")
Back to Journals » Journal of Inflammation Research » Volume 17
Is Lipopolysaccharide-Induced Lipid Metabolism Disorder in Testis of Rats a Consequence of Plasma Lipid Changes?
Authors Zheng X, Li Y , Shang X, Liu R
Received 24 September 2023
Accepted for publication 13 January 2024
Published 7 February 2024 Volume 2024:17 Pages 765—776
DOI https://doi.org/10.2147/JIR.S441840
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Xiaokang Zheng,1,2,* Yu Li,2,* Xuejun Shang,3 Ranlu Liu1
1Department of Urology, Tianjin Medical University General Hospital, Tianjin Medical University, Tianjin, 300052, People’s Republic of China; 2Department of Andrology, Henan Provincial People’s Hospital; Zhengzhou University People’s Hospital; Henan University People’s Hospital, Zhengzhou, Henan, 450003, People’s Republic of China; 3Department of Urology, Jinling Hospital, Affiliated Hospital of Medical School, Nanjing University, Nanjing, 210002, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ranlu Liu, Department of Urology, Tianjin Medical University General Hospital, Tianjin Medical University, No.154, Anshan Road, Tianjin, 300052, People’s Republic of China, Email [email protected] Yu Li, Department of Andrology, Henan Provincial People’s Hospital; Zhengzhou University People’s Hospital; Henan University People’s Hospital, No.7 Weiwu Road, Zhengzhou, Henan, 450003, People’s Republic of China, Email [email protected]
Purpose: The systemic infection and inflammation can result in testes injury, whereas the exact mechanism is unknown. The lipid metabolism has a dual impact on controlling metabolism and inflammation, which is a potential pathway. The objective of this study was to determine if changes in plasma lipids during lipopolysaccharide (LPS)-induced systemic inflammation affect the dysregulation of testes lipid metabolism.
Materials and Methods: LPS (5 mg/kg) was used to induce systemic inflammation in rats after a single intraperitoneal injection. After 4 weeks, the rats were sacrificed, and the serum and testes were used for laboratory measurements and histology examination. Plasma and testis were used for lipidomics analysis based the liquid chromatography-mass spectrometry. Spearman rank correlation analysis was used to compare the correlation of differential lipids in phospholipids, glycerolipids, and sphingolipids between testis and plasma.
Results: LPS raised the levels of cytokines in serum and testis, decreased the activities of antioxidant enzymes, increased the levels of lipid peroxidation products, and damaged testis tissue. In testis and plasma, 146 and 401 differential lipids, mostly phosphatidylcholine, phosphatidylethanolamine an so on, were found in comparison to the control group. Correlation analysis produced a total of 2528 correlation coefficients, 1150 of which were P< 0.05 and accounted for 45.49%.
Conclusion: The changes of lipid composition and content in the testis are related to cytokine overload and oxidative stress. Testis lipid metabolism disorders caused by LPS-induced systemic inflammation are lack of a correlation with plasma lipid changes, and are likely owing to interference with the testis itself.
Keywords: systemic inflammation, infertility, lipidomics, oxidative stress, correlation analysis
Introduction
Spermatogenesis is strictly controlled. In the testis, the macrophages, Leydig cells, and Sertoli cells generate cytokines under normal condition. These cytokines control spermatogenesis by influencing cell interaction, producing reactive oxygen species (ROS), and activating Toll-like receptors (TLRs). The pituitary-gonadal axis is affected by the simultaneous actions of cytokines and ROS at various levels to alter testosterone production.1 As a result, spermatogenesis requires a steady environment made up of a number of components, any imbalance of which might result in infertility in males.
Lipopolysaccharide (LPS) is a component of the outer membrane of Gram-negative bacteria. As an endotoxin, LPS can cause an inflammatory response similar to infection. Intraperitoneal injection of LPS can affect steroidogenesis and spermatogenesis, indicating that LPS-induced systemic inflammatory response can cause decreased testis function and lead to impairment of male fertility.2 Systemic inflammation may interfere with testis function because of cytokines indirect effects. LPS binds to LPS binding protein and the auxiliary protein CD14/MD2 to form a complex that activates the TLR4 immune response and produces cytokines. And inflammation and cytokines are regulated at the level of transcription and translation in a testis-specific manner.3 However, intense LPS stimulation leads to over-activation of the immune system via TLR4 and dysregulation of cytokine levels in the testis.4 In addition, during the acute inflammation induced by intraperitoneal injection of LPS, the testis experiences oxidative stress due to the excessive production of ROS, which is the primary factor causing testosterone synthesis disorder and fertility impairment.2 Clearly, LPS-induced systemic inflammatory response can destroy the stable state of spermatogenesis and impair male fertility by stimulating the production of cytokines and ROS, but the precise mechanism remains unclear.
Lipids are essential components of membrane structure, also potent signaling molecules that control a range of cellular reactions. Cytokine overproduction is related to the dysregulation of lipid metabolism induced by LPS.5 For example, when LPS stimulates macrophages, CD14 aggregates in the plasma membrane, causing bidirectional buildup of phosphatidylinositol 4,5-bisphosphate [PI (4,5) P2], which in turn regulates two important inflammatory signaling pathways of TLR4 (Myd88 and TRIF-dependent pathways). PI (4,5) P2 regulates cytokine production by modulating the TLR4 pro-inflammatory signaling pathway.6,7 In addition, LPS-induced lipid metabolism disorders can produce extra saturated fatty acids, such as palmitic acid (C16:0). Palmitic acid promotes the dimerization and translocation of TLR2/4 into lipid rafts at the plasma membrane, which recruit and activate the downstream signaling molecules in the lipid rafts. The downstream signaling molecules synergistically accelerate inflammatory responses with LPS.8 The spermatogenic cells and sperm have a specific lipid composition that mainly including phospholipids, glycerolipids and cholesterol. Throughout various stages of spermatogenesis, lipid content and composition vary and regulate production of ROS and cytokines.9 Thus, dysregulation of lipid metabolism is a potential pathway for testis dysfunction in LPS-induced systemic inflammation. In light of this, we inquire as to whether dysregulation of testis lipid metabolism during LPS-induced systemic inflammation is regulated by alterations in plasma lipids that interfere with testis function. This could be a mechanism of systemic infection or inflammation-induced reproductive reduction, and it is also useful for biomarker screening.
Liquid chromatography-mass spectrometry (LC-MS) is used to characterize the lipid composition, quantify individual lipids, and facilitate to revealing of pathological mechanisms and biomarkers.10 Therefore, by comparing the characteristics of non-targeted lipidomics in plasma and testis of LPS-induced rats and analyzing the correlation of key lipids, this study aims to clarify whether testis lipid metabolism disorder induced by systematic inflammation is secondary to plasma lipid changes or emanating from the testis itself.
Materials and Methods
Animals
A total of 15 sexually mature (6–8 weeks) male Sprague-Dawley rats (SD rats) weighing 260±10g were obtained from Jinling Hospital. After a two-week acclimatization period, the LPS group (n=5) and the control group (n=10) were randomly assigned to the animals. The LPS group rats were intraperitoneally injected with 0.5 mg/mL LPS at 1 mL/100 g body weight (dissolved in saline). The control group was injected with normal saline at 1 mL/100 g body weight. Animals were kept at a constant temperature of 24±2°C with a 12-hour light/dark cycle and free access to food and water. The rats were sacrificed after 4 weeks of continuous observation and samples for testing were obtained. Samples were stored at −80°C. Removed and thawed the samples when used for testing. All the experimental procedures on animals were approved by the Institutional Ethics Committee of Jinling Hospital (No. 2022JLHSXJDWLS-0021) and carried out according to the National Institutes of Health protocol to laboratory animal care and use.
Reagents and Biochemical Test
LPS (L2630-10MG) was purchased from Sigma-Aldrich (USA). The cytokines were measured in the testis and serum using ELISA kits. The kits were obtained from ELK Biotechnology (Wuhan, China). The levels of the cytokines interleukin 1alpha (IL-1α, Cat num: ELK1148), interleukin 1beta (IL-1β, Cat num: ELK1272), tumor necrosis factor-alpha (TNF-α, Cat num: ELK1396), monocyte chemotactic protein 1 (MCP-1, Cat num: ELK5504) and interleukin 6 (IL-6, Cat num: ELK1158) were assessed. Oxidative stress-related indicators assay kits were purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). Glutathione peroxidase (GSH-PX, Cat num: A005-1) and total antioxidant capacity (TAOC, Cat num: A015-1) were measured using colorimetry.11,12 Catalase (CAT, Cat num: A007-1), superoxide dismutase (SOD, Cat num: A001-1) and malondialdehyde (MDA, Cat num: A003-1) were measured by spectrophotometry.13,14
Lipidomics Test
Non-targeted lipidomics study was performed based on LC-MS technology.10 The samples were defrosted. Then 100 μL of sample was taken, and 1.5 mL chloroform/methanol (2/1, v/v) solution and 0.5 mL pure water were added to it. The mixed solution was vortexed for 1 minute. The organic phase was transferred into a new tube and dried. The organic phase was then resolved with 200 μL isopropanol/methanol (1/1, v/v), and mixed with 5 μL internal standard LPC (0.14 mg/mL). Centrifuging at 12,000 rpm for 10 minutes at 4 °C was then performed. Quality Control (QC) samples were created by combining equal metabolic samples. QC samples were inserted in the sample testing procedure to assess system stability throughout the experiment and conduct sample correlation analysis.
The mass spectrometry detection was based on LC-MS (Waters, UPLC; Thermo, Q Exactive Orbitrap).15 The column was ACQUITY UPLC BEH C18 (2.1*100mm 1.7 μm). The chromatographic separation conditions were a flow rate of 0.3 mL/min and a column temperature of 40 °C. Mobile phase elution was then performed according to the procedure. The positive mode mobile phase (A) was made up of acetonitrile and water (6:4, v/v, 10 mM ammonium), and the negative mode mobile phase (B) was made up of isopropyl alcohol and acetonitrile (9:1, v/v, 10 mM ammonium). The injection volume is 1 μL (positive ion mode) and 3 μL (negative ion mode), and the temperature was maintained at 4°C.
Multivariate Analysis
After the data preprocessing, the multivariate analyses, including Principal component analysis (PCA) and Orthogonal projections to Latent Structures-discriminant analysis (OPLS-DA), were performed using SIMCA-P 14.1 (Umeda, Umea, Sweden).16 The PCA model reflected the original state of the data. OPLS-DA was performed to obtain a higher level of group separation and get a better understanding of variables responsible for classification. To evaluate the model’s validity, R2Y and Q2 were obtained after 7-fold cross validation. And the permutation test was used to further test the original model’s reliability. The lipids with significant difference were identified based on the combination of a statistically significant threshold of variable influence on projection (VIP) values obtained from the OPLS-DA model and two-tailed Student’s t-test (P-value). The lipids with VIP>1.0 and P<0.05 were identified as significant.
Histology
The testis was preserved in EP tubes with 4% paraformaldehyde. Gradient dehydration was performed using different percentages of ethanol (80%, 90%, 95%, and 100%). Testis was then impregnated in xylene to transparent. Fully wax-soaked tissues were embedded and sectioned to slices thickness of 5 μm. The slices were immersed in xylene for 10 min to dewaxing. Then the hydration was performed. And the slices were stained with hematoxylin. The stained slices were washed off with water. Ethanol hydrochloride was then used for differentiation. The slices dipped back into the blue with ammonia, rinsed with running water, macerated in eosin staining solution for 3 min. The stained slices were dehydrated with ethanol and made transparent with xylene. After drying, the slices were sealed with gelatin and observed.
Statistical Analysis
The data were analyzed using the statistical software SPSS 26.0. The Analysis of Variance (ANOVA) were used to make comparisons, and differences were considered significant at P<0.05. The measurement data before the ANOVA analysis to normality test and homogeneity test. Spearman rank correlation analysis was used to analyze the correlation between lipids in plasma and testis. Origin 2021 was used to evaluate the significance of the correlation coefficient and draw the heat map of the correlation coefficient.
Results
Effect of LPS on Serum Cytokines
Serum cytokines were measured to assess systemic inflammation induced by LPS. Compared with the control group, intraperitoneal injection of LPS induced significant increases in the serum levels of inflammatory cytokines IL-1α, IL-1β, IL-6 and TNF-α, and chemokine MCP-1 (all P<0.001) (Figure 1).
 |
Figure 1 The cytokine levels in the serum between LPS group and control group. (A) IL-1α, (B) IL-1β, (C) IL-6, (D) TNF-α, (E) MCP-1 Compared with the control group (n=10), the levels of all cytokines in the LPS group rats (n=5) serum were significantly increased, with statistical differences, suggesting that LPS can induce systemic inflammatory response.***P<0.001. |
Effect of LPS-Induced Systemic Inflammation on Cytokines and Oxidative Stress in the Testis
Cytokines and oxidative stress-related indicators in the testis were detected to evaluate the effect of LPS-induced systemic inflammation on the testis. Compared with the control group, all cytokines were significantly increased in the LPS group (all P<0.001) (Figure 2A-E). Meanwhile, the activities of antioxidant enzymes GSH-PX, SOD and CAT were significantly decreased, the level of lipid peroxidation product MDA was significantly increased, and the total antioxidant capacity was decreased (all P<0.001) (Figure 2F-J).
 |
Figure 2 The levels of cytokines and oxidative status indicators in testis between LPS group and control group. (A) IL-1α, (B) IL-1β, (C) IL-6, (D) TNF-α, (E) MCP-1, (F) GSH-PX, (G) SOD, (H) CAT, (I) TAOC, (J) MDA In the testis of rats, the levels of all cytokines in the LPS group (n=5) comparing with the control group (n=10) were significantly increased, the activities of antioxidant enzymes and the total antioxidant capacity were significantly decreased, and the level of lipid peroxidation product MDA was significantly increased. ***P<0.001. |
Histology of Testis
The evaluation of testis histology to further validate that LPS-induced inflammation resulted in testis tissue destruction. Comparing with the control group (Figure 3A and B), the order of the germ lineage in the seminiferous tubules is disrupted in the LPS group. The number of germ cells was dramatically reduced, and normal germ cells vanished. Germ cells that had been damaged dropped off into the lumen. The LPS group had more prominent edema between germ tubules, and the interstitial space became more broadening than the control group (Figure 3C and D).
 |
Figure 3 Effect of LPS-induced systemic inflammation on testis histology between LPS group and control group. (A) Control group (×100), (B) Control group (×200), (C) LPS group (×100), (D) LPS group (×200) In the LPS group, there was a clear edema between the seminiferous tubules, the order of the seminiferous lineage was disappeared in the lumen, and the number of germ cells and sperm was decreased. Abbreviations: LU, lumen; SC, spermatogenic cell; IS, interstitial space. |
Results of Multivariate Analysis of Plasma and Testis
The PCA score plots, as the embodiment of the PCA results, evaluates the overall condition of the data (Figure 4A and B). In the PCA score plot, all scatter points are within the Hotelling T2 circle, indicating that there are no abnormal or outlier conditions in the tested samples. Meanwhile, the sample scatterings representing control and LPS groups were far from each other, indicating significant differences between LPS and control groups. In the OPLS-PA score plot (Figure 4C), the interpretability of the model R2Y=0.984, and the predictability of the model Q2=0.763, indicating that the model was effective and had good predictive ability and repeatability. Similarly, the same results were obtained in the OPLS-PA score plot (Figure 4D), the interpretability of the model was R2Y=0.997, and the predictability of the model was Q2=0.932. Permutation test results showed that the intercept R2Y=0.086 (close to 1), indicating that the model was consistent with the real situation of the sample, and the intercept Q2=−1.01 (less than 0), indicating that the model had strong stability and was effective (Figure 4E). The Permutation test result of testis showed that the intercept was R2Y=0.88, Q2=−1.03 (Figure 4F).
 |
Figure 4 The multivariate analysis of lipidomics in plasma and testis between LPS group and control group. Each sample represents one experiment replicate. (A) PCA score plot (plasma), (B) PCA score plot (testis), (C) OPLS-DA (plasma), (D) OPLS-DA (testis), (E) Permutation (plasma), (F) Permutation (testis) There were no anomalies or outliers in the data used for detection (A-B). The separation tendency between LPS (n=5) and control (n=10) groups was evident, showing that the lipid concentration and composition were significantly different between the two groups of samples and that LPS had a obvious modulation of lipids in plasma and testis (C-D). The validity of the model was further verified by permutation test (E-F) Green circles indicate control samples and red diamonds indicate LPS samples in score plots. |
Identify the Differential Lipids in Plasma and Testis
Differential lipids in plasma and testis were screened according to the criteria of VIP>1&P<0.05. After intraperitoneal injection of LPS, 146 lipids were differentially regulated in plasma, 103 (70.5%) of which were up-regulated and 43 (29.5%) were down-regulated. These lipids were mainly phosphatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidylinositol (PI), sphingomyelin (SM), diglyceride (DG) and triglyceride (TG) (Figure 5A). In testis, 401 lipids were differentially regulated in LPS group compared with control group, of which 174 (43.4%) were up-regulated and 227 (56.6%) were down-regulated. The lipids were Ceramides (Cer), dihexosylceramides (Hex2cer), DG, lysophosphatidylcholine (LPC), lysophosphatidylethanolamine (LPE), lysophosphatidylglycerol (LPG), monoglyceride (MG), PC, PE, phosphatidylglycerol (PG), PI, phosphatidylserine (PS), SM, TG (Figure 5B). These lipids are mainly classified as phospholipids, glycerolipids and sphingolipids.
 |
Figure 5 Comparison of differential lipids in plasma and testis between LPS group and control group. (A) Plasma, (B) Testis Based on the screening criteria of VIP>1andP<0.05, a total of 146 differential lipids were screened in plasma, 103 (70.5%) were up-regulated and 43 (29.5%) were down-regulated. A total of 401 differentially expressed lipids were screened in the testis, 174 (43.4%) were up-regulated and 227 (56.6%) were down-regulated. VIP is indicated by the size of the dot; the bigger the dot, the higher the value of VIP. The dots’ colors show state; red denotes upregulation and blue denotes downregulation. |
Correlation of Phospholipids Between Plasma and Testis
Plasma phospholipids, including PC, PE, LPC and PI, were correlated with testis phospholipids. The correlation coefficient of P<0.05 was considered to be significant, accounting for 53.84%, 36.86%, 39.39% and 36.11%, respectively. Plasma individual phospholipid PC (36:3), PC (24:0/12:4), PC (8:1e/9:0), PC (20:5e), PE (22:0e), LPC (17:0), LPC (26:1) were significantly associated with the majority (over 80%) of testis phospholipids (Figure 6).
 |
Figure 6 Correlation between differential phospholipids in plasma (x-axis) and testis (y-axis) was compared. (A) PC, (B) PE, (C) LPC, (D) PI/PIP The proportions of correlation coefficients of P<0.05 in different phospholipids were PC (53.84%, 533/990), PE (36.86%, 188/510), LPC (39.39%, 156/396) and PI (36.11%, 26/72). *P<0.05, **P<0.01, ***P<0.001. Abbreviations: PC, Phosphatidylcholine, PE, phosphatidylethanolamine, LPC, lysophosphatidylcholine, PI/PIP, Phosphatidylsphingosine. |
Correlation of Glycerolipids Between Plasma and Testis
The correlation analysis between plasma glycerolipids, mainly including DG and TG, and testis glycerolipids showed that the correlation coefficients of P<0.05 were 72.92% and 34.62%, respectively. In plasma, individual glycerolipid DG (18:1/18:2), DG (18:2/18:2); TG (12:0e/6:0), TG (38:7e) were significantly associated with most (beyond 80%) testis glycerolipids (Figure 7).
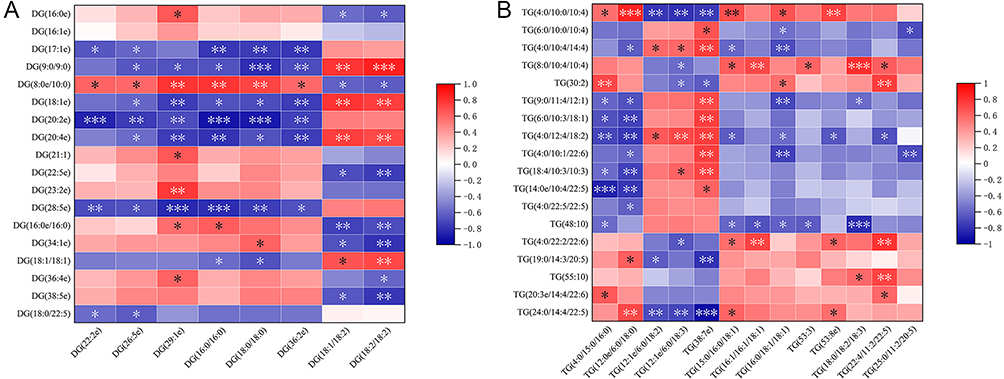 |
Figure 7 Correlation between differential glycerolipids in plasma (x-axis) and testis (y-axis) was compared. (A) DG, (B) TG The proportions of correlation coefficients of P<0.05 in different phospholipids were DG (72.92%, 70/96) and TG (34.62%, 81/234). *P<0.05, **P<0.01, ***P<0.001. Abbreviations: DG, diglyceride, TG, triglyceride. |
Correlation of Sphingolipids Between Plasma and Testis
The correlation between plasma sphingolipids, mainly including SM and Hex2Cer, and testis sphingolipids was analyzed, and the correlation coefficients of P<0.05 were 46.25% and 39.33%, respectively. In plasma, individual sphingolipid SM (s34:1) and SM (d42:1) were significantly associated with the majority (beyond 80%) of testis sphingolipids. Overall, among the 2528 correlation coefficients obtained by comparing plasma and testis lipids, 1150 correlation coefficients were P<0.05, accounting for 45.49% (Figure 8). Table 1 lists more details.
 |
Table 1 Proportion of Correlation Coefficients with P < 0.05 in Comparison of Differential Lipids Between Plasma and Testis |
 |
Figure 8 Correlation between differential sphingolipids in plasma (x-axis) and testis (y-axis) was compared. (A) SM, (B) Hex2Cer The proportions of correlation coefficients of P<0.05 in different phospholipids were SM (46.25%, 37/80) and Hex2Cer (39.33%, 59/150). *P<0.05, **P<0.01, ***P<0.001. Abbreviations: SM, sphingomyelin, Hex2Cer, dihexosylceramides. |
Discussion
In this study, we examined LPS-induced systemic inflammation and its impact on testis. In addition, lipids in plasma and testis were identified using LC-MS, and differential lipids were then screened using multivariate statistical analysis. Moreover, Spearman rank correlation analysis of differential lipids, primarily phospholipids, glycerolipids and sphingolipids, was used to compare the correlation of these lipids in plasma and testis.
The rise in serum cytokines after intraperitoneal injection of LPS suggested that a systemic inflammation had been induced. The testis’ stable environment for spermatogenesis was damaged by systemic inflammation, as evidenced by the rise in cytokines, decline in antioxidant enzyme activity, and rise in the amount of MDA, a lipid peroxidation product. The histology provided further evidence of structural and functional damages to the testis, and these damages are continuous. These results are consistent with reports from previous studies.2,17
Germ cells and sperm plasma membranes possess a specific lipid composition, mainly contains phospholipids (70%), glycerolipids (25%), and glycolipids (5%). Phospholipids containing choline and ethanolamine, which can be classified as PC, PE, PI, PG, etc., and are characterized by a high percentage of polyunsaturated fatty acids, such as arachidonic acid (20:4), eicosapentaenoic acid (DPA, 22:5), and docosahexaenoic acid (22:6, DHA).18 Germ cells and sperm have a high glycerolipids proportion in comparison to other type cells, especially containing high concentration of DG.19 This specific lipid composition affects sperm membrane fluidity, adhesion, and signaling.20 Plasma lipid content and composition changed as result of systemic inflammation induced by LPS, which was related to cytokine overload and oxidative stress. Our other study revealed a correlation between LPS-induced differential lipids and cytokines and oxidative status indicators, indicating that the regulation of lipid content and composition to cytokines and oxidative status is the potential pathway to control epididymis inflammation and oxidative stress injury.21 In this research, we studied the testis lipidomics, which were differentially regulated during LPS-induced systemic inflammation, including Cer, Hex2cer, DG, LPC, LPE, LPG, MG, PC, PE, PG, PI, PS, SM, TG. This forced regulation concerned the testis cytokines and ROS production. The male reproductive organs can secrete some proteins with antioxidant activity, such as GSH-PX, and bind to the sperm plasma membrane,22 suggesting that GSH-PX on the sperm plasma membrane can defend the membrane PC from ROS attack and subsequent oxidative stress damage. It has been shown that GSH-PX activity and PC content are positively correlated in healthy male seminal plasma, while LPC, the PC peroxidation product, is negatively correlated.18 Also, SOD positively correlated with linolenic acid, arachidonic acid, and DHA in the seminal plasma of healthy men. According to research, the phospholipids-based specific membrane structures adsorb GSH-PX and SOD in seminal plasma, strengthening sperm potential for antioxidant defense.23 At present study, LPS-induced changes in the lipids of the testis may have decreased the activity of antioxidant enzyme, exposing the more PUFA in sperm membranes to ROS. In another study, Xu et al used LC-MS to detect the seminal plasma of different types of infertile patients, and screened 17 metabolites related to sperm parameters. Among them, lipids lysoPE (16:0/0:0) and lysoSM (d18:0) were related to sperm co ncentration and sperm motility, and unsaturated fatty acids were related to sperm malformation and sperm motility.24 These studies suggest that cytokine production and oxidative stress through dysregulation of testis lipid metabolism impact spermatogenesis, sperm structure, and function. However, these studies mainly concentrated on seminal plasma, testis lipidomics still lacks a thorough illustration. In our research, changes of testis lipid characteristics resulted from LPS-induced systemic inflammation were well presented using LC-MS.
Plasma lipids were simultaneously changed under LPS-induced systemic inflammation. Compared with the plasma differential lipid distribution, the amount and type of differential lipids in the testis are more complex, and the dominant regulation state of differential lipids is also significantly different. Down-regulated lipids made up the majority of the differential lipids in the testis (56.6%), but up-regulated lipids predominated in the plasma (70.5%). By comparing the distribution of differential lipids between plasma and testis, the relationship between plasma lipid changes and testis lipid metabolism disorders was preliminarily estimated.
For further correlation analysis, the key differential lipid types—PC, PE, LPC, PI, DG, TG, SM, and Hex2Cer—were chosen. They were chosen since they differ from other lipids by one or more orders of magnitude. Its use as indicators to assess relationships is therefore persuasive and representative. The results showed that PC and DG in plasma above a 50% correlation with testis differential lipids (53.84% and 72.92%, respectively). A total of 45.49% of the correlation coefficients were P<0.05, indicating that disordered testis lipid metabolism caused by LPS-induced systemic inflammation was few related to plasma lipid changes. There are several explanations for this. The testis, like the brain, is an immunologically privileged organ with special inherent protective barriers (the blood-testis barrier and the blood-brain barrier), which insulate the protected organ from outside influences.25 When comparing PCs concentrations in paired cerebrospinal fluid (CSF) and serum samples, levels of the top 10 PCs increased significantly in CSF, but their concentrations in serum did not change significantly, suggesting that the changes in CSF do not reflect systemic inflammation but are instead directly caused by local disease.26 In addition, the imbalance of lipid concentrations in organs and peripheral blood may also result from altered membrane signaling, catabolism of free phospholipase and other enzymes in the blood,27 although the precise mechanisms still need to be clarified in further expanded experiments.
Conclusions
LPS induces systemic and testicular inflammation, 146 and 401 differential lipids were detected in the plasma and testis, respectively. Changes in lipid content and composition were linked to impaired testis function. The correlation of key differential lipids in plasma and testis was then assessed, with PC and DG, showing the highest correlation. Overall, out of 2528 correlation coefficients, 45.49% had the significance (less than 50%). Conclusively, the testis lipid metabolism disorders induced by LPS-induced systemic inflammation maybe not resulted from plasma lipid changes, but the impacts on the testis itself.
Abbreviations
ROS, reactive oxygen species; TLRs, Toll-like receptors; LPS, Lipopolysaccharide; PI (4,5) P2, phosphatidylinositol 4,5-bisphosphate; LC-MS, Liquid chromatography-mass spectrometry; PCA, Principal component analysis; OPLS-DA, Orthogonal projections to Latent Structures-discriminant analysis; VIP, variable influence on projection; PC, phosphatidylcholine; PE, phosphatidylethanolamine; PI, phosphatidylinositol; SM, sphingomyelin; DG, diglyceride; TG, triglyceride; Cer, Ceramides; Hex2cer, dihexosylceramides; LPC, lysophosphatidylcholine; LPE, lysophosphatidylethanolamine; LPG, lysophosphatidylglycerol; MG, monoglyceride; PG, phosphatidylglycerol; PS, phosphatidylserine.
Data Sharing Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Ethics Approval for Animal Study
The experiment was authorized by the Experimental Animal Ethics Committee of East Region Military Command General Hospital (Jinling Hospital) and carried out according to the National Institutes of Health protocol to laboratory animal care and use.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (NO. 81771646).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Syriou V, Papanikolaou D, Kozyraki A, Goulis DG. Cytokines and male infertility. Eur Cytokine Netw. 2018;29(3):73–82. doi:10.1684/ecn.2018.0412
2. Reddy MM, Mahipal SV, Subhashini J, et al. Bacterial lipopolysaccharide-induced oxidative stress in the impairment of steroidogenesis and spermatogenesis in rats. Reprod Toxicol. 2006;22(3):493–500. doi:10.1016/j.reprotox.2006.03.003
3. O’Bryan GO, Nikolic-Paterson DJ, et al. Cytokine profiles in the testes of rats treated with lipopolysaccharide reveal localized suppression of inflammatory responses. Am J Physiol Regulatory Integr Comp Physiol. 2005;288(6):56.
4. Sahnoun S, Sellami A, Chakroun N, et al. Human sperm Toll-like receptor 4 (TLR4) mediates acrosome reaction, oxidative stress markers, and sperm parameters in response to bacterial lipopolysaccharide in infertile men. J Assist Reprod Genet. 2017;34(8):1067–1077. doi:10.1007/s10815-017-0957-8
5. Kwiatkowska K, Ciesielska A. Lipid-mediated regulation of pro-inflammatory responses induced by lipopolysaccharide. Postepy Biochem. 2018;64(3):175–182. doi:10.18388/pb.2018_129
6. Plociennikowska A, Zdioruk MI, Traczyk G, Swiatkowska A, Kwiatkowska K. LPS-induced clustering of CD14 triggers generation of PI(4,5)P2. J Cell Sci. 2015;128(22):4096–4111. doi:10.1242/jcs.173104
7. Plociennikowska A, Hromada-Judycka A, Dembinska J, Roszczenko P, Ciesielska A, Kwiatkowska K. Contribution of CD14 and TLR4 to changes of the PI(4,5)P2 level in LPS-stimulated cells. J Leukoc Biol. 2016;100(6):1363–1373. doi:10.1189/jlb.2VMA1215-577R
8. Hwang DH, Kim JA, Lee JY. Mechanisms for the activation of Toll-like receptor 2/4 by saturated fatty acids and inhibition by docosahexaenoic acid. Eur J Pharmacol. 2016;785:24–35. doi:10.1016/j.ejphar.2016.04.024
9. Collodel G, Moretti E, Noto D, Iacoponi F, Signorini C. Fatty Acid Profile and Metabolism Are Related to Human Sperm Parameters and Are Relevant in Idiopathic Infertility and Varicocele. Mediators Inflamm. 2020;2020:3640450. doi:10.1155/2020/3640450
10. Dunn WB, Broadhurst D, Begley P, et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat Protoc. 2011;6(7):1060–1083. doi:10.1038/nprot.2011.335
11. Long SH, Yu ZQ, Shuai L, et al. The hypoglycemic effect of the kelp on diabetes mellitus model induced by alloxan in rats. Int J Mol Sci. 2012;13(3):3354–3365. doi:10.3390/ijms13033354
12. Mahfouz R, Sharma R, Sharma D, Sabanegh E, Agarwal A. Diagnostic value of the total antioxidant capacity (TAC) in human seminal plasma. Fertil Steril. 2009;91(3):805–811. doi:10.1016/j.fertnstert.2008.01.022
13. Pakpahan C, Rezano A, Margiana R, Amanda B, Agustinus A, Darmadi D. The Association Between Lipid Serum and Semen Parameters: a Systematic Review. Reprod Sci. 2022;30(3):761–771. doi:10.1007/s43032-022-01040-8
14. Unsal V, Kolukcu E, Gevrek F, Firat F. Sinapic acid reduces ischemia/reperfusion injury due to testicular torsion/detorsion in rats. Andrologia. 2021;53(8):e14117. doi:10.1111/and.14117
15. Wang J, Christison TT, Misuno K, et al. Metabolomic profiling of anionic metabolites in head and neck cancer cells by capillary ion chromatography with Orbitrap mass spectrometry. Anal Chem. 2014;86(10):5116–5124. doi:10.1021/ac500951v
16. Wiklund S, Johansson E, Sjostrom L, et al. Visualization of GC/TOF-MS-based metabolomics data for identification of biochemically interesting compounds using OPLS class models. Anal Chem. 2008;80(1):115–122. doi:10.1021/ac0713510
17. O’Bryan MK, Schlatt S, Phillips DJ, de Kretser DM, Hedger MP. Bacterial lipopolysaccharide-induced inflammation compromises testicular function at multiple levels in vivo. Endocrinology. 2000;141(1):238–246. doi:10.1210/endo.141.1.7240
18. Alvarez JG, Storey BT. Differential incorporation of fatty acids into and peroxidative loss of fatty acids from phospholipids of human spermatozoa. Mol Reprod Dev. 1995;42(3):334–346. doi:10.1002/mrd.1080420311
19. Wood PL, Scoggin K, Ball BA, Troedsson MH, Squires EL. Lipidomics of equine sperm and seminal plasma: identification of amphiphilic (O-acyl)-omega-hydroxy-fatty acids. Theriogenology. 2016;86(5):1212–1221. doi:10.1016/j.theriogenology.2016.04.012
20. Van Tran L, Malla BA, Kumar S, Tyagi AK. Polyunsaturated Fatty Acids in Male Ruminant Reproduction - A Review. Asian-Australas J Anim Sci. 2017;30(5):622–637. doi:10.5713/ajas.15.1034
21. Li Y, Zhu J, Zhao X, et al. Oral Lycopene Administration Attenuates Inflammation and Oxidative Stress by Regulating Plasma Lipids in Rats with Lipopolysaccharide-Induced Epididymitis. J Inflamm Res. 2022;15:6517–6531. doi:10.2147/JIR.S380785
22. Rejraji H, Vernet P, Drevet JR. GPX5 is present in the mouse caput and cauda epididymidis lumen at three different locations. Mol Reprod Dev. 2002;63(1):96–103. doi:10.1002/mrd.10136
23. Tavilani H, Goodarzi MT, Doosti M, et al. Relationship between seminal antioxidant enzymes and the phospholipid and fatty acid composition of spermatozoa. Reprod Biomed Online. 2008;16(5):649–656. doi:10.1016/S1472-6483(10)60478-6
24. Xu Y, Lu H, Wang Y, Zhang Z, Wu Q. Comprehensive metabolic profiles of seminal plasma with different forms of male infertility and their correlation with sperm parameters. J Pharm Biomed Anal. 2020;177:112888. doi:10.1016/j.jpba.2019.112888
25. Qu N, Ogawa Y, Kuramasu M, Nagahori K, Sakabe K, Itoh M. Immunological microenvironment in the testis. Reprod Med Biol. 2020;19(1):24–31. doi:10.1002/rmb2.12293
26. Al-Mekhlafi A, Suhs KW, Schuchardt S, et al. Elevated Free Phosphatidylcholine Levels in Cerebrospinal Fluid Distinguish Bacterial from Viral CNS Infections. Cells. 2021;10(5):1115. doi:10.3390/cells10051115
27. Arshad H, Alfonso JCL, Franke R, et al. Decreased plasma phospholipid concentrations and increased acid sphingomyelinase activity are accurate biomarkers for community-acquired pneumonia. J Transl Med. 2019;17(1):365. doi:10.1186/s12967-019-2112-z
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.