in osteoclasts. Biochem Biophys Res Commun. 2009;378(1):1–5. | ||
Hayden MS, Ghosh S. Signaling to NF-kappaB. Genes Dev. 2004;18(18):2195–2224. | ||
Takayanagi H. Osteoimmunology: shared mechanisms and crosstalk between the immune and bone systems. Nat Rev Immunol. 2007;7(4):292–304. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.