")
Back to Journals » International Journal of Women's Health » Volume 16
Genetic Analysis of a Mosaic Fra(16)(q22)/Del(16)(q22) Karyotype in a Primary Infertile Woman
Authors He G , Wang X, Li B, Wang L, Zhang J, Shi Y , Zhu W, Shi M
Received 1 December 2023
Accepted for publication 29 March 2024
Published 16 April 2024 Volume 2024:16 Pages 637—644
DOI https://doi.org/10.2147/IJWH.S450272
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Everett Magann
Guiyuan He,1,* Xi Wang,1,* Beiqing Li,1 Lei Wang,1 Jing Zhang,2 Yang Shi,1 Wenxiu Zhu,1 Ming Shi1,3
1Centre for Reproductive and Genetic Medicine, Dalian Women and Children’s Medical Group, Dalian, People’s Republic of China; 2Department of Clinical Laboratory, Central Hospital of Dalian University of Technology, Dalian Municipal Central Hospital, Dalian, People’s Republic of China; 3Department of Clinical Laboratory, Dalian Women and Children’s Medical Group, Dalian, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ming Shi, Centre for Reproductive and Genetic Medicine & Department of Clinical Laboratory, Dalian Women and Children’s Medical Group, Dalian, 116037, People’s Republic of China, Email [email protected]
Purpose: Fragile sites are specific chromosomal regions showing gaps, poor staining, contractions, or even breaks in the chromosomes. These spontaneous and heritable fragile sites are prone to structural variations which can lead to adverse reproductive outcomes. This paper aims to present a specific case study of a female patient, with a mosaic karyotype involving chromosome 16q22 fragile site which is very rare in clinic and her experience of infertility.
Case Presentation: A 37-year-old woman is diagnosed with ten-year primary infertility. She worked in a factory, and she was occasionally exposed to paint. She underwent two cycles of follicular monitoring with intrauterine insemination (IUI) using her husband’s sperm six years ago but failed. Most of her prepregnancy tests were normal, except a not smooth right fallopian tube. Her G-band karyotype of peripheral blood lymphocytes was mos 46, XX, del(16)(q22)[40]/46, XX, fra(16)(q22)[29]/46, XX, fra(16)tr(16)(q22)[3]/46, XX[28] which inherited from her mother. The SCE assay detected a significantly higher frequency of SCEs in the 16q region of the patient’s chromosomes compared to her mother and a healthy control. However, the average SCEs per chromosome were quite close. Moreover, copy number variation (CNV) sequencing showed no deletion nor duplication at 16q22.
Conclusion: Infertility cannot be completely attributed to the fragile site on chromosome 16q22. Assisted reproductive technology combined with preimplantation genetic testing may help in achieving a healthy live birth.
Keywords: fragile sites, infertility, fra(16)(q22), del(16)(q22), SCE, CNV
Introduction
Fragile sites are specific chromosomal regions that can be inherited and may lead to a partial inhibition of DNA replication when chromosomes are exposed to certain chemicals. This inhibition can result in gaps, poor staining, contractions, or even breaks in the chromosomes. During mitosis, these regions of chromatin do not bind tightly.1,2 In 1965, Dekaban reported the first fragile site on a human chromosome,3 which has since led to two significant discoveries in recent years. The first discovery involves a fragile site on the X chromosome, specifically in the q27.3 region. This fragile site is associated with conditions such as X-linked mental retardation, primary ovarian insufficiency, and fragile X-associated tremor/ataxia syndrome.4,5 The second discovery is that different fragile sites can be induced through chemical treatment during cell culture karyotyping.6 Chromosomal fragile sites are categorized as common or rare based on their prevalence in the population. Moreover, fragile sites in each category can be subdivided. Certain common fragile sites can be induced by aphidicolin, while others are sensitive to bromodeoxyuridine (BrdU) or 5-azacytidine induction.1 In addition to chemically induced fragile sites, there are spontaneous fragile sites that occur without external induction. These spontaneous and heritable fragile sites are prone to structural variations.7
Most known inducers of fragile sites have the potential to hinder DNA replication elongation, particularly at fragile sites.7 Common fragile sites are found in nearly all individuals and are known to coincide with breakpoints in non-random chromosome rearrangements in cancer. On the other hand, rare fragile sites are present in about 5% of the population, inherited in a Mendelian fashion, and are characterized by nucleotide repeat amplification.8
Fragile sites on chromosome 16 include five distinct locations designated as FRA16A, FRA16B, FRA16C, FRA16D, and FRA16E. Among these, FRA16A, FRA16B, and FRA16E are three rare fragile sites situated at 16p13.11, 16q22.1, and 16p12.1, respectively. FRA16A is folate-sensitive, FRA16B can be induced by distamycin-A or BrdU, while FRA16C and FRA16D are common fragile sites located at 16q22.1 and 16q23.2, respectively, and can be induced by aphidicolin.1 FRA16B was identified through positional cloning, revealing an expanded 33-bp AT-rich minisatellite repeat, p(ATATATTATATATTATATCTAATAATATATC/ATA)n.9
A study by Schmid M10 involving 350 unselected individuals indicated that fra(16)(q22) is the most common among all rare autosomal fragile sites in humans, with a heterozygous frequency of 5.1% in the European population.10 Another study by Japanese researchers found that the incidence of fra(16)(q22) induced by distamycin A was 1.42%.11 Homozygosity for fra(16)(q22) cases was exceptionally rare, with only a few reported instances.12 Statistical analysis revealed that the genotype of fra(16)(q22) on both fragile and non-fragile chromosomes in the population was in Hardy-Weinberg equilibrium.10
In general, individuals carrying fra(16)(q22) do not exhibit significant clinical differences compared to those with normal chromosomes. Some cases have reported a potential link between fra(16)(q22) and benign chronic neutropenia, a condition characterized by low levels of neutrophils in the blood. This suggests that fra(16)(q22) may be involved in the etiology of this condition.13 There are reports suggesting a connection between fra(16)(q22) and acute nonlymphocytic leukemia.14 Studies have indicated that fra(16)(q22) might serve as an induction factor for balanced translocations and inversions.15 These chromosomal rearrangements can lead to genetic diversity and can be associated with certain genetic disorders. Cases of individuals carrying fra(16)(q22) and its relationship to fertility are rarely reported. This paper aims to present a specific case study of a female patient, with a mosaic karyotype involving chromosome 16q22 fragile site and her experience of infertility.
Patient and Methods
Patient Presentation
The female patient, along with her husband, had been experiencing difficulty conceiving for 10 years despite not using any contraception. The couple are both Chinese and 37 years old. Several medical examinations and treatments have been conducted. The findings are listed as below.
Fallopian Tube Examination (2017): A salpingography in 2017 showed that the patient’s left fallopian tube was smooth, but the right fallopian tube was not smooth when the iodine-containing contrast medium was passed through. Fertility Treatments: The couple had undergone two cycles of follicular monitoring with intrauterine insemination (IUI) using her husband’s sperm in other hospital, but these attempts were not successful. Menstrual Cycle and Ovarian Health: The patient’s menstrual cycle was reported as normal, and transvaginal ultrasound scanning revealed that the bilateral ovaries had about 6–7 antral follicles with a dominant follicle and evidence of ovulation. The endometrium was also reported to be in good condition. Hormone Levels: Biochemical analyses indicated that the patient’s blood levels of estradiol, progesterone, testosterone, thyroid-stimulating hormone, prolactin, follicle-stimulating hormone, luteinizing hormone, and anti-Mullerian hormone were all within the normal range. Autoimmune and Infectious Disease Testing: Testing for various antibodies, including antinuclear antibody, anticardiolipin antibody IgM and IgG, and beta-2 glycoprotein 1 antibody IgM, IgG, and IgA, all returned negative results. The patient had a history of mumps infection during her childhood. Occupational Exposure: The patient worked in a factory and was occasionally exposed to paint, suggesting a potential occupational exposure risk factor that might be relevant to her fertility concerns. Husband’s Sperm Analysis: The total sperm count of the patient’s husband was below the normal range (≥39.0 × 10^6), with a count of 11.6 × 10^6, but the motility of the sperm was reported as normal.
Material Methods
G-Band Karyotype of Peripheral Blood Lymphocytes
Informed consents were signed by the patient and her husband, and the case information was only allowed to be used for academic study and publication purposes.
Two-milliliter sample of peripheral blood was collected from the patient and her family members. A sample of 0.5 mL peripheral blood was inoculated into 4mL pre-warmed Roswell Park Memorial Institute (RPMI) 1640 medium containing phytohaemagglutinin (PHA) (Weihaixinpu, Shandong, China). Cell cultures were incubated at 37°C in a CO2 cell culture incubator (Sanyo Electric Biomedical, Japan) for 72hrs. After the 72-hour incubation, 20 µL of colchicine (Weihaixinpu, Shandong, China) was added to the cultures to achieve a final concentration of 0.1μg/mL. After 2 hrs of incubation, samples were swelled with 0.075 M KCI solution and fixed in 3:1 methanol: glacial acetic acid. Cell suspensions were dropped onto glass slides and allowed to dry. These slides were then placed in an oven set at 60°C for overnight. The dried slides were digested in 0.05% trypsin solution (Weihaixinpu, Shandong, China) at 37°C for 2 to 5 minutes and stained with Giemsa stain (Baso Diagnostics Inc. Zhuhai, China) for 1 to 2 minutes. The slides were then rinsed with tap water and allowed to dry. Metaphases cell imagines were acquired with a Leica CytoVision GSL 120 cytogenetics platform (Leica, Germany). One hundred metaphases were analyzed for each sample using Cytovision® karyotyping software version 7.4.1. The analysis was performed according to the International System for Human Cytogenomics Nomenclature 2020 (ISCN, 2020).16
Copy Number Variation (CNV) Sequencing
CNV sequencing libraries were constructed from the DNA of the patient. DNA fragmentation and ligation with the adapters were according to the standard procedure.17 The constructed libraries were sequenced using BGISEQ-500 sequencing platforms as paired-end 100bp reads. High-quality sequencing reads were processed using DRAGEN software, including alignment to the human reference genome, sorting and duplicate marking.
Sister Chromatid Exchange (SCE)
Cell cultures of samples from the patient, the patient’s mother and a female healthy control were carried out as described in G-band karyotyping method mentioned above. Peripheral blood sample (0.8mL) was inoculated in 10mL RPMI1640 medium containing PHA. BrdU (Solarbio, Beijing, China) was added to the cultures at a final concentration of 10 µg/mL, 24hrs prior to cell harvest. Colchicine treatment, hypotonicity, fixation, cell dropping and oven heating were the same as in karyotyping. The dried glass slides were covered with 2 × saline sodium citrate (SSC) buffer and exposed under a 30W UV light for 30 min. Metaphase cell imagines were acquired with a Leica CytoVision GSL 120 cytogenetics platform (Leica, Germany). For each sample, 30 metaphases were analyzed, and the exchange times of sister chromatids were recorded according to the method described by M. Schmid.18
Results
G-Band Karyotype of Peripheral Blood Lymphocytes
Both the husband and the father of the patient had a normal 46, XY karyotype. Karyotype of patient was analyzed twice. The first result indicated a mosaic karyotype:
mos 46, XX, del(16)(q22)[40]/46, XX, fra(16)(q22)[29]/46, XX, fra(16)tr(16)(q22)[3]/46, XX[28].
(Figure 1a–d). The patient had 46 chromosomes per cell. There were four different cell lines present, and the most common cell line had a deletion in the q22 region of chromosome 16 and constituted 40% of the analyzed cells. The next most common cell line had a fragile site in the same region and constituted 29% of the cells. Another cell line had a triplication of 16q22 and constituted 3% of the cells. The remaining 28% of the cells had a normal karyotype. One month later, the patient was offered a second karyotyping and the cell line with del(16)(q22) was decreased to the second most and the percentage of the normal 46, XX cell was increased to 42%. The karyotype was
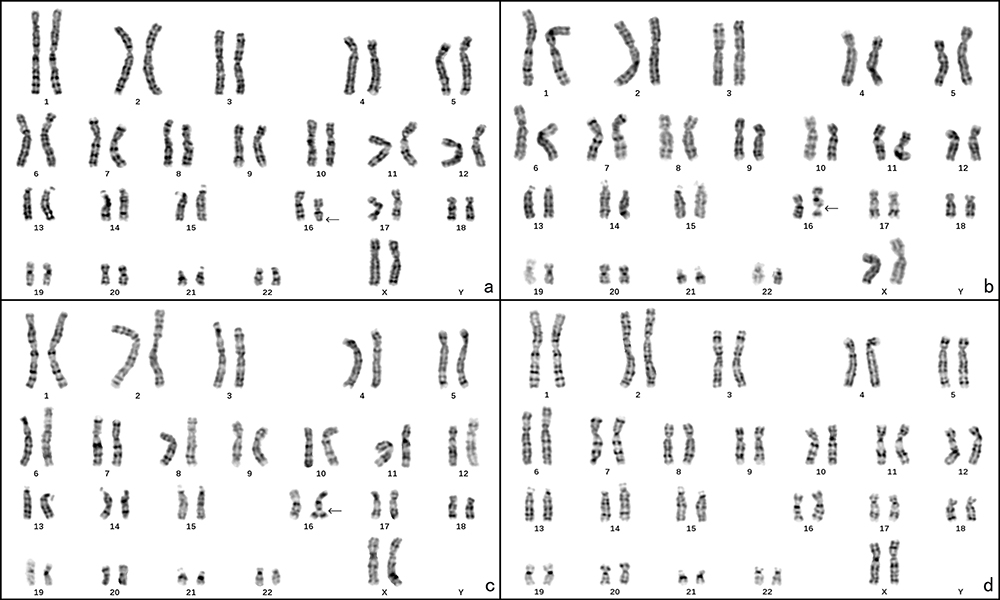 |
Figure 1 The chromosome karyotype of the patient. (a) 46, XX, del(16)(q22), black arrow points the deletion at 16q22; (b) 46, XX, fra(16)(q22), black arrow points the fragile site at 16q22; (c) 46, XX, fra(16)tr(16)(q22), black arrow points the duplication at 16q22; (d) 46, XX. |
mos 46, XX, del(16)(q22)[26]/46, XX, fra(16)(q22)[24]/46, XX, fra(16)tr(16)(q22)[8]/46, XX[42].
The mother of the patient also displayed a mosaic karyotype which was: mos 46, XX, del(16)(q22)[16]/46, XX, fra(16)(q22)[46]/46, XX, fra(16)tr(16)(q22)[6]/46, XX[32] (Figure 2a–d and Table 1).
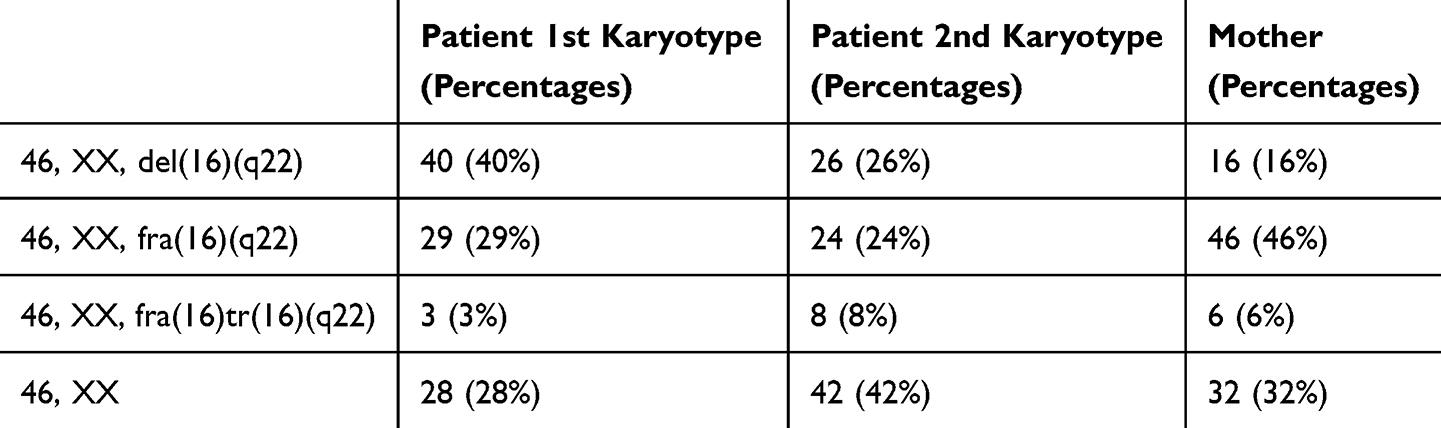 |
Table 1 The Percentages of Different Cell Lines in the Mosaic Karyotypes of the Patient and Her Mother |
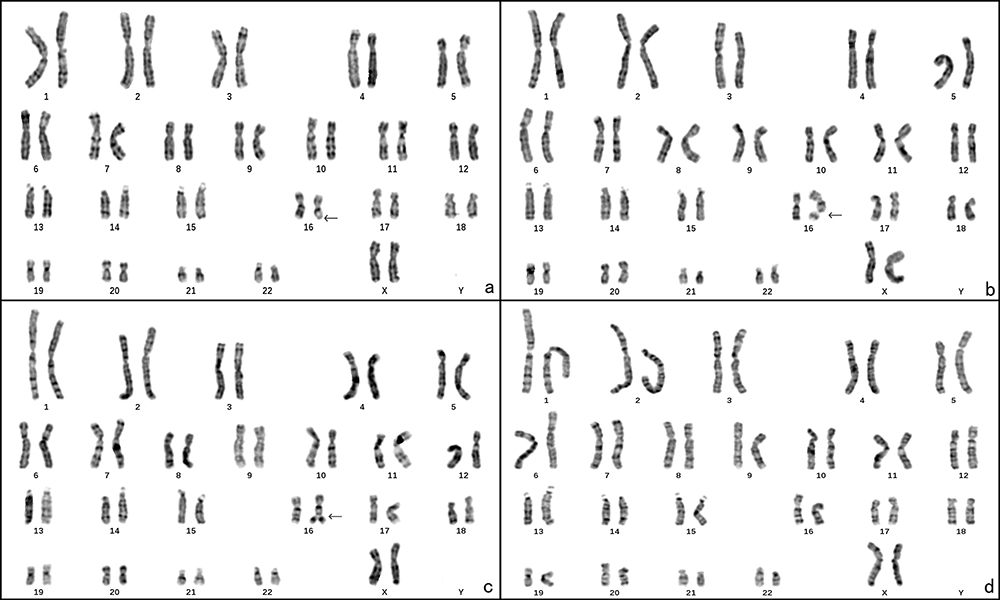 |
Figure 2 The chromosome karyotype of the patient’s mother. (a) 46, XX, del(16)(q22), black arrow points the deletion at 16q22; (b) 46, XX, fra(16)(q22), black arrow points the fragile site at 16q22; (c) 46, XX, fra(16)tr(16)(q22), black arrow points the duplication of 16q22; (d) 46, XX. |
Copy Number Variation (CNV) Sequencing
As the 46, XX, del(16)(q22) cells took up a quite high percentage in the patient’s cell culture, a copy number variation test was offered to the patient to identify any deletion in the genome. Surprisingly, the result of CNV sequencing indicated that the copy number of the patient was a normal diploid. No pathogenic or likely pathogenic deletion and duplication was detected at 16q22. A 239.07Kb duplication in chromosomal region 1q21.3q21.3 (sseq[GRCh37]1q21.3q21.3(152339130_152578202)×3 chr1:g.152339130_152578202dup) was detected which contained 7 protein coding genes. According to the scoring system of ClinGen, Decipher and DGV, the duplication was a variation of unknown clinical significance.
The CNV sequencing also discovered a 162.36Kb of duplication in the 2p22.3p22.3 chromosomal region (sseq[GRCh37]2p22.3p22.3(32689866_32852229)×3 chr2:g.32689866_32852229dup) which contained 1 protein coding gene. According to the scoring system of ClinGen, Decipher and DGV, the duplication was also a variation of unknown significance.
Sister Chromatid Exchange (SCE) Assay
The SCE assay showed that the patient, patient’s mother and a female healthy control all displayed certain exchanges between sister chromatids (Figure 3). The SCE assay detected a significantly higher frequency of SCEs in the 16q region of the patient’s chromosomes compared to her mother and a healthy control (Table 2). The average SCEs per chromosome were quite close in all the tested subjects and ranged from 0.11 to 0.13 (Table 2). These SCEs were in good agreement with the baseline frequencies of SCEs in BrdU-treated human lymphocytes reported in the literature references.19,20
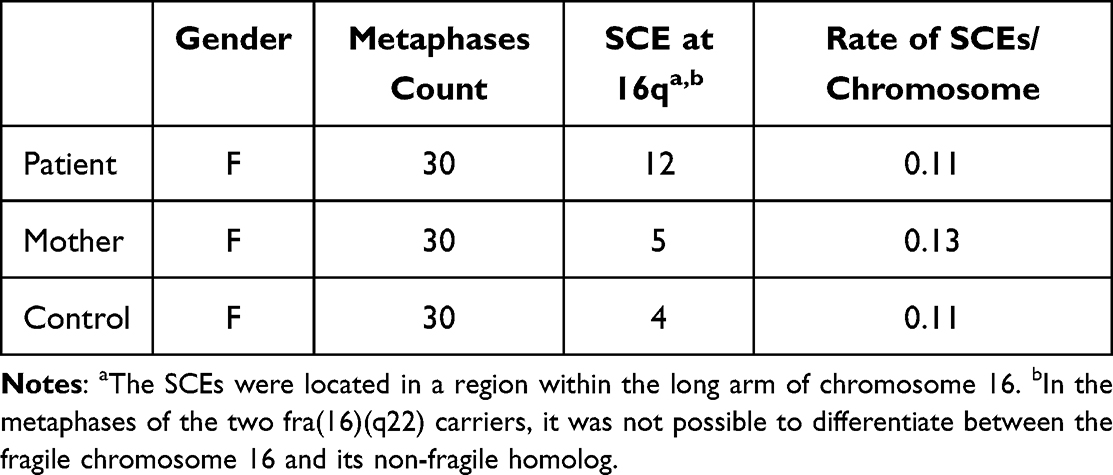 |
Table 2 Frequencies of Sister Chromatid Exchanges in Metaphases of Patient, Patient’s Mother and a Healthy Control |
 |
Figure 3 The SCE experiments of patient, patient’s mother and a healthy control. (a) A metaphase of the patient’s SCE; (b) A metaphase of the patient’s mother’s SCE; (c) A metaphase of the control’s SCE; (d) The patient’s SCE were located in a region within the long arm of chromosome 16, arrow points the SCE site on 16q; (e) The patient’s mother’s SCE were located in a region within the long arm of chromosome 16, arrow points the SCE site on 16q; (f) The normal control’s SCE were located in a region within the long arm of chromosome 16, arrow points the SCE site on 16q. |
Discussion
Spontaneous fragile site is extremely rare, and this patient is the first case we have encountered in more than 27,000 patients since 2014, when we started performing karyotyping in our hospital. The incidence of such spontaneous fragile sites has not been previously reported. Since karyotypes of the patient and her mother showed a significant proportion of cells with del(16)(q22), to pinpoint the exact location of the fragile site and the underlying genetic abnormality, a CNV sequencing test was performed on the patient. However, the results were a normal diploid person. This outcome is surprising given the presence of the mosaic fra(16)(q22). Two potential explanations were offered for the negative CNV sequencing results. First, it is suggested that cells with deletion or duplication of 16q22 may have a shorter lifespan compared to normal leukocytes, which means that most of the leukocytes in the tested sample may have a normal karyotype. Second, it is hypothesized that the deletion and duplication of 16q22 cells may have arisen from the same mitotic error, resulting in similar proportions of these two cell lines. When CNV sequencing was carried out, these two cell lines could potentially compensate for each other, leading to the observed normal diploid result. A FISH experiment could verify this hypothesis well. However, currently, we cannot precisely locate the fragile site, a site-specific fluorescent probe for 16q22 is needed. Since cells with del(16)(q22) in both patient and her mother take more percentage than cells with dup(16)(q22), it is suggested that deletion cells have good viability in in vitro culture.
The Sister Chromatid Exchange (SCE) assay is a cytogenetic technique used to assess the genetic stability of a person’s chromosomes, particularly to measure the occurrence of exchanges between sister chromatids. An increased frequency of SCEs can be an indicator of genomic instability and potentially an underlying genetic condition or exposure to certain genotoxic agents. Our results suggested that the frequency of SCEs in chromosome 16 indeed is higher in patient than in her mother and healthy control; however, there was no significant difference of average ratio of SCE in each person.
According to the literature, fra(16)(q22) is usually benign, without pathogenic effect. The patient in this case is an infertile woman. Based on the results of the various clinical tests she and her husband carried out; it is important to consider the possibility of issues related to fallopian tube functionality, sperm count, and potential occupational factors affecting fertility. Further evaluation and investigation may be necessary to determine the specific cause of the couple’s infertility and to explore appropriate treatment options. The infertility of the couple could not completely be attributed to the mosaic fragile site of 16q22, as the mother of the patient shared this karyotype, and she could conceive naturally. In addition, if there were 16q22 deletion or duplication in her oocytes, probably most embryos of these fertilized oocytes will lead to spontaneous abortion or abnormal fetal development. This woman had never being pregnant since married for the past 10 years. Previous case report and clinical study both indicated that embryos from mosaic fragile carriers have certain ratio of dup or del chromosome which is related with its fragile sites.21,22 Luo et al studied 12 couples with heterozygous mosaic fragile 16q22 on their chromosomes. These couples underwent preimplantation genetic testing for their embryos. The results indicated that 5 embryos out of 56 (8.93%) showed an extra 16q21-q24.3 copy number variant (CNV) associated with their parental fragile site at 16q22. The results indicate that fragile sites on chromosome 16q could slightly increase the risk of forming embryos with chromosomal abnormalities.
By sequencing, the molecular basis for most of fragile sites was elucidated, constituted by certain expanded tandem repeat of DNAs.7 The constitution of most folate sensitive fragile sites is in the form of CGG/CCG tandem repeat expansion. Typical example is FRAXA, which is genetic basis for fragile X syndrome. Another form of fragile site’s constitution is the expansion of a 33 bp AT-rich minisatellite repeat (which is found in fra 16q22). Fragile sites are prone to chromosomal rearrangements-deletions or translocations which found in many cancers.2,7,8 Fragile sites are also correlated with chromosomal rearrangements that might contribute to the development of vertebrate lineage and genome reorganization during evolution, although the carrier of the fragile(16)(q22) did not show.7 To sum up, fra(16)(q22) appears to have complex and potentially diverse health influences, including associations with blood disorders, cancer, and fertility issues. However, further research is needed to fully understand the implications of this chromosomal abnormality.
Conclusion
This paper presents a case of an individual with a high percentage of del(16)(q22)/fra(16)(q22) cells on her peripheral blood culture and her history of infertility. According to current evidence, infertility cannot be completely attributed to the mosaic fragile site in chromosome 16q22. However, the abnormal karyotype may increase the risk of chromosome structural variations and the production of unbalanced gametes. To enhance the chances of a successful pregnancy, assisted reproductive technology combined with preimplantation genetic testing may help.
Abbreviations
SCE, sister chromatid exchange; CNV, copy number variation; PGT, preimplantation genetic testing; MOF, monitor of follicle.
Ethical Approval
The study was approved by the ethics committee on ART of Dalian Municipal Women and Children’s Medical Center Group (No. 2023014).
Declaration of Patient Consent
Written informed consent was obtained from the patient for publication of this case report and any accompanying images.
Acknowledgments
We thank our patient for permitting us using her clinical data in this case report.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
PhD career start-up foundation of Dalian Women and Children’s Medical Group, Dalian, China, 2022-BSQD-0156.
Disclosure
The authors report no conflict of interest for this work.
References
1. Lukusa T, Fryns JP. Human chromosome fragility. Biochim Biophys Acta. 2008;1779(1):3–16. doi:10.1016/j.bbagrm.2007.10.005
2. Kumar R, Nagpal G, Kumar V, et al. HumCFS: a database of fragile sites in human chromosomes. BMC Genomics. 2019;19(Suppl 9):985–992. doi:10.1186/s12864-018-5330-5
3. Dekaban A. Persisting clone of cells with an abnormal chromosome in a woman previously irradiated. J Nucl Med. 1965;6(10):740–746. PMID:5837920.
4. Mulligan LM, Phillips MA, Forster-Gibson CJ, et al. Genetic mapping of DNA segments relative to the locus for the fragile-X syndrome at Xq27.3. Am J Hum Genet. 1985;37(3):463–472. PMID:2988332.
5. Goonewardena P, Gustavson KH, Holmgren G, et al. Analysis of fragile X-mental retardation families using flanking polymorphic DNA probes. Clin Genet. 1986;30(4):249–254. doi:10.1111/j.1399-0004.1986.tb00604.x
6. Zlotorynski E, Rahat A, Skaug J, et al. Molecular basis for expression of common and rare fragile sites. Mol Cell Biol. 2003;23(20):7143–7151. doi:10.1128/MCB.23.20.7143-7151.2003
7. Mirceta M, Shum N, Schmidt MHM, Pearson CE. Fragile sites, chromosomal lesions, tandem repeats, and disease. Front Genet. 2022;13:985975. doi:10.3389/fgene.2022.985975
8. Debacker K, Kooy RF. Fragile sites and human disease. Hum Mol Genet. 2007;16(2):150–158. doi:10.1093/hmg/ddm136
9. Yu S, Mangelsdorf M, Hewett D, et al. Human chromosomal fragile site FRA16B is an amplified AT-rich minisatellite repeat. Cell. 1997;88(3):367–374. doi:10.1016/s0092-8674(00)81875-9
10. Schmid M, Feichtinger W, Jessberger A, Köhler J, Lange R. The fragile site (16)(q22). I. Induction by AT-specific DNA-ligands and population frequency. Hum Genet. 1986;74(1):67–73. doi:10.1007/BF00278788
11. Takahashi E, Hori T, Murata M. Population cytogenetics of rare fragile sites in Japan. Hum Genet. 1988;78(2):121–126. doi:10.1007/BF00278179
12. Hocking T, Feichtinger W, Schmid M, et al. Homozygotes for FRA16B are normal. Chromosome Res. 1999;7(7):553–556. doi:10.1023/a:1009293613064
13. Glasser L, Meloni-Ehrig A, Joseph P, Mendiola J. Benign chronic neutropenia with abnormalities involving 16q22, affecting mother and daughter. Am J Hematol. 2006;81(4):262–270. doi:10.1002/ajh.20550
14. Murata M, Takahashi E, Ishihara T, et al. Heritable fragile sites and cancer: fra(16)(q22) in lymphocytes of an acute nonlymphocytic leukemia patient with inv(16)(p13q22). Cancer Genet Cytogenet. 1987;25(1):81–86. doi:10.1016/0165-4608(87)90162-2
15. García-Sagredo JM, San Román C, Gallego Gómez ME, Lledo G. Fragile chromosome 16(q22) cause a balanced translocation at the same point. Hum Genet. 1983;65(2):211–213. doi:10.1007/BF00286668
16. McGowan-Jordan J, Hastings RJ, Moore S. ISCN 2020: an international system for human cytogenomic nomenclature; 2020. Available from: https://www.karger.com/Book/Home/279152.
17. Wu K, Zhang X, Li F, et al. Frequent alterations in cytoskeleton remodelling genes in primary and metastatic lung adenocarcinomas. Nat Commun. 2015;6:10131. doi:10.1038/ncomms10131
18. Schmid M, Feichtinger W, Haaf T. The fragile site (16)(q22). II. Sister chromatid exchanges. Hum Genet. 1987;76(4):365–368. doi:10.1007/BF00272446
19. Morgan WF, Crossen PE. The frequency and distribution of sister chromatid exchanges in human chromosomes. Hum Genet. 1977;38(3):271–278. doi:10.1007/BF00402152
20. Tucker JD, Christensen ML, Strout CL, Carrano AV. Determination of the baseline sister chromatid exchange frequency in human and mouse peripheral lymphocytes using monoclonal antibodies and very low doses of bromodeoxyuridine. Cytogenet Cell Genet. 1986;43(1–2):38–42. doi:10.1159/000132295
21. Martorell MR, Martínez-Pasarell O, Lopez O, et al. Chromosome 16 abnormalities in embryos and in sperm from a male with a fragile site at 16q22.1. Cytogenet Genome Res. 2014;142(2):134–139. doi:10.1159/000357411
22. Luo K, Lan Y, Xie P, et al. Next-generation sequencing analysis of embryos from mosaic patients undergoing in vitro fertilization and preimplantation genetic testing. Fertil Steril. 2019;112(2):291–297. doi:10.1016/j.fertnstert.2019.03.035
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.