")
Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 17
Essential Oil of Matricaria chamomilla Alleviate Psoriatic-Like Skin Inflammation by Inhibiting PI3K/Akt/mTOR and p38MAPK Signaling Pathway
Authors Chen G , Lv C, Nie Q, Li X, Lv Y, Liao G, Liu S, Ge W , Chen J , Du Y
Received 9 November 2023
Accepted for publication 13 December 2023
Published 8 January 2024 Volume 2024:17 Pages 59—77
DOI https://doi.org/10.2147/CCID.S445008
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Anne-Claire Fougerousse
Guang Chen,1,* Caohua Lv,2,* Qing Nie,3 Xin Li,1 Yinyi Lv,1 Guoyan Liao,1 Shuangchun Liu,4 Weiwei Ge,2 Jinguang Chen,1 Yunting Du5
1Taizhou Central Hospital (Taizhou University Hospital), Taizhou University, Taizhou, 318000, People’s Republic of China; 2Department of Dermatology, Taizhou Second People’s Hospital, Taizhou, 317200, People’s Republic of China; 3Weifang Centers for Disease Control and Prevention, Weifang, 261061, People’s Republic of China; 4Municipal Hospital Affiliated to Medical School of Taizhou University, Taizhou, 318000, People’s Republic of China; 5Department of Laboratory Medicine, Cancer Hospital of China Medical University, Liaoning Cancer Hospital & Institute, Shenyang, 110042, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jinguang Chen; Yunting Du, Tel/Fax +86 576 88661988, Email [email protected]; [email protected]
Background: The traditional Matricaria chamomilla L. has been used to treat dermatitis for thousands of years. Due to emerging trends in alternative medicine, patients prefer natural remedies to relieve their symptoms. Therefore, finding safe and effective plant medicines for topical applications on the skin is an important treatment strategy for dermatologists. German chamomile (Matricaria chamomilla L.) from the Compositae family is a famous medicinal plant, often known as the “star of medicinal species.”However, the function of Matricaria chamomilla essential oil on skin inflammation has not been thoroughly examined in earlier research.
Methods: GC-MS analyzed the components of MCEO, and this study explored the anti-inflammation effects of MCEO on psoriasis with network pharmacological pathway prediction. Following this, we used clinical samples of psoriasis patients to confirm the secretory characteristic of relative inflammatory markers. The therapeutic effect of MCEO on skin inflammation was detected by examination of human keratinocytes HaCaT. At the same time, we prepared imiquimod-induced psoriatic-like skin inflammation in mice to investigate thoroughly the potential inhibition functions of MCEO on psoriatic skin injury and inflammation.
Results: MCEO significantly reduced interleukin-22/tumor necrosis factor α/lipopolysaccharide-stimulated elevation of HaCaT cell inflammation, which was correlated with downregulating PI3K/Akt/mTOR and p38MAPK pathways activation mediated by MCEO in HaCaT cells treated with IL-22/TNF-α/LPS. Skin inflammation was evaluated based on the PASI score, HE staining, and relative inflammatory cytokine levels. The results showed that MCEO could significantly contribute to inflammatory skin disease treatment.
Conclusion: MCEO inhibited inflammation in HaCaT keratinocytes induced by IL-22/TNF-α/LPS, the potential mechanisms associated with inhibiting excessive activation and crosstalk between PI3K/Akt/mTOR and p38MAPK pathways. MCEO ameliorated skin injury in IMQ-induced psoriatic-like skin inflammation of mice by downregulating the levels of inflammatory cytokines but not IL-17A. Thus, anti-inflammatory plant drugs with different targets with combined applications were a potential therapeutic strategy in psoriasis.
Keywords: Matricaria chamomilla, essential oil, psoriasis, HaCaT, IMQ, mouse
Introduction
Psoriasis is a chronic immune-mediated disease influencing 100 million individuals worldwide. Hence, action is needed to provide care for patients with psoriasis and eliminate discrimination.1 It is associated with multiple comorbidities and seriously affects patients’ quality of life. It is mainly manifested as epidermal hyperplasia, abnormal keratinocyte differentiation, marked Artery dilatation, and marked seep inflammatory cells around dermal capillaries in psoriatic histomorphology. The pathogenesis of psoriasis is complex and remains unelucidated. Stimulation of keratinocytes has been considered to lead to the generation of secondary mediums, for example, cell factors, chemokines, and antibacterial peptides. Thus, psoriasis can be induced by the dysregulation of immune and keratinocyte differentiation accompanied by persistent inflammation.2,3 At present, in addition to traditional immunosuppressants, for example, methotrexate (MTX) and cyclosporine, new therapeutic advances, including selective inhibitors, such as Phosphodiesterase 4 (PDE4) inhibitor, Janus kinase (JAK) inhibitor, and mAbs, anti-cytokine/T cell therapy, and relative gene therapy, was applied to clinical psoriasis.3,4 However, with the increase in the frequency of clinical use of biological agents for patients with prolonged drug use, the clinical effects of biological agents may be reduced. Furthermore, various adverse reactions limit their clinical application.5 Also, as the cost of psoriasis treatment is a major challenge, we require efficient, safe, and cost-effective therapies.
Despite advances in drug application, natural herbal alternative therapy has become a key factor for skin disease therapy due to the increasing demand for herbs, associated low costs, and the continuing adverse effects of modern medicines.6 Moreover, herbal remedies, especially those derived from plants, are effective against various skin conditions.7 Some investigations have found that natural plant extracts from Aloe vera, Mahonia aquifolium, Matricaria recutita, Silybum marianum, Gaultheria procumbens, special populations are effective against a variety of skin conditions.7
German chamomile (Matricaria chamomilla L.) from the Compositae family is a famous medicinal plant called “a star of medicinal species.” These herbs have been used in Greece, Rome, and ancient Egypt for thousands of years. Dai et al found that there are many kind of chemical constituents in German chamomile, including organic acids, flavonoids, coumarins, volatile oil, monoterpenes, sesquiterpenes, diterpenes and triterpenes ect.8 German chamomile has definite therapeutic effects on treating stomach problems, cramps, dermatitis, minor infections,8–12 bactericidal and muscle-relaxant activities. In addition, some animal studies have certified anti-anxiety,13 anti-cancer, cholesterol-lowering, wound-healing,14 anti-diabetic properties15 of this plant. However, its application in the field of inflammatory skin diseases is just emerging. Lin et al showed that besides anti-inflammatory and antioxidant, M. chamomilla essential oil (MCEO) can promote wound healing and repair the skin barrier.9 Moreover, Wang et al found that MCEO decreased the proinflammatory factors (TNF-α and IL-6) to inhibit the Th17 cell differentiation during eczema.16 In China and Japan, it was usually used externally in the skin field, including anti-itch and anti-inflammatory;17 Sun protection,18 relieves skin irritation19 and antiviral or prevents viral diseases.20 To date, no experimental reports on the application of German chamomile in psoriasis have been published.
Network pharmacology is a new strategy to systematically study the relationship between diseases, drugs, and targets.21 A growing body of evidence supports network pharmacology as an effective tool for studying the molecular mechanisms of herbs containing complex multi-compounds.22 The pharmacological effects and potential targets of MCEO on psoriasis were systematically discussed utilizing network pharmacology. The anti-inflammatory effect of MCEO on psoriatic-like skin inflammation was validated in vivo and in vitro based on network pharmacological prediction. In addition, Luminex evaluated the levels of relative inflammatory cytokines in psoriatic patients to identify potential targets for the anti-inflammatory effects of MCEO in humans. According to the analysis results of network pharmacology, we evaluated the anti-inflammation effects and regulating mechanism of MCEO in interleukin-22 (IL-22)/tumor necrosis factor α (TNF-α)/lipopolysaccharide (LPS)-stimulated human keratinocytes (HaCaT) in vitro and in the rodent imiquimod (IMQ)-induced psoriatic skin inflammation model mice, to clarify the potential value of MCEO in clinical application of inflammatory skin diseases from the perspective of anti-inflammatory.
Materials and Methods
MCEO Extraction and GC-MS Analysis
MCEO was obtained by steam distillation of flowers from Matricaria chamomilla. Qualitative analyses of MCEO were carried out by Gas Chromatography(GC)-mass spectrometry (MS) using a 7890A-5975C type GC-MS (Agilent Technology Co., LTD, California, USA). Chromatographic conditions: HP-5MS quartz capillary column (30m×0.25mm×0.25μm), inlet temperature 250°C; The carrier gas was helium (99.999%), the shunt ratio was 10:1, and the sample size was 1μL. Mass spectrometric conditions: electron bombardment ion source (EI), ion source temperature 230°C, interface temperature 280°C; The electron energy is 70Ev, the four-pole temperature is 150°C, and the mass scanning range is 20–650Amu. Heating procedure: the initial temperature is 60°C; keep it for 5min, and rise to 280°C at the rate of 5°C/min. Hold for 10 minutes.
Analysis of Network Pharmacology
The MCEO network was constructed with MCEO compounds. According to the results of GC-MS analysis, Genes related to MCEO compounds were searched with PubChem (https://pubchem.ncbi.nlm.nih.gov/) and target genes using a compound database. Psoriasis-related genes were gathered using GeneCards (http://www.genecards.org/) with the keyword “psoriasis.” Common genes of the MCEO network and psoriasis-related gene set were organized. A protein-protein interaction network (PPIN) was constructed with target genes between MCEO and psoriasis with the STRING database (https://string-db.org/) and then visualized by Cytoscape (3.9.1). DAVID (https://david.ncifcrf.gov/) database was used to perform Gene Ontology (GO) annotation analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of the targets. Psoriasis-related KEGG pathways and GO process were organized, and cut-off criteria were p-value < 0.05.
Patient
Patient Recruitment and Samples
A total of 31 psoriasis vulgaris patients, whose diagnoses were based on the Guideline for the diagnosis and treatment of psoriasis in China (2021 edition) and has not been treated with biological agents, were recruited from Taizhou Second People’s hospital (Zhejiang, China) from February 2021 to December 2021, along with 35 presenting non-inflammatory skin conditions as controls (Supplement Table 1).
Research work was approved by the Ethics Review Committee on Human Research of Taizhou Second People’s Hospital (tzey-ky202101) and executed based on the Declaration of Helsinki Principles. Taizhou Second People’s Hospital (Taizhou) recruited all participants in this study. All the patients signed informed consent forms.
Sterile venous blood was obtained by standard venepuncture and stored overnight in a 4°C refrigerator. The next day, we separated serum by centrifuging the blood samples at 1500g for 15 min at 4°C. All serum samples were kept frozen at −80 °C until assay.
Inflammatory Cytokines Assay by Luminex in Clinical Sample
Serum multiple cytokine analysis was executed using Bio-Plex Pro Human Cytokine 27-plex (M500KCAF0Y, Bio-rad systems), measure the concentrations of Interleukin (IL)-1β, IL-10, IL-17A, IFN-γ and Tumor Necrosis Factor-alfa (TNF-α) in one sample at the same time, follow the manufacturer’s instructions.
Chemicals, Antibodies, and Reagents
The following materials were used in the present study: MCEO (Jiaxing Hanjiangfangcao Bioengineering Co. Ltd., Zhangjiang, China); DMSO (20220813); FBS (10099-141C, Gibco, Australia); DMEM high glucose medium (Hyclone, USA); Penicillin-Streptomycin (Gibco, Australia); 0.25% trypsin-EDTA (Gibco, Australia); recombinant human TNF-α (R&D system, America); recombinant human IL-22 (R&D system, America); LPS (L2630, Sigma-Aldrich, Merck); Cell Counting Kit-8 (MedChemExpress, America); propidium iodide (BIO BASIC INC, Canada); PMSF, RIPAlysis buffer, BCA protein assay kit, BeyoECL Star, primary antibody dilution buffer, hematoxylin and eosin staining kit (Beyotime, China); multicolor prestained protein ladder (Shanghai Epizyme Biomedical Technology, China); Antibodies for Phospho-mTOR (Ser2448)(D9C2) Rabbit mAb #2475, mTOR (7C10) Rabbit mAb #2983, Phospho-p38 MAPK (Thr180/Tyr182) (D3F9) Rabbit mAb #4511, p38 MAPK (D13E1) Rabbit mAb #8690, Phospho-Akt (Ser473) (D9E) Rabbit mAb #4060, Akt (pan) (C67E7) Rabbit mAb #4691, Phospho-p44/42 MAPK (Erk1/2) (Thr202/Tyr204) (D13.14.4E) Rabbit mAb #4370, p44/42 MAPK (Erk1/2) (137F5) Rabbit mAb #4695, Bcl-2 (124) Mouse mAb #15071, Caspase-3 Antibody #9662, β-Actin (D6A8) Rabbit mAb (HRP Conjugate) #12620, Anti-mouse IgG, HRP-linked Antibody #7076, Anti-rabbit IgG, and HRP-linked Antibody #7074 (Cell Signaling Technology, Danvers, MA, USA); Caspase-8 Antibody, Caspase-9 Antibody, Bax Antibody, Goat anti-Mouse IgG HRP (TS0002), and Goat anti-Rabbit IgG HRP (TS0001)(Abmart Technology, Shanghai, China); Anti-PI 3 Kinase p85 alpha (phospho Y607)(ab182651), Anti-PI 3 Kinase p85 alpha antibody [6G10] (ab189403)(Abcam TechnologyCambridge, England); SAv HRP (Streptavidin-Horseradish Peroxidase Pre-diluted) (51–75477E) (BD Pharmingen); PrimeScript RT Reagent Kit (Perfect Real time, TaKaRa, Japan); SYBR Green Realtime PCR MasterMix (TOYOBO, Japan); Mouse ACTB Endogenous Reference Genes Primers (B661302), and Human ACTB Endogenous Reference Genes Primers (B661102) (Sangon Biotech, China); DEPC treated Water and TRIzol Reagent (Ambion, America).
Cell Experiment
General Cell Culture and Treatment
We obtained immortalized human HaCaT keratinocytes from Wuhan University Cell Bank (Wuhan, China). Dulbecco’s modified Eagle’s medium (DMEM), added into 10% fetal bovine serum (FBS), 100 μg/mL of streptomycin, and 100 U/mL of penicillin, were used for cell culture. Cells were incubated at 37°C with 5% CO2. We used rhIL-22 (20ng/mL), rhTNF-α (10ng/mL), or LPS (10μg/mL) to treat HaCaT cells. We prepared the stock solution of MCEO in dimethyl sulfoxide (DMSO, 0.1%v/v) and, according to experimental design, diluted the differential concentration in the medium for dealing with HaCaT cells. Control cells were treated with the same amount of DMEM (DMSO 0.1%v/v); this concentration of DMSO did not affect the cell viability.
For rhIL-22/rhTNF-α/LPS with or without MCEO treatment scheme, HaCaT cells were divided into normal control group, rhIL-22 (24h, 20ng/mL)/rhTNF-α (24h, 10ng/mL)/LPS (1h, 10 µg/mL) treatment group, and pre-MCEO (24h)+rhIL-22 (24h)/TNF-α (24h)/LPS (1h) treatment group. First, MCEO treated HaCaT cells for 24h, and then PBS washed the cells twice. Subsequently, rhIL-22 (20ng/mL, 24h), rhTNF-α (10ng/mL, 24h), or LPS (10μg/mL, 1h) were added, respectively. Cells were harvested and lysed for Western blot, and qRT-PCR was performed as described below.
Cell Viability Assay
CCK-8 method was adopted to calculate the cell survival rate. HaCaT cells were grown in 96-well plates for 24h or 48h. Cells were harvested after they reached 50–60% confluence and were treated with MCEO for 24h and 48h. CCK-8 solution (10 μL) was added into each well and then incubated at 37 °C with 5% CO2 for 2h. We used Multiskan Spectrum (Thermo Fisher Scientific, USA) to measure absorbance at 450 nm.
Flow Cytometric Analysis
Cells (5×105/plate) were seeded and attached in 60 mm culture dishes for 24h. The cells were harvested using trypsin after MCEO treatment for 24h. We used PBS to wash the collected cells and ethanol to fix them at 4 °C for 2h. We incubated fixed cells with RNase (10μg/mL) at 37 °C for 30min. Subsequently, PBS washed the fixed cells twice and then stained them with propidium iodide (PI, 10μg/mL) for 30 min in the dark. The cell cycle was monitored using CytoFLEX (BECKMAN CULTER, CA, USA).
Animals
Female BALB/c mice (eight-to-nine-week-old; raised under specific pathogen-free (SPF) conditions) were purchased from Shanghai Vital River Laboratory Animal Technology Co., Ltd. (Shanghai, China). Mice were bred under constant temperature and humidity conditions with a 12-h light/dark cycle. Food and water are freely afforded. All experiment schemes were agreed upon by the Ethical Committees of Taizhou University (Taizhou, China) (TZXY-2022-20221027). All animal procedures were conducted in compliance with the Regulations for the Administration of Affairs Concerning Experimental Animals (1988.11.1), and humanely treated. Animals were anesthetized by isoflurane (Riward Life Science Co., Ltd, Shenzhen, China)(3% for induction; 1–3% for installation) in a mixture of 0.25% air and 0.5% O2.
BALB/c mice were divided into three groups (n=6) randomly: Olive oil group (Con), IMQ group (IMQ), and IMQ+MCEO group (IMQ+MCEO). MCEO (1:10 dilution of olive oil) was applied on the bare back for seven days in a row (twice a day, once in the morning and once in the afternoon; 200μL/time/mice). Subsequently, mice were administered a daily 62.5 mg of commercially available IMQ cream (5%) (Sichuan Mingxin Pharmaceutical Co. LTD, H20030128) on the bare back for seven days in a row, concurrently with MCEO treatment as mentioned above. The control group and IMQ group were given similarly with a control cream (olive oil; fagron). On day eight, all mice were euthanized. Serum samples were collected, and the back skin was obtained for subsequent detection.
Measurement of Indicators
Scoring Severity of Skin Inflammation
Skin lesions were evaluated by Psoriasis Area and Severity Index (PASI). We calculated the overall score based on erythema, scaling, and thickening but excluding the affected skin area. According to the severity of each item, scored independently (from 0 to 4): 0, none; 1, mild; 2, general; 3, typical; 4, very specific. Inflammation severity was determined by cumulative score (erythema, scaling, and thickening) (scale 0–12).
Histological Examination
We fixed the skin tissues using a 4% paraformaldehyde solution and embedded skin tissues in paraffin. After sectioning the paraffin-embedded tissues, hematoxylin and eosin (H&E) stain was performed for histological evaluation. An Olympus microscope under a 100×light microscope got pictures, and epidermal thickness was calculated by the Image J software (NIH Image, USA).
Quantitative Real-Time PCR Assay (qRT-PCR)
The mRNA expressions of IL-6, IL-1β, TGF-β, IL-10, TNF-α, IL-17A, and IL-22 in HaCaT cells and skin sections were detected using the qRT-PCR method. We used TRIzol®reagent to extract RNA and a reverse transcription kit (Takara Bio, Inc.) to synthesize cDNA. qPCR was operated using SYBR premixed EX TaqTMcDNA and specific gene primers. We used the LightCycler480 system (Roche) to amplify target genes. We used the standardized ACTIN gene to determine the relative expression quantity of each gene, which was calculated via the 2−ΔΔCq method. The following primers were used: Human IL-6-forward, 5’-GACAGCCACTCACCT CTTCAGAAC’ and reverse, 5’-CCAGGCAAGTCTCCTCATTGAATCC’; Human IL-1β-forward, 5’-TACGAATCTCCGACCACCACTACAG’and reverse, 5’-ACACCACTTGTTGCTCCATATC CTG-3’; Human TGF-β-forward, 5’-ACTACTACGCCAAGGAGGTCACC’ and reverse, 5’-TG AGGTATCGCCAGGAATTGTTGC-3’; Human IL-10-forward, 5’-GGACAACATACTGCTAAC CGACTCC-3’ and reverse, 5’-CTTCACCTGCTCCACTGCCTTG-3’; Human IL-17A-forward, 5’-GGTGTCACTGCTACTGCTGCTG-3’ and reverse, 5’-GGTCCTCATTGCGGTGGAGATTC −3’; Human ATCB-forward, 5’-GTGCTATGT TGCTCTAGACTTCG-3’ and reverse, 5’-ATGCC ACAGGATTCCATACC-3’; Mouse IL-6-forward, 5’-CTTCTTGGGACTGATGCTGGTGAC-3’ and reverse, 5’-CTCTCTGAAGGACTCTGGCTTTGTC-3’; Mouse IL-1β-forward, 5’-TTCAGG CAGGCAGTATCACTCATTG-3’and reverse, 5’-TGTCGTTGCTTGGTTCTCCTTGTAC-3’; Mouse TGF-β-forward, 5’-ACCGCAACAACGCCATCTATGAG-3’ and reverse, 5’-AGCCCTG TATTCCGTCTCCTTGG-3’; Mouse IL-10-forward, 5’-GGACAACATACTGCTAACCGACTC C-3’ and reverse, 5’-CTTCACCTGCTCCACTGCCTTG-3’; Mouse TNF-α-forward, 5’-CACCA CGCTCTTCTGTCTACTGAAC-3’ and reverse, 5’-TGACGGCAGAGAGGAGGTTGAC-3’; Mouse IL-17A -forward, 5’-TGATGCTGTTGCTGCTGCTGAG-3’ and reverse, 5’-GGTCTTCA TTGCGGTGGAGAGTC-3’; Mouse ATCB-forward, 5’-CCTGGCACCCAGCACAAT-3’, and reverse, 5’-GGGCCGGACTGCTCATAC-3’
Western Blot Analysis
Proteins were extracted from HaCaT cells, then after 8 or 10% SDS-PAGE resolved, transferred to PVDF membranes (EMD Millipore). After blocking membranes using 5% nonfat dry milk, was incubated overnight with primary antibodies against Bax, Caspase 8, Caspase 9 (1:1000; Abmart); Caspase 3, Bcl-2, p-mTOR/mTOR, p-P38/P38, β-actin, p-P44/42/P44/42, and GAPDH (1:1000; CST); p-PI3K/PI3K (1:500; Abcam). We used HRP-conjugated goat anti-rabbit IgG or goat anti-mouse IgG (1:8000; Abmart Biological Reagents Co., Ltd.) as the secondary antibodies to label the antibodies above.
Immunofluorescence (IF) Studies
HaCaT cells (2×104/well) were inoculated into a 4-well glass plate and subjected to signal-direct IF staining of p-Akt/Akt (1:200), p-P38/P38 (1:200), and p-mTOR/mTOR (1:100), followed by incubation with Alexa Fluor 488-conjugated secondary antibodies (OriGene Technologies, Inc.). DAPI (Beyotime Biotechnology, Inc.) was used to counterstain nuclei. We used a laser scanning confocal microscope (IX-83 FV1000; Olympus Corporation) to check the staining section and save all images.
Inflammatory Cytokines Assay by Luminex in Psoriatic-Like Skin Inflammation Mice
Blood samples were collected from Con, IMQ, and IMQ+MCEO group mice. Separated serum was stored at −80°C. Bio-Plex Pro Mouse Cytokine 1 6plx EXP (17007935, Bio-rad systems) was used to analyze the concentrations of IL-1β, IFN-γ, IL-10, IL-17A, and TNF-α in one sample at the same time, follow the manufacturer’s instructions.
Statistical Analysis
We analyzed all data using GraphPad Prism version 8.0 (GraphPad Software, Inc. CA, USA). All indicators are represented as the mean ± standard deviation, Using a one-way ANOVA or a two-tailed unpaired Student’s t-test to calculate the difference. P-values were adjusted for multiple comparisons using Bonferroni correction. Values with P<0.05 were defined as statistically significant.
Results
Analysis of Matricaria Chamomilla Essential Oil Composition
The components of Matricaria chamomilla essential oil were analyzed by GC-MS method, which mainly included 15 components (Table 1). Among them, Azulene accounted for the largest amount, about 88.9%.
 |
Table 1 Composition of Matricaria chamomilla Essential Oil |
Analysis of Network Pharmacology
To explore the relationship between the active component of MCEO and psoriasis, the 12 components of MCEO were analyzed with PubChem. The intersection genes corresponding to the targets of MCEO and psoriasis were retrieved using the GeneCards database. The “Drug-target-disease” protein-protein interaction network (PPIN) was built using the STRING database. GO and KEGG enrichment analyses of common genes between MCEO and psoriasis were performed for further study. As shown in Figure 1A, 130 terms about MCEO and psoriasis were included, and PPIN was obtained using a common gene between MCEO and psoriasis (Figure 1B). According to the GO analysis of molecular function (Figure 1C), the active targets are connected to inflammation response and signal transduction. KEGG analysis included the PI3K/Akt signaling pathway, MAPK signaling pathway, and Th17 cell differentiation (Figure 1D). These results guided further inflammation response and signaling pathway regulation assay.
 |
Figure 1 Analysis of network pharmacology. (A) Venn diagram of intersection genes between MCEO and psoriasis; (B) PPIN was constructed with target genes between MCEO and psoriasis; (C) GO analysis of intersection genes between MCEO and psoriasis; (D) KEGG analysis of intersection genes between MCEO and psoriasis. |
Analysis of Inflammatory Phenotype in Psoriatic Patients
To clarify the expression characteristic of inflammatory cytokines in psoriatic patients, we collected the blood samples of 35 healthy and 31 psoriasis vulgaris patients and detected levels of IL-1β, TNF-α, IFN-γ, IL-10, and IL-17A in the peripheral blood by Luminex (Supplement Table 1). Our results found that the levels of relative inflammatory factors appeared to increase in the peripheral blood of patients, except for IL-10 (Supplement Table 1), which grew gradually with the prolongation of the course of psoriasis, except for IL-10 (Supplement Figure 1).
Drug-Regulated Verification in vitro
Effect of MCEO on the Viability of HaCaT Cells
The viability of MCEO-treated HaCaT cells for 24h and 48h was tested using the CCK-8 assay. MCEO showed no obvious toxicity upon treatment, with concentrations ranging from 30–60 μg/mL for 24h and 48h. A marked reduction in the activity of HaCaT cells was discovered upon treatment of cells with MCEO at concentrations above 63.18 µg/mL for 24h and 82.66µg/mL for 48h (Figure 2A and B).
 |
Figure 2 Treatment with MCEO had no significant effect on the growth and viability of HaCaT keratinocytes. Relative numbers of viable HaCaT treated with or without MCEO (30–80 mg/mL) for 24 h (A) and 48 h (B) as determined by the CCK8 assay. Mean percentage viability of HaCaT plotted against the indicated doses of MCEO. Experiments were performed thrice with each concentration in triplicate wells, and the IC50 values calculated from these plots are shown. Abbreviations: MCEO, Matricaria chamomilla essential oil; NC, normal control; IC50, half-maximal inhibitory concentration. |
Effect of MCEO on Cell Cycle and Apoptosis in HaCaT
To explore whether the effect of MCEO on cell viability was correlated with cell cycle arrest or apoptosis, we used flow cytometric analysis to measure the percentage of cells containing diploid (sub-G1) levels of DNA by PI staining. We used different concentrations of MCEO (30, 50, 60, and 70 µg/mL) to treat HaCaT. Treatment with 50 µg/mL and 60 µg/mL of MCEO increased the proportion of HaCaT in the G1 phase (60.4±2.46% and 67.0±5.58%, respectively) compared with normal control (0 µg/mL) (41.3±2.7%)(P<0.01) (Figure 3A). Western blot (WB) analysis revealed that at a concentration of 60 µg/mL, MCEO treatment for 24h induced expression of caspase 9 apoptosis protein in HaCaT cells. At a 50µg/mL concentration, MCOE treatment for 24h did not induce apoptosis in HaCaT, as evidenced by the expression of Caspases 3, 8, and 9, and Bax protein levels (Figure 3B).
 |
Figure 3 Effects of MCEO on the percentage of cell population in different phases of cell cycle and the expression of apoptosis markers. (A) Histogram and column diagram show levels or percentages of distribution of MCEO-treated cells based on cell cycle, as assessed by flow cytometry analysis. (B) Expression levels of caspase 3, caspase 8, caspase 9, and Bax proteins, as assessed by Western blotting. Equal protein loading was confirmed using β-actin as the loading control.Bar graphs represent the mean ± SD of results obtained from three independent experiments performed in triplicate. *P<0.05 **P<0.01 vs.NC. Abbreviations: MCEO, Matricaria chamomilla essential oil; NC, normal control. |
MCEO Regulates IL-22-Induced HaCaT Proliferation and Inflammation by Inhibiting the PI3K/Akt/mTOR Pathway
In recent years, the immunological and pathological mechanisms have been clarified, which Th17 cells produce various cytokines, such as IL-17 and IL-22, which can induce proliferation, migration, or invasion of keratinocytes, release a potential ability in the pathogenesis of psoriasis.23–25 IL-22 can also activate the Akt/mTOR pathway.26 The PI3K/Akt/mTOR pathway up-regulates Th17 differentiation by promoting the nuclear translocation of RORγt.27 Hence, we measured the expression of IL-6, IL-1β, IL-17A, IL-10, and TGF-β mRNAs by qRT-PCR (Figure 4B, some of the unchanged data were not shown). MCEO treatment decreased the levels of IL-6 and IL-1β mRNAs in HaCaT cells compared to those observed in the IL-22 treatment group, as shown by qRT-PCR (P<0.01; P<0.01). However, MCEO treatment did not change the expression of IL-17A mRNA in HaCaT. To clarify the anti-inflammation effect of MCEO exerted, the expression of PI3K, Akt, and mTOR proteins was detected by WB and IF in HaCaT treated with IL-22 (20ng/mL) for 24h after pretreatment with MCEO 24h (Figure 4A and C). Data showed that MCEO treatment significantly suppressed IL-22-mediated increase in the expression of p-Akt (at Ser473) and p-mTOR (at Ser2448) proteins compared with the control group, as assessed by WB and IF (P<0.05; P<0.01).
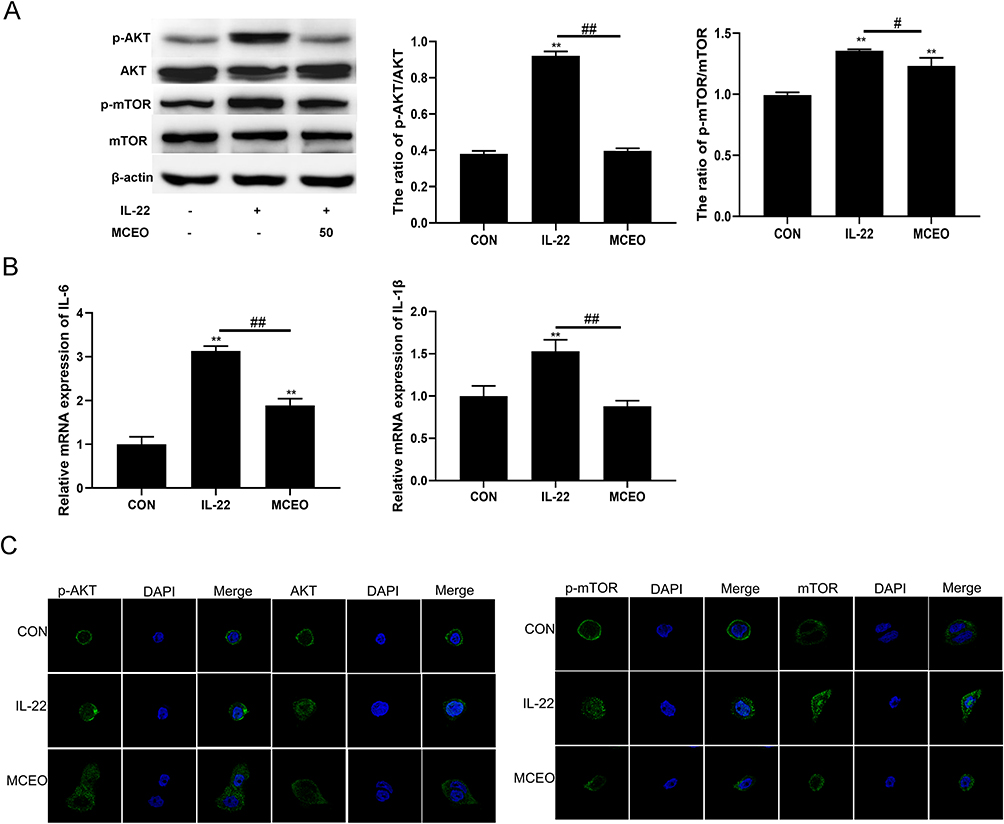 |
Figure 4 MCEO regulates IL-22-induced HaCaT proliferation by inhibiting the PI3K/Akt/mTOR pathway. (A) HaCaT cells treated with or without IL-22 and MCEO were subjected to Western blotting using specific antibodies against p-Akt, Akt, p-mTOR, and mTOR. (B) The mRNA expression levels of IL-6 and IL-1β were analyzed by qRT-PCR. (C) HaCaT cells treated with or without IL-22 and MCEO were stained with p-Akt, Akt, p-mTOR, and mTOR and assessed using immunofluorescence. p-Akt, Akt, p-mTOR, and mTOR (green), nuclei (blue). Magnification, ×2070. Bar graphs represent mean ± SD of results derived from three independent experiments performed in triplicate. **P<0.01 vs NC; #P<0.05 and ##P<0.01 vs IL-22 group. Abbreviations: MCEO, Matricaria chamomilla essential oil; NC, normal control; IL-22, interleukin-22. |
MCEO Regulates TNF-α-Induced Inflammation by Inhibiting the PI3K/Akt/mTOR and p38MAPK Signaling Pathway
TNF-α is considered to be a key cytokine mediating skin inflammation during the development process of psoriasis.28 It can accelerate lymphocyte, monocyte, and neutrophil infiltration, activated by dendritic cells and transferred from the peripheral blood into the skin.23 This is related to the activation of Akt/mTOR and p38 MAPK pathways.26,29,30 HaCaT cells were pretreated with 50µg/mL of MCEO for 24h before stimulation with 10ng/mL of rhTNF-α. We measured the expression of IL-6, IL-1β, IL-17A, IL-10, and TGF-β mRNAs by qRT-PCR (Figure 5B, some of the unchanged data were not shown). Simultaneously, the activation of PI3K/Akt/mTOR and p38MAPK pathway was detected by WB and IF (Figure 5A and C). Our results clarified that TNF-α treatment increased the expression of IL-1β and TGF-β mRNAs and p-Akt, p-mTOR, and p-P38 proteins compared with the control group (P<0.05; P<0.01). However, pretreatment with MCEO for 24h markedly inhibited these TNF-α-mediated effects in HaCaT. The results clarified that MCEO possesses an anti-inflammation-specific impact, which is correlated with the downregulation of Akt/mTOR and p38MAPK pathways activation.
 |
Figure 5 MCEO regulates TNF-α-induced inflammation by inhibiting the PI3K/Akt/mTOR andp38MAPK signaling pathways. (A) HaCaT cells treated with or without TNF-α and MCEO were subjected to Western blotting using specific antibodies against p-P38, P38, p-mTOR, and mTOR. (B) The mRNA expression levels of IL-6, IL-1β, IL-17A, IL-10, and TGF-β were analyzed by qRT-PCR. (C) HaCaT cells treated with or without TNF-α and MCEO were stained with p-Akt, Akt, p-mTOR and mTOR, and immunofluorescence was assessed. p-Akt, Akt, p-mTOR, mTOR (green), nuclei (blue). Magnification, ×2070. Bar graphs represent mean ± SD of results derived from three independent experiments performed in triplicate. **P<0.01 vs NC; #P<0.05 and ##P<0.01 vs TNF-α group. Abbreviations: MCEO, Matricaria chamomilla essential oil; NC, normal control; TNF-α, tumor necrosis factor-α. |
MCEO Regulates LPS-Induced Inflammation by Inhibiting the PI3K/Akt/mTOR and p38MAPK Signaling Pathways
Cell cultures stimulated with LPS are routinely used to elucidate mechanisms of molecular immunity, and a model to assess the anti-inflammatory properties of new agents has been developed.31–33 In our study, we used LPS (10μg/mL, 1h) to stimulate HaCaT cells pretreated with 50µg/mL of MCEO for 24h. We then detected the expression of IL-6, IL-1β, IL-17A, IL-10, and TGF-β mRNAs by qRT-PCR (Figure 6B, some of the unchanged data were not shown). At the same time, the activity of PI3K/Akt/mTOR and p38MAPK pathways was monitored by WB and IF (Figure 6A and C). Our results clarified that LPS treatment increased IL-6 and IL-1β mRNA levels and p-Akt, p-mTOR, and p-P38 proteins compared to those observed with normal control (P<0.01). However, these LPS-induced effects were markedly suppressed upon pretreatment with MCEO for 24h. The results clarified that MCEO possesses an anti-inflammation-specific impact, which is correlated with the downregulation of Akt/mTOR and p38MAPK pathways activity.
 |
Figure 6 MCEO regulates LPS-induced inflammation by inhibiting the PI3K/Akt/mTOR andp38MAPK signaling pathways. (A) HaCaT cells treated with or without LPSand MCEO were subjected to Western blotting using specific antibodies against p-Akt, Akt, p-P38, P38, p-mTOR, and mTOR. (B) mRNA expression levels of IL-6, IL-1β, IL-17A, IL-10, and TGF-β were analyzed by qRT-PCR. (C) HaCaT cells treated with or without LPS and MCEO were stained with p-P38, P38, p-mTOR, and mTOR, and immunofluorescence was assessed; p-Akt, Akt, p-mTOR, mTOR (green), nuclei (blue). Magnification, ×2070. Bar graphs represent mean ± SD of results derived from three independent experiments performed in triplicate. **P<0.01 vs NC; ##P<0.01 vs TNF-α group. Abbreviations: MCEO, Matricaria chamomilla essential oil; NC, normal control; LPS, lipopolysaccharide. |
Drug-Regulated Verification in vivo
MCEO Ameliorates IMQ-Induced Psoriatic-Like Skin Inflammation in Mice
To ascertain the function of MCEO on skin inflammation, we utilized an IMQ-induced psoriatic-like skin inflammation model in BALB/c mice, which were treated with or without MCEO (Figure 7A). Compared with the IMQ group, MCEO (16mg/time/mice) treatment alleviated the clinical symptoms of IMQ-induced psoriasiform dermatitis, including erythema, thickening, and scaling (Figure 7B). HE staining showed that MCEO treatment significantly ameliorated the histological changes mediated by IMQ compared to those observed in the IMQ group not treated with MCEO (Figure 7D). PASI scores in the IMQ+MCEO group were consistently lower than in the IMQ group (Figure 7C), while the IMQ group showed higher epidermal thickening than that observed in the IMQ+MCEO group (Figure 7E).
 |
Figure 7 MCEO ameliorates psoriatic-like skin lesion in IMQ-induced mice. (A) Mice were pretreated with MCEO daily for seven consecutive days, followed by treatment with MCEO +IMQ daily for seven consecutive days and observation on day eight. (B) Photos of the dorsal skin of mice were captured on the eighth day after daily administration of IMQ+MCEO. (C) The cumulative PASI scores of all groups were also calculated daily. (D) H&E staining of the dorsal skin (magnification: 100×, 400×), the distance between the red arrows represents the acanthoderm thickness and (E) epidermal thickness of the dorsal skin on day eight (scale bar: 100 μm). Data are presented as mean ± SD (n=6), **P<0.01 and ***P<0.001 vs Psoriasis model group. ##P<0.01 vs IMQ+MCEO group. One representative of three separate experiments is shown. Abbreviations: MCEO, Matricaria chamomilla essential oil; NC, normal control. |
MCEO Regulates IMQ-Induced Skin Inflammation by Inhibiting the PI3K/Akt/mTOR and p38MAPK Signaling Pathways in Mice
Comparative research clarified that activation of PI3K/Akt/mTOR and p38MAPK signaling pathways correlated with skin inflammation. In this study, we detected the activation of PI3K/Akt/mTOR and p38MAPK pathway was detected by WB (Figure 8A), and the levels of IL-6, IL-1β, TNF-α, IL-17A, IL-10, and TGF-β mRNA was analyzed by qRT-PCR in the skin tissues of BALB/c mice (Figure 8B, some of the unchanged data were not shown). To further confirm the levels of inflammatory cytokines in IMQ-induced psoriatic-like dermatitis mice, IL-1β, TNF-α, IFN-γ, IL-17A, and IL-10 were detected by Luminex in the serum of BALB/c mice (Figure 8C). Our data show that MCEO pretreatment downregulated the expressions of p-PI3K/PI3K, p-mTOR/mTOR, p-P38/P38, and p-ERK1/2/ERK1/2 protein in skin inflammation mice. At the same time, we found that the expression of IL-6, IL-1β, TNF-α, IL-10, and TGF-β mRNAs in skin tissue and levels of IL-1β, TNF-α, and IFN-γ in serum were decreased by MCEO pretreatment. These results imply that MCEO ameliorates IMQ-induced psoriatic-like skin inflammation in mice, which is associated with inhibiting the activation of PI3K/Akt/mTOR and p38MAPK signaling pathways.
 |
Figure 8 Mechanism of MCEO in ameliorating psoriatic-like skin inflammation in IMQ-induced mice. (A) The expression levels of p-PI3K/PI3K, p-mTOR/mTOR, and p-P38/P38 in the skin of mice with psoriasis were analyzed eight days after the treatment with MCEO by WB. (B) The mRNA expression levels of IL-6, IL-1β, IL-17A, IL-10, and TGF-β in the skin of mice with psoriasis were analyzed eight days after the treatment with MCEO by qRT-PCR. (C) The levels of IL-1β, IFN-γ, TNF-α, IL-10 and IL-17A in serum of mice with psoriasis were analyzed eight days after the treatment with MCEO by Luminex. Data are presented as mean ± SD (n=6), **P<0.01 and *P<0.05 vs Psoriasis model group. #P<0.05 and ##P<0.01 vs IMQ+MCEO group. One representative of three separate experiments is shown. Abbreviations: MCEO, Matricaria chamomilla essential oil; NC, normal control. |
Discussion
In this research, we first analyzed the components of MCEO by GC-MS; the intersection genes corresponding to the targets of MCEO and psoriasis were retrieved using the GeneCards database. GO and KEGG enrichment analyses of MCEO to psoriasis found that the active targets essentially connected to inflammation response and signal transduction, closely correlated with the PI3K/Akt signaling pathway, MAPK signaling pathway, and Th17 cell differentiation (Figure 1). These results guided further cytokines and pathway regulation assay. Following this, we evaluated the levels of relative inflammatory cytokines in psoriatic patients to define potential targets for the anti-inflammatory effects of MCEO in humans. We verified for the first time the anti-inflammation effect of MCEO in vitro and vivo experiments according to the predictive results of network pharmacology. Data showed that MCEO pretreatment inhibited inflammation of the HaCaT human keratinocytes induced by TNF-α, IL-22, and LPS, which was, at least partly, correlated with the regulation of PI3K/Akt/mTOR and p38MAPK pathway activity.
Moreover, MCEO exerted favorable effects for IMQ-induced psoriatic dermatitis in BALB/c mice, mainly reflected by the remission of erythema, thickening, induration, and scaling, and alleviating general skin injury. Even MCEO downregulated the levels of IL-1β, IL-6, TNF-α, and IL-10, but not IL-17A. These results indicated that MCEO has shown great value in preventing and treating psoriasis. In addition, MCEO, combined with different target anti-inflammatory plant drugs, could become a star in psoriatic skin inflammation treatment. Although the specific mechanism of action needs to be investigated further, MCEO may be an excellent agent for an anti-psoriatic-like skin inflammation drug.
Psoriasis is an autoimmune disease; however, its exact pathogenesis remains unknown. Currently, synthetic drugs are commonly used to treat psoriasis, either by systemic injection or topical administration. However, the application of synthetic drugs shows many serious side effects in patients. As a result, novel, safer, and more effective natural botanical medicines have garnered the significant interest of researchers worldwide.34 Plants and their secondary metabolites have a unique advantage in discovering potential anti-psoriasis agents.34,35 Moreover, medicinal plants are easy to obtain and are cheap and effective. Treatment of psoriasis is reliant on them globally.36 Matricaria chamomilla, a kind of ancient aromatic plant of the family Compositae, its base volatile oil, has shown potent effects in cosmetics, including eliminating skin inflammation and irritation and reducing skin redness.11,12,16 Essential oils are a complex natural mixture of volatile secondary metabolites isolated from plants. The main components of essential oils are terpenes, and terpenes have been extensively studied for their anti-inflammation properties.37–39 MCEO display a unique advantage in wound healing and skin barrier repair, which is related to their anti-inflammation, antioxidant, and antibacterial properties.10,16,40 There are many kinds of ingredients in chamomile, such as cyclic ether, flavonoids, and total volatile oil, which can inhibit the growth of fungus. In addition, polysaccharides in the inflorescences have also shown significant anti-inflammatory effects, and MCEO also demonstrates anti-eczema effects.16,41–43 Regrettably, the specific mechanism and functional active ingredients of MCEO in dermatology remain unelucidated.
Network pharmacology is a new strategy to systematically study the relationship between diseases, drugs, and targets.21 MCEO include 16 components (Table 1). We obtained 130 MCEO network genes matched with the psoriasis gene set according to the results of GC-MS analysis. The PPIN network of MCEO was further analyzed with Cytoscape to explore psoriasis-related pathways. The network analysis using the MCEO compound network and psoriasis gene set demonstrated that the possible molecular mechanism of MCEO was highly associated with the PI3K/Akt, MAPK, and Th17 cell differentiation signaling pathway (Figure 1).
Inflammation is one of the important pathological features of psoriasis.44 Our study used clinical patient samples to analyze inflammatory phenotype, including IL-1β, IFN-γ, TNF-α, IL-10, and IL-17A. The levels of inflammatory cytokines increased gradually with the prolongation of the course of psoriasis, but not IL-10 (Supplement Table 1)(Supplement Figure 1). Cytokine IL-10 is a key anti-inflammatory mediator that protects the host from overreaction by pathogens and the microbiome, while playing an important role in other Settings such as sterile wound healing, autoimmunity, cancer, and homeostasis.45 Maybe the downregulation of anti-inflammatory cytokines is also a cause of enhanced inflammatory response in psoriasis patients. These results indicated that inflammation is a determining factor during the progress of psoriasis. Therefore, anti-inflammatory therapy has become an effective entry point for exploring new therapeutic drugs.
Until now, no relative research has analyzed the effect of MCEO on inflammation in keratinocytes. First, we confirmed the optimal concentration of MCEO and analyzed its function on the viability and cell cycle of HaCaT cells by CCK8 and flow cytometry. Our data found that MCEO (50µg/mL and 60µg/mL) had no cytotoxic effect on HaCaT. Subsequently, we detected the expression of relative apoptotic proteins in HaCaT cells treated with MCEO (Figures 2 and 3). The expression of caspase 9 protein was obviously increased in HaCaT cells treated with MCEO (60µg/mL). We aimed to eliminate background effects to elucidate the mechanisms of MCEO underlying anti-inflammation effects. Hence, we selected 50µg/mL of MCEO for subsequent experiments.
We used IL-22, TNF-α, and LPS cytokines to stimulate HaCaT cells and establish an in vitro inflammatory keratinocyte model.46–48 In this study, the mRNA expression of cytokines IL-1β, IL-6, TNF-α, IL-17A, TGF-β, and IL-10 was analyzed by qRT-PCR. MCEO treatment significantly decreased IL-1β, IL-6, and TGF-β mRNAs induced by IL-22, TNF-α, and LPS stimulation (Figures 4B, 5B, and 6B). In addition, IL-22 can activate the Akt/mTOR pathway,26 TNF-α induces inflammation and the recruitment of immune cells, which is related to the activation of the MAPK/Akt pathway.49 Liu et al found that LPS can act as a simulator, inducing inflammatory cytokines production in HaCaT cells, which is related to the induction of phosphorylation of p38MAPK.47 p38MAPK participate in a broad spectrum of cellular responses induced by external stimulus. External stimuli include proinflammatory factors and stress, which mediates the release of IL-6, IL-1, IL-8, or TNF-α proinflammatory cytokines.50 Consistently, our results clarified that MCEO treatment downregulated inflammation by controlling PI3K/Akt/mTOR and p38MAPK pathway activity and blocking the entry of p-mTOR and p-P38 into the nucleus in IL-22/TNF-α/LPS-induced HaCaT (Figures 4, 5 and 6).
Inflammation in dermatology is closely related to the PI3K/AKT/mTOR signaling axis, which is highly associated with the pathogenesis of skin disorders.51 The mTOR pathway in pathogenic skin diseases perhaps participates in crosstalk with other signaling pathways directly or indirectly, such as MAPK, PTEN, AMPK, and PKC.51 Consistently, we also found that MCEO cross-regulated the activation between PI3K/Akt/mTOR and p38MAPK pathway, which suppressed inflammation of HaCaT keratinocytes, perhaps via relative cytokines.
Since 2009, the mouse with IMQ-induced psoriatic-like skin inflammation has been widely used in preclinical psoriasis studies. Researchers usually use this animal model to study the efficacy, molecular mechanism, and absorption path of new drugs.52,53 IMQ was smeared on the skin of mice to induce psoriasis-like symptoms. Many clinical symptoms were similar to psoriasis, including rash, scales, hyperplasia of the epidermis, and inflammatory cell infiltration in model mice were detected. MCEO pretreatment significantly ameliorated those symptoms (Figure 7A and B). The PASI scores and H&E staining of skin tissue showed that MCEO treatment relieved skin lesions of IMQ-induced BALB/c mice (Figure 7C and D). Moreover, MCEO treatment decreased the levels of IL-1β, TNF-α, IFN-γ, and TGF-β, but not IL-17A in mice with IMQ-induced skin inflammation, as assessed by qRT-PCR assay and Luminex, correlated with inhibiting PI3K/Akt/mTOR and p38MAPK pathway activity (Figure 8A and B). Therefore, we considered that MCEO could ameliorate skin inflammation in IMQ-induced psoriatic-like skin inflammation mice, which is involved in downregulating the activity of PI3K/Akt/mTOR and p38MAPK pathway. Wang et al found that German chamomile volatile oil regulated T-cell lymphatic subpopulations to inhibit the Th17 cell differentiation signaling pathway, resulting in a reduction of interleukin 17 (IL-17), thereby inhibiting the activation of NF-κB and MAPK pathways, decreasing the secretion of the proinflammatory factors TNF-α and IL-6, and reducing inflammation.16 In addition, extract besides the anti-inflammatory effect of Matricaria chamomilla extracts, we found MC aqueous extract effects as disrupting Th1/Th2 balance to Th1 upregulation.54 Interesting, accumulated literature clarified that the IL-17/T-helper 17 (Th17) axis, a well-established proinflammatory cytokine, is involved in the pathophysiology of psoriasis.55,56 We detected the level of IL-17A in clinical patient samples by Luminex and found that the secretion level of IL-17A increased gradatim with the prolongation of the disease course (Supplement Figure 1). We did not find the change tendency of IL-17A mRNA in HaCaT cells treated with TNF-α, IL-22, and LPS compared with normal control cells. However, in psoriatic-like skin inflammation model mice, we only observed that the level of IL-17A production in serum increased. MCEO treatment significantly reduced IL-1β, IFN-γ, and TNF-α expression in the skin tissue and serum, but not IL-17A. This phenomenon indicates that IL-17A production is not associated with the activation of PI3K/Akt or MAPK signaling pathways. The way in which MCEO down-regulates inflammatory response does not affect IL-17A expression. There are reports that IL-17 inhibitors (ixekizumab and secukinumab) treated psoriasis as first-line clinical drugs. According to clinician experience and relative reports, targeting IL-17A obtained some surprising therapeutic effects in psoriasis.56 Ho et al discovered that rosmarinic acid, a natural plant extract, is an IL-17A-targeting inhibitor that can block the IL-17A/IL-17RA axis to improve psoriatic skin inflammation in mice.57 These new findings give us a new revelation: the combination of anti-inflammatory plant drugs with different targets can be selected, which may be a potential new approach for psoriasis therapy in the future field of psoriasis treatment.
To be mentioned, Azulene is one of the main components in MCEO, which has been recognized for its application in medicinal therapy against inflammation.58,59 Azulene is one of these compounds whose anti-inflammatory potential have been shown by the previous studies, but a detailed analysis of its effect at the cellular level in terms of proinflammatory cytokine production has not been studied yet. In the following experiments, we will investigate the anti-inflammatory effect of Azulene on HaCaT cells in vitro and on IMQ-induced psoriatic-like skin inflammation.
Conclusion
In the present study, we first discovered that MCEO, a plant oil, possesses a natural advantage in terms of skin barrier repair, which treatment inhibits inflammation in HaCaT keratinocytes, the potential mechanisms correlated with the activation and crosstalk of PI3K/Akt/mTOR and p38MAPK pathways. In addition, MCEO ameliorates the skin lesions in IMQ-induced psoriatic-like skin inflammation mice, downregulating the levels of inflammatory cytokines.
Finding novel, safe, and potential agents from natural plants for treating inflammatory skin disorders is a novel strategy for future research. Anti-inflammatory plant drugs with different targets with combined applications were a potential therapeutic strategy in psoriasis. Our new findings provide a sufficient experimental and theoretical basis for the clinical application of MCEO.
Ethics Approval and Consent to Participate
The human study was approved by the Ethics Review Committee on Human Research of Taizhou Second People’s Hospital (tzey-ky202101). The animal studies were agreed upon by the Ethical Committees of Taizhou University (Taizhou, China) (TZXY-2022-20221027).
Acknowledgment
We thank Jiaxing Hanjingfangcao Bioengineering Company Limited for providing MCEO.
Funding
This research was supported by grants from the Taizhou Science and Technology Planning Project (20ywa57, 21ywa61, 20ywb59), the natural science foundation in Zhejiang Province (LGD21H100002). The funders had no role in the study design, data collection and analysis, publication decision, or manuscript preparation. All authors had full access to the full data in the study and accepted responsibility to submit for publication.
Disclosure
The authors have no conflicts of interest to declare for this work.
References
1. World Health Organization. Global Report on Psoriasis. Geneva: World Health Organization; 2016.
2. Branisteanu DE, Cojocaru C, Diaconu R, et al. Update on the etiopathogenesis of psoriasis (Review). Exp Ther Med. 2022;23(3):201. doi:10.3892/etm.2022.11124
3. Rapalli VK, Waghule T, Gorantla S, et al. Psoriasis: pathological mechanisms, current pharmacological therapies, and emerging drug delivery systems. Drug Discov Today. 2020;25(12):2212–2226. doi:10.1016/j.drudis.2020.09.023
4. Gladman DD. Should methotrexate remain the first-line drug for psoriasis? Lancet. 2017;389(10068):482–483. doi:10.1016/S0140-6736(16)32574-0
5. Ronholt K, Iversen L. Old and new biological therapies for psoriasis. Int J Mol Sci. 2017;18(11):2297. doi:10.3390/ijms18112297
6. Ahuja A, Gupta J, Gupta R. Miracles of herbal phytomedicines in treatment of skin disorders: natural healthcare perspective. Infect Disord Drug Targets. 2021;21(3):328–338. doi:10.2174/1871526520666200622142710
7. Bonesi M, Loizzo MR, Provenzano E, et al. Anti-psoriasis agents from natural plant sources. Curr Med Chem. 2016;23(12):1250–1267. doi:10.2174/0929867323666160321121819
8. Dai YL, Li Y, Wang Q, et al. Chamomile: a review of its traditional uses, chemical constituents, pharmacological activities and quality control studies. Molecules. 2022;28(1):133. doi:10.3390/molecules28010133
9. Lin TK, Zhong L, Santiago JL. Anti-inflammatory and skin barrier repair effects of topical application of some plant oils. Int J Mol Sci. 2018;19(1):70. doi:10.3390/ijms19010070
10. Hajaji S, Jabri M-A, Alimi D, et al. Chamomile methanolic extract mitigates small bowel inflammation and ROS overload related to the intestinal nematodes infection in mice. Acta Parasitol. 2019;64(1):152–161. doi:10.2478/s11686-019-00027-x
11. Weber L, Kuck K, Jurgenliemk G, et al. Anti-Inflammatory and barrier-stabilising effects of Myrrh, coffee charcoal and chamomile flower extract in a co-culture cell model of the intestinal mucosa. Biomolecules. 2020;10(7):1033. doi:10.3390/biom10071033
12. Majidi M, Heidarnejad F, Naseri M, et al. Marham-Mafasel decrease joint inflammation and IL-1β gene expression in rheumatoid arthritis animal model. Vet Med Sci. 2021;7(4):1417–1425. doi:10.1002/vms3.430
13. Amsterdam JD, Li QS, Xie SX, et al. Putative antidepressant effect of chamomile (Matricaria chamomilla L.) oral extract in subjects with comorbid generalized anxiety disorder and depression. J Altern Complement Med. 2020;26(9):813–819. doi:10.1089/acm.2019.0252
14. Niknam S, Tofighi Z, Faramarzi MA, et al. Polyherbal combination for wound healing: matricaria chamomilla L. and Punica granatum L. Daru. 2021;29(1):133–145. doi:10.1007/s40199-021-00392-x
15. Bakun P, Czarczynska-Goslinska B, Goslinski T, et al. In vitro and in vivo biological activities of azulene derivatives with potential applications in medicine. Med Chem Res. 2021;30(4):834–846. doi:10.1007/s00044-021-02701-0
16. Wang W, Wang Y, Zou J, et al. The mechanism action of German chamomile (Matricaria recutita L.) in the treatment of eczema: based on dose-effect weight coefficient network pharmacology. Front Pharmacol. 2021;12:706836. doi:10.3389/fphar.2021.706836
17. Ma DW. A traditional Chinese medicine composition for sunscreen cosmetics containing marigold. CN105796448A; 2016.
18. Lu HM. A kind of compound plant extract repair liquid and preparation method thereof. CN112754979A; 2021.
19. Chen GR, Xin YH. Antiviral traditional Chinese medicine composition and its application. CN113797266A; 2020.
20. Chung JY, Park N, Kim MH, Yang WM. Abies holophylla leaf essential oil alleviates allergic rhinitis based on network pharmacology. Pharmaceutics. 2023;15(4):1195. doi:10.3390/pharmaceutics15041195
21. Yao T, Yan J, Li Y, et al. An integrated approach based on phytochemistry, network pharmacology and metabolomics reveals the mechanism of action of Xanthium strumarium L. for allergic rhinitis. RSC Adv. 2020;10(67):41154–41163. doi:10.1039/D0RA06763F
22. Ogawa E, Sato Y, Minagawa A, et al. Pathogenesis of psoriasis and development of treatment. J Dermatol. 2018;45(3):264–272. doi:10.1111/1346-8138.14139
23. He Q, Liu N, Hu F, et al. Circ_0061012 contributes to IL-22-induced proliferation, migration and invasion in keratinocytes through miR-194-5p/GAB1 axis in psoriasis. Biosci Rep. 2021a;41(1). doi:10.1042/BSR20203130
24. Honma M, Hayashi K. Psoriasis: recent progress in molecular-targeted therapies. J Dermatol. 2021;48(6):761–777. doi:10.1111/1346-8138.15727
25. Wang X, Yang L, Yang L, et al. Gypenoside IX suppresses p38 MAPK/Akt/NFkappaB signaling pathway activation and inflammatory responses in astrocytes stimulated by proinflammatory mediators. Inflammation. 2017;40(6):2137–2150. doi:10.1007/s10753-017-0654-x
26. Mitra A, Raychaudhuri SK, Raychaudhuri SP. IL-22 induced cell proliferation is regulated by PI3K/Akt/mTOR signaling cascade. Cytokine. 2012;60(1):38–42. doi:10.1016/j.cyto.2012.06.316
27. Chima M, Lebwohl M. TNF inhibitors for psoriasis. Semin Cutan Med Surg. 2018;37(3):134–142. doi:10.12788/j.sder.2018.039
28. Kim A, Nam YJ, Shin YK, et al. Rotundarpene inhibits TNF-alpha-induced activation of the Akt, mTOR, and NF-kappaB pathways, and the JNK and p38 associated with production of reactive oxygen species. Mol Cell Biochem. 2017;434(1–2):113–125. doi:10.1007/s11010-017-3041-x
29. Leng H, Pu L, Xu L, et al. Effects of aloe polysaccharide, a polysaccharide extracted from Aloe vera, on TNFalphainduced HaCaT cell proliferation and the underlying mechanism in psoriasis. Mol Med Rep. 2018;18(3):3537–3543. doi:10.3892/mmr.2018.9319
30. Park K, Lee JH, Cho HC, et al. Down-regulation of IL-6, IL-8, TNF-α and IL-1β by glucosamine in HaCaT cells, but not in the presence of TNF-α. Oncol Lett. 2010;1(2):289–292. doi:10.3892/ol_00000051
31. Balato A, Lembo S, Mattii M, et al. IL-33 is secreted by psoriatic keratinocytes and induces proinflammatory cytokines via keratinocyte and mast cell activation. Exp Dermatol. 2012;21(11):892–894. doi:10.1111/exd.12027
32. Gvirtz R, Ogen-Shtern N, Cohen G. Kinetic cytokine secretion profile of LPS-induced inflammation in the human skin organ culture. Pharmaceutics. 2020;12(4):299. doi:10.3390/pharmaceutics12040299
33. Daniyal M, Akram M, Zainab R, et al. Progress and prospects in the management of psoriasis and developments in phyto-therapeutic modalities. Dermatol Ther. 2019;32(3):e12866. doi:10.1111/dth.12866
34. Deng S, May BH, Zhang AL, et al. Phytotherapy in the management of psoriasis: a review of the efficacy and safety of oral interventions and the pharmacological actions of the main plants. Arch Dermatol Res. 2014;306(3):211–229. doi:10.1007/s00403-013-1428-4
35. Nowak-Perlak M, Szpadel K, Jablonska I, et al. Promising strategies in plant-derived treatments of psoriasis-update of in vitro, in vivo, and clinical trials studies. Molecules. 2022;27(3):591. doi:10.3390/molecules27030591
36. Thitinarongwate W, Nimlamool W, Khonsung P, et al. Anti-Inflammatory activity of essential oil from Zingiber ottensii valeton in animal models. Molecules. 2022;27(13):4260. doi:10.3390/molecules27134260
37. Wong CF, Poon CK, Ng TW, et al. Anti-inflammatory, antipyretic efficacy and safety of inhaled Houttuynia cordata thunb. essential oil formulation. J Ethnopharmacol. 2022;297:115541. doi:10.1016/j.jep.2022.115541
38. Zhao C, Zhang Z, Nie D, et al. Protective effect of lemon essential oil and its major active component, D-Limonene, on intestinal injury and inflammation of E. coli-challenged mice. Front Nutr. 2022;9:843096. doi:10.3389/fnut.2022.843096
39. Vaughn AR, Clark AK, Sivamani RK, et al. Natural oils for skin-barrier repair: ancient compounds now backed by modern science. Am J Clin Dermatol. 2018;19(1):103–117. doi:10.1007/s40257-017-0301-1
40. Anderson C, Lis-Balchin M, Kirk-Smith M. Evaluation of massage with essential oils on childhood atopic eczema. Phytother Res. 2000;14(6):452–456. doi:10.1002/1099-1573(200009)14:6<452::AID-PTR952>3.0.CO;2-4
41. El-Salamouni NS, Ali MM, Abdelhady SA, et al. Evaluation of chamomile oil and nanoemulgels as a promising treatment option for atopic dermatitis induced in rats. Expert Opin Drug Deliv. 2020;17(1):111–122. doi:10.1080/17425247.2020.1699054
42. Patzelt-Wenczler R, Ponce-Poschl E. Proof of efficacy of Kamillosan(R) cream in atopic eczema. Eur J Med Res. 2000;5(4):171–175.
43. Zhou X, Chen Y, Cui L, et al. Advances in the pathogenesis of psoriasis: from keratinocyte perspective. Cell Death Dis. 2022;13(1):81. doi:10.1038/s41419-022-04523-3
44. Guilloteau K, Paris I, Pedretti N, et al. Skin inflammation induced by the synergistic action of IL-17A, IL-22, oncostatin M, IL-1{alpha}, and TNF-{alpha} recapitulates some features of psoriasis. J Immunol. 2010;184(9):5263–5270. doi:10.4049/jimmunol.0902464
45. Saraiva M, Vieira P, O’Garra A. Biology and therapeutic potential of interleukin-10. J Exp Med. 2020;217(1):e20190418. doi:10.1084/jem.20190418
46. Li C, Xiao L, Jia J, et al. Cornulin is induced in psoriasis lesions and promotes keratinocyte proliferation via phosphoinositide 3-Kinase/Akt pathways. J Invest Dermatol. 2019;139(1):71–80. doi:10.1016/j.jid.2018.06.184
47. Liu Y, Zhao C, Ma Q, et al. Sinomenine retards LPS-elicited inflammation via down-regulating CCAT1 in HaCaT cells. Life Sci. 2019;233:116703. doi:10.1016/j.lfs.2019.116703
48. Kalliolias GD, Ivashkiv LB. TNF biology, pathogenic mechanisms and emerging therapeutic strategies. Nat Rev Rheumatol. 2016;12(1):49–62. doi:10.1038/nrrheum.2015.169
49. Pahlavani HA. Exercise-induced signaling pathways to counteracting cardiac apoptotic processes. Front Cell Dev Biol. 2022;10:950927. doi:10.3389/fcell.2022.950927
50. Karagianni F, Pavlidis A, Malakou LS, et al. Predominant role of mTOR signaling in skin diseases with therapeutic potential. Int J Mol Sci. 2022;23(3):1693. doi:10.3390/ijms23031693
51. Lin YK, Yang SH, Chen CC, et al. Using imiquimod-induced psoriasis-like skin as a model to measure the skin penetration of anti-psoriatic drugs. PLoS One. 2015;10(9):e0137890. doi:10.1371/journal.pone.0137890
52. Chuang SY, Lin CH, Sung CT, et al. Murine models of psoriasis and their usefulness for drug discovery. Expert Opin Drug Discov. 2018;13(6):551–562. doi:10.1080/17460441.2018.1463214
53. Wu S, Zhao M, Sun Y, et al. The potential of Diosgenin in treating psoriasis: studies from HaCaT keratinocytes and imiquimod-induced murine model. Life Sci. 2020;241:117115. doi:10.1016/j.lfs.2019.117115
54. Asadi Z, Ghazanfari T, Hatami H. Anti-inflammatory effects of Matricaria chamomilla extracts on BALB/c mice macrophages and lymphocytes. Iran J Allergy Asthma Immunol. 2020;19(S1):63–73. doi:10.18502/ijaai.v19i(s1.r1).2862
55. Zhang B, Domling A. Small molecule modulators of IL-17A/IL-17RA: a patent review (2013–2021). Expert Opin Ther Pat. 2022;32(11):1161–1173. doi:10.1080/13543776.2022.2143264
56. Lin TK, Zhong L, Santiago JL. Anti-inflammatory and skin barrier repair effects of topical application of some plant oils. Int J Mol Sci. 2017;19(1):70.
57. Ho TY, Lo HY, Liu IC, Huang SL, Hsiang CY. Rosmarinic acid ameliorated psoriatic skin inflammation in mice through the novel inhibition of the interleukin-17A/interleukin-17A receptor interaction. Food Funct. 2022;13(12):6802–6812. doi:10.1039/D2FO00417H
58. Nolting DD, Nickels M, Price R, Gore JC, Pham W. Synthesis of bicyclo[5.3.0]azulene derivatives. Nat Protoc. 2009;4(8):1113–1117. doi:10.1038/nprot.2009.99
59. Ayaz F, Yuzer A, Ince T, Ince M. Anti-cancer and anti-inflammatory activities of bromo- and cyano-substituted azulene derivatives. Inflammation. 2020;43(3):1009–1018. doi:10.1007/s10753-020-01186-0
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.