")
Back to Journals » Blood and Lymphatic Cancer: Targets and Therapy » Volume 14
AC024896.1/miR-363-3p Axis Regulates the Malignant Progression of Acute Myeloid Leukemia by Cuproptosis-Related Gene MYO1B
Authors Zhang J, Zheng Y, Liu H, Liu B
Received 9 December 2023
Accepted for publication 10 March 2024
Published 24 March 2024 Volume 2024:14 Pages 17—30
DOI https://doi.org/10.2147/BLCTT.S454115
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Wilson Gonsalves
Jie Zhang,1 Yuhuan Zheng,1 Hongjuan Liu,2 Bo Liu3
1Department of Blood Transfusion, Ankang Central Hospital, Ankang, Shannxi, 725000, People’s Republic of China; 2Operating Room, Ankang Central Hospital, Ankang, Shannxi, 725000, People’s Republic of China; 3Department of Blood Transfusion, Ankang Hospital of Traditional Chinese Medicine, Ankang, Shannxi, 725000, People’s Republic of China
Correspondence: Bo Liu, Ankang Hospital of Traditional Chinese Medicine, Ankang Municipality of Bashan East Road No. 47, Ankang, Shannxi, 725000, People’s Republic of China, Email [email protected]
Background: Acute myeloid leukemia (AML) is a hematological malignancy with poor patient prognosis. Cuprotosis is a newly discovered cell death that regulates the proliferation and progression of tumor cells. Long non-coding RNAs (lncRNAs) are key molecules and potential biomarkers for the diagnosis and treatment of various diseases. However, the effect of cuprotosis-associated lncRNAs on AML remains unclear.
Objective: The aim of this study was to investigate the relationship between the expression of cuprotosis-related gene and the prognosis of AML.
Methods: Consensus cluster analysis was performed on AML patients according to the cuprotosis-related gene expression matrix, and survival analysis and differential gene analysis were performed. Then lncRNA and miRNA related to AML tumor progression were screened according to univariate COX regression analysis. After that, Kaplan-Meier analysis, correlation analysis, and AUC curve were used to determine the ceRNA network that might regulate AML. The regulatory relationship of ceRNA was verified in AML cell lines by RT-qPCR and Western blotting.
Results: The AC024896.1/miR-363-3p axis drives MYO1B to promote the malignant progression of AML. First, a change in the expression of AC024896.1 and miR-363-3p can affect the proliferation of AML by regulating MYO1B. Mechanistically, AC024896.1 regulates the expression of MYO1B as the ceRNA of miR-363-3p. Moreover, the regulation of AC024896.1 in the malignant progression of AML depends partly on miR-363-3p.
Conclusion: In summary, our study reveals AC024896.1/miR-363-3p/MYO1B Axis in AML, which can be regarded as a new potential target for the diagnosis and treatment of AML.
Keywords: acute myeloid leukemia, cuprotosis, ceRNA network, bioinformatics analysis
Introduction
Acute myeloid leukemia (AML) is a type of acute leukemia that develops in the blood and bone marrow, most commonly occurs in the elderly, and is more common in men compared to women.1 AML causes excessive production of abnormal myeloblasts, crowds out the bone marrow, and prevents the generation of normal blood cells, making severe cases life-threatening. The prognoses of these patients vary considerably depending on the subtype.2,3 In 2015, nearly one million people worldwide developed acute myelogenous leukemia, and approximately 15% died.4 The diagnosis of AML usually includes a physical examination (blood tests and bone marrow biopsy), laboratory examination (flow cytometry and cytogenetic testing), and imaging examination (CT, MRI, and X-ray scans).5 Although existing diagnostic methods for AML are generally effective, these have several limitations. For example, invasive bone marrow biopsies and blood tests that cause discomfort to patients do not necessarily detect AML, especially in the early stages, which implies that the diagnosis and treatment may be delayed.6 Unlike most cancers with solid tumors, surgical methods are unsuitable for AML, and the first treatment is chemotherapy.7 In specific cases where conditions permit, stem cell transplantation is also a better choice, and the targeted therapy methods that have emerged in recent years bear potential.8 However, long-term chemotherapy can easily lead to drug resistance and toxic side effects.9 Even if the treatment is successful, there is a possibility of recurrence, requiring additional treatment intervention. Earlier reports have been strengthening the study of the diagnosis and treatment of AML, in an attempt to obtain a combination of novel and effective diagnosis and treatment methods.
Non-coding RNA (ncRNA) is an RNA molecule that does not code for protein but has various regulatory functions in cells.10 MicroRNAs (miRNAs) are small ncRNAs that can control gene expression by binding to the target messenger RNA (mRNA), thus inhibiting its translation.11 Altered expression of miRNAs can contribute to cancer development by affecting the genes related to cell cycle regulation, apoptosis, and other key cellular processes.12 The study of miRNAs in cancer has led to the recognition of novel diagnostic and prognostic biomarkers and potential targets for cancer therapy.13 miRNAs are dysregulated in AML.14 Thus, these can be targeted by anti-miRNA oligonucleotides or miRNA mimics to suppress leukemia cell growth and sensitize them to chemotherapy.15 MiR-363-3p is a miRNA that is dysregulated as both a tumor suppressor and an oncogene in various cancers, including lung, gastric, and colorectal cancers.16 Long non-coding RNAs (lncRNAs) are crucial in gene expression regulation, cell differentiation, and development.17 LncRNAs can act as “molecular sponges” to adsorb proteins and miRNAs in large quantities and bind to the latter as competing endogenous RNAs (ceRNAs), thus affecting the gene expression.18 Besides their function in cancer development and progression, lncRNAs may also have diagnostic and prognostic value.19 Studies have found that the expression patterns of lncRNAs can distinguish diverse cancer subtypes, and high expression of certain lncRNAs is related to poor prognosis.20,21 An in-depth study of the role of lncRNAs in cancer may yield therapeutic targets.22 Additional research is needed to adequately elucidate alternative lncRNAs in AML and develop effective therapeutics targeting these molecules. AC024896.1 is a lncRNA located on human chromosome 1.23 The function of AC024896.1 is unclear but recent studies suggest that it may play a non-negligible role in cancer,24 while some studies proposed that AC024896.1 might be a potential biomarker and therapeutic target for certain types of cancers. AC024896.1 has not yet been implicated in the field of AML, and further research is needed to fully understand the function of this lncRNA and develop effective treatments. ceRNAs are RNA molecules that regulate gene expression by competing for shared miRNA binding sites.25 In cancer, abnormal expression of ceRNAs can promote tumorigenesis by altering the expression of cell proliferation, apoptosis, and metastasis-related genes, including those in AML.26 The study of ceRNAs has provided new insights into the complex mechanisms of gene expression regulation.
Cuproptosis, also known as ferroptosis-like cell death, is a form of programmed cell death triggered by copper accumulation in cells.27 In cancer, copper accumulation can promote tumor growth and resistance to chemotherapy, and targeting cuproptosis pathways may offer a novel approach to cancer therapy.28 Further research on the role of cuproptosis in AML is warranted. However, based on the basic research of cuproptosis on different tumors, understanding its mechanism in AML is expected to provide new ideas for its treatment.
The MYO1B gene encodes Myosin IB, which functions in various cellular processes.29 MYO1B is upregulated in many cancers, including breast, colorectal, and lung cancers.30 Although the exact mechanism by which MYO1B promotes cancer progression is not fully understood, earlier reports generally believe that MYO1B may promote tumor growth and metastasis by enhancing cell migration and invasion.31 A previous study found that MYO1B may regulate cellular responses to cuproptosis and protect cells from copper-induced cell death.32 Nevertheless, the function of MYO1B in AML and its regulatory mechanism of receiving ncRNA need to be elucidated.
In this study, we found that AC024896.1, as the ceRNA of miR-363-3p, was involved in regulating the expression of the cuproptosis-related gene, MYO1B, in AML. The direct relationship among AC024896.1, miR-363-3p, and MYO1B is key in controlling the malignant progression of AML. Finally, we predicted and fully validated that AC024896.1 binds to miR-363-3p and acts as its molecular sponge to regulate MYO1B levels. This study laid a solid foundation for the research and application of ncRNA and copper death in AML and pointed out the direction for further development for the diagnosis and treatment of these patients.
Materials and Methods
Data Collection and Differential Gene Expression Analysis
Using The Cancer Genome Atlas (TCGA) database, 150 AML transcriptomes and clinically relevant information (survival time, age, sex, etc.) were downloaded. The differentially expressed miRNAs, mRNAs, and lncRNAs were analyzed and screened. The “ggplot2” and “pheatmap” packages were used to create volcano maps and heat maps to visualize the levels of differentially expressed genes.
Consistent Cluster Analysis
The “ConsensusClusterPlus” package was used to analyze the mRNA expression matrix of AML patients in TCGA cohort, and unsupervised cluster analysis (k-means clustering) was performed with 100 repeats to ensure the stability of the classification.
Batch Univariate Cox Analysis
To screen the prognostic differentially expressed genes, univariate Cox analysis was performed to compare and evaluate the ROC curves and survival analysis was conducted between the high-risk group and the low-risk group with P < 0.05 as the significance criterion. In this study, the “survivalROC” package was used for analysis, while the “survminer” and “pheatmap” packages were used for plotting.
Construction of the lncRNA-microRNA-mRNA Network
First, the miRcode database was used to predict interactions between lncRNAs and miRNAs. Subsequently, miRNA-mRNA interactions were searched using TargetScan, mirTarBase, and miRDB databases. Furthermore, we analyzed the expression of lncRNAs in ceRNA networks by examining their association with targeted miRNAs. Finally, the identified co-expression competing triplets were used to construct the mRNA-miRNA-lncRNA networks. Cytoscape v3.7.2 was used for visualizing the ceRNA network.
DAVID Enrichment Analysis
The enrichment analysis of differentially expressed genes was conducted using the DAVID database; GeneRatio and enrichment factors were calculated and visualized using“ggplot”. The larger the enrichment factor, the more significant the enrichment level of differentially expressed genes in that particular pathway.
Cell Culture and Cell Transfection
AML cell lines, THP1 and Kasumi were purchased from ATCC. THP1 and Kasumi cells were cultured in RPMI 1640 media, and HEK293T cells were cultured in 10% DMEM. Both media were supplemented with 10% FBS and cells were grown in an incubator at 37 °C with 5% CO2. According to the instructions, psPAX2 and pMD2.G plasmids were transfected into 293T cells. After 48 hours, the supernatant was collected and filtered through a 0.45 µm filter membrane. Next, THP1 and Kasumi cells were infected with virus solution, and 48 hours later, fresh medium was replaced and 5 mg/mL puromycin was added to screen stably transfected cell lines.
CCK8 Assay
The CCK8 kit (Beyotime, China) was used to detect cell proliferation. Briefly, the cells were seeded in 96-well plates and incubated at 37 °C for 48 hours, followed by 2 hours of incubation with 10 μL of CCK-8 reagent. Finally, optical density (OD) values were measured at 450 nm using a Thermo Scientific enzyme marker.
RNA Extraction and qRT-PCR
Total cellular RNA was extracted using the TRIzol reagent (TaKaRa, Japan), and mRNA was reverse-transcribed into cDNA following the manufacturer’s instructions. Next, the cDNA was amplified using SYBR Premix Ex Taq kit (TaKaRa). The experiments were repeated at least thrice and the primer sequences used in the study were as follows:
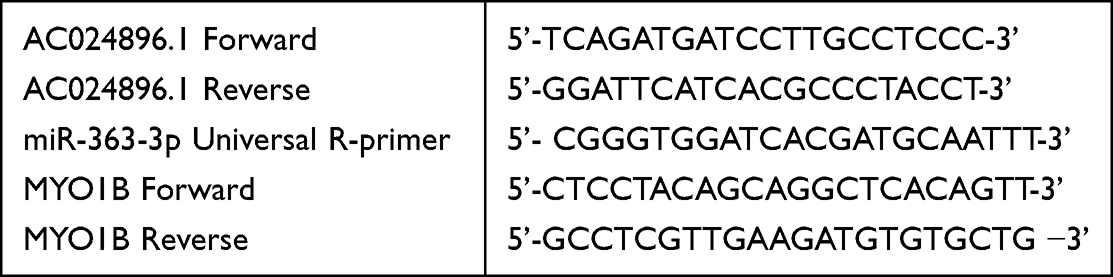 |
|
Luciferase Assay
Wild-type vector and mutant vector carrying AC024896.1 were constructed on PGL4.15, and HEK293T cells were co-transfected with the specified luciferase reporter gene and sea kidney luciferase reporter gene for 24 hours. Luciferase activity in cell lysates was measured using a dual luciferase reporter assay kit (Beyotime, China).
Statistical Analysis
Two-tailed Student’s t-test was used for the comparison between the two groups. Spearman correlation analysis was used for the gene expression correlation study. All data were analyzed using GraphPad Prism 8 software, and P < 0.05 was considered statistically significant.
Results
Consensus Cluster Analysis of Genes Related to Cuproptosis
Cuproptosis plays a crucial role in AML. It is a programmed cell death triggered by copper accumulation in cells. In order to divide AML patients into groups with different levels of cuproptosis, we performed cluster analysis according to the expression matrix of 13 cuproptosis-related genes. We divided 150 AML patients from TCGA database into k groups (k = 2–9). At k = 2, the optimal data classification was obtained (Figure 1A and Figure S1A-B). The overall survival rates between the two groups were compared, and Kaplan-Meier results showed a significantly longer survival in group 1 than in group 2 (Figure 1B). A total of 248 up-regulated and 50 down-regulated differentially expressed genes were obtained between the two groups (Figure 1C–D and Figure S1C).
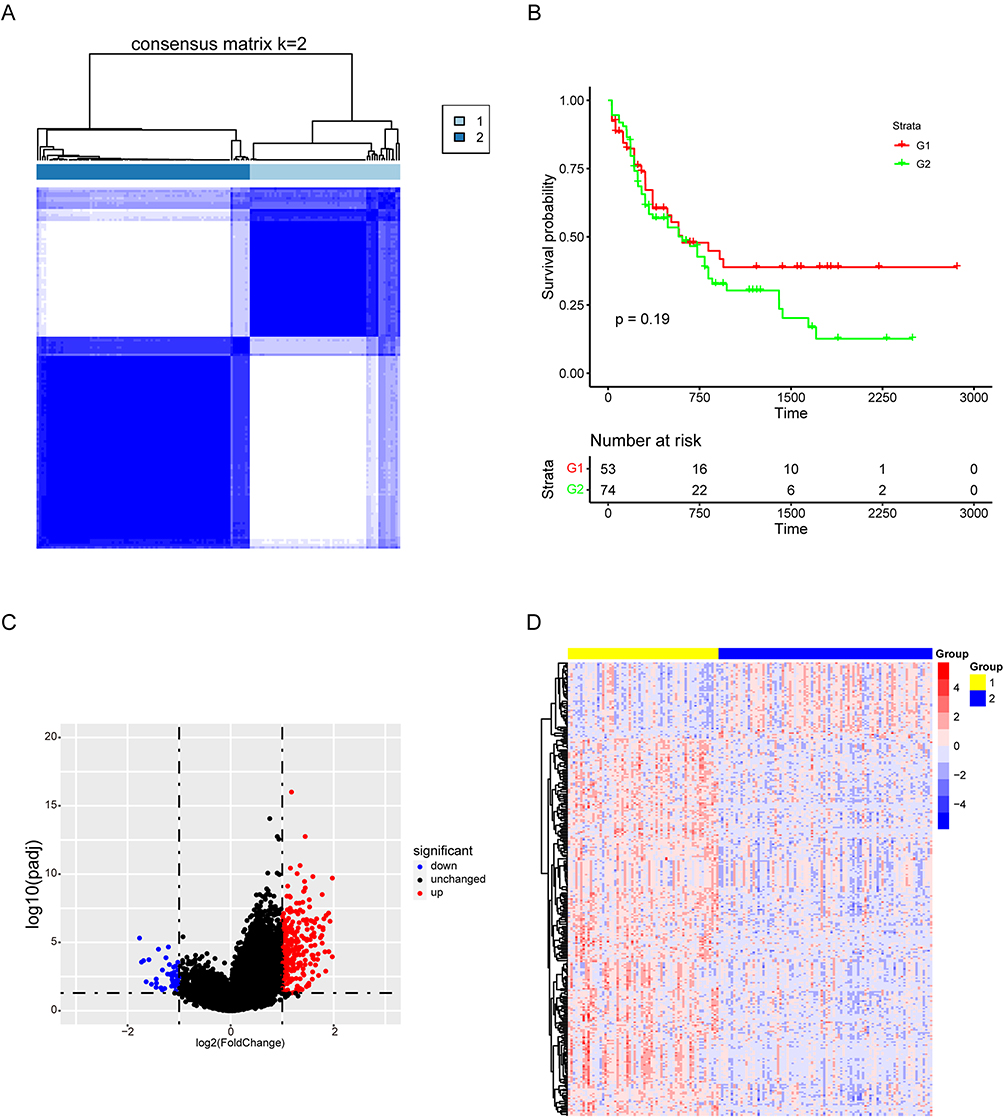 |
Figure 1 Consensus cluster analysis of genes related to cuproptosis. (A) Consensus clustering heatmap of genes associated with cuproptosis with k = 2. (B) Kaplan-Meier survival curve between cluster 1 and cluster 2. (C) The volcanic map showed the differential genes of the two clusters. (D) Heatmap of differential genes. |
Screening of lncRNAs and miRNAs Associated with AML Tumor Progression
The level of expression of tumor markers changed significantly during the progression of malignant tumors, and a large proportion of them may be oncogenes and tumor suppressor genes that dynamically regulate tumors. However, the role of the ceRNA axis of these genes in AML tumors has not been elucidated. To explore the underlying mechanisms, univariate cox analysis was performed for lncRNAs and miRNAs of AML samples, and the top 10 among each were obtained (Figure 2A–B).
 |
Figure 2 Screening of LncRNAs and miRNAs associated with AML tumor progression. (A–B) Forest maps showed the prognostic value of lncRNAs and miRNAs based on univariate cox proportional hazard regression analysis (P < 0.05). |
AC024896.1/miR-363-3p/MYO1B May Be an Important ceRNA Axis Regulating AML
In accordance with the above results, the ceRNA network regulating the malignant progression of AML was constructed using lncRNAs and miRNAs obtained by cox analysis and mRNAs obtained by differential analysis of cuproptosis-related genes. The network contained 29 lncRNAs, 2 miRNAs, 7 mRNAs, and 137 sets of interaction axes (Figure 3A and Figure S2A). Among mRNAs, MYO1B is a member of the actin motor protein myosin superfamily, which promotes cancer cell proliferation by regulating various signaling pathways. Hence, we focused our research on the ceRNA axis regulating MYO1B. Figure 3B illustrates the signaling axes that may be regulated by MYO1B.
 |
Figure 3 AC024896.1/miR−363−3p/MYO1B may be an important ceRNA axis in the regulation of AML. (A) Sankey diagram for the ceRNA network in ALL. Each rectangle represents a gene, and the connection degree of each gene is visualized based on the size of the rectangle. (B) The ceRNA regulatory network. |
AC024896.1 May Regulate the Nucleic Acid and Protein Levels of MYO1B
To identify the major lncRNAs regulating MYO1B, survival analysis was performed for AC006064.2, AC024896.1, AC066613.2, AC073842.2, AC091117.2, AC093423.2, AC103876.1, AL139193.2, AL683813.1, LINC02280 and Z97989.1. The results of survival analysis and correlational analysis showed that among the 11 lncRNAs, the expression of AC024896.1 was highly correlated with worse survival and the expression of MYO1B (Figure 4A–B), and the AUC curve showed the sensitivity of the survival (Figure S3A). To further screen the results, ASO knockout of these lncRNAs was used to detect the mRNA levels of MYO1B were evaluated. The results showed that AC024896.1 significantly regulated the expression of MYO1B, while no statistically significant difference was observed when other lncRNAs were knocked down (Figure 4C). In summary, AC024896.1 may be the main lncRNA regulating MYO1B.
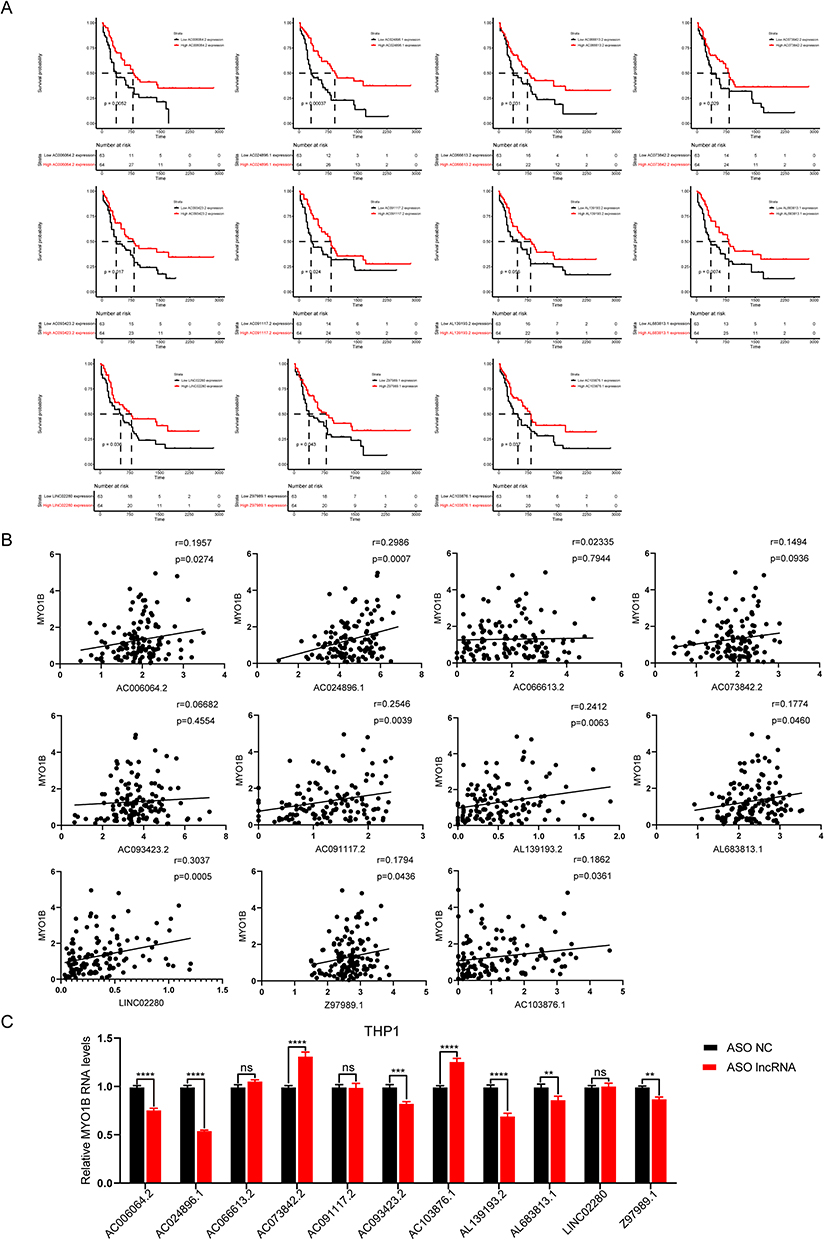 |
Figure 4 AC024896.1 may regulate the nucleic acid and protein levels of MYO1B. (A) Survival analysis in the high and low expression lncRNAs groups. (B) Correlation analysis between these lncRNAs and MYO1B. (C) The expression level of MYO1B after knockdown of these lncRNAs using ASO was examined by qPCR. (**P < 0.01, and ***P < 0.001, ****P < 0.0001). |
AC024896.1, miR-363-3p and MYO1B Regulate the Malignant Progression of AML
According to the above prediction results, AC024896.1/miR-363-3p/MYO1B may be the ceRNA axis regulating the development of AML but its role in the malignant progression of AML has not been elucidated. Therefore, AML samples were divided into the AC024896.1-high expression group and AC024896.1-low expression group for differential and enrichment analyses (the same analyses were also performed for miR-363-3p and MYO1B). Moreover, the regulatory genes of the three were primarily enriched in cell proliferation pathways (Figure 5A and Figure S4A-C). To further verify the regulatory effects of the three genes on AML proliferation, CCK8 experiments were performed following the inhibition of each of the three genes, and the results were consistent with the findings from enrichment analysis (Figure 5B–C). These results indicated that the three genes could regulate the malignant progression of AML.
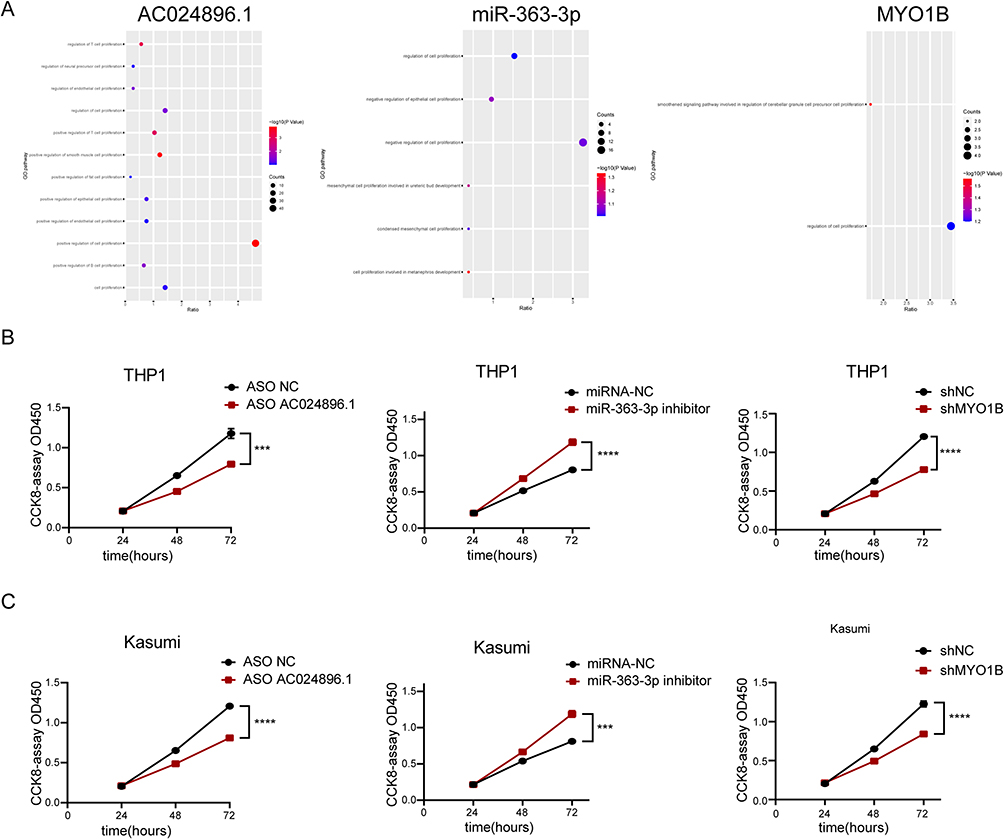 |
Figure 5 AC024896.1, miR-363-3p and MYO1B regulate the malignant progression of AML. (A) GSEA enrichment map. (B–C) CCK-8 assay showed that AC024896.1, miR-363-3p or MYO1B knockdown inhibited THP1 and Kasumi cells activity. (***P < 0.001, ****P < 0.0001). |
LncRNA AC024896.1 and miR-363-3p Can Regulate MYO1B in AML
To further explore the relationship between AC024896.1, miR-363-3p and MYO1B protein, the three genes were subjected to pearson correlation analysis. The results showed a high R-value and a certain correlation between the three (Figure 6A). After knocking down AC024896.1 with ASO, the changes in miR-363-3p and MYO1B RNA were detected. MYO1B was downregulated, while miR-363-3p was upregulated. Next, the changes in the levels of AC024896.1 and MYO1B RNA were detected after overexpression and inhibition of miR-363-3p with corresponding mimic and inhibitor, respectively. The results showed that AC024896.1 and MYO1B were negatively regulated by miR-363-3p (Figure 6B). These results indicated that lncRNA AC024896.1 and miR-363-3p could regulate MYO1B in AML, following the trend of the ceRNA axis.
 |
Figure 6 LncRNA AC024896.1 and miR-363-3p could regulate MYO1B protein in AML. (A) Correlation analysis of AC024896.1, miR-363-3p, and MYO1B. (B) RT-qPCR was used to detect the expression levels of AC024896.1, MYO1B and miR-363-3p after knocking down AC024896.1 with ASO or adding miR-363-3p mimic or inhibitor, respectively. (****P < 0.0001). |
AC024896.1 as a ceRNA of miR-363-3p Regulates MYO1B to Affect the Malignant Progression of AML
The predicted binding sites of AC024896.1 and miR-363-3p were complementarily mutated to determine whether AC024896.1 acts as a ceRNA of miR-363-3p (Figure 7A). Luciferase assays were performed to detect the effect of changes in miR-363-3p before and after mutation on luciferase activity. We found that miR-363-3p regulated the luciferase activity of wild-type AC024896.1, while mutant AC024896.1 was unaffected (Figure 7B). Next, we predicted the enrichment of genes co-regulated by AC024896.1 and miR-363-3p in the malignant progression of AML tumors, and the results showed that the enrichment was particularly significant in the proliferation pathway (Figure 7C–D). The knockdown of AC024896.1 inhibited tumor cell proliferation in control cells but not in mimic cells (Figure 7E–F). Briefly, it appears that AC024896.1 affected AML tumor malignant progression by acting as a ceRNA of miR-363-3p, thus regulating MYO1B.
 |
Figure 7 AC024896.1 as a ceRNA of miR-363-3p regulates MYO1B protein to affect the malignant progression of AML. (A) Bioinformatics analysis demonstrated that miR-363-3p directly targeted the AC024896.1 sequence. (B) Luciferase reporter plasmid containing AC024896.1-WT or AC024896.1-MUT was co-transfected into 293T cells with miR-363-3p mimic. (C) Venn diagram showed the overlapping genes of AC024896.1 groups and miR-363-3p groups. (D) Pathway enrichment of genes co-regulated by AC024896.1 and miR-363-3p. (E–F) The CCK-8 results of THP1 and Kasumi cells with knockdown AC024896.1 treated with NC mimic or miR-363-3p mimic. Data were expressed as the mean ± standard deviation of three independent experiments (n=3).(*P < 0.05, ****P < 0.0001). |
Discussion
Among the four major blood cancers, AML has high malignancy and shows rapid disease development.33 The diagnostic approach for AML typically involves several steps, including a physical exam, molecular testing, and biopsy. Since the treatment of AML needs to fully consider the subtype, age, and overall health of the patient, as well as specific gene mutations, completely effective treatments, are needed.34 Therefore, effective treatment for diagnosing and treating AML is still an important topic in the medical field. Many scientists continue finding new diagnoses and treatment methods. With the development of ncRNA research over the past few years, a vital role of ncRNAs in AML has also been discovered.35 One class of ncRNAs extensively studied in AML is miRNAs. For example, miR-155 is upregulated in AML which results in promoting cell proliferation in leukemia. It also regulates the differentiation and immune responses of myeloid cells.36 miR-181 a/b is downregulated in AML and participates in the differentiation of hematopoietic stem cells.37 It also promotes apoptosis and sensitizes leukemia cells to chemotherapy. In addition to miRNAs, another class of ncRNAs is lncRNAs. For instance, among lncRNAs, Linc-MAF-4 is upregulated in AML, and knocking it down inhibits cell growth in AML.38 miRNAs and lncRNAs can regulate the pathogenesis of various cancers, including AML, through ceRNA. In AML, lncRNA H19 acts as a ceRNA for miR-29b, thus upregulating the oncogene HMGA2 and promoting proliferation and survival.39 Additionally, lncRNA NEAT1 acts as a ceRNA for miR-34a in AML, leading to the upregulation of Bcl-2, an anti-apoptotic protein, and inducing resistance to chemotherapy-induced apoptosis.40 Herein, we constructed the ceRNA regulatory network for AML and fully verified that AC024896.1, as the ceRNA of miR-363-3p, controls its progression. These research results can greatly expand the ceRNA regulatory network of AML and provide a new research direction for its diagnosis and treatment.
Cuproptosis is a type of programmed cell death triggered by copper accumulation in cells. This process is thought to involve the accumulation of copper in lysosomes, which are organelles within cells that break down and recycle cellular waste. Due to the accumulation of copper in lysosomes, reactive oxygen species (ROS) are produced, resulting in oxidative stress and cell death. Cuproptosis has been linked to several different diseases and conditions, including Wilson’s disease, a genetic disorder that causes copper accumulation in the liver and other organs, and cancer. It may contribute to the effectiveness of certain chemotherapeutic treatments. In recent years, cuproptosis has emerged as a potential therapeutic target for cancer treatment since cancer cells are more sensitive to copper-induced cell death than normal cells. However, studies on the role of cuproptosis in AML are limited. Studies have shown that AML cells have higher levels of copper than normal hematopoietic cells.41 Copper chelation or inhibition of copper uptake can induce cell death of AML cells. Copper chelation can sensitize AML cells to chemotherapy and improve treatment outcomes in vivo.42 Herein, according to 13 cuproptosis genes, AML sampled were subjected to consensus clustering. The patients were separated into two groups based on the differential degrees of cuproptosis, and further analysis was conducted to obtain cuproptosis-related genes in AML.
Myosin IB encoded by MYO1B is a member of the myosin superfamily of actin-based motor proteins.43 MYO1B may promote cancer cell proliferation by regulating various signaling pathways that control cell growth and division. Furthermore, MYO1B also promotes cell cycle progression by regulating the expression and localization of the key regulator of G1-S transition and cyclin D1.44 Some evidence suggests that MYO1B may indirectly contribute to cuproptosis by regulating the cellular processes that are affected by copper metabolism. For example, MYO1B regulates the trafficking and recycling of membrane proteins, including CTR1 which is a copper transporter.45 CTR1 is responsible for the uptake of copper into cells and its expression and localization are tightly regulated by various mechanisms of trafficking and recycling.46 MYO1B interacts with CTR1 and regulates its localization and activity, suggesting that MYO1B has a role in copper metabolism. One study indicated that MYO1B is involved in the ceRNA network in hepatocellular carcinoma, where it competes with other RNAs for binding to miR-150-5p.47 A molecular sponge known as Myo1B negatively regulates miR-150-5p expression, leading to increased expression of the miR-150-5p target gene, c-Myb, thus promoting the proliferation of HCC cells.48 However, neither MYO1B nor its role as a link in the ceRNA network has been addressed in AML. Herein, we presented, for the first time, that MYO1B contributes to the malignant progression of AML. The more innovative establishment of ceRNA between MYO1B and ncRNAs in AML provides a solid theoretical basis for its diagnosis and treatment.
In summary, we demonstrated for the first time that AC024896.1, a ceRNA of miR-363-3p, regulates the cuproptosis-related gene MYO1B in AML. Among them, we demonstrated that AC024896.1, miR-363-3p, and MYO1B have regulator effects y on the progression of AML. In addition, AC024896.1 regulates MYO1B expression via miR-363-3p and serves as the molecular sponge of MYO1B. This study revealed a novel ceRNA regulatory network in AML, thus providing new potential targets and a theoretical basis for its diagnosis and treatment.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethical Approval
Our research followed the guidelines outlined by Article 32 of the “Measures for Ethical Review of Human Life Sciences and Medical Research” issued by the National Health Commission of the People’s Republic of China, which exempted the use of legally obtained public data from ethical review.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The research received no external funding.
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Khwaja A, Bjorkholm M, Gale RE, et al. Acute myeloid leukaemia. Nat Rev Dis Primers. 2016;2(1):16010. doi:10.1038/nrdp.2016.10
2. Blackburn LM, Bender S, Brown S. Acute leukemia: diagnosis and treatment. In: Seminars in Oncology Nursing. Elsevier; 2019.
3. Pui CH, Nichols KE, Yang JJ. Somatic and germline genomics in paediatric acute lymphoblastic leukaemia. Nat Rev Clin Oncol. 2019;16(4):227–240. doi:10.1038/s41571-018-0136-6
4. Buschner G, Feuerecker B, Spinner S, et al. Differentiation of acute myeloid leukemia (AML) cells with ATRA reduces 18 F-FDG uptake and increases sensitivity towards ABT-737-induced apoptosis. Leuk Lymphoma. 2021;62(3):630–639. doi:10.1080/10428194.2020.1839648
5. Tamamyan G, Kadia T, Ravandi F, et al. Frontline treatment of acute myeloid leukemia in adults. Crit Rev Oncol Hematol. 2017;110:20–34. doi:10.1016/j.critrevonc.2016.12.004
6. Estey E, Levine RL, Lowenberg B. Current challenges in clinical development of ”targeted therapies”: the case of acute myeloid leukemia. Blood. 2015;125(16):2461–2466. doi:10.1182/blood-2015-01-561373
7. Ge S, Zhu G, Yi Y. Extramedullary plasmacytoma of the larynx: literature review and report of a case who subsequently developed acute myeloid leukemia. Oncol Lett. 2018;16(3):2995–3004. doi:10.3892/ol.2018.8992
8. Kayser S, Levis MJ. Advances in targeted therapy for acute myeloid leukaemia. Br J Haematol. 2018;180(4):484–500. doi:10.1111/bjh.15032
9. Goto H. Childhood relapsed acute lymphoblastic leukemia: biology and recent treatment progress. Pediatr Int. 2015;57(6):1059–1066. doi:10.1111/ped.12837
10. Guo Y, Luo F, Liu Q, et al. Regulatory non-coding RNA s in acute myocardial infarction. J Cell Mol Med. 2017;21(5):1013–1023. doi:10.1111/jcmm.13032
11. Kharel P, Balaratnam S, Beals N, et al. The role of RNA G-quadruplexes in human diseases and therapeutic strategies. Wiley Interdiscip Rev RNA. 2020;11(1):e1568. doi:10.1002/wrna.1568
12. Almamun M, Levinson BT, van Swaay AC, et al. Integrated methylome and transcriptome analysis reveals novel regulatory elements in pediatric acute lymphoblastic leukemia. Epigenetics. 2015;10(9):882–890. doi:10.1080/15592294.2015.1078050
13. Falzone L, Lupo L, Salemi R, et al. Identification of novel microRNAs and their diagnostic and prognostic significance in oral cancer. Cancers. 2019;11(5):610. doi:10.3390/cancers11050610
14. Mesrian Tanha H, Mojtabavi Naeini M, Rahgozar S, et al. Integrative computational in-depth analysis of dysregulated miRNA-mRNA interactions in drug-resistant pediatric acute lymphoblastic leukemia cells: an attempt to obtain new potential gene-miRNA pathways involved in response to treatment. Tumour Biol. 2016;37(6):7861–7872. doi:10.1007/s13277-015-4553-1
15. Trino S, Lamorte D, Caivano A, et al. MicroRNAs as new biomarkers for diagnosis and prognosis, and as potential therapeutic targets in acute myeloid leukemia. Int J Mol Sci. 2018;19(2):460. doi:10.3390/ijms19020460
16. Wang LJ, Qiu B-Q, Yuan -M-M, et al. Identification and validation of dilated cardiomyopathy-related genes via bioinformatics analysis. Int J Gen Med. 2022;15:3663–3676. doi:10.2147/IJGM.S350954
17. Lopez-Pajares V. Long non-coding RNA regulation of gene expression during differentiation. Pflugers Arch. 2016;468(6):971–981. doi:10.1007/s00424-016-1809-6
18. Koscianska E, Kozlowska E, Fiszer A. Regulatory potential of competing endogenous RNAs in myotonic dystrophies. Int J Mol Sci. 2021;22(11):6089. doi:10.3390/ijms22116089
19. Yuan CL, Li H, Zhu L, et al. Aberrant expression of long noncoding RNA PVT1 and its diagnostic and prognostic significance in patients with gastric cancer. Neoplasma. 2016;63(3):442–449. doi:10.4149/314_150825N45
20. Guo Q, Cheng Y, Liang T, et al. Comprehensive analysis of lncRNA-mRNA co-expression patterns identifies immune-associated lncRNA biomarkers in ovarian cancer malignant progression. Sci Rep. 2015;5(1):17683. doi:10.1038/srep17683
21. Zhou M, Zhao H, Xu W, et al. Discovery and validation of immune-associated long non-coding RNA biomarkers associated with clinically molecular subtype and prognosis in diffuse large B cell lymphoma. Mol Cancer. 2017;16(1):16. doi:10.1186/s12943-017-0580-4
22. Chandra Gupta S, Nandan Tripathi Y. Potential of long non-coding RNAs in cancer patients: from biomarkers to therapeutic targets. Int J Cancer. 2017;140(9):1955–1967. doi:10.1002/ijc.30546
23. Sebastiani P, Gurinovich A, Nygaard M, et al. APOE alleles and extreme human longevity. J Gerontol a Biol Sci Med Sci. 2019;74(1):44–51. doi:10.1093/gerona/gly174
24. Chen D, Wang M, Jiang X, et al. Comprehensive analysis of ZFPM2-AS1 prognostic value, immune microenvironment, drug sensitivity, and co-expression network: from gastric adenocarcinoma to pan-cancers. Discov Oncol. 2022;13(1):24. doi:10.1007/s12672-022-00487-0
25. Qu J, Li M, Zhong W, et al. Competing endogenous RNA in cancer: a new pattern of gene expression regulation. International Journal of Clinical and Experimental Medicine. 2015;8(10):17110.
26. Cheng Y, Su Y, Wang S, et al. Identification of circRNA-lncRNA-miRNA-mRNA competitive endogenous RNA network as novel prognostic markers for acute myeloid leukemia. Genes. 2020;11(8):868. doi:10.3390/genes11080868
27. Cobine PA, Brady DCJMC. Cuproptosis: cellular and molecular mechanisms underlying copper-induced cell death. Molecular Cell. 2022;82(10):1786–1787. doi:10.1016/j.molcel.2022.05.001
28. da Silva DA, De Luca A, Squitti R, et al. Copper in tumors and the use of copper-based compounds in cancer treatment. Journal of Inorganic Biochemistry. 2022;226:111634. doi:10.1016/j.jinorgbio.2021.111634
29. Chen YH, Xu N-Z, Hong C, et al. Myo1b promotes tumor progression and angiogenesis by inhibiting autophagic degradation of HIF-1alpha in colorectal cancer. Cell Death Dis. 2022;13(11):939. doi:10.1038/s41419-022-05397-1
30. Wang A, Dai H, Gong Y, et al. ANLN-induced EZH2 upregulation promotes pancreatic cancer progression by mediating miR-218-5p/LASP1 signaling axis. J Exp Clin Cancer Res. 2019;38(1):347. doi:10.1186/s13046-019-1340-7
31. Wen L-J, Hu X-L, Li C-Y, et al. Myosin 1b promotes migration, invasion and glycolysis in cervical cancer via ERK/HIF-1α pathway. American Journal of Translational Research. 2021;13(11):12536.
32. Sharma A, Yadav D, Rao P, et al. Identification of potential therapeutic targets associated with diagnosis and prognosis of colorectal cancer patients based on integrated bioinformatics analysis. Computers in Biology and Medicine. 2022;146:105688. doi:10.1016/j.compbiomed.2022.105688
33. Piddock RE, Bowles KM, Rushworth SA. The role of PI3K isoforms in regulating bone marrow microenvironment signaling focusing on acute myeloid leukemia and multiple myeloma. Cancers. 2017;9(4):29. doi:10.3390/cancers9040029
34. Newell LF, Cook RJJB. Advances in acute myeloid leukemia. BMJ. 2021;375.
35. Pan JQ, Zhang Y-Q, Wang J-H, et al. lncRNA co-expression network model for the prognostic analysis of acute myeloid leukemia. Int J Mol Med. 2017;39(3):663–671. doi:10.3892/ijmm.2017.2888
36. Liang C, Li Y, Wang L-N, et al. Up-regulated miR-155 is associated with poor prognosis in childhood acute lymphoblastic leukemia and promotes cell proliferation targeting ZNF238. Hematology. 2021;26(1):16–25. doi:10.1080/16078454.2020.1860187
37. Zhao TF, Jia H-Z, Zhang -Z-Z, et al. LncRNA H19 regulates ID2 expression through competitive binding to HSA-miR-19a/b in acute myelocytic leukemia. Mol Med Rep. 2017;16(3):3687–3693. doi:10.3892/mmr.2017.7029
38. Uthaya Kumar DB, Williams A. Long non-coding RNAs in immune regulation and their potential as therapeutic targets. Int Immunopharmacol. 2020;81:106279. doi:10.1016/j.intimp.2020.106279
39. Zhao T, Liu XJERMPS. LncRNA-H19 inhibits apoptosis of acute myeloid leukemia cells via targeting miR-29a-3p. European Review for Medical and Pharmacological Sciences. 2019;23(3 Suppl):224–231. doi:10.26355/eurrev_201908_18651
40. Ng M, Heckl D, Klusmann JH. The regulatory roles of long noncoding RNAs in acute myeloid leukemia. Front Oncol. 2019;9:570. doi:10.3389/fonc.2019.00570
41. Hassani S, Ghaffari P, Chahardouli B, et al. Disulfiram/copper causes ROS levels alteration, cell cycle inhibition, and apoptosis in acute myeloid leukaemia cell lines with modulation in the expression of related genes. Biomed Pharmacother. 2018;99:561–569. doi:10.1016/j.biopha.2018.01.109
42. Banerjee K, Das S, Sarkar A, et al. A copper chelate induces apoptosis and overcomes multidrug resistance in T-cell acute lymphoblastic leukemia through redox imbalance and inhibition of EGFR/PI3K/Akt expression. Biomed Pharmacother. 2016;84:71–92. doi:10.1016/j.biopha.2016.08.056
43. Mentes A, Huehn A, Liu X, et al. High-resolution cryo-EM structures of actin-bound myosin states reveal the mechanism of myosin force sensing. Proc Natl Acad Sci U S A. 2018;115(6):1292–1297. doi:10.1073/pnas.1718316115
44. Li SH, Qian L, Chen Y-H, et al. Targeting MYO1B impairs tumorigenesis via inhibiting the SNAI2/cyclin D1 signaling in esophageal squamous cell carcinoma. Journal of Cellular Physiology. 2022;237(9):3671–3686. doi:10.1002/jcp.30831
45. Naughton FB, Becker P, Brotherton D, et al. Substrate binding and conformational changes of the bile acid symporter ASBTNM. Biophys J. 2019;116(3):552a–553a. doi:10.1016/j.bpj.2018.11.2972
46. Wee NK, Weinstein DC, Fraser ST, et al. The mammalian copper transporters CTR1 and CTR2 and their roles in development and disease. Int J Biochem Cell Biol. 2013;45(5):960–963. doi:10.1016/j.biocel.2013.01.018
47. Yamada Y, Koshizuka K, Hanazawa T, et al. Passenger strand of miR-145-3p acts as a tumor-suppressor by targeting MYO1B in head and neck squamous cell carcinoma. Int J Oncol. 2018;52(1):166–178. doi:10.3892/ijo.2017.4190
48. Shimonosono M, Idichi T, Seki N, et al. Molecular pathogenesis of esophageal squamous cell carcinoma: identification of the antitumor effects of miR‑145‑3p on gene regulation. Int J Oncol. 2019;54(2):673–688. doi:10.3892/ijo.2018.4657
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.