")
Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 16
Transcriptome Analysis Identifies Biomarkers for the Diagnosis and Management of Psoriasis Complicated with Depression
Authors Xia X , Yu H, Li Y , Liang Y , Li G , Huang F
Received 6 April 2023
Accepted for publication 13 May 2023
Published 18 May 2023 Volume 2023:16 Pages 1287—1301
DOI https://doi.org/10.2147/CCID.S413887
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Jeffrey Weinberg
Xichun Xia,1,2,* Hai Yu,3,* Yanxiang Li,2 Yunting Liang,1 Guangqiang Li,2 Fang Huang1
1Department of Dermatology, Zhuhai People’s Hospital (Zhuhai Hospital Affiliated with Jinan University), Jinan University, Zhuhai, 519050, People’s Republic of China; 2The Biomedical Translational Research Institute, Health Science Center (School of Medicine), Jinan University, Guangzhou, 510632, People’s Republic of China; 3Department of Dermatology, The First Affiliated Hospital of Jinan University, Guangzhou, Guangdong, 510632, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Guangqiang Li, No. 601, West Huangpu Road, Tianhe, Guangzhou, Guangdong, 510632, People’s Republic of China, Tel +86 15521006500, Fax +8620-85226420, Email [email protected] Fang Huang, No. 79, Kangning Road, Xiangzhou District, Zhuhai, Guangdong, 519050, People’s Republic of China, Tel +86 13923373864, Fax +8620-85226420, Email [email protected]
Purpose: Psoriasis is a systemic inflammatory disease, and the mechanism that links psoriasis to depression is still elusive. Hence, this study aimed to elucidate the potential pathogenesis of psoriasis and depression comorbidity.
Methods: The gene expression profiles of psoriasis (GSE34248, GSE78097 and GSE161683) and depression (GSE39653) were downloaded from the Gene Expression Omnibus (GEO) DataSets. Functional annotation, protein‐protein interaction (PPI) network and module construction, and hub gene identification and co-expression analysis were performed, following identification of the common differentially expressed genes (DEGs) of psoriasis and depression.
Results: A total of 115 common DEGs (55 up-regulated and 60 down-regulated) were identified between psoriasis and depression. Functional analysis indicated that T cell activation and differentiation were predominantly implicated in the potential pathogenesis of these two diseases. In addition, Th17 cell differentiation and cytokines is closely related to both. Finally, 17 hub genes were screened, including CTLA4, LCK, ITK, IL7R, CD3D, SOCS1, IL4R, PRKCQ, SOCS3, IL23A, PDGFB, PAG1, TGFA, FGFR1, RELN, ITGB5 and TNXB, which re-emphasized the importance of the immune system in psoriasis and depression.
Conclusion: Our study reveals the common pathogenesis of psoriasis and depression. These common pathways and hub genes may apply to a molecular screening tool for depression in psoriasis patients, which could help dermatologists optimize patient management in routine care.
Keywords: psoriasis, depression, bioinformatics, differentially expressed genes, hub genes
Introduction
Psoriasis is a common, chronic and systemic inflammatory skin disease with multiple comorbidities, leading to a substantial burden for individuals and society.1,2 It is a consequence of genetic susceptibility, particularly in the presence of the HLA-C*06:02 risk allele,3 and of environmental triggers such as stress, streptococcal infection, smoking, obesity, and alcohol consumption.4,5 Due to the appearance of skin manifestations of psoriatic patients, the common stigmatizing attitude of society contributes to the high psychosocial burden of psoriasis. More and more evidence showed that psoriasis is frequently associated with psychological comorbidities such as depression, which can interact negatively with psoriasis to produce a dangerous vicious circle.6–8 Screenings of patients with psoriasis in dermatological clinics found prevalence rates for clinical depression of approximately 12–13% using International Classification of Diseases (ICD) codes.9,10 However, depending on the screening methodology, estimated comorbidities of depressive symptoms are described as present in up to 28–55% of patients with psoriasis.11,12 Given the high numbers of depressive symptoms that is a major source of suicidal ideation in psoriatic patients,13 comprehensive management of patients with psoriasis in dermatological practice is critical. To this end, the establishment of an easy-to-use screening tool for affected patients would be an important complement to routine clinical practice.
Emerging evidence shows that psoriasis and depression have many overlapping biological mechanisms, which are, to a substantial degree, responsible for the pathophysiological link between the two diseases. In depression, proinflammatory cytokines mediated communication between the immune system and the brain by crossing the blood-brain barrier (BBB) plays a central role in the pathogenesis of depression.14,15 Stress triggers the production of various proinflammatory cytokines, including tumor necrosis factor alpha (TNF-α), interleukins (IL)-1β, IL-2, IL-6, IL-8, IL-17, IL-21 and C-reactive protein (CRP).16 These cytokines can impair the BBB,17 damage neurons,18 decrease neurogenesis,19 and cause IL-6 trans-signalling in the CNS.20 Additionally, some proinflammatory cytokines activate plastic Th17 cells, which can disrupt BBB permeability,21 promote neuroinflammation,22 and cause neuronal toxicity.23 The IL-17A produced by Th17 cells can further worsen neuroinflammation and synaptic dysfunction, along with the kynurenine pathway in microglia and astrocytes, which is directly related to the aetiology of depression.24 In psoriasis, a complex interplay between genetic and environmental factors activates plasmacytoid DCs (pDCs) and promotes myeloid dendritic cells (mDC) phenotypic maturation.25 The activated DCs then produce IFN-α, IFN-β, IL-12, IL-23, IL-6, and TNF-α, which activate and polarize auto-aggressive Th cells toward Th1, Th17, and Th22 cell subsets, as well as γδ T cells toward the γδ T17 (IL-17A–producing γδ T cells) subset.26 Th1 cells produce TNF-α and IFN-β to stimulate keratinocyte activation and proliferation, and expression of intercellular adhesion molecule 1. Th17 and γδ T17 cells produce IL17 or IL22 to enhance keratinocyte proliferation and angiogenesis in the lesion.27 Overall, inflammatory cytokines and immune cells released in psoriasis are also released in depression, which suggests that there is a possible common pathogenesis of the two diseases.
Since transcriptome analysis is an effective tool for exploring the pathogenesis of complex human diseases, the common transcriptional characteristic might provide new perspectives on the common pathophysiology of psoriasis and depression. The goal of this research is to identify the common DEGs, hub genes and TFs involved in the pathogenesis of psoriasis and depression. We downloaded and analyzed gene expression data sets from the GEO database. The common DEGs and their functions in psoriasis and depression were determined by using comprehensive bioinformatics and enrichment analysis. In addition, using the STRING database and the Cytoscape software, a PPI network was built to analyze gene modules and identify hub genes. Finally, we identified 17 important hub genes, which were investigated their transcription factors and verified their expression. The hub genes determined here are expected to shed light on the biological mechanisms of psoriasis complicated with depression.
Materials and Methods
Data Source
GEO (http://www.ncbi.nlm.nih.gov/geo) is a microarray and high-throughput sequencing database. The inclusion criteria were set as follows: (1) independent expression profiles of psoriasis or depression; (2) datasets containing patients and healthy controls and including the largest possible sample size; (3) the test specimens in datasets derived from human tissues. Based on the inclusion criteria, 6 gene datasets were chosen from GEO. GSE34248 (14 psoriasis patients and 14 controls), GSE78097 (13 psoriasis patients and 6 controls), GSE161683 (9 psoriasis patients and 9 controls) and GSE137218 (14 psoriasis patients and 14 controls) were screened as psoriasis datasets,28–30 in which the tissue type is skin lesion from psoriasis patients. Meanwhile, GSE39653 (24 depression patients and 21 controls) and GSE98793 (64 depression patients and 64 controls) were screened as depression datasets,31,32 in which the tissue type is peripheral blood mononuclear cells (PBMC) from patients with depression. GSE34248, GSE78097, GSE161683 and GSE39653 were used to screen DEGs, while GSE137218 and GSE98793 were used for hub gene expression validation.
Identification of DEGs
GEO2R (www.ncbi.nlm.nih.gov/geo/ge2r) is an interactive web tool developed based on GEOquery and Limma package. DEGs of psoriasis and depression group were respectively identified using GEO2R with adjusted P-value < 0.05 and |logFC (fold change)| ≥ 1. Genes with multiple probe sets were averaged, and probe sets without a corresponding gene symbol were eliminated. Their common DEGs were detected and visualized by constructing VENN diagrams.
Enrichment Analyses of DEGs
Gene Ontology (GO) enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway analysis were subsequently implemented to explore the biological functions of the overlap DEGs. As described in previous study,33 the enrichment analysis results of GO and KEGG Pathway were obtained from the KEGG Orthology Based Annotation System (KOBAS) (http://kobas.cbi.pku.edu.cn), a Web server for gene/protein functional annotation and functional enrichment developed by Peking University, which collects 4325 species functional annotation information. Adjusted P-value < 0.05 was considered significant.
PPI Network Construction and Module Analysis
Search Tool for the Retrieval of Interacting Genes (STRING; http://string-db.org) (version 11.0)34 is an online database used for predicting direct and indirect protein-protein functional interactions through correlation analyses. Upon the condition of interaction combined score > 0.4, the PPI network of common DEGs was constructed based on the STRING database. Cytoscape (http://www.cytoscape.org) (version 3.7.2)35 was used to visualize this PPI network. Molecular complex detection technology (MCODE), a plugin in Cytoscape, was used to investigate the key functional modules in the PPI network. Set the selection criteria as: K-core = 2, degree cutoff = 2, max depth = 100, and node score cutoff = 0.2. Then the GO and KEGG analysis of the involved modular genes were performed with KOBAS 3.0.
Selection and Analysis of Hub Genes
CytoHubba is a Cytoscape plugin that could calculate the hub genes of the PPI network. In this case, seven algorithms (MCC, DMNC, MNC, Degree, Closeness, EPC, Radiality) were used in CytoHubba to evaluate and select hub genes. Then, GeneMANIA (http://www.genemania.org/),36 a dependable online tool for exploring internal associations in gene sets, was implemented in construction of a co-expression network of these hub genes.
Validation of Hub Genes Expression in Other Data Sets
The expression of hub genes was verified in GSE13721830 and GSE98793.32 The GSE137218 dataset contains 14 paired psoriasis patients with skin lesions and adjacent normal tissues. GSE98793 dataset contains 64 paired PBMC samples from depression patients and healthy control. The comparison between the two sets of data was conducted with the T-test. A P-value < 0.05 was considered significant.
Prediction and Verification of Transcription Factors (TFs)
Transcriptional Regulatory Relationships Unraveled by Sentence-based Text mining (TRRUST), containing the target genes corresponding to TFs and the regulatory relationships between TFs, is a database for the prediction of transcriptional regulatory networks.37 Here, we adopted it to predict TFs that regulate the hub genes, with adjusted P-value < 0.05. Subsequently, we verified the expression levels of these TFs in GSE34248 and GSE39653 with the T-test.
Results
Identification of DEGs
A detailed flowchart of the research is shown in Figure 1. After standardizing the microarray results, DEGs of psoriasis and depression were screened respectively. In the psoriasis group, we obtained 2784 DEGs (1515 up-regulated and 1269 down-regulated) after taking the intersection of the DEGs in GSE34248, GSE78097 and GSE161683 (Figure 2A–E). In depression group, there were a total of 1833 DEGs (915 up-regulated and 896 down-regulated) (Figure 2F–H). After taking the intersection of the Venn diagram, we obtained 115 common DEGs (50 upregulated and 65 downregulated) between psoriasis and depression (Figure 2G and H).
 |
Figure 1 Research design flow chart. |
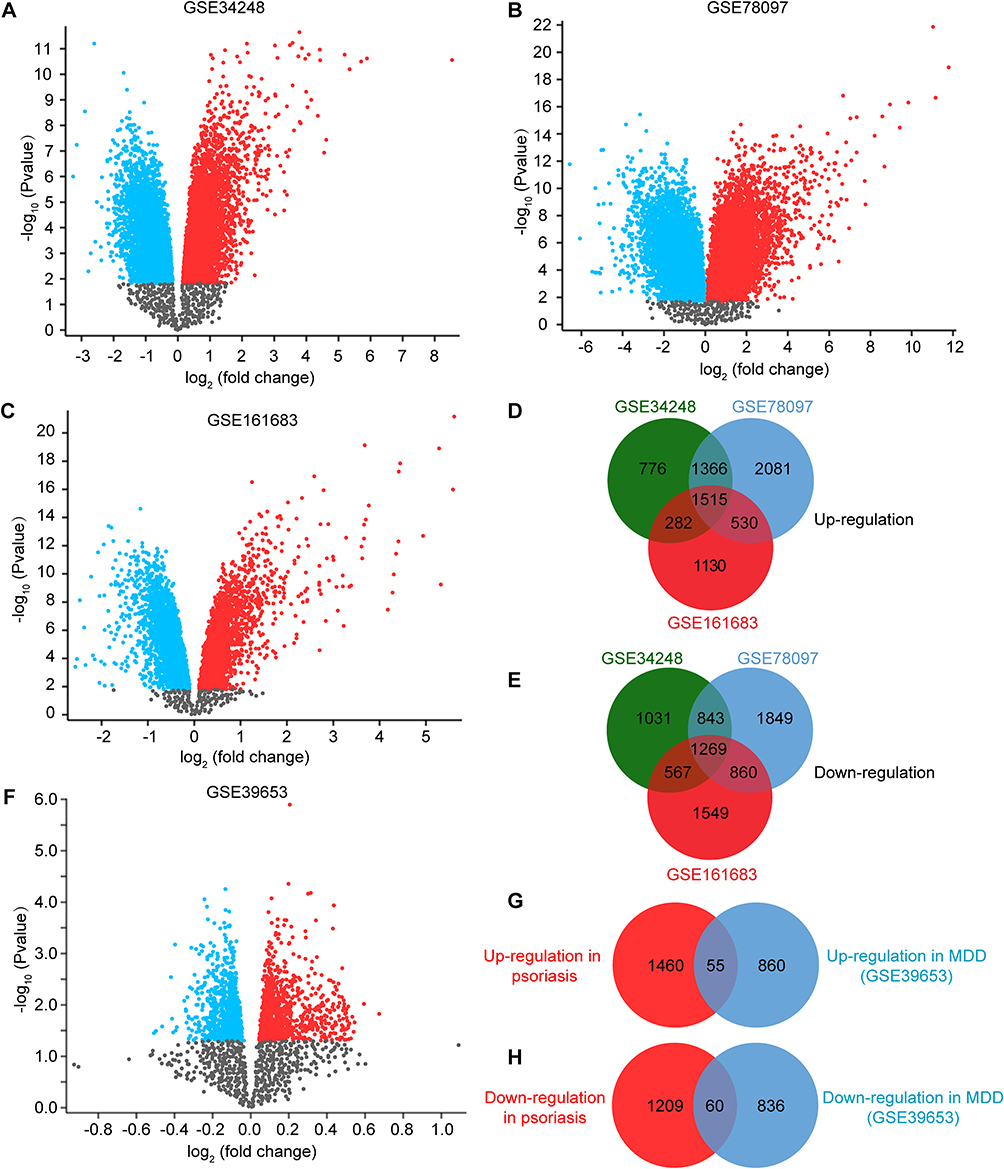 |
Figure 2 Volcano diagram and Venn diagram. (A–C) The volcano map of GSE34248 (A), GSE78097 (B) and GSE161683 (C). Up-regulated genes are marked in red; down-regulated genes are marked in blue. (D and E) Venn diagram of the 1515 common up-regulated DEGs (D) and the 1269 common down-regulated DEGs (E) in psoriasis (PS). (F) The volcano map of GSE39653. Up-regulated genes are marked in red; down-regulated genes are marked in blue. (G and H) Venn diagram of the 55 common up-regulated (G) and 60 common down-regulated (H) DEGs between PS and MDD. |
Functional and Pathway Enrichment Analysis of Common DEGs
GO and KEGG Pathway enrichment analysis were performed to examine the biological characteristic and pathways connected to the 115 common DEGs. The GO result was classified into three functional categories: biological processes (BP), cell component (CC), and molecular function (MF) (Figure 3A–C). In the BP category, changes included significant enrichment of T cell activation, T cell differentiation, positive regulation of cell-cell adhesion, regulation of adaptive immune response and immune response-activating signal transduction (Figure 3A). Within the CC category, these DEGs were mainly enriched in cornified envelope, protein complex involved in cell adhesion, immunological synapse, clathrin-coated vesicle membrane and peroxisomal matrix (Figure 3B). Regarding the MF category, these DEGs were significantly involved in sterol transporter activity, transcription coactivator binding, chemoattractant activity, ligand-activated transcription factor activity and cell adhesion mediator activity (Figure 3C). In terms of KEGG Pathway, PI3K-Akt signaling pathway, T cell receptor signaling pathway and Th17 cell differentiation occupied the top significant enrichment pathways (Figure 3D), which are also the most prominent signaling pathways implicated in human inflammatory diseases. These DEGs were also enriched in other pathways, including the calcium signaling pathway, FoxO signaling pathway and NF-kappa B signaling pathway (Figure 3D). These findings overwhelmingly supported that T cell activation and differentiation could contribute substantially to the onset and progression of both psoriasis and depression.
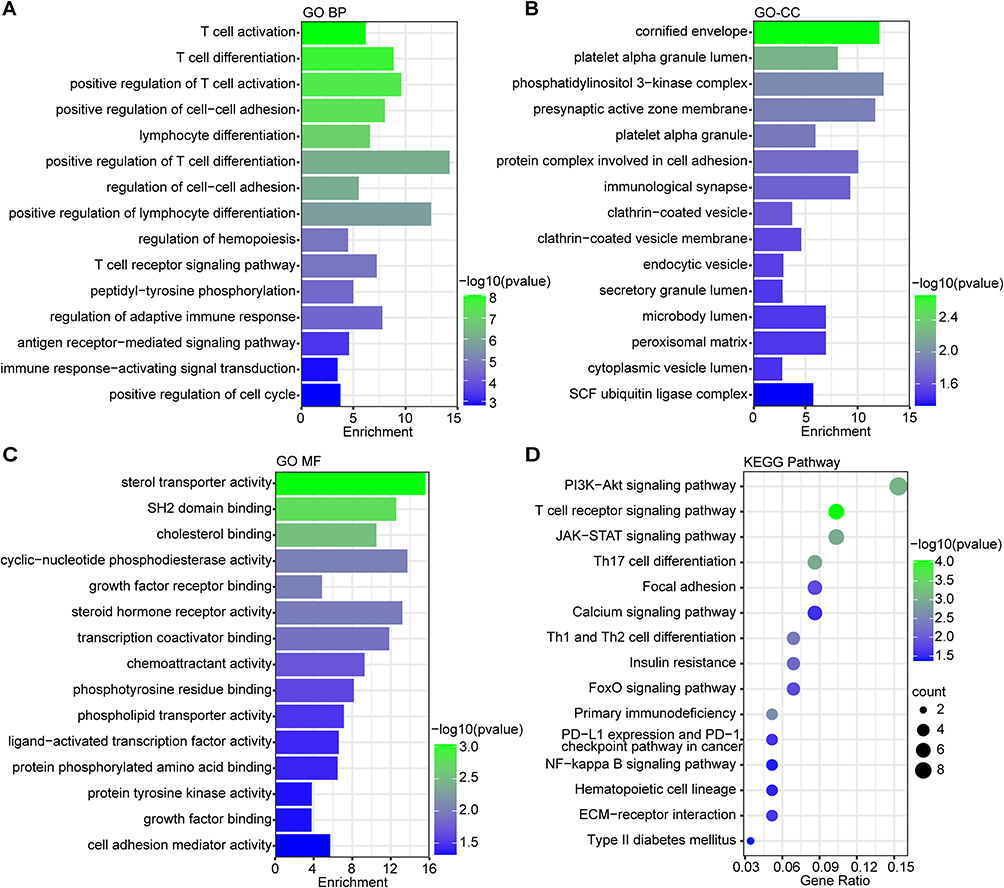 |
Figure 3 The enrichment analysis of common DEGs between PS and MDD. A-C. The GO enrichment analyses of common DEGs with P-value based on the KOBAS 3.0 database. Top 15 BP terms (A), Top 15 CC terms (B) and Top 15 MF terms (C) were shown. (D) The KEGG pathway enrichment analysis of communal DEGs on the ground of KOBAS 3.0 database. The size of the circle represents the number of genes involved, and the abscissa represents the frequency of the genes involved in the term total genes. P-value < 0.05 was considered significant. |
PPI Network Construction and Module Analysis
The PPI network of common DEGs was constructed using Cytoscape on the ground of the STRING database. We found 34 nodes and 132 edges in the network (Figure 4A) under the condition of combined scores of >0.4 points. The top two significant modules were extracted through MCODE plug-in of Cytoscape, of which, module 1 contained 7 nodes and 28 edges (Figure 4B) and module 2 contained 3 nodes and 6 edges (Figure 4C). GO analysis showed that these genes were related to T cell-mediated inflammation and immune response (Figure 4D). KEGG Pathway analysis showed that they were mainly involved in PI3K-Akt signaling pathway, T cell receptor signaling pathway and Th17 cell differentiation (Figure 4E). Both PPI network and significant modules indicated that psoriasis and depression share some common potential molecular mechanisms.
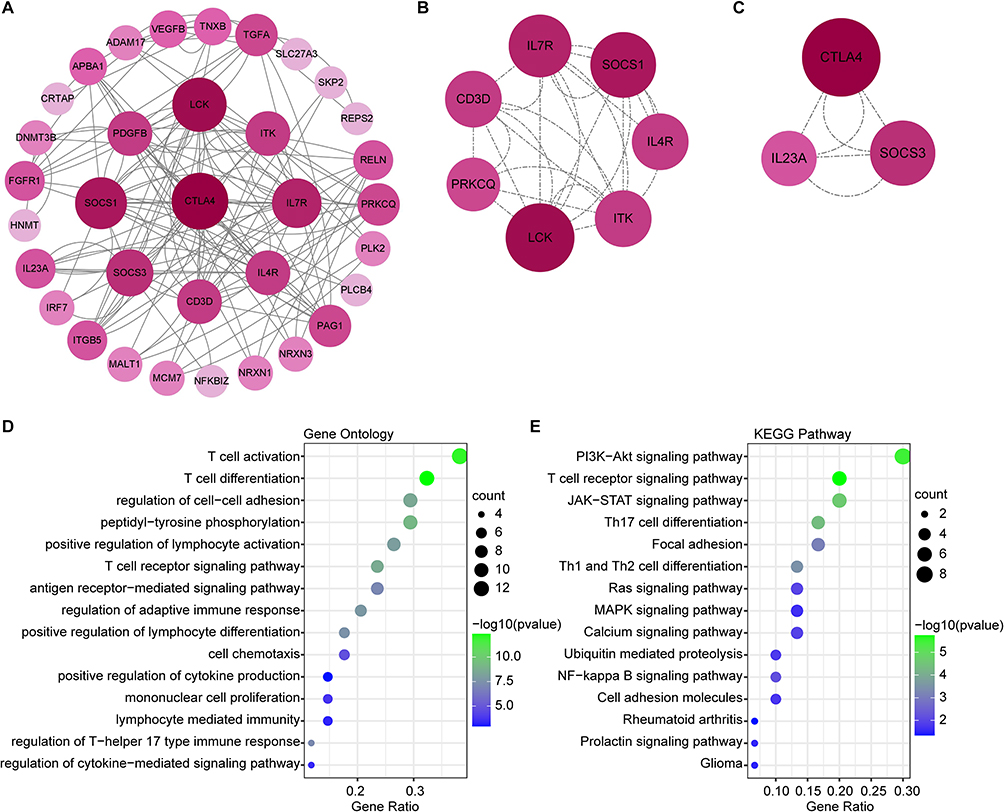 |
Figure 4 The results of PPI network and significant modules. (A) PPI network of common DEGs on the ground of STRING database. (B and C) Two significant gene clustering modules. Significant module 1 (B) and significant module 2 (C) were shown. (D and E) GO and KEGG enrichment analysis of the modular genes. The size of the circle represents the number of genes involved, and the abscissa represents the frequency of the genes involved in the term total genes. P-value < 0.05 was considered significant. |
Hub Gene Identification and Analysis
The top 20 hub genes were screened out based on the seven algorithms of plug-in CytoHubba (Supplementary Table 1). After taking the intersection of the Venn diagrams, we found 17 hub genes (Figure 5A), including CTLA4, LCK, ITK, IL7R, CD3D, SOCS1, IL4R, PRKCQ, SOCS3, IL23A, PDGFB, PAG1, TGFA, FGFR1, RELN, ITGB5 and TNXB (Figure 5B). Among them, CTLA4, LCK, ITK, IL7R, CD3D, SOCS1, IL4R, PRKCQ, SOCS3, IL23A and TGFA were up-regulated genes, while PDGFB, PAG1, FGFR1, RELN, ITGB5 and TNXB were down-regulated genes. Subsequently, the network of the hub genes and their co-expression genes were generated on the ground of the GeneMANIA platform (Figure 5C). These genes showed the complex PPI network with the co-expression of 65.06%, physical interactions of 13.85%, pathway of 10.22%, predicted of 3.06% and co-localization of 2.14% (Figure 5C). As expected, the biological function and roles of the hub genes re-emphasized the importance of the Th17 cells, cytokines and mononuclear cells in psoriasis and depression. To explore the biological functions and pathways related to these hub genes, GO and KEGG analyses were performed. GO analysis showed that these genes were significantly enriched in T cell activation, T cell differentiation, cell chemotaxis, regulation of T-helper 17 type immune response and regulation of cytokine-mediated signaling pathway (Figure 5D). Besides, KEGG analysis showed that they are mainly involved in PI3K-Akt signaling pathway, T cell receptor signaling pathway, Cytokine-cytokine receptor interaction, Ras signaling pathway and NF-kappa B signaling pathway (Figure 5E). Interestingly, five genes (LCK, CD3D, IL4R, PRKCQ and IL23A) were also involved in Th17 cell differentiation. Consistent with the analysis outcome of common DEGs, the gene annotation analysis revealed that hub genes were associated with the process of the immune system reacting.
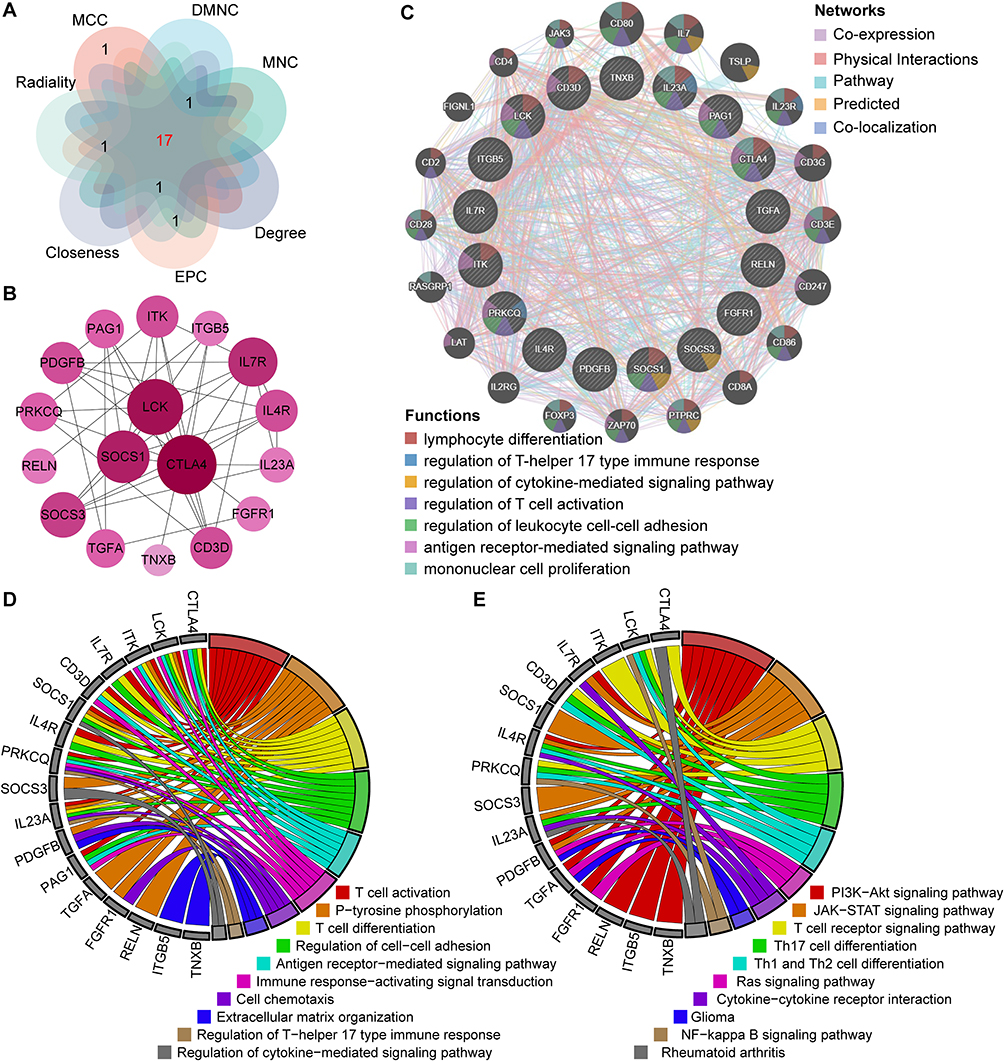 |
Figure 5 Co-expression network and enrichment analysis of hub genes. (A) 17 hub genes were determined by taking the intersection of the top 20 genes in the seven algorithms of cytoHubba plugin (MCC, DMNC, MNC, Degree, EPC, Closeness and Radiality). (B) PPI network of 17 hub genes. (C) The network of hub genes and their co-expression genes were constructed and analyzed by GeneMANIA. (D and E) GO (D) and KEGG (E) enrichment analysis of the hub genes. The outermost circle is term on the right and the inner circle on the left represents the significant P-value of the corresponding pathway of the gene. |
Verification of Hub Genes Expression
In order to verify the reliability and accuracy of bioinformatics analysis results, two other data sets were adopted to verify the expression levels of hub genes in psoriasis (GSE137218) and depression (GSE98793) samples by independence testing analysis respectively. Encouragingly, the results showed that CD3D, CTLA4, IL4R, IL7R, IL23A, LCK, PRKCQ, SOCS1, SOCS3 and TGFA were significantly up-regulated, while FGFR1, ITGB5, PDGFB and TNXB were significantly down-regulated in psoriasis patients from GSE137218 (Figure 6), which is consistent with aforementioned results from GSE34248, GSE78097 and GSE161683. Interestingly, these hub genes significantly conversely expressed in psoriatic patients during anti-IL-17A (secukinumab) treatment (Figure 6). In addition, the expression of three hub genes (ITK, PAG1 and RELN) was no significant difference in GSE137218 (Figure 6). Similarly, the expression of CTLA4, IL7R, IL23A, LCK, SOCS1, SOCS3 and TGFA in depression samples was higher, whereas the expression of FGFR1 and PDGFB was lower, than in normal samples from GSE98793 (Figure 7). Although the expression of eight hub genes (CD3D, IL4R, ITGB5, ITK, PAG1, PRKCQ, RELN, and TNXB) was no significant difference in GSE98793, these results increased our confidence in the findings.
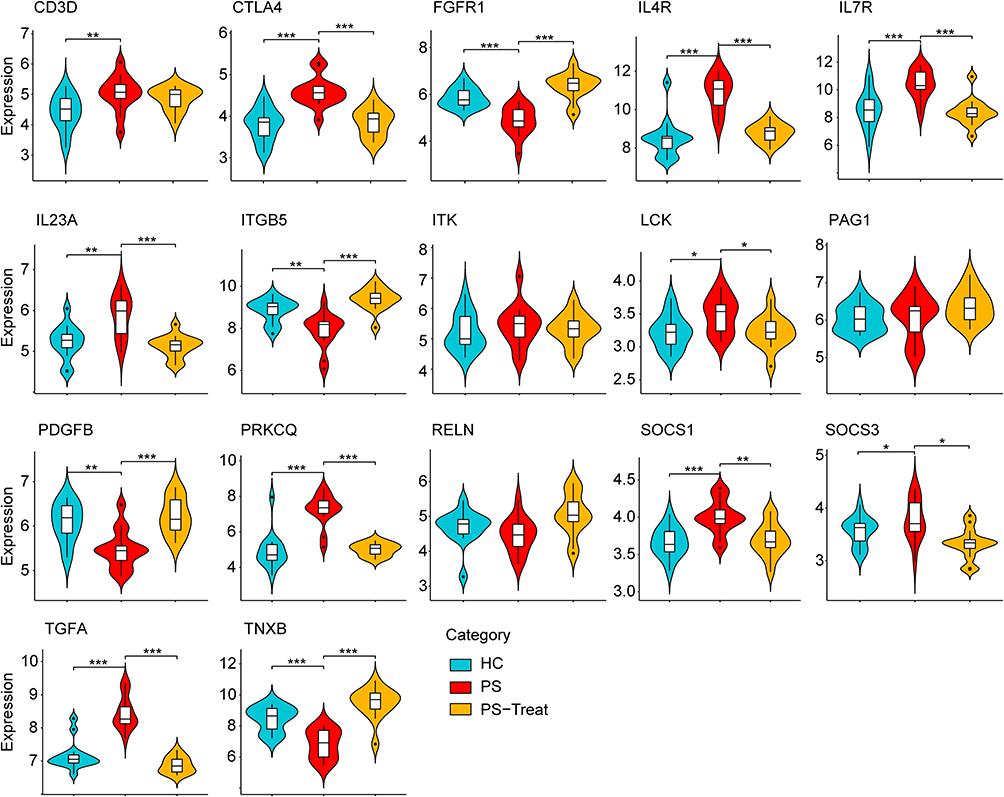 |
Figure 6 The expression level of hub gene in GSE137218. The comparison between the two sets of data uses the mean T-test. P-value < 0.05 was considered statistically significant. *p < 0.05; **p < 0.01; ***p < 0.001. Abbreviations: HC, healthy controls; PS, psoriasis patients; PS-Treat, psoriasis patients with secukinumab treatment. |
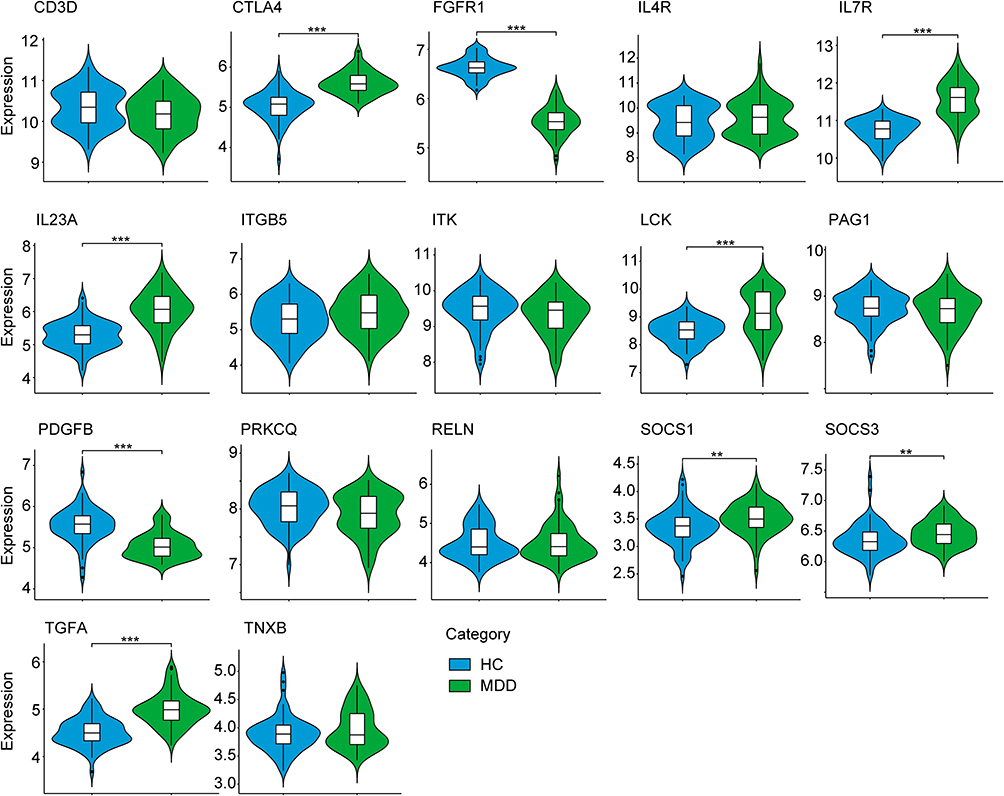 |
Figure 7 The expression level of hub gene in GSE98793. The comparison between the two sets of data uses the mean T-test. P-value < 0.05 was considered statistically significant. **p < 0.01; ***p < 0.001. Abbreviations: HC, healthy controls; MDD, major depression disorder. |
Prediction and Validation of TFs
On the ground of the TRRUST database predicting TFs, we obtained 5 TFs that may regulate the expression of these hub genes, as shown in the regulatory network of TF-hub genes constructed using Cytoscape software (Figure 8A). Supplementary Table 2 shows their full names and related functions. Further verification, we found that 4 TFs (E2F1, NFKB1, RELA and STAT3) are highly expressed in psoriasis (Figure 8B) and depression (Figure 8C). Only one TF (SP1) was not differentially expressed both in psoriasis (Figure 8B) and depression (Figure 8C). Taken together, they coordinately participated in the regulation of 6 hub genes (IL23A, SOCS1, SOCS3, TGFA, FGFR1 and PDGFB).
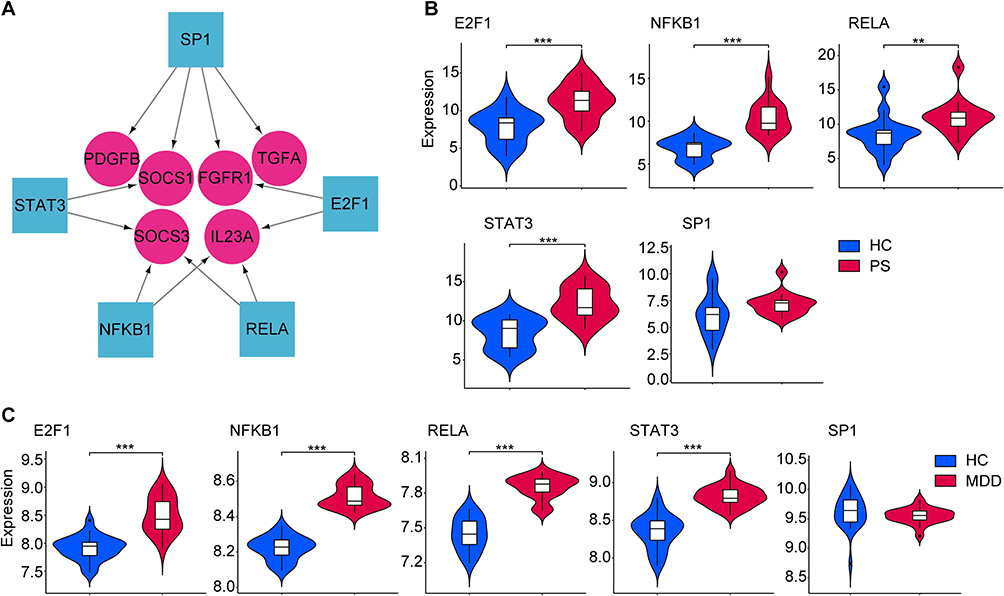 |
Figure 8 TFs regulatory network and its expression in GSE34248 and GSE39653. (A) TFs regulatory network. TFs were marked as square, and the hub genes were marked as circle. (B and C) The expression level of TFs in GSE34248 and GSE39653. The comparison between the two sets of data uses the mean T-test. P-value < 0.05 was considered statistically significant. **p < 0.01; ***p < 0.001. Abbreviations: HC, healthy controls; PS, psoriasis patients; MDD, major depression disorder. |
Discussion
Psoriasis is a chronic, inflammatory skin disease with a range of comorbidities. The common comorbidities are mental disorders, especially depression, which can interact negatively with psoriasis to produce a dangerous vicious circle.38 Pruritus and sleep disorders are also common comorbidities of psoriasis, which is known to be a risk factor for depression in psoriasis patients.39,40 It is believed that the chronicity and severity of pruritus may contribute to the development of depression through alterations in the brain, whereas sleep dysfunction and psoriasis are thought to be linked by pruritus and depression.41 However, a recent theory connecting depression and psoriasis through chronic inflammation offers new insights into our comprehension of these conditions.7 Increased proinflammatory mediators, such as C-reactive protein or interleukin-6, are present in both psoriasis and depression.42 Biologics that target inflammatory cytokines treat the clinical manifestations of psoriasis, but also resulted in significantly improved the associated depressive symptoms.43 Recently, the inflammatory overlap between psoriasis and depression were highlighted by examining the pathophysiology of depression,7 indicating a common pathogenic mechanism between psoriasis and depression.
Previous studies have separately explored the hub genes associated with psoriasis and depression. However, few investigations have described the common molecular mechanism between them by advanced bioinformatics techniques. In this study, we identified 115 common DEGs in both, of which 17 were hub genes, including CTLA4, LCK, ITK, IL7R, CD3D, SOCS1, IL4R, PRKCQ, SOCS3, IL23A, PDGFB, PAG1, TGFA, FGFR1, RELN, ITGB5 and TNXB. According to the outcomes of GO and KEGG Pathway enrichment analysis, the hub genes were strongly enriched in immune and inflammatory processes. Cytokines, chemokines and adhesion molecules are jointly implicated in the pathogenesis of these two diseases, such as serum TNF-α, IL-1β, IL-2, IL-6, IL17A, prostaglandin E2, C-reactive protein and MCP-1.44 Interestingly, the hub genes were also enriched in regulation of cytokine-mediated signaling pathway, cytokine-cytokine receptor interaction, cell chemotaxis and regulation of cell-cell adhesion. In addition, most of these functional molecules are regulated by NF-κB signaling pathway, in which the hub genes were also enriched. By validating the hug genes in other datasets, we further screened out 9 hub genes with higher confidence, including CTLA4, FGFR1, IL7R, IL23A, LCK, PDGFB, SOCS1, SOCS3 and TGFA. However, 8 hub genes (CD3D, IL4R, ITGB5, ITK, PAG1, PRKCQ, RELN, and TNXB), most of them from depression group, was not successfully authenticated, which may be due to the high heterogeneity of depression.45 Moreover, we found that 5 TFs may regulate the expression of the hub genes. By further verification, we found that four TFs are highly expressed in psoriasis and depression, including E2F1, STAT3, NFKB1 and RELA. They coordinately participated in the regulation of six hub genes (IL23A, TGFA, FGFR1, PDGFB, SOCS1 and SOCS3). Interestingly, in psoriasis and depression, FGFR1 and PDGFB are found to be downregulated, despite their transcription factors being upregulated. This could be due to post-transcriptional modification or negative feedback mechanisms, or the competitive occupancy of transcription sites by SP1.
IL-23A belongs to the IL-12 pro-inflammatory cytokine family, which is necessary for early local inflammatory response by connecting innate and adaptive immune system. It is considered as a key cytokine for the proper functioning of IL-17 producing Th17 cells by strongly enhancing the expansion of Th17 cells, indicating importance of IL-23A for many inflammatory autoimmune diseases.46 Psoriasis is a disease in which IL-23A/IL-17 axis plays an important role, hence the significance of IL-23 in the pathogenesis of this disease is not surprising. Studies have confirmed IL-17A produced by Th17 cells provokes the deterioration of neuroinflammation and synaptic dysfunction, and the kynurenine pathway in microglia and astrocytes, which is directly related to the aetiology of depression.24 Interestingly, we observed a statistically significant increase of IL-23A expression in peripheral blood from patients with depression. Ustekinumab is a monoclonal antibody targeting the common subunit p40 of IL-12 and IL-23, which can prevent later signaling, differentiation and production of the cytokines that are crucial for psoriasis. Strikingly, patients taking ustekinumab for psoriasis showed a significant improvement in depressive symptoms.47 These observations indicated that elevated expression of IL-23A in patients with psoriasis could contribute to the increased risk for depressive symptoms. TGFA is believed to play an important part in cell proliferation and differentiation through the epithelial growth factor receptor (EGFR). It is reported that TGFA production by keratinocytes is increased in psoriatic skin lesions.48 Although TGFA did not induce psoriasis-related gene expression, it could significantly enhance the IL-17A-mediated induction of various psoriasis signature genes, including antimicrobial peptides, cytokines, and chemokines.49 Thus, TGFA production in psoriatic lesions may amplify the inflammation in the lesions. In depression, data regarding role of TGFA is lacking, however, it could be advantageous in supporting the neuroprotective effect in the CNS. Specifically, TGFA has been reported to contribute to enhancing the repair after nervous system post-stroke and post-traumatic injury.50 Therefore, it might play a beneficial role in maintaining brain plasticity in MDD. FGFR1, a member of the receptor tyrosine kinase family, was found in dermatitis by a global transcriptome analysis.51 Although combined stimulation via FGFR1 and the TCR synergistically enhances the T-cell activation,52 decreased expression of FGFR1 was associated with psoriasis. Loss of FGFR1 in keratinocytes caused psoriasis-like inflammation by activation of keratinocytes and epidermal γδ T cells, which produce interleukin-1 family member 8 and S100A8/A9 proteins.53 On the other hand, FGFR1 is widely distributed throughout the nervous system and is negatively related to depression status.54,55 FGF2, a ligand of FGFR1, was down-regulated in the hippocampus and the serum with high consistency in patients with MDD.56 Notably, we also discovered FGFR1 was significantly decreased in peripheral blood in patients with MDD, which suggests that low FGFR1 and FGF2 expression in peripheral blood is involved in the pathogenesis of depression. Thus, considering the difficulty of obtaining brain tissue samples, FGFR1 and FGF2 has the potential to be a good serum biomarker for the detection of depression. PDGFB is considered as a mitogenic hormone, which attracts monocytes, promotes neovascularization, and stimulates connective tissue cells to proliferate, migrate, and synthesize extracellular matrix components. PDGF ligands and their corresponding receptors are associated with skin repair.57 However, little is known about the regulatory role of PDGFB in psoriasis. Given that keratinocytes and macrophages residing in skin contribute to PDGF production in the injured skin, which plays the chemotaxis role for monocytes.57 Thus, PDGFB might amplify the inflammation in the psoriatic lesions. Peripheral inflammation can cause BBB disruption/dysfunction, thereby increasing PDGF levels, which were inversely associated with integrity of myelin sheaths in depressed patients.58 Hence, elevated levels of PDGF in periphery may reflect neuroinflammation. SOCS1 and SOCS3 are members of the SOCS family, which has a vital role in the regulation of the immune system and resolution of inflammatory cascades. Since SOCS1 is a classical regulator of IFN-γ and IL-6 signaling, SOCS1 can skew T cells responses away from Th1 and Th17 cells.59 In addition, SOCS1 and SOCS3 suppress TLR7-mediated type I IFN production of pDCs, another vital element in psoriasis pathogenesis.60 In depression, study has demonstrated that blood SOCS1 and SOCS3 mRNA were significantly decreased in the MDD group compared to the NC group.61 Of particular interest, a significant finding was a decrease in SOCS3 mRNA after remission from MDD, implying that SOCS3 is a trait marker indicating a tendency for inflammatory cytokine in depressive symptoms.61 Prior study explored the inflammatory genes related to depression in skin and serum from patients with psoriasis. It found an increase in skin and serum gene expression for TNF-α, IL1β, IL-6 and IL-17 in psoriasis versus healthy controls. Our study found that new genes like CTLA4, FGFR1, IL7R, IL23A, LCK, PDGFB, SOCS1, SOCS3 and TGFA associated with inflammation present abnormal expression both in psoriasis and depression.
Screening for depressive symptoms led to increased utilization of mental health care and improvement of psoriasis, depressive symptoms, and quality of life.62 As a result, it is recommended that dermatologists screen for depression in order to ensure the caution in medication and patient management. The Whooley questions, a convenient two-question case-finding instrument, might be easy to use as a screening tool for depression in psoriasis patients.63 However, there are currently no molecular biomarker-based screening techniques available to identify depression in psoriasis patients. In this study, we used advanced bioinformatics methods to identify the common DEGs, hub genes and TFs implicated in the pathogenesis of psoriasis and depression for the first time, which helps to further elucidate the molecular mechanism of the two diseases. In addition, the potential molecular biomarkers we explored might be incorporated into existing diagnostic procedures to diagnose psoriasis patients with depression. Nevertheless, it is important to acknowledge the limitations of our research. Firstly, as a retrospective study, our findings require external validation to ensure their reliability and generalizability. Additionally, it is crucial to ensure that the verification databases selected are appropriate for studying the comorbidity of psoriasis and depression. Finally, while our study identified hub genes associated with psoriasis and depression, further experimental verification is needed to confirm their functional significance. Therefore, future research will focus on conducting experiments to validate the roles of these hub genes in the pathogenesis of psoriasis and depression.
Conclusions
In summary, our study successfully identified 115 common DEGs associated with both psoriasis and depression. Additionally, psoriasis and depression share common pathogenic mechanisms, such as T cell activation and Th17 cell differentiation, which are mediated by 17 specific hub genes. These findings contribute to a deeper understanding of the molecular mechanisms underlying the comorbidity of psoriasis and depression, and provide important insights for further research in this area. Overall, our study represents an important step towards the development of new therapies and treatments for individuals affected by psoriasis complicated with depression.
Ethics
The conduct of this study complies with the Declaration of Helsinki. For ethics approval waived, this study was approved by IRB of The First Affiliated Hospital of Jinan University, Guangzhou, China. The ethics waiver reason is described below. GEO belongs to public databases. The patients involved in the database have obtained ethical approval. Users can download relevant data for free for research and publish relevant articles. Our study is based on the open source data downloaded from GEO, so there are no ethical issues.
Acknowledgment
We thank the researchers who are generous and have uploaded their experimental data onto online databases. We thank Professor Yang (The Second Affiliated Hospital of Soochow University, Suzhou, China) for his methodological assistances.
Funding
This work is supported by grants from the National Natural Science Foundation of China (grant 82103722, to XX, grant 82103542, to GL), GuangDong Basic and Applied Basic Research Foundation (grant 2020A1515110052, to XX), China Postdoctoral Science Foundation (2022M723699, to XX) and the Postdoctoral Fund of Zhuhai People’s Hospital (grant BSHQD2022090006, BSHQD2023020001 to XX).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Griffiths CEM, Armstrong AW, Gudjonsson JE, Barker J. Psoriasis. Lancet. 2021;397(10281):1301–1315. doi:10.1016/S0140-6736(20)32549-6
2. Damiani G, Radaeli A, Olivini A, Calvara-Pinton P, Malerba M. Increased airway inflammation in patients with psoriasis. Br J Dermatol. 2016;175(4):797–799. doi:10.1111/bjd.14546
3. Dand N, Duckworth M, Baudry D, et al. HLA-C*06:02 genotype is a predictive biomarker of biologic treatment response in psoriasis. J Allergy Clin Immunol. 2019;143(6):2120–2130. doi:10.1016/j.jaci.2018.11.038
4. Griffiths CE, Barker JN. Pathogenesis and clinical features of psoriasis. Lancet. 2007;370(9583):263–271. doi:10.1016/S0140-6736(07)61128-3
5. Kumar S, Han J, Li T, Qureshi AA. Obesity, waist circumference, weight change and the risk of psoriasis in US women. J Eur Acad Dermatol Venereol. 2013;27(10):1293–1298. doi:10.1111/jdv.12001
6. Zafiriou E, Daponte AI, Siokas V, et al. Depression and obesity in patients with psoriasis and psoriatic arthritis: is IL-17-mediated immune dysregulation the connecting link? Front Immunol. 2021;12:699848. doi:10.3389/fimmu.2021.699848
7. Patel N, Nadkarni A, Cardwell LA, et al. Psoriasis, depression, and inflammatory overlap: a review. Am J Clin Dermatol. 2017;18(5):613–620. doi:10.1007/s40257-017-0279-8
8. Wang X, Li Y, Wu L, et al. Dysregulation of the gut-brain-skin axis and key overlapping inflammatory and immune mechanisms of psoriasis and depression. Biomed Pharmacother. 2021;137:111065. doi:10.1016/j.biopha.2020.111065
9. Dalgard FJ, Gieler U, Tomas-Aragones L, et al. The psychological burden of skin diseases: a cross-sectional multicenter study among dermatological out-patients in 13 European countries. J Invest Dermatol. 2015;135(4):984–991. doi:10.1038/jid.2014.530
10. Dowlatshahi EA, Wakkee M, Arends LR, Nijsten T. The prevalence and odds of depressive symptoms and clinical depression in psoriasis patients: a systematic review and meta-analysis. J Invest Dermatol. 2014;134(6):1542–1551. doi:10.1038/jid.2013.508
11. Lamb RC, Matcham F, Turner MA, et al. Screening for anxiety and depression in people with psoriasis: a cross-sectional study in a tertiary referral setting. Br J Dermatol. 2017;176(4):1028–1034. doi:10.1111/bjd.14833
12. Korman AM, Hill D, Alikhan A, Feldman SR. Impact and management of depression in psoriasis patients. Expert Opin Pharmacother. 2016;17(2):147–152. doi:10.1517/14656566.2016.1128894
13. Singh S, Taylor C, Kornmehl H, Armstrong AW. Psoriasis and suicidality: a systematic review and meta-analysis. J Am Acad Dermatol. 2017;77(3):425–440 e422. doi:10.1016/j.jaad.2017.05.019
14. Miller AH, Raison CL. The role of inflammation in depression: from evolutionary imperative to modern treatment target. Nat Rev Immunol. 2016;16(1):22–34. doi:10.1038/nri.2015.5
15. Roman M, Irwin MR. Novel neuroimmunologic therapeutics in depression: a clinical perspective on what we know so far. Brain Behav Immun. 2020;83:7–21. doi:10.1016/j.bbi.2019.09.016
16. Beurel E, Toups M, Nemeroff CB. the bidirectional relationship of depression and inflammation: double trouble. Neuron. 2020;107(2):234–256. doi:10.1016/j.neuron.2020.06.002
17. Cheng Y, Desse S, Martinez A, et al. TNFα disrupts blood brain barrier integrity to maintain prolonged depressive-like behavior in mice. Brain Behav Immun. 2018;69:556–567. doi:10.1016/j.bbi.2018.02.003
18. Varatharaj A, Galea I. The blood-brain barrier in systemic inflammation. Brain Behav Immun. 2017;60:1–12. doi:10.1016/j.bbi.2016.03.010
19. Borsini A, Alboni S, Horowitz MA, et al. Rescue of IL-1β-induced reduction of human neurogenesis by omega-3 fatty acids and antidepressants. Brain Behav Immun. 2017;65:230–238. doi:10.1016/j.bbi.2017.05.006
20. Baran P, Hansen S, Waetzig GH, et al. The balance of interleukin (IL)-6, IL-6·soluble IL-6 receptor (sIL-6R), and IL-6·sIL-6R·sgp130 complexes allows simultaneous classic and trans-signaling. J Biol Chem. 2018;293(18):6762–6775. doi:10.1074/jbc.RA117.001163
21. Schiweck C, Valles-Colomer M, Arolt V, et al. Depression and suicidality: a link to premature T helper cell aging and increased Th17 cells. Brain Behav Immun. 2020;87:603–609. doi:10.1016/j.bbi.2020.02.005
22. Beurel E, Lowell JA. Th17 cells in depression. Brain Behav Immun. 2018;69:28–34. doi:10.1016/j.bbi.2017.08.001
23. Siffrin V, Radbruch H, Glumm R, et al. In vivo imaging of partially reversible th17 cell-induced neuronal dysfunction in the course of encephalomyelitis. Immunity. 2010;33(3):424–436. doi:10.1016/j.immuni.2010.08.018
24. Anderson G. Editorial (Thematic Issue: the kynurenine and melatonergic pathways in psychiatric and CNS disorders). Curr Pharm Des. 2016;22(8):947–948. doi:10.2174/1381612822999160104143932
25. Rendon A, Schakel K. Psoriasis pathogenesis and treatment. Int J Mol Sci. 2019;20(6):1475. doi:10.3390/ijms20061475
26. Xia X, Cao G, Sun G, et al. GLS1-mediated glutaminolysis unbridled by MALT1 protease promotes psoriasis pathogenesis. J Clin Invest. 2020;130(10):5180–5196. doi:10.1172/JCI129269
27. Liu T, Li S, Ying S, et al. The IL-23/IL-17 pathway in inflammatory skin diseases: from bench to bedside. Front Immunol. 2020;11:594735. doi:10.3389/fimmu.2020.594735
28. Bigler J, Rand HA, Kerkof K, Timour M, Russell CB, Brandner JM. Cross-study homogeneity of psoriasis gene expression in skin across a large expression range. PLoS One. 2013;8(1):e52242. doi:10.1371/journal.pone.0052242
29. Kim J, Bissonnette R, Lee J, et al. The spectrum of mild to severe psoriasis vulgaris is defined by a common activation of IL-17 pathway genes, but with key differences in immune regulatory genes. J Invest Dermatol. 2016;136(11):2173–2182. doi:10.1016/j.jid.2016.04.032
30. Bertelsen T, Ljungberg C, Litman T, et al. IκBζ is a key player in the antipsoriatic effects of secukinumab. J Allergy Clin Immunol. 2020;145(1):379–390. doi:10.1016/j.jaci.2019.09.029
31. Savitz J, Frank MB, Victor T, et al. Inflammation and neurological disease-related genes are differentially expressed in depressed patients with mood disorders and correlate with morphometric and functional imaging abnormalities. Brain Behav Immun. 2013;31:161–171. doi:10.1016/j.bbi.2012.10.007
32. Leday GGR, Vértes PE, Richardson S, et al. Replicable and coupled changes in innate and adaptive immune gene expression in two case-control studies of blood microarrays in major depressive disorder. Biol Psychiatry. 2018;83(1):70–80. doi:10.1016/j.biopsych.2017.01.021
33. Su W, Zhao Y, Wei Y, et al. Exploring the pathogenesis of psoriasis complicated with atherosclerosis via microarray data analysis. Front Immunol. 2021;12:667690. doi:10.3389/fimmu.2021.667690
34. Szklarczyk D, Gable AL, Lyon D, et al. STRING v11: protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019;47(D1):D607–D613. doi:10.1093/nar/gky1131
35. Smoot ME, Ono K, Ruscheinski J, Wang P-L, Ideker T. Cytoscape 2.8: new features for data integration and network visualization. Bioinformatics. 2011;27(3):431–432. doi:10.1093/bioinformatics/btq675
36. Warde-Farley D, Donaldson SL, Comes O, et al. The GeneMANIA prediction server: biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010;38(suppl_2):W214–W220. doi:10.1093/nar/gkq537
37. Han H, Cho J-W, Lee S, et al. TRRUST v2: an expanded reference database of human and mouse transcriptional regulatory interactions. Nucleic Acids Res. 2018;46(D1):D380–D386. doi:10.1093/nar/gkx1013
38. Gonzalez-Parra S, Dauden E. Psoriasis and depression: the role of inflammation. Actas Dermosifiliogr. 2019;110(1):12–19. doi:10.1016/j.ad.2018.05.009
39. Damiani G, Cazzaniga S, Conic RR, Naldi L, Griseta V. Pruritus characteristics in a large Italian cohort of psoriatic patients. J Eur Acad Dermatol Venereol. 2019;33(7):1316–1324. doi:10.1111/jdv.15539
40. Nowowiejska J, Baran A, Lewoc M, et al. The assessment of risk and predictors of sleep disorders in patients with psoriasis—a questionnaire-based cross-sectional analysis. J Clin Med. 2021;10(4):664. doi:10.3390/jcm10040664
41. Myers B, Reddy V, Brownstone N, et al. Sleep and the gut microbiome in psoriasis: clinical implications for disease progression and the development of cardiometabolic comorbidities. J Psoriasis Psoriatic Arthritis. 2021;6(1):27–37. doi:10.1177/2475530320964781
42. Holsken S, Krefting F, Schedlowski M, Sondermann W. Common fundamentals of psoriasis and depression. Acta Derm Venereol. 2021;101(11):adv00609. doi:10.2340/actadv.v101.565
43. Talamonti M, Malara G, Natalini Y, et al. Secukinumab improves patient perception of anxiety and depression in patients with moderate to severe psoriasis: a post hoc analysis of the SUPREME study. Acta Derm Venereol. 2021;101(3):adv00422. doi:10.2340/00015555-3712
44. Aleem D, Tohid H. Pro-inflammatory cytokines, biomarkers, genetics and the immune system: a mechanistic approach of depression and psoriasis. Rev Colomb Psiquiatr. 2018;47(3):177–186. doi:10.1016/j.rcp.2017.03.002
45. Cai N, Choi KW, Fried EI. Reviewing the genetics of heterogeneity in depression: operationalizations, manifestations and etiologies. Hum Mol Genet. 2020;29(R1):R10–R18. doi:10.1093/hmg/ddaa115
46. Galecka M, Bliźniewska-Kowalska K, Orzechowska A, et al. Inflammatory versus anti-inflammatory profiles in major depressive disorders—the role of IL-17, IL-21, IL-23, IL-35 and Foxp3. J Pers Med. 2021;11(2):66. doi:10.3390/jpm11020066
47. Langley RG, Feldman SR, Han C, et al. Ustekinumab significantly improves symptoms of anxiety, depression, and skin-related quality of life in patients with moderate-to-severe psoriasis: results from a randomized, double-blind, placebo-controlled Phase III trial. J Am Acad Dermatol. 2010;63(3):457–465. doi:10.1016/j.jaad.2009.09.014
48. Sergi C, Kahl P, Otto HF. Immunohistochemical localization of transforming growth factor-α and epithelial growth factor receptor in human fetal developing skin, psoriasis and restrictive dermopathy. Pathol Oncol Res. 2000;6(4):250–255. doi:10.1007/BF03187327
49. Dai X, Murakami M, Shiraishi K, et al. EGFR ligands synergistically increase IL-17A-induced expression of psoriasis signature genes in human keratinocytes via IκBζ and Bcl3. Eur J Immunol. 2022;52(6):994–1005. doi:10.1002/eji.202149706
50. Lee KM, Kim YK. The role of IL-12 and TGF-beta1 in the pathophysiology of major depressive disorder. Int Immunopharmacol. 2006;6:1298–1304. doi:10.1016/j.intimp.2006.03.015
51. Chen B, Li C, Chang G, Wang H. Dihydroartemisinin targets fibroblast growth factor receptor 1 (FGFR1) to inhibit interleukin 17A (IL-17A)-induced hyperproliferation and inflammation of keratinocytes. Bioengineered. 2022;13(1):1530–1540. doi:10.1080/21655979.2021.2021701
52. Byrd VM, Kilkenny DM, Dikov MM, et al. Fibroblast growth factor receptor-1 interacts with the T-cell receptor signalling pathway. Immunol Cell Biol. 2003;81(6):440–450. doi:10.1046/j.1440-1711.2003.01199.x
53. Yang J, Meyer M, Müller AK, et al. Fibroblast growth factor receptors 1 and 2 in keratinocytes control the epidermal barrier and cutaneous homeostasis. J Cell Biol. 2010;188(6):935–952. doi:10.1038/jid.2013.508
54. Turner CA, Gula EL, Taylor LP, Watson SJ, Akil H. Antidepressant-like effects of intracerebroventricular FGF2 in rats. Brain Res. 2008;1224:63–68. doi:10.1016/j.bbi.2018.02.003
55. Aurbach EL, Inui EG, Turner CA, et al. Fibroblast growth factor 9 is a novel modulator of negative affect. Proc Natl Acad Sci USA. 2015;112(38):11953–11958. doi:10.1111/bjd.14546
56. Xu Y-H, Zhu Y, Zhu YY, et al. Abnormalities in FGF family members and their roles in modulating depression-related molecules. Eur J Neurosci. 2021;53(1):140–150. doi:10.1172/JCI129269
57. Rollman O, Jensen UB, Östman A, et al. Platelet Derived Growth Factor (PDGF) responsive epidermis formed from human keratinocytes transduced with the PDGFβ receptor gene. J Invest Dermatol. 2003;120(5):742–749. doi:10.1093/nar/gky1131
58. Jha MK, Minhajuddin A, Gadad BS, Trivedi MH. Platelet-derived growth factor as an antidepressant treatment selection biomarker: higher levels selectively predict better outcomes with bupropion-SSRI combination. Int J Neuropsychopharmacol. 2017;20(11):919–927. doi:10.1016/S0140-6736(07)61128-3
59. Sharma J, Larkin J
60. Yu C-F, Peng W-M, Schlee M, et al. SOCS1 and SOCS3 target IRF7 degradation to suppress TLR7-mediated type I IFN production of human plasmacytoid dendritic cells. J Immunol. 2018;200(12):4024–4035. doi:10.4049/jimmunol.1700510
61. Kobayashi N, Shinagawa S, Nagata T, Shigeta M, Kondo K. Suppressors of cytokine signaling are decreased in major depressive disorder patients. J Pers Med. 2022;12(7):1040. doi:10.3390/jpm12071040
62. Kromer C, Mohr J, Celis D, et al. Screening for depression in psoriasis patients during a dermatological consultation: a first step towards treatment. J Dtsch Dermatol Ges. 2021;19:1451–1461. doi:10.1111/ddg.14532
63. Gerdes S, Wilsmann-Theis D, Celis D, Kromer C, Mossner R. Two questions may be enough - screening for depression in patients with psoriasis: a multicenter study. J Dtsch Dermatol Ges. 2020;18:1115–1125. doi:10.1111/ddg.14203
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.