")
Back to Journals » OncoTargets and Therapy » Volume 13
Upregulated lncRNA CASC9 Contributes to Progression of Non-Small Cell Lung Cancer Through Inhibition of miR-335-3p and Activation S100A14 Expression
Authors Zhao W, Chen T , Zhao Y
Received 15 February 2020
Accepted for publication 14 May 2020
Published 24 June 2020 Volume 2020:13 Pages 6027—6036
DOI https://doi.org/10.2147/OTT.S249973
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Carlos E Vigil
Weigang Zhao,1,* Tangbing Chen,2,* Yonghong Zhao1
1Department of Thoracic Surgery, Shanghai Jiao Tong University Affiliated Sixth People’s Hospital, Shanghai 200030, People’s Republic of China; 2Department of Thoracic Surgery, Shanghai Chest Hospital, Shanghai Jiao Tong University, Shanghai 200030, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yonghong Zhao
Department of Thoracic Surgery, Shanghai Jiao Tong University Affiliated Sixth People’s Hospital, 600 Yishanlu Road, Xuhui District, Shanghai 200030, People’s Republic of China
Email [email protected]
Introduction: Non-small cell lung cancer (NSCLC) is a deadly cancer type worldwide and the main sub-type of lung cancer. Cancer susceptibility candidate-9 (CASC9) was reported to be a key player in cancer progression. However, its function and underlying mechanism in NSCLC remain unclear.
Materials and Methods: Expression level of CASC9 in NSCLC tissues and cells was measured with RT-qPCR. Biological roles of CASC9 in NSCLC were analyzed with a series of in vitro experiments. Potential mechanisms of CASC9 in NSCLC were analyzed by predicting and validating the possible targets of CASC9 in NSCLC.
Results: In this study, we found CASC9 expression was upregulated in NSCLC tissues and cell lines. High CASC9 expression was identified as a predictor for poorer overall survival of NSCLC patients. Furthermore, functional assays showed CASC9 knockdown suppressed NSCLC cell proliferation, migration, and invasion, while CASC9 overexpression caused opposite effects. We also found microRNA-335-3p (miR-335-3p) could act as a target of CASC9 in NSCLC and the inhibition effect of CASC9 knockdown on NSCLC progression required the activity of miR-335-3p. In addition, we identified S100 calcium-binding protein A14 (S100A14) acts as a target of miR-335-3p.
Discussion: Taken together, our study suggested CASC9 could promote NSCLC progression via miR-335-3p/S100A14 axis. The CASC9/miR-335-3p/S100A14 regulatory triplets identified in this work might provide new therapeutic strategies for NSCLC treatment.
Keywords: non-small cell lung cancer, CASC9, miR-335-3p, S100A14, ceRNA
Introduction
Non-small cell lung cancer (NSCLC) represents about 85% of all lung cancer cases.1 NSCLC patients were often diagnosed at late tumor stages, which results in poorer overall survival of cancer patients.2
Long non-coding RNA (lncRNA, above 200 nucleotides in length) is regarded as crucial regulators for cancer expression in recent years.3,4 LncRNA cancer susceptibility candidate-9 (CASC9), located at chromosome 8q21.11, was found closely associated with cancer progression. In colorectal cancer, CASC9 expression was found elevated in tumor tissues and correlated with poorer overall survival.5 Functionally, CASC9 knockdown inhibits colorectal cancer growth through regulating cleavage and polyadenylation specificity factor subunit 3 and transforming growth factor-β signaling pathway.5 Zhang et al reported high CASC9 level was a predictor for poorer overall survival of patients with cervical cancer.6 Hu and co-authors reported CASC9 could promote ovarian cancer progression in vitro and in vivo through upregulating LIN7A expression.7 Moreover, a previous study revealed CASC9 could promote esophageal squamous cell carcinoma metastasis through binding with CREB-binding protein to promote LAMC2 expression.8 However, the expression and regulatory mechanism of CASC9 in NSCLC remains to be explored.
S100 calcium-binding protein A14 (S100A14) is a member of S100 protein family and consisted of 104 amino acids.9 Meanwhile, the gene that encodes S100A14 protein was mapped to chromosome 1q21.9 Several studies have conducted to analyze the roles of S100A14 in cancers and revealed S100A14 functions as an oncogenic role in lung adenocarcinoma and serous ovarian carcinoma.10–12 However, the molecules that can regulate S100A14 in NSCLC remain to be explored.
Our study aimed to investigate CASC9 levels in NSCLC tissues and cell lines. Prognostic value of CASC9 on NSCLC patients was analyzed with Kaplan-Meier curve. Effects of CASC9 on NSCLC cell proliferation, migration, and invasion were explored in vitro. Moreover, we further investigated underlying mechanisms by which CASC9 regulates NSCLC progression.
Materials and Methods
Patients
43 NSCLC patients who did not receive anti-cancer treatment were enrolled in this work. Study protocol was approved by the ethics committee of Shanghai Jiao Tong University Affiliated Sixth People’s Hospital. Tumor tissues and normal tissues obtained from these patients were snap-frozen in liquid nitrogen and stored at −80°C for further usage. Informed written consent was obtained from all patients. This work was performed in compliance with Declaration of Helsinki. Clinicopathological data including age, gender, tobacco smoking status, alcohol drinking status, tumor stage and tumor size are summarized in Table 1.
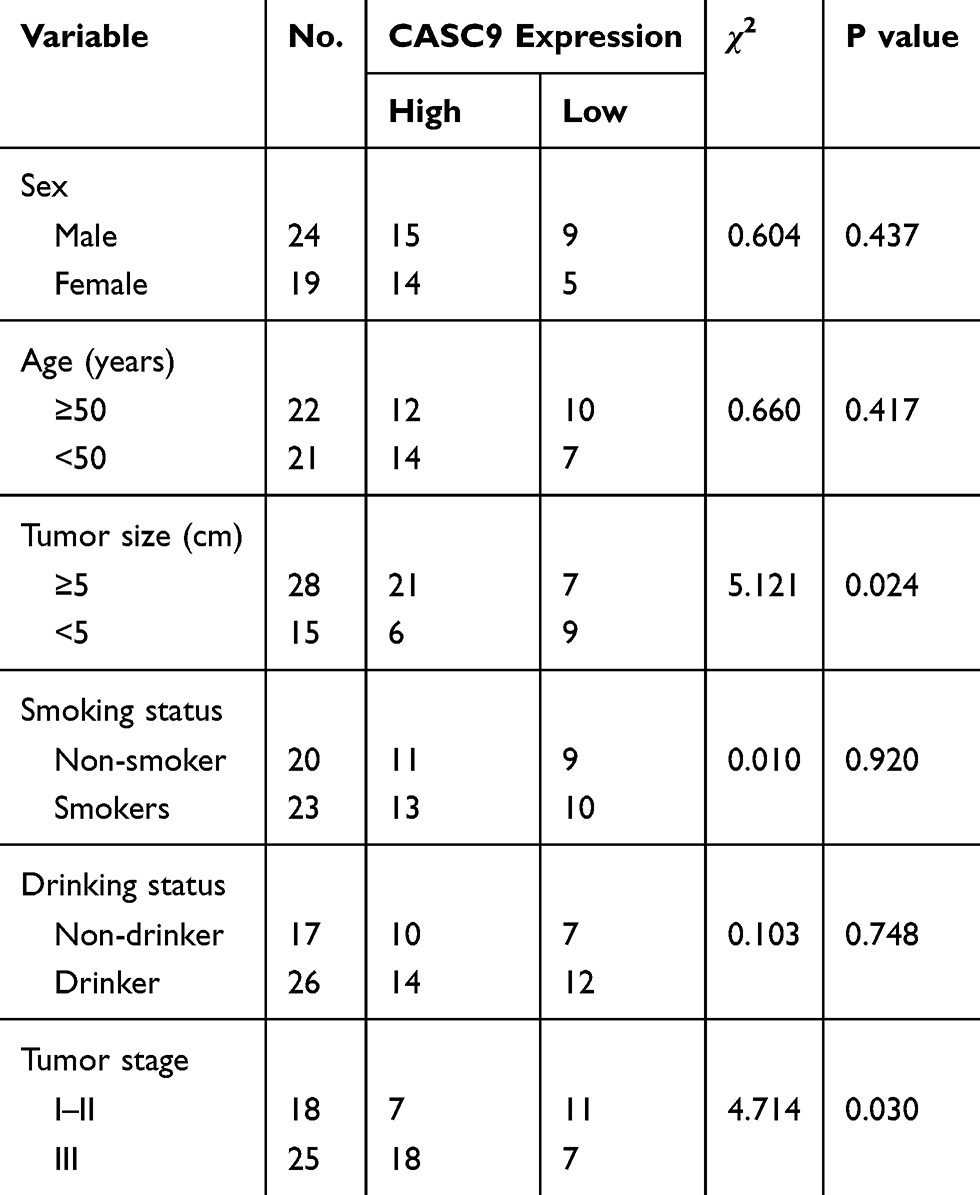 |
Table 1 Correlations of CASC9 Expression and Clinicopathological Features of NSCLC Patients |
Cell Lines
NSCLC cells (A549, H1299) and lung epithelial cell line (BEAS-2B) were obtained from American Type Culture Collection (ATCC). These cells were incubated in DMEM contains 10% fetal bovine serum (FBS, Gibco, Thermo Fisher Scientific, Inc., Waltham, MA, USA) at a 37°C humidified incubator with 5% CO2.
Cell Transfection
pcDNA3.1 contains sequence of CASC9 or S100A14 expression vector (pCASC9 or pS100A14) was constructed by GenScript (Nanjing, Jiangsu, China). miR-335-3p mimic (5ʹ-UUUUUCAUUAUUGCUCCUGACC-3ʹ) and corresponding negative control (miR-NC, 5ʹ-GUCUCAUCGUCCUUAUUCUUAU-3ʹ) were purchased from GenePharma (Shanghai, China). Small interfering RNA targeting CASC9 (si-CASC9, 5ʹ-CAGCAAAGCAAUGGAAGCAUGUAUU-3ʹ) and siR-NC (5ʹ-UUCUCCGAACGUGUCACGUTT-3ʹ) were also bought from GenePharma. Cell transfection was conducted using Lipofectamine 2000 (Invitrogen, Thermo Fisher Scientific, Inc.) after incubation these cells to about 80% confluence. Cells were collected for following analyses after 48 h transfection.
RNA Extraction and Quantitative Real-Time PCR (RT-qPCR)
RNA was isolated from tissues and cells with Trizol reagent (Invitrogen). After concentration quantification using NanoDrop-1000 (Thermo Fisher Scientific, Inc.), RNA was reverse transcribed into complementary DNA using Reverse Transcription kit (Takara, Dalian, Liaoning, China). RT-qPCR was performed at ABI 7500 system (Applied Biosystems, Foster City, CA, USA) using SYBR Green (Takara) with primers: CASC9: F: 5ʹ-TTGGTCAGCCACATTCATGGT-3ʹ, R: 5ʹ-AGTGCCAATGACTCTCCAGC-3ʹ; S100A14: F: 5ʹ-GTCGGTCAGCCAACGCAGAG-3ʹ, R: 5ʹ-CAGGCCACAGTTGCTCGG-3ʹ; MMP-2: F:5ʹ-TGATTCTGGTCGCTCAGATG-3ʹ, R: 5ʹ-CTTGTTTCCCAGGAAGGTGA-3ʹ; MMP-9:F: 5ʹ-AGGACCATGGGGATCCTTAC-3ʹ, R: 5ʹ-AACACAAGGCTGCCCATTAC-3ʹ; E-Cadherin: F: 5ʹ-TAACCGATCAGAATGAC-3ʹ, R: 5ʹ-TTTGTCAGGGAGCTCAGGAT-3ʹ; N‐Cadherin: F: 5ʹ-CAACTTGCCAGAAAACTCCAGG-3ʹ, R: 5ʹ-ATGAAACCGGGCTATCTGCTC-3ʹ; GAPDH: F: 5ʹ-GGGAGCCAAAAGGGTCATCA-3ʹ, R: 5ʹ-TGATGGCATGGACTGTGGTC-3ʹ; miR-335-3p: F: 5ʹ-UUUUUCAUUAUUGCUCCUGACC-3ʹ, R: 5ʹ-CCAGTCTCAGGGTCCGAGGTATTC-3ʹ; U6 snRNA: F: 5ʹ‐CTCGCTTCGGCAGCACA‐3ʹ, R: 5ʹ‐AACGCTTCACGAATTTGCGT‐3ʹ. Relative expression level was calculated using 2−ΔΔCt method.
Western Blot
Tissues and cells were collected and lysed with RIPA buffer to extract protein samples. BCA kit (Beyotime) was used to quantify protein concentration. Then, protein samples were isolated at SDS-PAGE, transferred to PVDF membrane, and treated with anti-S100A14 (ab251779, Abcam, Cambridge, MA, USA) and anti-GAPDH (ab181602, Abcam) at 4°C for overnight. Then, secondary antibody (ab6721) was added and incubated for 4 h. Band signals were visualized using BeyoECL kit and analyzed with Image J software.
Immunohistochemistry (IHC) Analysis of S100A14 in NSCLC Tissues
Protein levels of S100A14 in NSCLC tissues and normal tissues analyzed with the IHC method were analyzed at Human Protein Atlas.
Cell Proliferation Assay
Cell Counting Kit-8 (CCK-8, Beyotime, Haimen, Jiangsu, China) assay was used to measure cell viability. Cells were seeded into 96-well plate at the density of 5 × 103 cells/well and incubated for 0, 24, 48, and 72 hrs. At these time points, CCK-8 reagent was added to each well and incubated for another 4 hrs. Microplate reader was used to measure optical density at 450 nm.
Wound Healing Assay
Cell surface was scratched with pipette tip after incubation of these cells in serum-free medium to 90% confluence. PBS was used to remove cell debris, and then observed under a microscope at 0 hrs and 24 hrs after wound creation.
Transwell Invasion Assay
DMEM was used to resuspend collected cells and then filled into upper chamber of Matrigel-coated insert (BD Bioscience, San Jose, CA, USA), while DMEM contains 10% FBS was filled into lower chamber. 48 hrs later, non-invasive cells were removed, while invasive cells were fixed by methanol, stained with crystal violet, and counted under microscope.
Bioinformatic Analysis
miRNA targets of CASC9 were predicted at DIANA LncBase v_2, and we found miR-335-3p was a putative target. Subsequently, TargetScan was used to predict targets of miR-335-3p and revealed S1000A14 was a potential target.
Luciferase Activity Reporter Assay
Wild-type (wt) sequence of CASC9 or S100A14 contains a binding region for miR-335-3p was inserted into psi-CHECK-2 (Promega. Madison, WI, USA) to generate wt-CASC9 or wt-S100A14. Mutant luciferase vectors (mt-CASC9 or mt-S100A14) were constructed using site-direct mutagenesis kit (Takara) to lose miR-335-3p binding sites. Cells were co-transfected with luciferase vectors and miRNAs using Lipofectamine 2000. 48 hrs later, dual-luciferase activity system (Promega) was utilized to measure relative luciferase activity using Renilla luciferase as internal control.
RNA Immunoprecipitation (RIP) Assay
RIP assay was performed using Magna RIP Kit (EMD Millipore, Billerica, MA, USA) to analyze the co-enrichment of CASC9, miR-335-3p, and S100A14. Cells were lysed using RIP buffer to obtain cell pellets. Then, anti-Ago2 antibody-conjugated magnetic beads were used to incubate with these pellets. RNA sample was isolated from these pellets after treatment with Proteinase K and RNase-free DNase. qRT-PCR was performed to detect CASC9, miR-335-3p, and S100A14 expression.
Statistical Analysis
Results obtained from three independent experiments were analyzed with GraphPad Prism (GraphPad Prism Software Inc, San Diego, CA, USA) and then expressed as mean ± SD. Student’s t-test and one-way analysis of variance with Tukey post-hoc test were used for difference analyses. Kaplan-Meier curve with Log-rank test was used to analyze the effect of CASC9 on overall survival of NSCLC patients. Pearson’s coefficient was used to analyze correlations of miR-335-3p with CASC9 or S100A14. Chi-square test was used to analyze the correlation of CASC9 expression and clinicopathological parameters. P value of less than 0.05 was regarded as statistically significant.
Results
CASC9 Expression Level Was Elevated in NSCLC
Firstly, we found CASC9 was highly expressed in NSCLC tissues compared with normal tissues (Figure 1A). Using the median expression level of CASC9 as cut-off value, these patients were classified into high or low groups. Kaplan-Meier curve showed high CASC9 expression was a predictor for poorer overall survival of cancer patients (Figure 1B). Then, we analyzed the correlations of CASC9 expression and clinicopathological parameters. It was found the high CASC9 expression was significantly correlated with tumor size and tumor stage, while it did not have correlations with other collected clinicopathological parameters (Table 1). In addition, we showed CASC9 expression level was elevated in NSCLC cells compared with normal cell (Figure 1C).
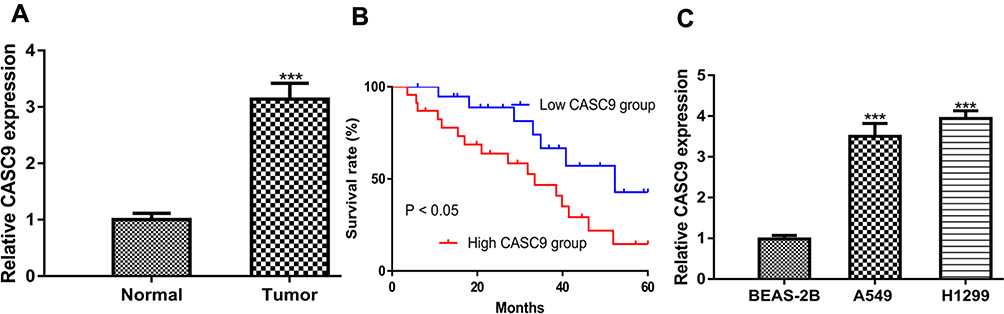 |
Figure 1 CASC9 was elevated expression in NSCLC. Notes: (A) RT-qPCR results showed CASC9 was upregulated in NSCLC tissues compared with normal tissues. (B) Kaplan-Meier curve showed that high CASC9 expression was a predictor for poor overall survival of cancer patients. (C) RT-qPCR results showed CASC9 was upregulated in NSCLC cell lines compared with normal cell line. ***P < 0.001. Abbreviations: RT-qPCR, quantitative real-time PCR; NSCLC, non-small cell lung cancer; CASC9, cancer susceptibility candidate-9. |
CASC9 Overexpression Promotes NSCLC Cell Proliferation, Migration, and Invasion
To explore the roles of CASC9 in NSCLC, gain-of-function experiments were performed at first. The results showed pCASC9 transfection significantly increased CASC9 level in NSCLC cells (Figure 2A). CCK-8 assay showed CASC9 overexpression promotes NSCLC cell proliferation (Figure 2B). Additionally, wound-healing assay and transwell invasion assay showed CASC9 overexpression promoted NSCLC cell migration and invasion abilities compared with control groups (Figure 2C and D). Analysis of the related markers including MMP-2, MMP-9, E-Cadherin, and N-Cadherin in these groups showed that MMP-2, MMP-9, N-Cadherin expressions were increased, while E-Cadherin expression was decreased by pCASC9 (Figure 2E).
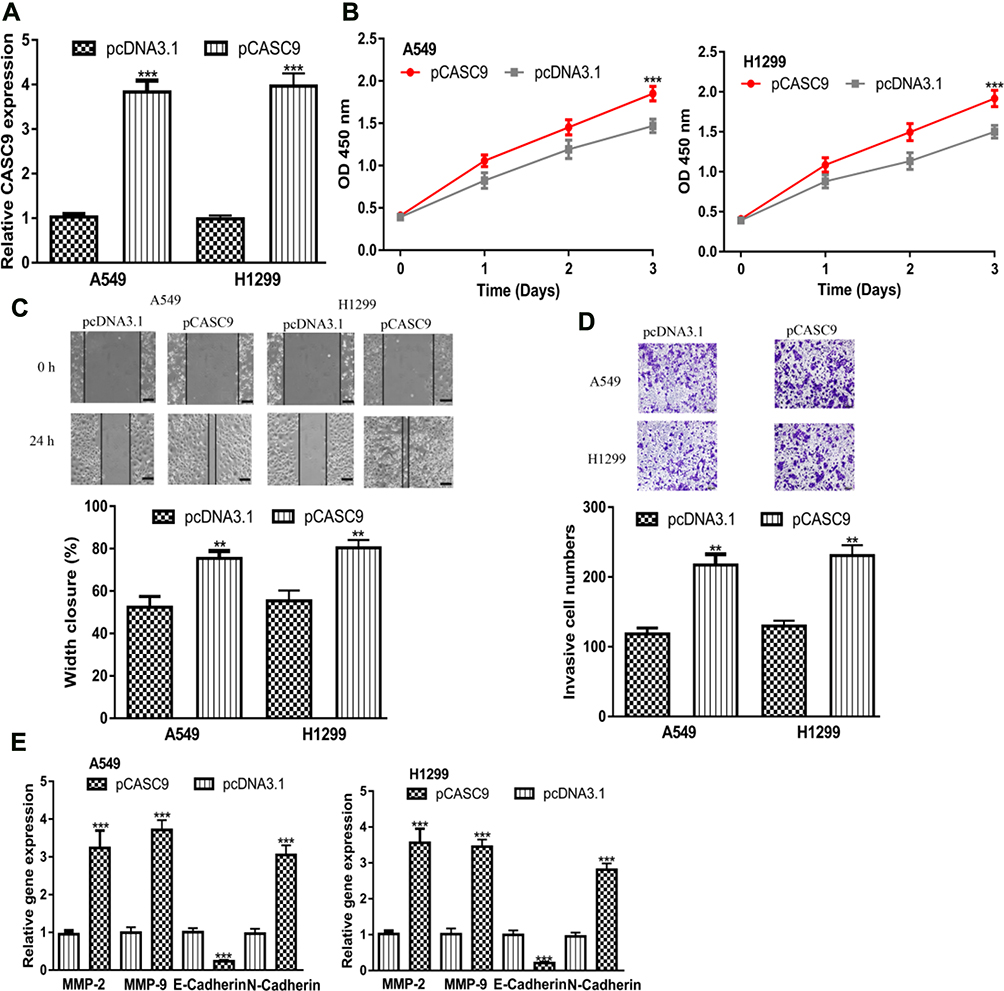 |
Figure 2 CASC9 overexpression promotes NSCLC cell proliferation, migration, and invasion. Notes: (A) RT-qPCR results showed CASC9 was upregulated in NSCLC cell lines after pCASC9 transfection. (B) Cell counting kit-8 assay showed that CASC9 overexpression accelerates NSCLC cell proliferation. (C) Wound-healing assay showed CASC9 overexpression increased cell migration ability. (D) Transwell assay showed pCASC9 transfection enhanced invasion ability. (E) Expression of MMP-2, MMP-9, N-Cadherin, and E-Cadherin in NSCLC cells after pCASC9 transfection. ***P < 0.001; **P < 0.01. Abbreviations: RT-qPCR, quantitative real-time PCR; NSCLC, non-small cell lung cancer; CASC9, cancer susceptibility candidate-9. |
CASC9 Knockdown Inhibits NSCLC Cell Proliferation, Migration, and Invasion
To further investigate the functions of CASC9, we knockdown its expression in NSCLC cell using si-CASC9 (Figure 3A). CCK-8 assay demonstrated CASC9 knockdown inhibits NSCLC cell growth (Figure 3B). Moreover, the introduction of si-CASC9 hinders NSCLC cell migration and invasion abilities (Figure 3C and D). After the knockdown of CASC9, MMP-2, MMP-9, N-Cadherin expressions were decreased, while E-Cadherin expression was increased in NSCLC cells (Figure 3E).
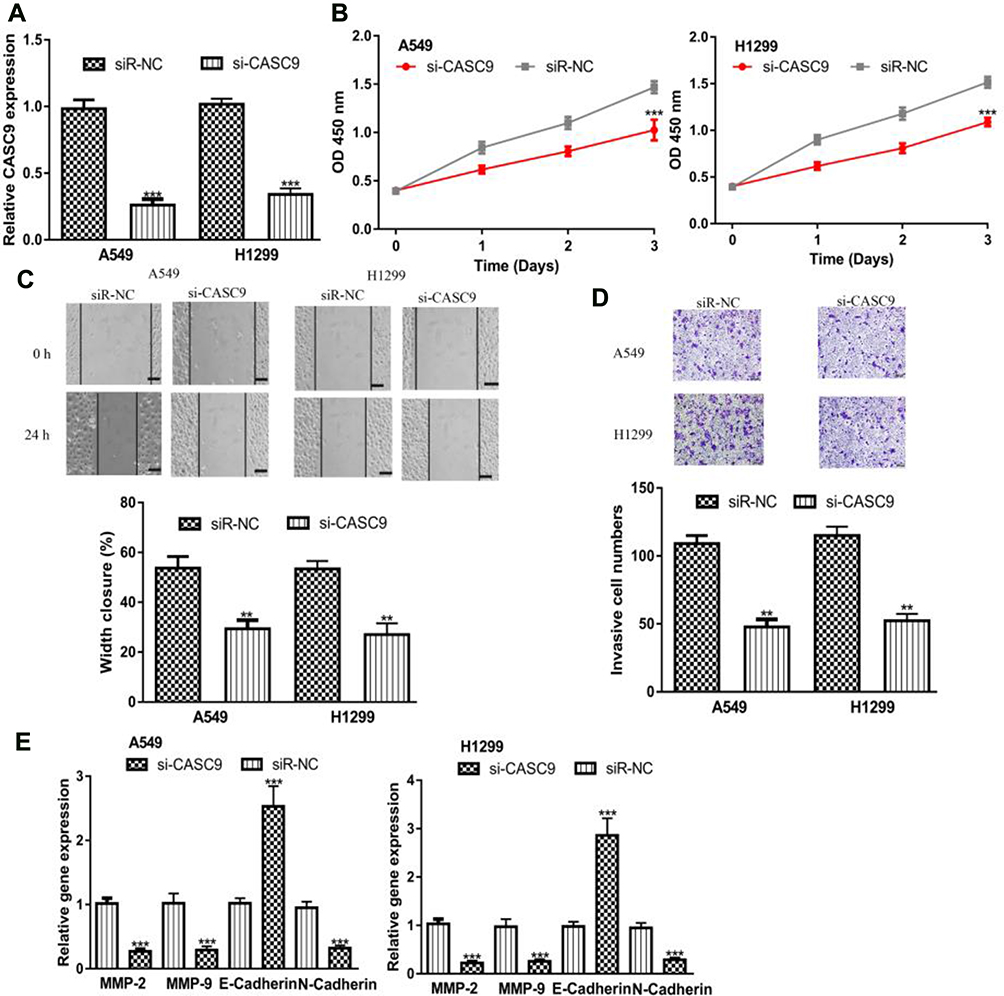 |
Figure 3 CASC9 knockdown inhibits NSCLC cell proliferation, migration, and invasion. Notes: (A) RT-qPCR results showed CASC9 was downregulated in NSCLC cell lines after si-CASC9 transfection. (B) Cell counting kit-8 assay showed that CASC9 downregulation inhibits NSCLC cell proliferation. (C) Wound-healing assay showed CASC9 downregulation decreased cell migration ability. (D) Transwell assay showed si-CASC9 transfection decreased invasion ability. (E) Expression of MMP-2, MMP-9, N-Cadherin, and E-Cadherin in NSCLC cells after si-CASC9 transfection. ***P < 0.001; **P < 0.01. Abbreviations: RT-qPCR, quantitative real-time PCR; NSCLC, non-small cell lung cancer; CASC9, cancer susceptibility candidate-9; si-CASC9, small interfering RNA targeting cancer susceptibility candidate-9; siR-NC, negative control small interfering RNA. |
CASC9 Directly Regulate miR-335-3p Expression
Then, we analyzed mechanisms of CASC9 in NSCLC, and we showed miR-335-3p was a putative target for CASC9 (Figure 4A). RT-qPCR showed transfection of miR-335-3p mimic decreases CASC9 levels in NSCLC cells (Figure 4B). Dual-luciferase activity reporter assay validated interactions between CASC9 and miR-335-3p (Figure 4C). Moreover, we showed miR-335-3p expression was decreased in NSCLC tumor tissues and cell lines compared with normal counterparts (Figure 4D and E). Importantly, we showed CASC9 and miR-335-3p were inversely correlated in NSCLC tissues (Figure 4F).
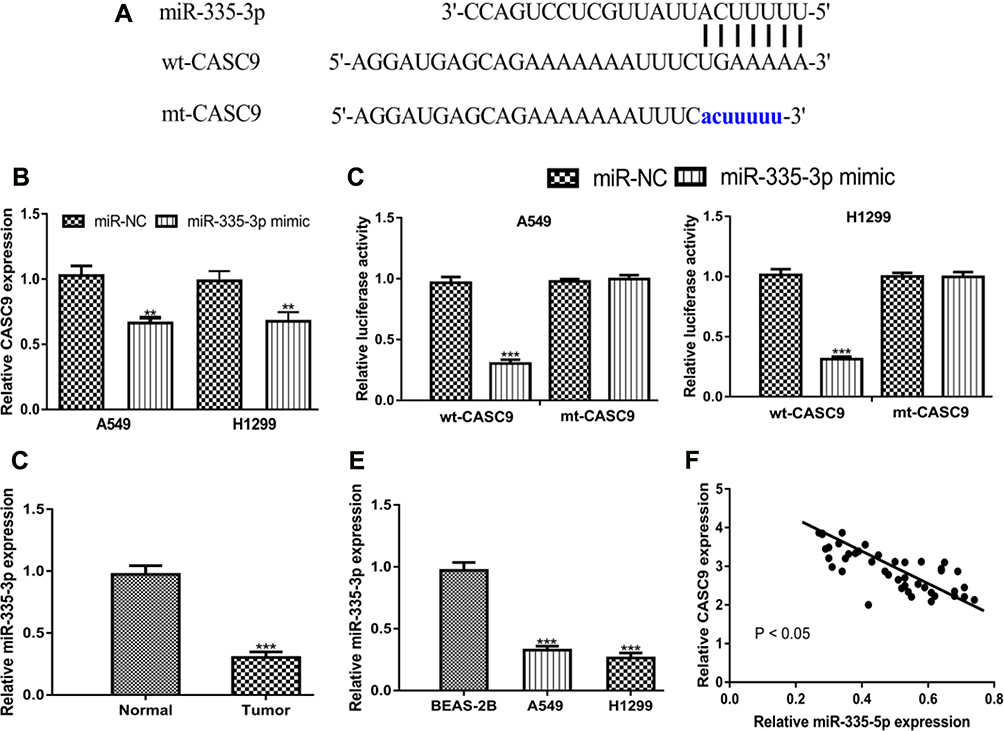 |
Figure 4 CASC9 acts as competing endogenous RNA for miR‐335-5p. Notes: (A) Binding module between CASC9 and miR-335-3p. (B) CASC9 expression in NSCLC cells with synthetic miRNAs transfection. (C) Relative luciferase activity in NSCLC cells with synthetic miRNAs and luciferase activity vectors transfection. RT-qPCR results showed miR-335-3p was downregulated in NSCLC (D) tissues and (E) cell lines. (F) Correlation of CASC9 and miR-335-3p in NSCLC tissues. ***P < 0.001; **P < 0.01. Abbreviations: RT-qPCR, quantitative real-time PCR; NSCLC, non-small cell lung cancer; CASC9, cancer susceptibility candidate-9; miR-335-3p, microRNA-335-5p; miR-NC, negative control microRNA; wt, wild-type; mt, mutant. |
S100A14 Was the Direct Target of miR-335-3p
Afterward, we explored targets of miR-335-3p using TargetScan. We found S100A14 ranks top among all the predicted targets and therefore was selected for analyses (Figure 5A). RT-qPCR indicated miR-335-3p overexpression decreased S100A14 levels in NSCLC cells (Figure 5B). Western blot indicated S100A14 protein levels were also suppressed by miR-335-3p mimic (Figure 5C). To further confirm the connection of miR-335-3p and S100A14, dual-luciferase activity reporter assay was performed. We showed miR-335-3p mimic inhibited luciferase activity in cells transfected with wt-S100A14 (Figure 5D). RIP assay further indicated that CASC9, miR-335-3p, and S100A14 were co-enriched by anti-Ago2 (Figure 5E). The analysis of S100A14 expression in NSCLC tissues and cells using qRT-PCR indicated S100A14 expression was elevated in tumor tissues and cell lines compared with normal groups (Figure 5F and G). Western blot showed S100A14 protein levels were also increased in both NSCLC tissues and cells investigated (Figure 5H and I). We further analyzed the protein level of S100A14 in tumor tissues using IHC assay and confirmed S100A14 was increased expression in NSCLC tissues (Figure 5J). Moreover, we found a significant inversely relationship between miR-335-3p and S100A14 in tumor tissues (Figure 5K).
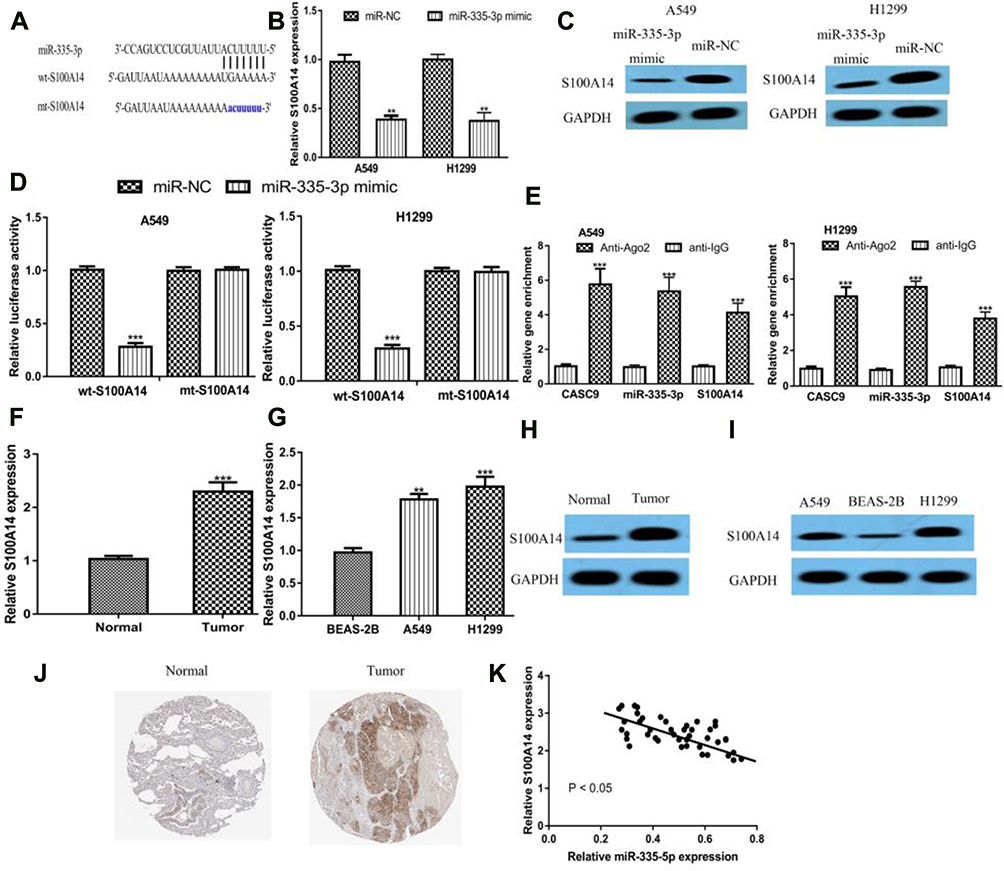 |
Figure 5 S100A14 was the direct target for miR‐335-5p. Notes: (A) Binding module between S100A14 and miR-335-3p. (B) S100A14 expression in NSCLC cells with synthetic miRNAs transfection. (C) S100A14 protein expression in NSCLC cells with synthetic miRNAs transfection. (D) Relative luciferase activity in NSCLC cells with synthetic miRNAs and luciferase activity vectors transfection. (E) RIP assay revealed CASC9, miR-335-3p, and S100A14 were co-enriched in anti-Ago2 pellets. RT-qPCR results showed S100A14 was increased in NSCLC (F) tissues and (G) cell lines. Western blot to analyze S100A14 protein expression in NSCLC (H) tissues and (I) cell lines. (J) IHC assay to detect S100A14 expression in NSCLC tissues and normal tissues. (K) Correlation of S100A14 and miR-335-3p in NSCLC tissues. ***P < 0.001; **P < 0.01. Abbreviations: RT-qPCR, quantitative real-time PCR; NSCLC, non-small cell lung cancer; S100A14, S100 calcium binding protein A14; miR-335-3p, microRNA-335-5p; miR-NC, negative control microRNA; wt, wild-type; mt, mutant. |
CASC9 Promotes NSCLC Progression via S100A14
At length, we investigated whether S100A14 was responsible for the roles of CASC9 in NSCLC through co-transfecting of si-CASC9 and pS100A14. We showed S100A14 level could be elevated by pS100A14 and inhibited by si-CASC9 (Figure 6A). S100A14 overexpression promoted NSCLC cell proliferation, migration, and invasion (Figure 6B–D). We also showed restoration of S100A14 partially reversed the effects of si-CASC9 on NSCLC progression (Figure 6B–D).
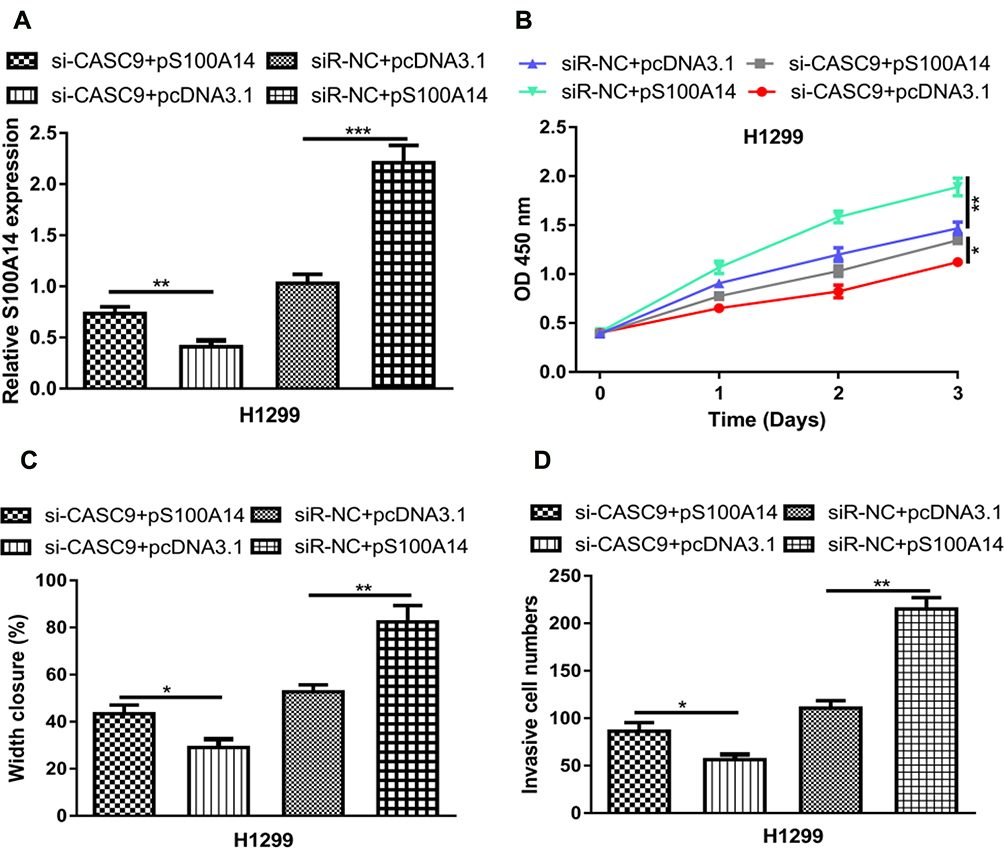 |
Figure 6 CASC9 promotes NSCLC progression through S100A14. Notes: (A) RT‐qPCR to detect S100A14 expression after si-CASC9+pcDNA3.1, si-CASC9+pS100A14, siR-NC+pcDNA3.1, or siR-NC+pS100A14 transfection. (B) Cell counting kit‐8 assay showed that S100A14 overexpression accelerates NSCLC cell proliferation. (C) Wound-healing assay showed S100A14 overexpression increased cell migration ability. (D) Transwell assay showed pS100A14 transfection enhanced invasion ability. ***P < 0.001; **P < 0.01; *P < 0.05. Abbreviations: RT-qPCR, quantitative real-time PCR; NSCLC, non-small cell lung cancer; CASC9, cancer susceptibility candidate-9; S100A14, S100 calcium-binding protein A14; si-CASC9, small interfering RNA targeting cancer susceptibility candidate-9; siR-NC, negative control small interfering RNA. |
Discussion
lncRNAs have been revealed to play crucial roles in contributing NSCLC progression, however only few of them have been functionally characterized.4 For instance, Li et al showed LINC00968 was elevated expression in NSCLC.13 Xia et al showed lncRNA PLAC2 was decreased expression in NSCLC tissues and correlated with poorer overall survival of cancer patients.14 Also, they reported force PLAC2 expression could inhibit NSCLC cell migration and invasion via targeting miR-21.14 Another work showed knockdown of cancer susceptibility 15 could inhibit NSCLC progression via regulating epithelial-mesenchymal transition associated molecules.15
In our work, we showed CASC9 level was elevated in both tumor tissues and cell lines. Also, we showed high CASC9 expression was an indicator for poorer overall survival of NSCLC patients, suggesting CASC9 might be a prognostic biomarker. Through a series of in vitro experiments, we showed CASC9 could promote NSCLC cell proliferation, migration, and invasion. By analyzing the epithelial‐mesenchymal transition (EMT) markers and two matrix metalloproteinases (MMPs), we showed MMP-2, MMP-9, N-Cadherin expression can be increased, while E-Cadherin was decreased by CASC9 overexpression. These results suggested CASC9 might function as an oncogenic lncRNA in NSCLC.
lncRNA-miRNA-mRNA regulatory triplets proposed by Salmena have been well recognized.2,16 For instance, CASC9 functions as competing endogenous RNA to regulate TWIST2 expression through sponging miR-215 in breast cancer.6 According to bioinformatic analysis results and luciferase activity reporter assay, we identified miR-335-3p as a target of CASC9. Through RT-qPCR analysis, we showed miR-335-3p mimic transfection could decrease CASC9 expression level in NSCLC cells. miR-335-3p has reported decreased expression in childhood acute lymphoblastic leukemia, and correlated with poor overall survival of cancer patients.17 Also, miR-335-3p was revealed can be regulated by lncRNAs EAT1 and MALAT1 to affect cancer progression and drug resistant.17 Moreover, miR-335-3p was reported could be regulated by LINC00969 to affect intervertebral disk degeneration.18 We further investigated the targets of miR-335-3p using TargetScan. We revealed miR-335-3p may directly target S100A14, an oncogene in lung adenocarcinoma.10,11 Additionally, we showed S100A14 was increased expression in NSCLC tissues and cell lines using RT-qPCR analysis, Western blot, and IHC assay. RIP assay further confirmed the connections of CASC9, miR-335-3p, and S100A14. Functionally experiments showed CASC9 could promote NSCLC malignant behaviors via regulating S100A14.
Conclusion
In conclusion, our work showed CASC9 functions as an oncogenic lncRNA in NSCLC through regulating miR-335-3p/S100A14 axis. Our work may provide novel therapeutic targets for NSCLC treatment.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Chheang S, Brown K. Lung cancer staging: clinical and radiologic perspectives. Semin Intervent Radiol. 2013;30(2):99–113. doi:10.1055/s-0033-1342950
3. Zhao X, Tang DY, Zuo X, Zhang TD, Wang C. Identification of lncRNA-miRNA-mRNA regulatory network associated with epithelial ovarian cancer cisplatin-resistant. J Cell Physiol. 2019;234(11):19886–19894. doi:10.1002/jcp.28587
4. Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77(15):3965–3981. doi:10.1158/0008-5472.CAN-16-2634
5. Luo K, Geng J, Zhang Q, et al. LncRNA CASC9 interacts with CPSF3 to regulate TGF-β signaling in colorectal cancer. J Exp Clin Cancer Res. 2019;38(1):249. doi:10.1186/s13046-019-1263-3
6. Zhang J, Wang Q, Quan Z. Long non-coding RNA CASC9 enhances breast cancer progression by promoting metastasis through the meditation of miR-215/TWIST2 signaling associated with TGF-β expression. Biochem Biophys Res Commun. 2019;515(4):644–650. doi:10.1016/j.bbrc.2019.05.080
7. Hu X, Li Y, Kong D, Hu L, Liu D, Wu J. Long noncoding RNA CASC9 promotes LIN7A expression via miR-758-3p to facilitate the malignancy of ovarian cancer. J Cell Physiol. 2019;234(7):10800–10808. doi:10.1002/jcp.27903
8. Liang Y, Chen X, Wu Y, et al. LncRNA CASC9 promotes esophageal squamous cell carcinoma metastasis through upregulating LAMC2 expression by interacting with the CREB-binding protein. Cell Death Differ. 2018;25(11):1980–1995. doi:10.1038/s41418-018-0084-9
9. Pietas A, Schlüns K, Marenholz I, Schäfer BW, Heizmann CW, Petersen I. Molecular cloning and characterization of the human S100A14 gene encoding a novel member of the S100 family. Genomics. 2002;79(4):513–522. doi:10.1006/geno.2002.6744
10. Katono K, Sato Y, Kobayashi M, et al. Clinicopathological significance of S100A14 expression in lung adenocarcinoma. Oncol Res Treat. 2017;40(10):594–602. doi:10.1159/000478100
11. Ding F, Wang D, Li XK, et al. Overexpression of S100A14 contributes to malignant progression and predicts poor prognosis of lung adenocarcinoma. Thorac Cancer. 2018;9(7):827–835. doi:10.1111/1759-7714.12654
12. Qian J, Ding F, Luo A, Liu Z, Cui Z. Overexpression of S100A14 in human serous ovarian carcinoma. Oncol Lett. 2016;11(2):1113–1119. doi:10.3892/ol.2015.3984
13. Li DY, Chen WJ, Luo L, et al. Prospective lncRNA-miRNA-mRNA regulatory network of long non-coding RNA LINC00968 in non-small cell lung cancer A549 cells: a miRNA microarray and bioinformatics investigation. Int J Mol Med. 2017;40(6):1895–1906. doi:10.3892/ijmm.2017.3187
14. Xia H, Xiu M, Gao J, Jing H. LncRNA PLAC 2 downregulated miR-21 in non-small cell lung cancer and predicted survival. BMC Pulm Med. 2019;19(1):172. doi:10.1186/s12890-019-0931-6
15. Li M, Chen Y, Zhu J, Gao Z, Wang T, Zhou P. Long noncoding RNA CASC15 predicts unfavorable prognosis and exerts oncogenic functions in non-small cell lung cancer. Am J Transl Res. 2019;11(7):4303–4314.
16. Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the rosetta stone of a hidden RNA language? Cell. 2011;146(3):353–358. doi:10.1016/j.cell.2011.07.014
17. Pouyanrad S, Rahgozar S, Ghodousi ES. Dysregulation of miR-335-3p, targeted by NEAT1 and MALAT1 long non-coding RNAs, is associated with poor prognosis in childhood acute lymphoblastic leukemia. Gene. 2019;692:35–43. doi:10.1016/j.gene.2019.01.003
18. Yu L, Hao Y, Xu C, Zhu G, Cai Y. LINC00969 promotes the degeneration of intervertebral disk by sponging miR-335-3p and regulating NLRP3 inflammasome activation. IUBMB Life. 2019;71(5):611–618. doi:10.1002/iub.1989
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.