")
Back to Journals » International Journal of Nanomedicine » Volume 18
Therapeutic Potential of Nanomedicine in Management of Alzheimer’s Disease and Glioma
Authors Anwar F, Al-Abbasi FA , Naqvi S, Sheikh RA , Alhayyani S, Asseri AH, Asar TO, Kumar V
Received 23 January 2023
Accepted for publication 28 April 2023
Published 22 May 2023 Volume 2023:18 Pages 2737—2756
DOI https://doi.org/10.2147/IJN.S405454
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yan Shen
Firoz Anwar,1 Fahad A Al-Abbasi,1 Salma Naqvi,2 Ryan Adnan Sheikh,1 Sultan Alhayyani,3 Amer H Asseri,1 Turky Omar Asar,1 Vikas Kumar4
1Department of Biochemistry, Faculty of Science, King Abdulaziz University, Jeddah, Saudi Arabia; 2Department of Biomedical Sciences, College of Medicine, Gulf Medical University, Ajman, United Arab Emirates; 3Department of Chemistry, College of Sciences & Arts, Rabigh King Abdulaziz University, Jeddah, Saudi Arabia; 4Natural Product Discovery Laboratory, Department of Pharmaceutical Sciences, Shalom Institute of Health and Allied Sciences, SHUATS, Prayagraj, India
Correspondence: Firoz Anwar; Vikas Kumar, Email [email protected]; [email protected]
Abstract: Neoplasm (Glioblastoma) and Alzheimer’s disease (AD) comprise two of the most chronic psychological ailments. Glioblastoma is one of the aggressive and prevalent malignant diseases characterized by rapid growth and invasion resulting from cell migration and degradation of extracellular matrix. While the latter is characterized by extracellular plaques of amyloid and intracellular tangles of tau proteins. Both possess a high degree of resistance to treatment owing to the restricted transport of corresponding drugs to the brain protected by the blood–brain barrier (BBB). Development of optimized therapies using advanced technologies is a great need of today. One such approach is the designing of nanoparticles (NPs) to facilitate the drug delivery at the target site. The present article elaborates the advances in nanomedicines in treatment of both AD as well as Gliomas. The intention of this review is to provide an overview of different types of NPs with their physical properties emphasizing their importance in traversing the BBB and hitting the target site. Further, we discuss the therapeutic applications of these NPs along with their specific targets. Multiple overlapping factors with a common pathway in development of AD and Glioblastoma are discussed in details that will assist the readers in developing the conceptual approach to target the NP for an aging population in the given circumstances with limitations of currently designed NPs, and the challenges to meet and the future perspectives.
Keywords: therapeutic potential, Alzheimer’s disease, glioma, nanomedicine, blood–brain barrier
Introduction
Neoplasia and Alzheimer’s disease (AD) comprise two of the most chronic psychological ailments. Age is a major risk factor associated with the deterioration of psychological functions in both diseases.1,2 Multiple factors including uncontrolled proliferative signals, downregulation of growth suppressors, development of immortal characters, resistance to apoptosis, development of angiogenesis, activation of invasion and metastasis are a few hallmarks of neoplasm.3 Further epigenetics alterations, genomic instability, avoidance of immune destruction, tumor microenvironment and inflammation associated with reactive oxygen species are other such markers associated with cancer.4,5 Alzheimer’s disease is one of the most common neurodegenerative disorders in the aged population, affecting about 36 million people around the globe and projected to impact 115 million people by the year 2050.6 Clinical manifestations include progressive dysfunction and loss of neurons, histological alterations, marked by the presence of intracellular tangles of neurofibrils along with extracellular amyloid plaques with reduced cognition functions,7,8 further characterized by loss of synaptic plasticity, misfold of amyloid β (Aβ) and Tau, hyperphosphorylated at various sites.9,10 Progressive and spontaneous aggregation of Aβ forming oligomers and fibrils with final deposition of senile plaques are the main products responsible for memory deficit and synaptic damage in AD patients.11,12 Along with Aβ proteins, metabolomics, proteomics and genomic studies have identified various markers that can predict disease development and progression from mild cognitive impairment (MCI) in AD.13,14 The multivalent cations in the blood plasma, including zinc, copper and iron, are important factors besides markers in the diagnosis of AD.15
Among all brain disorders from the family of cancer disease, Glioblastoma (GBM) (a type of Glioma) is one of the aggressive and prevalent types of malignant disease.16 Overuse of statins,17 hormonal including contraceptive pills and reproductive factors are associated with increased incidence of Glioma.18 Compared to other malignant tumours, very little progress is made in its clinical outcome due to limitations in the effective drug delivery mechanism.19 High invasiveness,20 frequent recurrence 21 and increased mortality rates22 made the treatment of Glioma a biggest challenge to neuro health scientists. The present therapeutic approach is limited to a combination of radiotherapy, chemotherapy and surgical resection.23 Though researchers have tried cancer-selective cell killing by boron neutron capture therapy (BNCT) it is still in the juvenile phase before it can be completely used in humans.24 Incomplete or ineffective treatment of Glioma can infiltrate the residual cells to penetrate the other parts of the brain, making the survival time-limited to 12–15 months. In one cohort study, Glioma (GBM) patients survive up to 5 years and only 0.7% of them can live to 10 years,25 making treatment of GBM one of the non-competitive trials for researchers with advanced drug delivery technology. Many signaling pathways are associated with GBM but the most important among all is the signal transducer and activator of transcription 3 (STAT3) pathway,26 involved in cancer proliferation, invasion and progression27 along with evasion to the immune system.28 The properties of evasion to the immune system and increased proliferation are assisted by cytokines like interleukin (IL)-6 and growth factors such as epidermal growth factor (EGF) and fibroblast growth factor (FGF) that can activate STAT329,30 through tyrosine phosphorylation.31 The activated STAT3 increases the expression of all genes that are involved in cell proliferation, inhibition of apoptosis and metastasis.32–34 Further, STAT3 is also associated with stemness and cell death of GBM.35
Inverse comorbidity between cancer and AD has been reported in many clinical and epidemiological studies. A transcriptomic meta-analysis of AD and cancer reported significant overlapping factors in association with genes enough to establish the relation between the two disorders.36–38 Despite advancements in technology and mammoth efforts, present diagnostic and therapeutic options are limited and ineffective in the treatment and prevention of AD and Glioma, making them a high-risk disorder for pharmaceutical and health scientists. Effective and safe development of a new strategy is paramount to understanding the etiology and molecular physiology involved in pathogenesis that can target the new drug entity. The underlying factor for a limited option in the treatment of AD is the presence of the blood–brain barrier (BBB),38 which protects the brain tissues from all toxic and perilous substances in the blood, retarding the activity of pharmaceutical compounds.39 The protection and control of solute movement toward the brain are strictly governed by the BBB, composed of basal membrane, neurons, pericytes, astrocytes, tight junctions and microvascular endothelial cells.40,41 The limitations of various molecules are very strict to cross the BBB including that the molecular weight should be ˂500 Da,42 with a varying degree of brain to plasma partition coefficient,43 high lipid solubility and non-charge at physiological pH.
The permeability to the BBB is dependent on the age factors and it is altered in AD both in structure and functions.44 The limited options and age of the patients prompted health scientists to develop on an urgent basis a new and effective drug delivery mechanism, that can easily cross the BBB, have minimum adverse effects and maximum bioavailability for treatment of AD. To overcome the limitation of the conventional approach, nano drug carriers were designed to deliver the therapeutic agent at the required site.45 This limitation of drug delivery therapy for AD and Gliomas can be overcome by nanotechnology in providing a better option and strategy in the field of CNS related diseases, and further the high biocompatibility, low toxicity and stability in the blood can be better hope in the field of therapeutics and for the pharmaceutical industry. Nanoparticles (NPs) facilitate the delivery of drugs to the brain with proper modification required by brain tissue. Table 1 details the selected NPs under investigation potential to cross the BBB. The present review provides some prospective application of nanomedicine in the treatment of AD and Glioma.
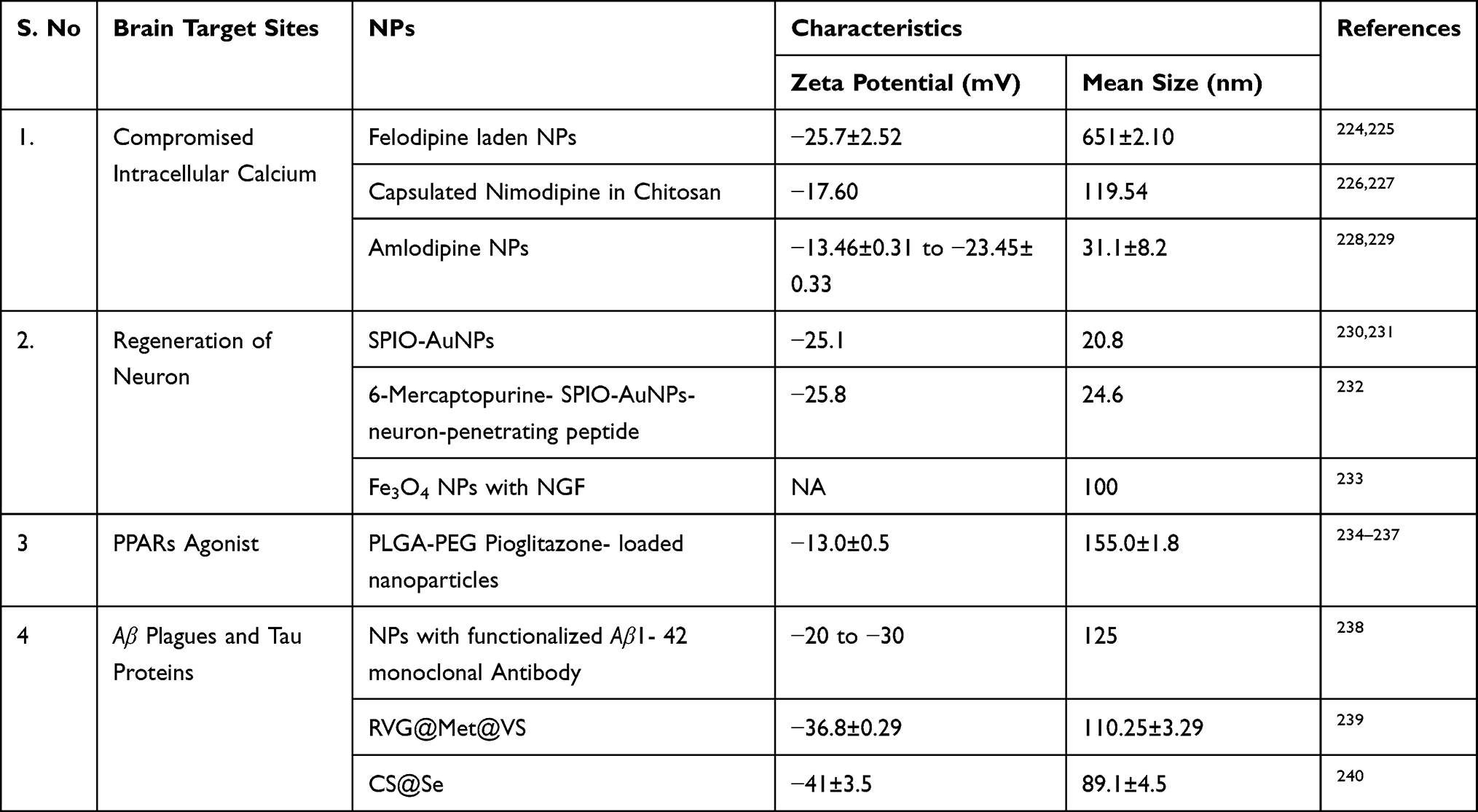 |
Table 1 Summary of Application of Drug Nanoparticles Delivery System with Characteristics in Crossing the BBB to Various Brain Targets |
Overlapping Biological Molecules Between Glioma and Alzheimer Disease
Tumor suppressor p53 contributes to around 50% of all malignancies46 including Glioma. The mutation in p53 facilitates angiogenesis,47 genomic instability,48 progression of cell cycle, cell survival and escape of cell death,49–51 migration and invasion,52 anchorage independence survival and growth.53 Further, it alters impaired detoxification of reactive oxygen species (ROS) via decreasing Phase 2 ROS-detoxifying enzymes, quinone oxidoreductase 1 (NQ01) and heme oxygenase-1 (HO-1), thus resulting in imbalanced redox homeostasis.54,55 In contrast to cancer, p53 expression increases in Alzheimer’s disease,56 and promotes apoptotic neural cell death.57,58 Accumulation of Aβ level with increased expression of mutated amyloid precursor protein/presenilin (APP/PS) strongly supports the correlation between p53 and AD59 in the transgenic mice model. Further, functionally altered tertiary structure, called conformational mutant p53, is distinctly observed in AD patients.60 It is reported that the expression of triggering receptor expressed on myeloid cells 2 (TREM2) in AD is regulated by an altered level of p53.61 Impaired redox status of Superoxide Dismutase (SOD) and Glutathione Reductase in neurodegenerative diseases like AD62 corresponds to an increase in the level of unfolded p53,63 which strongly suggests a possible role of ROS in conformational changes of this gene in AD patients.
Cyclins, the cell cycle regulators in the dysregulated state, lead to cancer initiation and progression, through cyclin-dependent kinase (CKDs) in humans.64,65 In addition to the cell cycle regulation, Cyclins also modulate and regulate the functions of terminally differentiated neurons, thereby imparting a significant contribution in the maintenance of the normal physiology of neurons.66 Most extensively studied are Cyclins D, E, F and Y for their role in human diseases. Cyclin D acts as a checkpoint in the cell cycle,67 controlling the entry of cells from the G0 to G1 phase in Glioma via CKD2/4/6.67 Cyclin D mutant mice were resistant to cancer via inactivation of CKD 4/6.68,69 Cyclin D knockdown induced oxidative imbalance in cancer cells by high ROS generation, which promoted the senescence of cancer cells, making it one of the essential targets for Glioma therapy,70 and besides this, the brains of AD patients have demonstrated high levels of CKD4. Studies have reported that Cyclin D upregulation in AD patients is associated with tau and caspase 3 proteins in cultured hippocampal neurons that are responsible for apoptosis.71 Recently, it has been deciphered that Cyclin D/CKD4-mediated ROS alters mitochondrial functions and facilitates neurodegeneration in AD.72
Cyclin E, a subunit of CDK2, is essential for DNA replication at G1/S checkpoints.73 Its over-expression in breast cancer,74 gastric cancer75 and many other neoplasms76,77 including Glioma78 causes genomic instability.79 Ubiquitin specific peptidase 27 (USP27), a novel therapeutic molecule, targets Cyclin E and retards the migration and metastasis of cancer cells.80,81 Its expression in AD regulates synaptic plasticity and memory formation,82 with induction of cell cycle activation in a Drosophila tauopathy model of AD.83 However, deficiency of Cyclin E reduced spine volume and synapses and potentiated the memory impairment84 key factors in AD pathogenesis.
Cyclin F (FBXO1), a motif of F box proteins, contributes to proliferation and invasion of cancer cells85 and regulates genome stability through ubiquitin-mediated proteolysis, involved in the production of deoxyribonucleotide triphosphate, centrosome duplication and spindle formation in cancer cells.86,87 Upregulation of Cyclin F under metabolic stress in Glioma inhibits tumorigenesis via mutation in isocitrate dehydrogenase-1,88,89 which makes it a potential target for nanomedicine. Missense mutations in the Cyclin F gene are causative of amyotrophic lateral sclerosis (ALS) – a motor neuron disease characterized by a decline in motor functions, due to its binding with valosin, a protein essential for the normal activity of motor neurons;90 with no specific underlying mechanism still to be deciphered, it is a potential candidate for further investigation to understand its relevance in AD and other neurodegenerative diseases.
Intercellular Communication Between Glioma and Alzheimer Disease
As discussed above, the intracellular molecules p53 and Cyclins have a significant contribution in maintaining a normal homeostatic pathway; any deregulation in these molecules may lead to Glioma and AD. Some research suggests that Glioma and AD can affect each other through intracellular molecules, which complicates the treatment of the two diseases. Recent studies demonstrated that Glioma cells secrete excessive glutamate via cystine/glutamate antiporter xCT,91,92 thereby changing the microenvironment of neurons in the vicinity of Glioma, resulting in neuronal degeneration and death.93,94 Glioma cells implanted in striata of experimental animals enhanced the release of glutamate causing rapid growth of Glioma and neuronal degeneration in the vicinity.95 Neuronal degradation and Glioma formation was countered by blocking the glutamate and N-methyl-
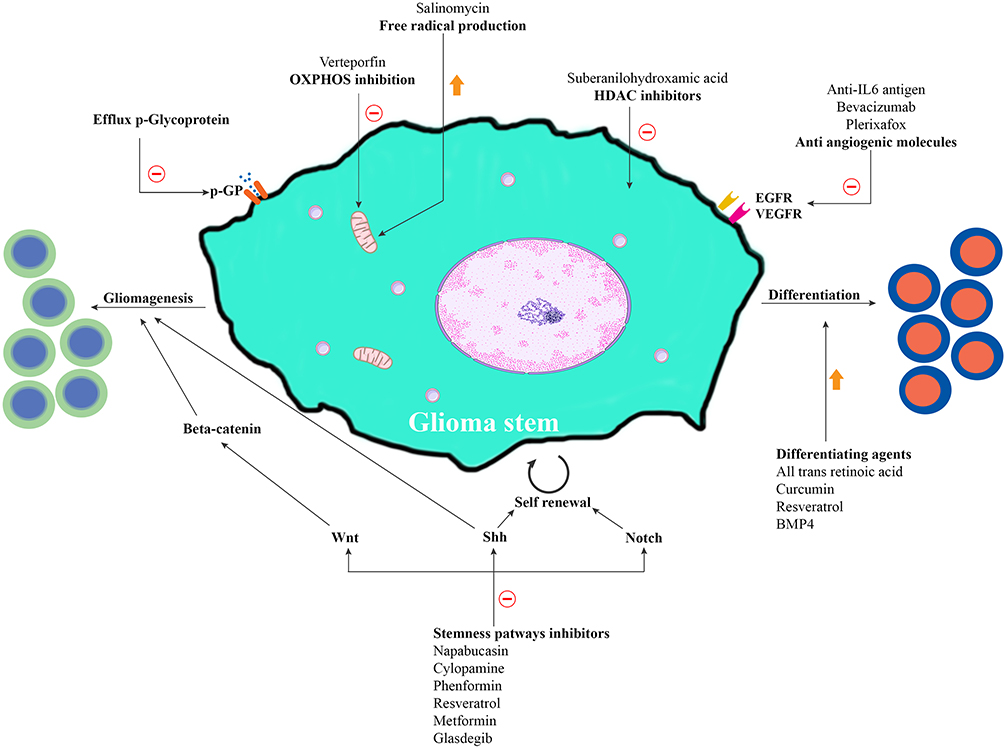 |
Figure 1 Possible approaches and their mechanisms that can probably eliminate Glioma stem cells. Glioma tumor cells and their metastases originate from stem cells possessing self-renewal and differentiation properties. Self-renewal is attributed to activation of alternative pathways like Wnt, Shh and Notch. Targeting these stemness pathways can eliminate Glioma stem cells. HDAC (Histone deacetylase) enzymes catalyze the deacetylation of histones, facilitate chromatin condensation and are associated with oncogenic transcription factors. HDAC inhibitors may target these enzymes and alter gene transcription. Cancer stem cells exhibit overexpression of OXPHOS (oxidative phosphorylation), which plays a key role in cellular energy. They use stored energy in mitochondrial ATP and generate free radicals, ROS (reactive oxygen species). |
Therapeutic Targets of NPs in Alzheimer Disease
The advancement in medical science has increased the life expectancy and consequently the prevalence of neurodegenerative diseases including AD. All the present treatments available today are effective but with limitations, thus scaling the complications of AD with age. Multiple molecular and cellular pathways overlap with each other that ultimately lead to neuronal apoptosis.104 Apoptosis, autophagy dysfunction, pathogenic proteins, impairment, oxidative damage and inflammatory processes are a few contributing factors for all neurodegenerative diseases.105 Inflammation and oxidative stress are interdependent and linked together for neurodegeneration. Generation and elimination of reactive oxygen species (ROS) both from exogenous and exogeneous sources play a crucial role in maintaining the redox balance.106 Inflammatory crosstalk between periphery and central nervous system via the blood–brain barrier is observed in Alzheimer’s disease particularly involving cathepsin.107 Activation and dysfunction of microglial disturbs the brain homeostasis, that directly enhances phagocytosis, increases proinflammatory cytokine secretion and increases the release of ROS.108 It is observed that lipid dysfunction or dyshomeostasis disturbs the regulation of microglial cells due to alteration in phosphoinositides (PiPs), a key molecule in regulation of neuroinflammation. Further, PiPs also regulate the activities of proteins and enzymes essential for Toll-like receptor signaling, endocytosis, purinergic signaling migration and chemotaxis,109,110 a possible reason for alteration in AD physiology. NPs with 1–100 nm of dimensions can easily traverse through the BBB and prevent aggregation of proteins, reduce inflammation and alleviate stress.
Specific Targets Linked to Aβ
Apart from the BBB, the brain parenchyma is the other major obstacle in the delivery of drugs in AD. The parenchymal cells reduce the effective drug concentration at the amyloid plaques, the specific target site in AD patients, thereby reducing its therapeutic value and efficacy.111 Spontaneous aggregation of Aβ monomers leads to the generation of fibrils and oligomers,112,113 a phenomenon causing neuronal malfunction and death.114 The interaction or Aβ monomers aggregation may be prevented by drugs that block such reactions. NPs in the form of liposomes and PEG-PLA have been used as conjugates to prevent this aggregation.115 KLVFF peptide is known to interfere with the Aβ aggregation,116 but its inability to cross the BBB and poor bioavailability have retarded its use. However, when loaded in polymeric nanoparticles, KLVFF gave promising results in reversing Aβ-induced pathology in AD. Likewise, nano-forms of Epigallocatechin–gallate resulted in sustained release of the drug that significantly inhibited Aβ42 protein and reduced cellular toxicity from metallic elements.117,118 Liposomes have attracted great attention in transportation of drugs in AD. Curcumin embedded anti-TrF liposomes have shown a high affinity for amyloid deposits in brain samples of AD patients.119 Similarly, in a mouse model of AD nasal administration of Quercetin,120 liposomes attenuated degeneration of cortical and cholinergic neurons in the hippocampus.121,122 Resveratrol is reported to exhibit the neuroprotective function in AD.123 Liposome formulation of resveratrol, in the treatment of AD, is now well documented.124 Immunotherapy using nano formulation with antibodies is receiving great attention in the treatment of AD.125,126 Immunoliposomes formulated as polyethylene glycol can act directly against the glial fibrillary acid protein.127 Monoclonal antibodies that can target the Aβ protein in NPs are under the state of research and are showing some promising results in AD patients.128 Graphene quantum dots (GQDs) and carbon nanomaterial are the two newly introduced nanomaterials which have shown some promising results in treatment of AD when combined with scavenging materials of peptide.129,130 The charge on the graphene plays a vital role in inhibition of fibril formation, the charge may be transferred to the aromatic residue of protein amino acid.131 Carbon dots (CDs) have demonstrated potential to cross the BBB, due to presence of amino and carboxylic acid group on their surface that can be conjugated with CNS drugs,132 making them an ideal nanocarrier to deliver the drugs in CNS to treat glioma and AD. Yellow-emissive CDs and graphene quantum dots were able to prevent the aggregation of Aβ in neuronal cells linked to tramiprosate.133,134 Similar results were obtained by Gong et al in glycine proline-glutamate loaded CDs to inhibit Aβ aggregation.135 Identical results were observed when the branched PEI loaded CDs synthesized by Chung et al exhibited cationic surface and were able to suppress the aggregation of Aβ.136 Preclinical and clinical research have demonstrated that some of the metallic ions including iron, zinc and copper play an important role in manifestation of AD137 with increased concentration above a certain limit. This can increase the deposition of Aβ138 and can promote the progression of disease.139 Zinc loaded nanoparticles in wild type (WT) and APP23 mice model alters the pathological conditions in the mice model by significant effect on proinflammatory cytokines IL-6 and IL-18 and reduction in plague size.140 Selenium-loaded nanoparticles with penicillamine can act as Aβ inhibitor, with no major toxicological effect on organs and systemic toxicity, making them an important product for biomedical use.141 Naresh et al successfully developed patient-friendly long-acting donepezil nanocrystals formulation, with a high payload for i.m administration, detectable even after 18 days in blood with improved spatial memory learning.142 Similar results were obtained for fabricated ApoE3 coated polymeric nanoparticles, enhancing the uptake of donepezil nanocarrier through oral delivery in treatment of AD.143 Drugs like rivastigmine formulated in novel
Specific Targets Linked to Aβ Production
Deregulation or dysregulation of β and γ secretase can lead to overproduction of Aβ protein, making a significant contribution to the etiology of AD.146 These enzymes can be appropriate pharmacological targets to develop new strategies for the management of AD. However, due to the broad range of proteolytic activity of these enzymes, the inhibition can favor the undesired adverse reactions or effects.147 To target β secretase, a new concept of RNA interference small interfering RNA (siRNA) was developed with great promising results on AD in the nanoform.148 They can directly block the causative gene expression with high targeting specificity, in low doses with a simple drug development process.149 The major challenge for the siRNA in the treatment of AD is their delivery via systemic circulation that can cross the BBB, overcome enzymatic degradation, cell endocytosis and impaired cytosolic transport along with short circulation time. Present nano technology has great potential to overcome these barriers. In a recent study BACE1 siRNA to mouse brain through systemic injection has partially reduced AD neuropathology with low therapeutic efficacy.150,151 The delivery was made through glycosylated NP siRNA, in transgenic mice targeting BACE1, which has a better potential for clinical translation. Exosomes are naturally occurring NPs with a diameter of 40–100 nm152 and loaded with siRNA against BACE1, these exosomes altered the expression and production of Aβ proteins in a transgenic mice model.153
Specific Targets Linked to Aβ Dispensation/Clearance
Aβ plaques and neurofibrillary tangles are the hallmark of neuropathological lesions of AD. Aβ immunotherapy was able to reduce both extracellular Aβ plaques and intracellular accumulation also leading to a reduction in tau pathology,154 indicating a direct correlation between accumulation of Aβ and tau155, where clearance is mediated by the proteasome and is associated with phosphorylation.156 In vivo antigens are prepared that can mimic the Aβ proteins, Abs targeting these antigens are products that can bind to cerebral Aβ and facilitate their dispense.157 After obtaining promising results at a preclinical level in animals, its translation into humans resulted in severe adverse effects including vasogenic edema, intracellular microhemorrhages and T cell-mediated meningoencephalopathy.158 Furthermore, Apolipoprotein (ApoE) and its isomeric forms APOE3 or APOE2 play a critical role in pathogenesis of neurodegenerative disease including AD risk.159 The incomplete structural information of ApoE limits its role in understanding the pathogenesis of AD. Single amino acid substitution of ApoE2 and ApoE4 differs from ApoE3, resulting in different impact on risk of disease and its outcome.160 The binding or interaction of ApoE proteins with Aβ, tau, and α-synuclein alters the response of brain to these aggregates.161,162 Lipidation of ApoE and the conformational changes that occurs in ApoE on the lipid surface is essential for its binding with the ApoE receptors.163 The interdomain interaction within ApoE is an essential driving factor for specific isoenzyme difference of activity including Aβ, the biochemical data suggest and indicate non-lapidated ApoE undergoes dimerization and tetramerization at higher concentration for effective pathological activity.164 Present techniques are insufficient to elucidate the exact interaction of ApoE with Aβ that can eliminate these proteins responsible for AD, however if isoform specific structures related to lipidation and non-lipidation of ApoE complex are solved then newer drugs can be designed that can directly modulate the ApoE–receptor and ApoE–protein interaction at the molecular and submolecular level.165 Structure alteration of ApoE has already shown potential to alleviate the toxic effects of ApoE.166 In order to counter the adverse effects of conventional immunotherapy, NPs could give a better advantage over it. Antibodies designed for Aβ are trapped in NPs, and deliver to specific targets.167 The studies have demonstrated partial fragments of Aβ consisting of 15 amino acids formulated with PLGA have shown full response toward complete Aβ plague proteins via subcutaneous or intranasal route, with minimum toxicity.168 Still, the delivery of antibodies or antigen for the treatment of AD is in the juvenile stage but gives better hope for AD patients if successful in human trials.
Therapeutic NPs in Management of Glioma/GBM
Preclinical studies on GBM models with NPs emerged with certain advantages compared to their soluble counterparts. Polymeric NPs can easily trap the drug molecule intended for GBM therapy and can exert the required effect on target tissue. NPs are either synthetic like PCL, PLA and PLGA with biodegradable and compatible properties169,170 or can be natural, viz albumin, chitosan or gelatin.171 The NPs are modified in order to generate the effective therapeutic concentration in the brain due to presence of macrophage in the liver and spleen that can engulf them.172 Bioavailability and distribution of NPs in the brain is enhanced by use of hydrophilic surfactant with an increase in half-life.173 Anticancer drugs like doxorubicin coated with Tween 80 as surfactant, in the form of NPs formulated from poly(n-butyl cyanoacrylate) (PBCA) (270±20 nm) exhibited potential therapeutic effect on GBM.174 PBCA NPs loaded with doxorubicin have increased the survival time by 85% compared to the untreated control 24% where drug was administered in solution form without NPs, and further without the Tween 80 the survival rate was 38% only.175 It was important to investigate the toxicological profile of DOX-loaded PBCA NPs (240±40 nm d.; injected IV) and DOX-loaded HAS (404±24 nm d.; injected IV) on healthy animals; both of the NP formulations were less toxic to cardiac and testicular tissues compared to DOX injection after 15 and 30 days respectively.176,177 Drug concentration in the brain was enhanced many fold administered in PBCA NP form176 compared to uncoated formulations. In spite of these encouraging results the specificity of the drugs to target the GBM remains a major challenge to health scientists.
Specific Target Sites in GSM
Targeted nanomedicines possess a unique advantage over a non-targeted form, increasing the amount of drug at cancer cells reducing the concentration at healthy cells.178 The target site achievement can be initiated by addition of target agent in the form of an antibody or ligand that selectively binds to a specific site or receptor on the cancer cells179 through endocytosis facilitating the cellular uptake of the cytotoxic agent.180 In case of Glioma cells it is the CD133 receptor that can easily bind with the antibody181 specific on them. The conjugation of anti-CD133 antibodies with polymeric dendrimers with mercapto-undecahydro-dodecaborate significantly increased the drug uptake.182 Receptor-mediated targets have great importance in target site delivery of polymeric NP. Transferrin receptors (TfR) are over-expressed in multiple cancers. Anti-transferrin receptor antibodies (anti-TfR) conjugated with resveratrol liposomes reduced the growth of Glioma cells.183 Paclitaxel loaded liposomes using arginine–glycine aspartic acid were able to initiate the excellent apoptosis on a Glioma cell line by binding to TfR receptor.184 Although much advancement and effort has been made to specifically target the tumor cells in the brain, limitations of in vivo results made the development of CDs (CD-Asp) with
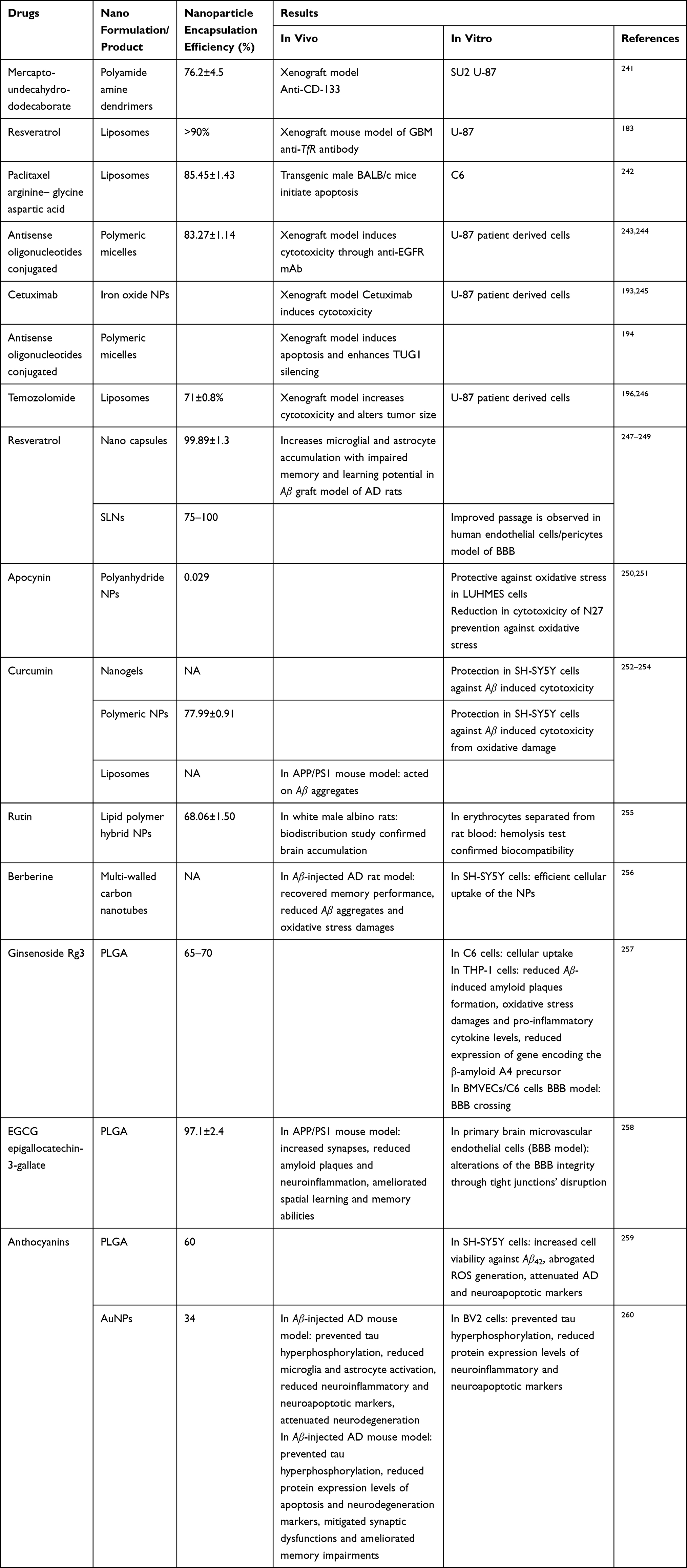 |
Table 2 Summary of In Vivo and In Vitro with Drug Nano Formulation Demonstrating the Encapsulation Efficiency in Treatment of Glioma and AD |
 |
Figure 2 Mechanisms of overlapping fragments in cancer and Alzheimer’s disease. Cyclin D1 endorses tau phosphorylation in presence of GSK3β (which is again dephosphorylated by PP2A), and induces apoptosis through a Caspase-3-mediated pathway. Reduced activity of SOD and GR tend to increase ROS production, which causes a conformational change in p53 by unfolding it. This unfolded p53 is also observed in Alzheimer’s disease. Mutant p53 decreases the expression of NQO1 and HO-1, the ROS- detoxifying enzymes, and thus induces ROS production. LMW cyclin E forms a complex with CDK2 in the cytoplasm, and activates oncogenic functions like cell invasion and metastasis. APP produces Aβ proteins, Aβ fibrils and plaques. Abbreviations: APP, Amyloid precursor proteins; Aβ, Amyloid β; CDK2, Cyclin-dependent Kinase 2; GR, Glutathione Reductase; GSK3β, Glycogen synthase kinase 3β; HO1, Heme Oxygenase 1; LMW, Low molecular weight; NQO1, Quinine-oxidoreductase 1; PP2A, Protein phosphatase 2A; ROS, Reactive Oxygen Species; SOD, Superoxide Dismutase. |
Multiple factors both intrinsic and extrinsic like high tumor heterogenicity, drug resistance, invasiveness, and targetable mutation are responsible for ineffective GB therapy; further, the design of drug delivery plays a major important role in crossing the BBB that can be specific to tumor site. In view to overcome these limitations Novel design of NPs has given new hope in effective treatment of GBM. Much of the NPs are already in various phase of clinical trials. Oligonucleotides (ONTs) are able to target the oncogenic mechanism delivered in the form of p53 mRNA or PTEN siRNA overcoming the limitations of tumor heterogeneity.204 Integrins and ApoE are targeted by Dox due to common EGFR by EGFR(V) antibody conjugated to an EnGeneIC delivery vehicle (EDV), loaded with DOX (EGFR(V)-EDV-Dox).205 A similar approach was reserved with Pseudomonas exotoxin with EGFR-targeted, convection enhanced delivery system.206 Proteins such as Selectins are found to express both on brain endothelial and glioma cells, and NPs loaded with doxorubicin or other chemotherapeutic agent possess a tyrosine kinase inhibition potential,207,208 that may improve treatment results with reduction in cell resistance.
It is still a topic of debate whether nano-formulations can eradicate Glioma and AD compared to conventional therapy.45 This system of delivery is much safer with reduced toxicity compared to conventional therapy.209 Drugs in the nano-formulation are known to improve saturation and maintain or enhanced permeability and retention effect (EPR) along with the concentration at the site of tumor with increase in retention time.210 The underlying mechanism with EPR is associated with rapid growth of tumor, blood vessels in a leaky state and low organized structure of blood vessels further the inefficient lymphatic drainage.211 It is observed that enhanced potency of 1,3 β-Glucan as an outer shell to chitosan nanoparticles loaded with paclitaxel can prevent hemolysis enabling effective therapeutic advantage against glioblastoma, thus overcoming the systemic toxicities due to paclitaxel alone with increased bioavailability.212 The safety profile of drugs in nano formulations provides additional advantage compared to free drugs,213 and further the cancer therapeutic is always at risk and disadvantage due to radiation toxicity, drugs like baicalein in its oral nanoform in preclinical evaluation have modulated the radiation response.214 Cytarabine loaded liposomes in phase I/II clinical trials have shown additional safety as compared to free drug215 in patients with secondary glioblastoma. Further, the NPs in case of delivery to glioblastoma gives the protection from enzymatic degradation, metabolism especially in the case of delivery of siRNAs, miRNA and other forms of nucleic acids.216 Therapeutic nucleic acids have been delivered in the form of polymeric NPs, lipid polymer NPs,217 gold NPs218 and superparamagnetic NPs of iron oxide.219 Such types of formulations increase the efficiency of the target drug to the target gene through enhanced internalization that can easily slice glioblastoma related genes, thus prolonging the survival time period of the model animals. Further, it has been observed that such type of delivery has retarded the efflux of medicine by efflux pumps220 in cancer cells including ABC proteins.
Future Direction and Limitations
The majority of these novel drug delivery system results available are only preliminary in vitro or in the mouse model. Many challenges may arise during clinical application of these NPs in humans. A poorly explored aspect is any change in the functional activities of a tissue or cell encountered by the nanoparticles while approaching their target. Also, it is not much reported whether and how the electrical impulse conduction of the neurons targeted by NPs are affected. Further, another important aspect to be considered is that nanomaterials may themselves be cytotoxic and their administration may cause neurotoxicity. Further, as these nanomaterials interfere with BBB integrity, they may create a passage not only for therapeutic drugs, but also favor the entry of toxic substances or pathogens to the brain. Additionally, NPs can interfere with normal cellular metabolism, resulting in increased ROS and altered gene expression. Alhough these challenges are still to be met, extensive research is going on, and every modification in nanotechnologies for drug delivery bypasses the presenting obstacles. Significance of nanoparticle driven drug delivery is increasing. New targets like mutant genes, DNA synthesis, hypoxia, neuroproteins, neuropilin-1, novel therapies including virus-based NPs, protein based NPs and nucleic acid based NPs with more effective penetration across the BBB have a great potential to unfold a promising era in the treatment of AD, glioblastoma as well as other brain diseases. Furthermore, mRNA (particularly non-invasive PTEN mRNA221) targeting Orthotopic Glioblastoma222 for prophylactic and therapeutics applications in the form of NPs have potential to change the course of many diseases including AD and Glioma.223
However, the bottom line still states that NPs need much more extensive research before they can be therapeutically used in humans, without any doubt of their drawbacks.
Conclusion
This article is crosstalk between nanoparticles with promising insight for the two diseases AD and glioblastoma with completely different pathology, where AD results from neuron degeneration while glioblastoma is characterized by rapid cell multiplication; however, the factor common in both is that their treatment is very difficult and unspecific. Development of nanoparticles loaded with drugs has provided a favorable approach to target and release the drugs at amyloid plaques, Aβ, the pathological site in AD patients. Similarly, the drugs in NPs can reach the brain parenchyma by EPR effect, an effective breakthrough in the treatment of glioblastoma. The particle size and physical properties of NPs are essential parameters that influence the penetration through biological membranes in order to obtain the best therapeutic effects of NPs.
Acknowledgment
This project was funded by the Deanship of Scientific Research (DSR) at King Abdulaziz University, Jeddah, under grant no. IFPRP: 148-130-1442. The authors, therefore, acknowledge with thanks the DSR for their technical and financial support.
Disclosure
The authors declare that they have no competing interests.
References
1. Prasad K. AGE–RAGE stress: A changing landscape in pathology and treatment of Alzheimer’s disease. Mol Cell Biochem. 2019;459(1):95–112. doi:10.1007/s11010-019-03553-4
2. Ladomersky E, Scholtens DM, Kocherginsky M, et al. The coincidence between increasing age, immunosuppression, and the incidence of patients with glioblastoma. Front Pharmacol. 2019;10:200. doi:10.3389/fphar.2019.00200
3. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. cell. 2011;144(5):646–674. doi:10.1016/j.cell.2011.02.013
4. Ziech D, Franco R, Pappa A, et al. Reactive Oxygen Species (ROS)––Induced genetic and epigenetic alterations in human carcinogenesis. Mutat Res. 2011;711(1–2):167–173. doi:10.1016/j.mrfmmm.2011.02.015
5. Nikitovic D, Tzardi M, Berdiaki A, et al. Cancer microenvironment and inflammation: role of hyaluronan. Front Immunol. 2015;6:169. doi:10.3389/fimmu.2015.00169
6. Franceschi AM. Hybrid PET/MR Neuroimaging: A Comprehensive Approach. Springer Nature; 2022.
7. Cheignon CM, Tomas M, Bonnefont-Rousselot D, et al. Oxidative stress and the amyloid beta peptide in Alzheimer’s disease. Redox Biol. 2018;14(p):450–464. doi:10.1016/j.redox.2017.10.014
8. Matej R, Tesar A, Rusina R. Alzheimer’s disease and other neurodegenerative dementias in comorbidity: a clinical and neuropathological overview. Clin Biochem. 2019;73:26–31. doi:10.1016/j.clinbiochem.2019.08.005
9. Balschun D, Rowan MJ. Hippocampal synaptic plasticity in neurodegenerative diseases: Amyloid-Β, tau and beyond that. Neuroforum. 2018;24(3):203–212. doi:10.1515/nf-2017-0063
10. Busche MA, Hyman BT. Synergy between amyloid-β and tau in Alzheimer’s disease. Nat Neurosci. 2020;23(10):1183–1193. doi:10.1038/s41593-020-0687-6
11. Balducci C, Beeg M, Stravalaci M, et al. Synthetic amyloid-β oligomers impair long-term memory independently of cellular prion protein. Proc Natl Acad Sci. 2010;107(5):2295–2300. doi:10.1073/pnas.0911829107
12. Viola KL, Klein WL. Amyloid β oligomers in Alzheimer’s disease pathogenesis, treatment, and diagnosis. Acta Neuropathol. 2015;129(2):183–206. doi:10.1007/s00401-015-1386-3
13. Saykin AJ, Shen L, Yao X, et al. Genetic studies of quantitative MCI and AD phenotypes in ADNI: Progress, opportunities, and plans. Alzheimers Dement. 2015;11(7):792–814. doi:10.1016/j.jalz.2015.05.009
14. Azhdarzadeh M, Noroozian M, Aghaverdi H, et al. Serum Multivalent Cationic Pattern: Speculation on the Efficient Approach for Detection of Alzheimer’s Disease. Sci Rep. 2013;3(1):1–6. doi:10.1038/srep02782
15. Nie J. Exposure to aluminum in daily life and Alzheimer’s disease, in Neurotoxicity of Aluminum. Springer; 2018:99–111.
16. Ladomersky E, Zhai L, Lauing KL, et al. Advanced age increases immunosuppression in the brain and decreases immunotherapeutic efficacy in subjects with glioblastoma. Clin Cancer Res. 2020;26(19):5232–5245. doi:10.1158/1078-0432.CCR-19-3874
17. Gaist D, Andersen L, Hallas J, et al. Use of statins and risk of glioma: a nationwide case–control study in Denmark. Br J Cancer. 2013;108(3):715–720. doi:10.1038/bjc.2012.536
18. Silvera SAN, Miller AB, Rohan TE. Hormonal and reproductive factors and risk of glioma: a prospective cohort study. Int J Cancer. 2006;118(5):1321–1324. doi:10.1002/ijc.21467
19. Cha GD, Kang T, Baik S, et al. Advances in drug delivery technology for the treatment of glioblastoma multiforme. J Control Release. 2020;328:350–367. doi:10.1016/j.jconrel.2020.09.002
20. Mecca C, Giambanco I, Bruscoli S, et al. PP242 counteracts glioblastoma cell proliferation, migration, invasiveness and stemness properties by inhibiting mTORC2/AKT. Front Cell Neurosci. 2018;12:99. doi:10.3389/fncel.2018.00099
21. Scoccianti S, Francolini G, Carta GA, et al. Re-irradiation as salvage treatment in recurrent glioblastoma: a comprehensive literature review to provide practical answers to frequently asked questions. Crit Rev Oncol Hematol. 2018;126(p):80–91. doi:10.1016/j.critrevonc.2018.03.024
22. Zhang P, Xia Q, Liu L, et al. Current opinion on molecular characterization for GBM classification in guiding clinical diagnosis, prognosis, and therapy. Front Mol Biosci. 2020;2020:241.
23. Buszek SM, Al Feghali KA, Elhalawani H, et al. Optimal timing of radiotherapy following gross total or subtotal resection of glioblastoma: a real-world assessment using the National Cancer Database. Sci Rep. 2020;10(1):1–9. doi:10.1038/s41598-020-61701-z
24. He H, Li J, Jiang P, et al. The basis and advances in clinical application of boron neutron capture therapy. Radiat Oncol. 2021;16(1):1–8. doi:10.1186/s13014-021-01939-7
25. Tykocki T, Eltayeb M. Ten-year survival in glioblastoma. A systematic review. J Clin Neurosci. 2018;54:7–13. doi:10.1016/j.jocn.2018.05.002
26. Arora L, Kumar A, Arfuso F, et al. The role of signal transducer and activator of transcription 3 (STAT3) and its targeted inhibition in hematological malignancies. Cancers. 2018;10(9):327. doi:10.3390/cancers10090327
27. Zhang J, Sikka S, Siveen KS, et al. Cardamonin represses proliferation, invasion, and causes apoptosis through the modulation of signal transducer and activator of transcription 3 pathway in prostate cancer. Apoptosis. 2017;22(1):158–168. doi:10.1007/s10495-016-1313-7
28. Grabowski MM, Sankey EW, Ryan KJ, et al. Immune suppression in gliomas. J Neurooncol. 2021;151(1):3–12. doi:10.1007/s11060-020-03483-y
29. West AJ, Tsui V, Stylli SS, et al. The role of interleukin‑6‑STAT3 signalling in glioblastoma. Oncol Lett. 2018;16(4):4095–4104. doi:10.3892/ol.2018.9227
30. Wu J, Feng X, Zhang B, et al. Blocking the bFGF/STAT3 interaction through specific signaling pathways induces apoptosis in glioblastoma cells. J Neurooncol. 2014;120(1):33–41. doi:10.1007/s11060-014-1529-8
31. Birner P, Toumangelova-Uzeir K, Natchev S, et al. STAT3 tyrosine phosphorylation influences survival in glioblastoma. J Neurooncol. 2010;100(3):339–343. doi:10.1007/s11060-010-0195-8
32. Zhang S, Zhao BS, Zhou A, et al. m6A demethylase ALKBH5 maintains tumorigenicity of glioblastoma stem-like cells by sustaining FOXM1 expression and cell proliferation program. Cancer cell. 2017;31(4):591–606. e6. doi:10.1016/j.ccell.2017.02.013
33. Dai Z, Wang L, Wang X, et al. Oxymatrine induces cell cycle arrest and apoptosis and suppresses the invasion of human glioblastoma cells through the EGFR/PI3K/Akt/mTOR signaling pathway and STAT3. Oncol Rep. 2018;40(2):867–876. doi:10.3892/or.2018.6512
34. Mashimo T, Pichumani K, Vemireddy V, et al. Acetate is a bioenergetic substrate for human glioblastoma and brain metastases. Cell. 2014;159(7):1603–1614. doi:10.1016/j.cell.2014.11.025
35. Han D, Yu T, Dong N, et al. Napabucasin, a novel STAT3 inhibitor suppresses proliferation, invasion and stemness of glioblastoma cells. J Exp Clin Cancer Res. 2019;38(1):1–12. doi:10.1186/s13046-019-1289-6
36. Ibáñez K, Boullosa C, Tabarés-Seisdedos R, et al. Molecular evidence for the inverse comorbidity between central nervous system disorders and cancers detected by transcriptomic meta-analyses. PLoS Genet. 2014;10(2):e1004173. doi:10.1371/journal.pgen.1004173
37. Zhang L, Silva TC, Young JI, et al. Epigenome-wide meta-analysis of DNA methylation differences in prefrontal cortex implicates the immune processes in Alzheimer’s disease. Nat Commun. 2020;11(1):1–13. doi:10.1038/s41467-020-19791-w
38. Wan Y-W, Al-Ouran R, Mangleburg CG, et al. Meta-analysis of the Alzheimer’s disease human brain transcriptome and functional dissection in mouse models. Cell Rep. 2020;32(2):107908. doi:10.1016/j.celrep.2020.107908
39. Reichel A. The role of blood-brain barrier studies in the pharmaceutical industry. Curr Drug Metab. 2006;7(2):183–203. doi:10.2174/138920006775541525
40. Campisi M, Shin Y, Osaki T, et al. 3D self-organized microvascular model of the human blood-brain barrier with endothelial cells, pericytes and astrocytes. Biomaterials. 2018;180:117–129. doi:10.1016/j.biomaterials.2018.07.014
41. Jamieson JJ, Linville RM, Ding YY, et al. Role of iPSC-derived pericytes on barrier function of iPSC-derived brain microvascular endothelial cells in 2D and 3D. Fluids Barriers CNS. 2019;16(1):1–16. doi:10.1186/s12987-019-0136-7
42. Gosselet F, Loiola RA, Roig A, et al. Central nervous system delivery of molecules across the blood-brain barrier. Neurochem Int. 2021;144:104952. doi:10.1016/j.neuint.2020.104952
43. Loryan I, Reichel A, Feng B, et al. Unbound Brain-to-Plasma Partition Coefficient, Kp, uu, brain—a Game Changing Parameter for CNS Drug Discovery and Development. Pharm Res. 2022;39(7):1321–1341. doi:10.1007/s11095-022-03246-6
44. Pan Y, Nicolazzo JA. Impact of aging, Alzheimer’s disease and Parkinson’s disease on the blood-brain barrier transport of therapeutics. Adv Drug Deliv Rev. 2018;135:62–74. doi:10.1016/j.addr.2018.04.009
45. Afzal M, Alharbi KS, Alruwaili NK, et al. Nanomedicine in treatment of breast cancer–A challenge to conventional therapy. In: Seminars in cancer biology. Elsevier; 2021.
46. Efeyan A, Serrano M. p53: guardian of the genome and policeman of the oncogenes. Cell cycle. 2007;6(9):1006–1010. doi:10.4161/cc.6.9.4211
47. Kamat CD, Green DE, Warnke L, et al. Mutant p53 facilitates pro-angiogenic, hyperproliferative phenotype in response to chronic relative hypoxia. Cancer Lett. 2007;249(2):209–219. doi:10.1016/j.canlet.2006.08.017
48. Fu X, Wu S, Li B, et al. Functions of p53 in pluripotent stem cells. Protein Cell. 2020;11(1):71–78. doi:10.1007/s13238-019-00665-x
49. Vancsik T, Forika G, Balogh A, et al. Modulated electro‐hyperthermia induced p53 driven apoptosis and cell cycle arrest additively support doxorubicin chemotherapy of colorectal cancer in vitro. Cancer Med. 2019;8(9):4292–4303. doi:10.1002/cam4.2330
50. Tsabar M, Mock CS, Venkatachalam V, et al. A switch in p53 dynamics marks cells that escape from DSB-induced cell cycle arrest. Cell Rep. 2020;32(5):107995. doi:10.1016/j.celrep.2020.107995
51. Das S, Shukla N, Singh SS, et al. Mechanism of interaction between autophagy and apoptosis in cancer. Apoptosis. 2021;26(9–10):512–533. doi:10.1007/s10495-021-01687-9
52. Liu L, Yan J, Cao Y, et al. Proliferation, migration and invasion of triple negative breast cancer cells are suppressed by berbamine via the PI3K/Akt/MDM2/p53 and PI3K/Akt/mTOR signaling pathways. Oncol Lett. 2021;21(1):1. doi:10.3892/ol.2020.12262
53. Deng Z, Wang H, Liu J, et al. Comprehensive understanding of anchorage-independent survival and its implication in cancer metastasis. Cell Death Dis. 2021;12(7):1–12. doi:10.1038/s41419-021-03890-7
54. Tauffenberger A, Magistretti PJ. Reactive oxygen species: beyond their reactive behavior. Neurochem Res. 2021;46(1):77–87. doi:10.1007/s11064-020-03208-7
55. Lisek K, Campaner E, Ciani Y, et al. Mutant p53 tunes the NRF2-dependent antioxidant response to support survival of cancer cells. Oncotarget. 2018;9(29):20508. doi:10.18632/oncotarget.24974
56. Jembrek MJ, Slade N, Hof PR, et al. The interactions of p53 with tau and Aß as potential therapeutic targets for Alzheimer’s disease. Prog Neurobiol. 2018;168:104–127. doi:10.1016/j.pneurobio.2018.05.001
57. Erekat NS. Apoptosis and its therapeutic implications in neurodegenerative diseases. Clin Anat. 2022;35(1):65–78. doi:10.1002/ca.23792
58. Tajbakhsh A, Read M, Barreto GE, et al. Apoptotic neurons and amyloid-beta clearance by phagocytosis in Alzheimer’s disease: Pathological mechanisms and therapeutic outlooks. Eur J Pharmacol. 2021;895:173873. doi:10.1016/j.ejphar.2021.173873
59. Singh BK, Vatsa N, Kumar V, et al. Ube3a deficiency inhibits amyloid plaque formation in APPswe/PS1δE9 mouse model of Alzheimer’s disease. Hum Mol Genet. 2017;26(20):4042–4054. doi:10.1093/hmg/ddx295
60. Yang DS, Saeedi A, Davtyan A, et al. Mesoscopic protein-rich clusters host the nucleation of mutant p53 amyloid fibrils. Proc Natl Acad Sci. 2021;118(10):1.
61. Zajkowicz A, Gdowicz-Kłosok A, Krześniak M, et al. The Alzheimer’s disease-associated TREM2 gene is regulated by p53 tumor suppressor protein. Neurosci Lett. 2018;681:62–67. doi:10.1016/j.neulet.2018.05.037
62. Zarrouk A, Hammouda S, Ghzaiel I, et al. Association between oxidative stress and altered cholesterol metabolism in Alzheimer’s disease patients. Curr Alzheimer Res. 2021;17(9):823–834. doi:10.2174/1567205017666201203123046
63. Amor-Gutiérrez O, Costa-Rama E, Arce-Varas N, et al. Competitive electrochemical immunosensor for the detection of unfolded p53 protein in blood as biomarker for Alzheimer’s disease. Anal Chim Acta. 2020;1093:28–34. doi:10.1016/j.aca.2019.09.042
64. Li X, Gong X, Chen J, et al. miR-340 inhibits glioblastoma cell proliferation by suppressing CDK6, cyclin-D1 and cyclin-D2. Biochem Biophys Res Commun. 2015;460(3):670–677. doi:10.1016/j.bbrc.2015.03.088
65. Sittithumcharee G, Suppramote O, Vaeteewoottacharn K, et al. Dependency of Cholangiocarcinoma on Cyclin D–Dependent Kinase Activity. Hepatology. 2019;70(5):1614–1630. doi:10.1002/hep.30704
66. Chu C, Geng Y, Zhou Y, et al. Cyclin E in normal physiology and disease states. Trends Cell Biol. 2021;31(9):732–746. doi:10.1016/j.tcb.2021.05.001
67. Bai J, Li Y, Zhang G. Cell cycle regulation and anticancer drug discovery. Cancer Biol Med. 2017;14(4):348. doi:10.20892/j.issn.2095-3941.2017.0033
68. Tan X, Luo Q, Zhou S, et al. Erchen plus huiyanzhuyu decoction inhibits the growth of laryngeal carcinoma in a mouse model of phlegm-coagulation-blood-stasis syndrome via the STAT3/Cyclin D1 pathway. Evid Based Complement Alternat Med. 2020;2020:1–14. doi:10.1155/2020/2803496
69. Bouclier C, Simon M, Laconde G, et al. Stapled peptide targeting the CDK4/Cyclin D interface combined with Abemaciclib inhibits KRAS mutant lung cancer growth. Theranostics. 2020;10(5):2008. doi:10.7150/thno.40971
70. Laphanuwat P, Likasitwatanakul P, Sittithumcharee G, et al. Cyclin D1 depletion interferes with oxidative balance and promotes cancer cell senescence. J Cell Sci. 2018;131(12):jcs214726. doi:10.1242/jcs.214726
71. Absalon S, Kochanek DM, Raghavan V, et al. MiR-26b, upregulated in Alzheimer’s disease, activates cell cycle entry, tau-phosphorylation, and apoptosis in postmitotic neurons. J Neurosci. 2013;33(37):14645–14659. doi:10.1523/JNEUROSCI.1327-13.2013
72. Ciapa B, Granon S. Expression of cyclin-D1 in astrocytes varies during aging. Front Aging Neurosci. 2018;10:104. doi:10.3389/fnagi.2018.00104
73. Heo SY, Jeong M-S, Lee HS, et al. Dieckol induces cell cycle arrest by down-regulating CDK 2/cyclin E in response to p21/p53 activation in human tracheal fibroblasts. Cell Biochem Funct. 2022;40(1):71–78. doi:10.1002/cbf.3675
74. Chen X, Low K-H, Alexander A, et al. Cyclin E overexpression sensitizes triple-negative breast cancer to Wee1 kinase inhibition. Clin Cancer Res. 2018;24(24):6594–6610. doi:10.1158/1078-0432.CCR-18-1446
75. Min A, Kim JE, Kim Y-J, et al. Cyclin E overexpression confers resistance to the CDK4/6 specific inhibitor palbociclib in gastric cancer cells. Cancer Lett. 2018;430:123–132. doi:10.1016/j.canlet.2018.04.037
76. Albero R, Enjuanes A, Demajo S, et al. Cyclin D1 overexpression induces global transcriptional downregulation in lymphoid neoplasms. J Clin Invest. 2018;128(9):4132–4147. doi:10.1172/JCI96520
77. Tao JL, Luo M, Sun H, et al. Overexpression of tripartite motif containing 26 inhibits non‐small cell lung cancer cell growth by suppressing PI3K/AKT signaling. Kaohsiung J Med Sci. 2020;36(6):417–422. doi:10.1002/kjm2.12194
78. Jian Z, Zhang L, Jin L, et al. Rab5 regulates the proliferation, migration and invasion of glioma cells via cyclin E. Oncol Lett. 2020;20(2):1055–1062. doi:10.3892/ol.2020.11660
79. Lee C, Fernandez KJ, Alexandrou S, et al. Cyclin E2 promotes whole genome doubling in breast cancer. Cancers. 2020;12(8):2268. doi:10.3390/cancers12082268
80. Dong L, Yu L, Bai C, et al. USP27-mediated Cyclin E stabilization drives cell cycle progression and hepatocellular tumorigenesis. Oncogene. 2018;37(20):2702–2713. doi:10.1038/s41388-018-0137-z
81. Li S, Zhang H, Wei X. Roles and Mechanisms of Deubiquitinases (DUBs) in Breast Cancer Progression and Targeted Drug Discovery. Life. 2021;11(9):965. doi:10.3390/life11090965
82. Glasgow SD, Ruthazer ES, Kennedy TE. Guiding synaptic plasticity: Novel roles for netrin‐1 in synaptic plasticity and memory formation in the adult brain. J Physiol. 2021;599(2):493–505. doi:10.1113/JP278704
83. Nangia V, O’Connell J, Chopra K, et al. Genetic reduction of tyramine β hydroxylase suppresses Tau toxicity in a Drosophila model of tauopathy. Neurosci Lett. 2021;755:135937. doi:10.1016/j.neulet.2021.135937
84. John A, Reddy PH. Synaptic basis of Alzheimer’s disease: Focus on synaptic amyloid beta, P-tau and mitochondria. Ageing Res Rev. 2021;65:101208. doi:10.1016/j.arr.2020.101208
85. Li Y, Guo H, Wang Z, et al. Cyclin F and KIF20A, FOXM1 target genes, increase proliferation and invasion of ovarian cancer cells. Exp Cell Res. 2020;395(2):112212. doi:10.1016/j.yexcr.2020.112212
86. Walter D, Hoffmann S, Komseli E-S, et al. SCFCyclin F-dependent degradation of CDC6 suppresses DNA re-replication. Nat Commun. 2016;7(1):1–10. doi:10.1038/ncomms10530
87. Galper J, Rayner SL, Hogan AL, et al. Cyclin F: a component of an E3 ubiquitin ligase complex with roles in neurodegeneration and cancer. Int J Biochem Cell Biol. 2017;89:216–220. doi:10.1016/j.biocel.2017.06.011
88. Deshmukh RS, Sharma S, Das S. Cyclin F-dependent degradation of RBPJ inhibits IDH1R132H-mediated tumorigenesis. Cancer Res. 2018;78(22):6386–6398. doi:10.1158/0008-5472.CAN-18-1772
89. Han CH, Batchelor TT. Isocitrate dehydrogenase mutation as a therapeutic target in gliomas. Chin Clin Oncol. 2017;6(3):33. doi:10.21037/cco.2017.06.11
90. Mackenzie IR, Nicholson AM, Sarkar M, et al. TIA1 mutations in amyotrophic lateral sclerosis and frontotemporal dementia promote phase separation and alter stress granule dynamics. Neuron. 2017;95(4):808–816. e9. doi:10.1016/j.neuron.2017.07.025
91. Sørensen MF, Heimisdóttir SB, Sørensen MD, et al. High expression of cystine–glutamate antiporter xCT (SLC7A11) is an independent biomarker for epileptic seizures at diagnosis in glioma. J Neurooncol. 2018;138(1):49–53. doi:10.1007/s11060-018-2785-9
92. Robert SM, Sontheimer H. Glutamate transporters in the biology of malignant gliomas. Cell Mol Life Sci. 2014;71(10):1839–1854. doi:10.1007/s00018-013-1521-z
93. Savaskan N E, Fan Z, Broggini T, et al. Neurodegeneration in the brain tumor microenvironment: glutamate in the limelight. Curr Neuropharmacol. 2015;13(2):258–265. doi:10.2174/1570159X13666150122224158
94. Belov Kirdajova D, Kriska J, Tureckova J, Anderova M. Ischemia-triggered glutamate excitotoxicity from the perspective of glial cells. Front Cell Neurosci. 2020;14:51. doi:10.3389/fncel.2020.00051
95. Lange F, Hörnschemeyer J, Kirschstein T. Glutamatergic Mechanisms in Glioblastoma and Tumor-Associated Epilepsy. Cells. 2021;10(5):1226. doi:10.3390/cells10051226
96. Tawfik A, Mohamed R, Kira D, et al. N-Methyl-D-aspartate receptor activation, novel mechanism of homocysteine-induced blood–retinal barrier dysfunction. J Mol Med. 2021;99(1):119–130. doi:10.1007/s00109-020-02000-y
97. Shafiei-Irannejad V, Abbaszadeh S, Janssen PML, et al. Memantine and its benefits for cancer, cardiovascular and neurological disorders. Eur J Pharmacol. 2021;910:174455. doi:10.1016/j.ejphar.2021.174455
98. Estrada LD, Oliveira-Cruz L, Cabrera D. Transforming Growth Factor Beta Type I Role in Neurodegeneration: Implications for Alzheimer´s Disease. Curr Protein Pept Sci. 2018;19(12):1180–1188. doi:10.2174/1389203719666171129094937
99. Lee M, Lin S-R, Chang J-Y, et al. TGF-β induces TIAF1 self-aggregation via type II receptor-independent signaling that leads to generation of amyloid β plaques in Alzheimer’s disease. Cell Death Dis. 2010;1(12):e110–e110. doi:10.1038/cddis.2010.83
100. Chang H-T, Liu -C-C, Chen S-T, et al. WW domain-containing oxidoreductase in neuronal injury and neurological diseases. Oncotarget. 2014;5(23):11792. doi:10.18632/oncotarget.2961
101. Chou P-Y, Lin S-R, Lee M-H, et al. A p53/TIAF1/WWOX triad exerts cancer suppression but may cause brain protein aggregation due to p53/WWOX functional antagonism. Cell Commun Signal. 2019;17(1):1–16. doi:10.1186/s12964-019-0382-y
102. Lee M-H, Shih YH, Yap YV, et al. Zfra restores memory deficits in Alzheimer’s disease triple-transgenic mice by blocking aggregation of TRAPPC6AΔ, SH3GLB2, tau, and amyloid β, and inflammatory NF-κB activation. Alzheimer's Dement. 2017;3(2):189–204.
103. Chang NS. Zfra regulates protein degradation and provides strong prevention against skin cancer. Cancer Research. 2011;71(8_Supplement):4621.
104. Guo T, Zhang D, Zeng Y, et al. Molecular and cellular mechanisms underlying the pathogenesis of Alzheimer’s disease. Mol Neurodegener. 2020;15(1):1–37. doi:10.1186/s13024-019-0350-4
105. Ruffini N, Klingenberg S, Schweiger S, et al. Common factors in neurodegeneration: a meta-study revealing shared patterns on a multi-omics scale. Cells. 2020;9(12):2642. doi:10.3390/cells9122642
106. Zhou Z, Ni K, Deng H, et al. Dancing with reactive oxygen species generation and elimination in nanotheranostics for disease treatment. Adv Drug Deliv Rev. 2020;158:73–90. doi:10.1016/j.addr.2020.06.006
107. Hook V, Yoon M, Mosier C, et al. Cathepsin B in neurodegeneration of Alzheimer’s disease, traumatic brain injury, and related brain disorders. Biochim Biophys Acta Proteins Proteom. 2020;1868(8):140428. doi:10.1016/j.bbapap.2020.140428
108. Hickman S, Izzy S, Sen P, Morsett L, El Khoury J. Microglia in neurodegeneration. Nat Neurosci. 2018;21(10):1359–1369. doi:10.1038/s41593-018-0242-x
109. Ernest James Phillips T, Maguire E. Phosphoinositides: roles in the development of microglial-mediated neuroinflammation and neurodegeneration. Front Cell Neurosci. 2021;15:90.
110. Desale SE, Chinnathambi S. Phosphoinositides signaling modulates microglial actin remodeling and phagocytosis in Alzheimer’s disease. Cell Commun Signal. 2021;19(1):1–12. doi:10.1186/s12964-021-00715-0
111. Thomsen MS, Routhe LJ, Moos T. The vascular basement membrane in the healthy and pathological brain. J Cereb Blood Flow Metab. 2017;37(10):3300–3317. doi:10.1177/0271678X17722436
112. Gaspar R, Meisl G, Buell AK, et al. Secondary nucleation of monomers on fibril surface dominates α -synuclein aggregation and provides autocatalytic amyloid amplification. Q Rev Biophys. 2017;50. doi:10.1017/S0033583516000172
113. Michaels TCT, Šarić A, Curk S, et al. Dynamics of oligomer populations formed during the aggregation of Alzheimer’s Aβ42 peptide. Nat Chem. 2020;12(5):445–451. doi:10.1038/s41557-020-0452-1
114. Malishev R, Nandi S, Śmiłowicz D, et al. Interactions between BIM protein and beta-amyloid may reveal a crucial missing link between Alzheimer’s disease and neuronal cell death. ACS Chem Neurosci. 2019;10(8):3555–3564. doi:10.1021/acschemneuro.9b00177
115. Tang Y, Gao J, Wang T, et al. The effect of drug loading and multiple administration on the protein corona formation and brain delivery property of PEG-PLA nanoparticles. Acta Pharm Sin B. 2021;2021:1.
116. Huang Q, Zhao Q, Peng J, et al. Peptide–polyphenol (KLVFF/EGCG) binary modulators for inhibiting aggregation and neurotoxicity of amyloid-β peptide. ACS Omega. 2019;4(2):4233–4242. doi:10.1021/acsomega.8b02797
117. Liu H, Yu L, Dong X, et al. Synergistic effects of negatively charged hydrophobic nanoparticles and (−)-epigallocatechin-3-gallate on inhibiting amyloid β-protein aggregation. J Colloid Interface Sci. 2017;491:305–312. doi:10.1016/j.jcis.2016.12.038
118. Zhang J, Zhou X, Yu Q, et al. Epigallocatechin-3-gallate (EGCG)-stabilized selenium nanoparticles coated with Tet-1 peptide to reduce amyloid-β aggregation and cytotoxicity. ACS Appl Mater Interfaces. 2014;6(11):8475–8487. doi:10.1021/am501341u
119. Mourtas S, Lazar AN, Markoutsa E, et al. Multifunctional nanoliposomes with curcumin–lipid derivative and brain targeting functionality with potential applications for Alzheimer disease. Eur J Med Chem. 2014;80:175–183. doi:10.1016/j.ejmech.2014.04.050
120. Papakyriakopoulou P, Manta K, Kostantini C, et al. Nasal powders of quercetin-β-cyclodextrin derivatives complexes with mannitol/lecithin microparticles for Nose-to-Brain delivery: In vitro and ex vivo evaluation. Int J Pharm. 2021;607:121016. doi:10.1016/j.ijpharm.2021.121016
121. Tong-un T, Muchimapura S, Wattanathorn J, Phachonpai W. Nasal Administration of Quercetin Liposomes Improves Memory Impairment and Neurodegeneration in Animal Model of Alzheimer’s Disease. Am J Agric Biol Sci. 2010;5(3):286–293. doi:10.3844/ajabssp.2010.286.293
122. Kuo Y-C, Chen I-Y, Rajesh R. Use of functionalized liposomes loaded with antioxidants to permeate the blood–brain barrier and inhibit β-amyloid-induced neurodegeneration in the brain. J Taiwan Inst Chem Eng. 2018;87:1–14. doi:10.1016/j.jtice.2018.03.001
123. Sawda C, Moussa C, Turner RS. Resveratrol for Alzheimer’s disease. Ann N Y Acad Sci. 2017;1403(1):142–149. doi:10.1111/nyas.13431
124. Santos AC, Pereira I, Pereira-Silva M, et al. Nanotechnology-based formulations for resveratrol delivery: Effects on resveratrol in vivo bioavailability and bioactivity. Colloids Surf B Biointerfaces. 2019;180:127–140. doi:10.1016/j.colsurfb.2019.04.030
125. Sharma HS, Muresanu DF, Castellani RJ, et al. Nanowired delivery of cerebrolysin with neprilysin and p-Tau antibodies induces superior neuroprotection in Alzheimer’s disease. Prog Brain Res. 2019;245:145–200. doi:10.1016/bs.pbr.2019.03.009
126. Li Q, Wu Y, Chen J, et al. Microglia and immunotherapy in Alzheimer’s disease. Acta Neurol Scand. 2022;145(3):273–278. doi:10.1111/ane.13551
127. Eloy JO, Petrilli R, Trevizan LNF, et al. Immunoliposomes: a review on functionalization strategies and targets for drug delivery. Colloids Surf B Biointerfaces. 2017;159:454–467. doi:10.1016/j.colsurfb.2017.07.085
128. Bachurin SO, Bovina EV, Ustyugov AA. Drugs in clinical trials for Alzheimer’s disease: the major trends. Med Res Rev. 2017;37(5):1186–1225. doi:10.1002/med.21434
129. Liu Y, Xu L-P, Dai W, et al. Graphene quantum dots for the inhibition of β amyloid aggregation. Nanoscale. 2015;7(45):19060–19065. doi:10.1039/C5NR06282A
130. Zhang L, Liu F, Sun X, et al. Engineering carbon nanotube fiber for real-time quantification of ascorbic acid levels in a live rat model of Alzheimer’s disease. Anal Chem. 2017;89(3):1831–1837. doi:10.1021/acs.analchem.6b04168
131. Gregory WE, Sharma B, Hu L, et al. Interfacial charge transfer with exfoliated graphene inhibits fibril formation in lysozyme amyloid. Biointerphases. 2020;15(3):031010. doi:10.1116/6.0000019
132. Zhang W, Sigdel G, Mintz KJ, et al. Carbon dots: A future Blood–Brain Barrier penetrating nanomedicine and drug nanocarrier. Int J Nanomedicine. 2021;16:5003. doi:10.2147/IJN.S318732
133. Zhou Y, Liyanage PY, Devadoss D, et al. Nontoxic amphiphilic carbon dots as promising drug nanocarriers across the blood–brain barrier and inhibitors of β-amyloid. Nanoscale. 2019;11(46):22387–22397. doi:10.1039/C9NR08194A
134. Liu Y, Xu L-P, Wang Q, et al. Synergistic inhibitory effect of GQDs–tramiprosate covalent binding on amyloid aggregation. ACS Chem Neurosci. 2018;9(4):817–823. doi:10.1021/acschemneuro.7b00439
135. Gong X, Zhang Q, Gao Y, et al. Phosphorus and nitrogen dual-doped hollow carbon dot as a nanocarrier for doxorubicin delivery and biological imaging. ACS Appl Mater Interfaces. 2016;8(18):11288–11297. doi:10.1021/acsami.6b01577
136. Chung YJ, Kim K, Lee BI, et al. Carbon Nanodot‐Sensitized Modulation of Alzheimer’s β‐Amyloid Self‐Assembly, Disassembly, and Toxicity. Small. 2017;13(34):1700983. doi:10.1002/smll.201700983
137. Stelmashook E, Isaev NK, Genrikhs EE, et al. Role of zinc and copper ions in the pathogenetic mechanisms of Alzheimer’s and Parkinson’s diseases. Biochemistry. 2014;79(5):391–396. doi:10.1134/S0006297914050022
138. James SA, Churches QI, de Jonge MD, et al. Iron, copper, and zinc concentration in Aβ plaques in the APP/PS1 mouse model of Alzheimer’s disease correlates with metal levels in the surrounding neuropil. ACS Chem Neurosci. 2017;8(3):629–637. doi:10.1021/acschemneuro.6b00362
139. Pithadia AS, Lim MH. Metal-associated amyloid-β species in Alzheimer’s disease. Curr Opin Chem Biol. 2012;16(1–2):67–73. doi:10.1016/j.cbpa.2012.01.016
140. Vilella A, Belletti D, Sauer AK, et al. Reduced plaque size and inflammation in the APP23 mouse model for Alzheimer’s disease after chronic application of polymeric nanoparticles for CNS targeted zinc delivery. J Trace Elem Med Biol. 2018;49:210–221. doi:10.1016/j.jtemb.2017.12.006
141. Sun D, Zhang W, Yu Q, et al. Chiral penicillamine-modified selenium nanoparticles enantioselectively inhibit metal-induced amyloid β aggregation for treating Alzheimer’s disease. J Colloid Interface Sci. 2017;505:1001–1010. doi:10.1016/j.jcis.2017.06.083
142. Mittapelly N, Thalla M, Pandey G, et al. Long acting ionically paired embonate based nanocrystals of donepezil for the treatment of Alzheimer’s disease: a proof of concept study. Pharm Res. 2017;34(11):2322–2335. doi:10.1007/s11095-017-2240-1
143. Krishna KV, Wadhwa G, Alexander A, et al. Design and biological evaluation of lipoprotein-based donepezil nanocarrier for enhanced brain uptake through oral delivery. ACS Chem Neurosci. 2019;10(9):4124–4135. doi:10.1021/acschemneuro.9b00343
144. Pagar KP, Sardar SM, Vavia PR. Novel L-Lactide-depsipeptide polymeric carrier for enhanced brain uptake of rivastigmine in treatment of Alzheimer’s disease. J Biomed Nanotechnol. 2014;10(3):415–426. doi:10.1166/jbn.2014.1719
145. Nguyen TT, Dung Nguyen TT, Vo TK, et al. Nanotechnology-based drug delivery for central nervous system disorders. Biomed Pharmacother. 2021;143:112117. doi:10.1016/j.biopha.2021.112117
146. MacLeod R, Hillert E-K, Cameron RT, et al. The role and therapeutic targeting of α-, β-and γ-secretase in Alzheimer’s disease. Future Sci OA. 2015;1(3). doi:10.4155/fso.15.9
147. Imamura Y, Umezawa N, Osawa S, et al. Effect of helical conformation and side chain structure on γ-secretase inhibition by β-peptide foldamers: insight into substrate recognition. J Med Chem. 2013;56(4):1443–1454. doi:10.1021/jm301306c
148. Rassu G, Soddu E, Posadino AM, et al. Nose-to-brain delivery of BACE1 siRNA loaded in solid lipid nanoparticles for Alzheimer’s therapy. Colloids Surf B Biointerfaces. 2017;152:296–301. doi:10.1016/j.colsurfb.2017.01.031
149. Zheng M, Tao W, Zou Y, et al. Nanotechnology-based strategies for siRNA brain delivery for disease therapy. Trends Biotechnol. 2018;36(5):562–575. doi:10.1016/j.tibtech.2018.01.006
150. Wang P, Zheng X, Guo Q, et al. Systemic delivery of BACE1 siRNA through neuron-targeted nanocomplexes for treatment of Alzheimer’s disease. J Control Release. 2018;279:220–233. doi:10.1016/j.jconrel.2018.04.034
151. Zhou Y, Zhu F, Liu Y, et al. Blood-brain barrier–penetrating siRNA nanomedicine for Alzheimer’s disease therapy. Sci Adv. 2020;6(41):eabc7031. doi:10.1126/sciadv.abc7031
152. Pegtel DM, Gould SJ. Exosomes. Annu Rev Biochem. 2019;88(1):487–514. doi:10.1146/annurev-biochem-013118-111902
153. Snyder A, Grunseich C. Hitching a ride on exosomes: a new approach for the delivery of siRNA-mediated therapies. Brain. 2021;144(11):3286–3287. doi:10.1093/brain/awab398
154. Urban AS, Pavlov KV, Kamynina AV, et al. Structural Studies Providing Insights into Production and Conformational Behavior of Amyloid-β Peptide Associated with Alzheimer’s Disease Development. Molecules. 2021;26(10):2897. doi:10.3390/molecules26102897
155. Song C, Shi J, Zhang P, et al. Immunotherapy for Alzheimer’s disease: targeting β-amyloid and beyond. Transl Neurodegener. 2022;11(1):1–17. doi:10.1186/s40035-022-00292-3
156. Farizatto KLG, Ikonne US, Almeida MF, et al. Aβ42-mediated proteasome inhibition and associated tau pathology in hippocampus are governed by a lysosomal response involving cathepsin B: Evidence for protective crosstalk between protein clearance pathways. PLoS One. 2017;12(8):e0182895. doi:10.1371/journal.pone.0182895
157. Xiao S, Song -L-L, Li J-T, et al. Intraperitoneal Administration of Monoclonal Antibody Against Pathologic Aβ 42 Aggregates Alleviated Cognitive Deficits and Synaptic Lesions in APP/PS1 Mice. J Alzheimer's Dis. 2020;73(2):657–670. doi:10.3233/JAD-190874
158. Usman M, Bhardwaj S, Roychoudhury S, et al. Immunotherapy for Alzheimer’s disease: current scenario and future perspectives. J Prev Alzheimer's Dis. 2021;8(4):534–551. doi:10.14283/jpad.2021.52
159. Yamazaki Y, Painter MM, Bu G, et al. Apolipoprotein E as a therapeutic target in Alzheimer’s disease: a review of basic research and clinical evidence. CNS drugs. 2016;30(9):773–789. doi:10.1007/s40263-016-0361-4
160. Huang Y-WA, Zhou B, Nabet AM, et al. Differential Signaling Mediated by ApoE2, ApoE3, and ApoE4 in Human Neurons Parallels Alzheimer’s Disease Risk. The Journal of Neuroscience. 2019;39(37):7408–7427. doi:10.1523/JNEUROSCI.2994-18.2019
161. Davis AA, Inman CE, Wargel ZM, et al. APOE genotype regulates pathology and disease progression in synucleinopathy. Sci Transl Med. 2020;12(529):eaay3069. doi:10.1126/scitranslmed.aay3069
162. Zhao N, Urban AS, Pavlov KV, et al. APOE4 exacerbates α-synuclein pathology and related toxicity independent of amyloid. Sci Transl Med. 2020;12(529):eaay1809. doi:10.1126/scitranslmed.aay1809
163. Chai AB, Lam HH, Kockx M, Gelissen IC. Apolipoprotein E isoform-dependent effects on the processing of Alzheimer’s amyloid-β. Biochim Biophys Acta Mol Cell Biol Lipids. 2021;1866(9):158980.
164. Frieden C, Wang H, Ho CM. A mechanism for lipid binding to apoE and the role of intrinsically disordered regions coupled to domain–domain interactions. Proc Natl Acad Sci. 2017;114(24):6292–6297. doi:10.1073/pnas.1705080114
165. Chen Y, Strickland MR, Soranno A, et al. Apolipoprotein E: structural insights and links to Alzheimer disease pathogenesis. Neuron. 2021;109(2):205–221. doi:10.1016/j.neuron.2020.10.008
166. Wang C, Najm R, Xu Q, et al. Gain of toxic apolipoprotein E4 effects in human iPSC-derived neurons is ameliorated by a small-molecule structure corrector. Nat Med. 2018;24(5):647–657. doi:10.1038/s41591-018-0004-z
167. Montazersaheb S, Ahmadian E, Dizaj SM, et al. Emerging Nanotherapeutic Alzheimer’s Disease. Front Clin Drug Res. 2021;2:173.
168. Martin-Rapun R, De Matteis L, Ambrosone A, et al. Targeted Nanoparticles for the Treatment of Alzheimer’s Disease. Curr Pharm Des. 2017;23(13):1927–1952. doi:10.2174/1381612822666161226151011
169. Kong N, Deng M, Sun X-N, et al. Polydopamine-functionalized CA-(PCL-ran-PLA) nanoparticles for target delivery of docetaxel and chemo-photothermal therapy of breast cancer. Front Pharmacol. 2018;9:125. doi:10.3389/fphar.2018.00125
170. Su S, Kang PM. Systemic review of biodegradable nanomaterials in nanomedicine. Nanomaterials. 2020;10(4):656. doi:10.3390/nano10040656
171. Griffin S, Masood M, Nasim M, et al. Natural nanoparticles: a particular matter inspired by nature. Antioxidants. 2017;7(1):3. doi:10.3390/antiox7010003
172. Ibarra LE, Beaugé L, Arias-Ramos N, et al. Trojan horse monocyte-mediated delivery of conjugated polymer nanoparticles for improved photodynamic therapy of glioblastoma. Nanomedicine. 2020;15(17):1687–1707. doi:10.2217/nnm-2020-0106
173. Casamonti M, Risaliti L, Vanti G, et al. Andrographolide loaded in micro-and nano-formulations: improved bioavailability, target-tissue distribution, and efficacy of the “king of bitters”. Engineering. 2019;5(1):69–75. doi:10.1016/j.eng.2018.12.004
174. Zhang C, Song J, Lou L, et al. Doxorubicin‐loaded nanoparticle coated with endothelial cells‐derived exosomes for immunogenic chemotherapy of glioblastoma. Bioeng Transl Med. 2021;6(3):e10203. doi:10.1002/btm2.10203
175. Kaushal N. Double-Coated Biodegradable Poly (Butyl Cyanoacrylate) Nanoparticulate Delivery Systems for Brain Targeting of Doxorubicin via Oral Administration. New York: St. John’s University; 2021.
176. Wiwatchaitawee K, Quarterman JC, Geary SM, et al. Enhancement of therapies for glioblastoma (GBM) using nanoparticle-based delivery systems. AAPS PharmSciTech. 2021;22(2):1–16. doi:10.1208/s12249-021-01928-9
177. Pereverzeva E, Treschalin I, Bodyagin D, et al. Influence of the formulation on the tolerance profile of nanoparticle-bound doxorubicin in healthy rats: focus on cardio-and testicular toxicity. Int J Pharm. 2007;337(1–2):346–356. doi:10.1016/j.ijpharm.2007.01.031
178. Muhamad N, Plengsuriyakarn T, Na-Bangchang K. Application of active targeting nanoparticle delivery system for chemotherapeutic drugs and traditional/herbal medicines in cancer therapy: a systematic review. Int J Nanomedicine. 2018;13:3921. doi:10.2147/IJN.S165210
179. Marques A, Costa PJ, Velho S, et al. Functionalizing nanoparticles with cancer-targeting antibodies: A comparison of strategies. J Control Release. 2020;320:180–200. doi:10.1016/j.jconrel.2020.01.035
180. Kaźmierczak Z, Szostak-Paluch K, Przybyło M, et al. Endocytosis in cellular uptake of drug delivery vectors: Molecular aspects in drug development. Bioorg Med Chem. 2020;28(18):115556. doi:10.1016/j.bmc.2020.115556
181. Vora P, Venugopal C, Salim SK, et al. The rational development of CD133-targeting immunotherapies for glioblastoma. Cell Stem Cell. 2020;26(6):832–844. e6. doi:10.1016/j.stem.2020.04.008
182. Zhu X, Prasad S, Gaedicke S, et al. Patient-derived glioblastoma stem cells are killed by CD133-specific CAR T cells but induce the T cell aging marker CD57. Oncotarget. 2015;6(1):171. doi:10.18632/oncotarget.2767
183. Jhaveri A, Deshpande P, Pattni B, et al. Transferrin-targeted, resveratrol-loaded liposomes for the treatment of glioblastoma. J Control Release. 2018;277:89–101. doi:10.1016/j.jconrel.2018.03.006
184. Devarajan PV, Dandekar P, D’Souza AA. Targeted Intracellular Drug Delivery by Receptor Mediated Endocytosis. Springer; 2019.
185. Zheng M, Ruan S, Liu S, et al. Self-targeting fluorescent carbon dots for diagnosis of brain cancer cells. ACS nano. 2015;9(11):11455–11461. doi:10.1021/acsnano.5b05575
186. Qiao L, Sun T, Zheng X, et al. Exploring the optimal ratio of d-glucose/l-aspartic acid for targeting carbon dots toward brain tumor cells. Mater Sci Eng C. 2018;85:1–6. doi:10.1016/j.msec.2017.12.011
187. Qian ZM, Li H, Sun H, Ho K. Targeted drug delivery via the transferrin receptor-mediated endocytosis pathway. Pharmacol Rev. 2002;54(4):561–587. doi:10.1124/pr.54.4.561
188. Hettiarachchi SD, Graham RM, Mintz KJ, et al. Triple conjugated carbon dots as a nano-drug delivery model for glioblastoma brain tumors. Nanoscale. 2019;11(13):6192–6205. doi:10.1039/C8NR08970A
189. Liyanage PY, Zhou Y, Al-Youbi AO, et al. Pediatric glioblastoma target-specific efficient delivery of gemcitabine across the blood–brain barrier via carbon nitride dots. Nanoscale. 2020;12(14):7927–7938. doi:10.1039/D0NR01647K
190. Sun T, Patil R, Galstyan A, et al. Blockade of a laminin-411–notch axis with CRISPR/Cas9 or a nanobioconjugate inhibits glioblastoma growth through tumor-microenvironment cross-talk. Cancer Res. 2019;79(6):1239–1251. doi:10.1158/0008-5472.CAN-18-2725
191. Ljubimova JY. In vivo targeting of laminin-411-β1 integrin-Notch signaling pathway using nanobioconjugate alters glioma microenvironment for effective treatment. AACR; 2017.
192. Canfarotta F, Lezina L, Guerreiro A, et al. Specific drug delivery to cancer cells with double-imprinted nanoparticles against epidermal growth factor receptor. Nano Lett. 2018;18(8):4641–4646. doi:10.1021/acs.nanolett.7b03206
193. Zhang Q, Liu Q, Du M, et al. Cetuximab and Doxorubicin loaded dextran-coated Fe3O4 magnetic nanoparticles as novel targeted nanocarriers for non-small cell lung cancer. J Magn Magn Mater. 2019;481:122–128. doi:10.1016/j.jmmm.2019.01.021
194. Katsushima K, Natsume A, Ohka F, et al. Targeting the Notch-regulated non-coding RNA TUG1 for glioma treatment. Nat Commun. 2016;7(1):1–14. doi:10.1038/ncomms13616
195. Hsu SPC, Dhawan U, Tseng -Y-Y, et al. Glioma-sensitive delivery of Angiopep-2 conjugated iron gold alloy nanoparticles ensuring simultaneous tumor imaging and hyperthermia mediated cancer theranostics. Appl Mater Today. 2020;18:100510. doi:10.1016/j.apmt.2019.100510
196. Wang M, Kuang R, Huang B, et al. Polylactic acid block copolymer grafted temozolomide targeted nano delivery in the treatment of glioma. Mater Express. 2021;11(5):627–633. doi:10.1166/mex.2021.1961
197. Xuan S, Shin DH, Kim J-S. Angiopep-2-conjugated liposomes encapsulating γ-secretase inhibitor for targeting glioblastoma stem cells. J Pharm Investig. 2014;44(7):473–483. doi:10.1007/s40005-014-0151-2
198. Xu H, Li C, Wei Y, et al. Angiopep-2-modified calcium arsenite-loaded liposomes for targeted and pH-responsive delivery for anti-glioma therapy. Biochem Biophys Res Commun. 2021;551:14–20. doi:10.1016/j.bbrc.2021.02.138
199. Liu Y, Mei L, Yu Q, et al. Multifunctional tandem peptide modified paclitaxel-loaded liposomes for the treatment of vasculogenic mimicry and cancer stem cells in malignant glioma. ACS Appl Mater Interfaces. 2015;7(30):16792–16801. doi:10.1021/acsami.5b04596
200. Wang Y, Ying X, Xu H, et al. The functional curcumin liposomes induce apoptosis in C6 glioblastoma cells and C6 glioblastoma stem cells in vitro and in animals. Int J Nanomedicine. 2017;12:1369. doi:10.2147/IJN.S124276
201. Shi M, Anantha M, Wehbe M, et al. Liposomal formulations of carboplatin injected by convection-enhanced delivery increases the median survival time of F98 glioma bearing rats. J Nanobiotechnology. 2018;16(1):1–12. doi:10.1186/s12951-018-0404-8
202. Xia P, Li Q, Wu G, et al. An immune-related lncRNA signature to predict survival in glioma patients. Cell Mol Neurobiol. 2021;41(2):365–375. doi:10.1007/s10571-020-00857-8
203. Sriraman A. Targeting MDM2, the antagonist of the tumor suppressor p53. Niedersächsische Staats-und Universitätsbibliothek Göttingen; 2018.
204. Yang R, Cai TT, Wu XJ, et al. Tumour YAP1 and PTEN expression correlates with tumour‐associated myeloid suppressor cell expansion and reduced survival in colorectal cancer. Immunology. 2018;155(2):263–72.
205. Kreatsoulas D, Bolyard C, Wu BX, et al. Translational landscape of glioblastoma immunotherapy for physicians: guiding clinical practice with basic scientific evidence. J Hematol Oncol. 2022;15(1):1–30.
206. Loya J, Zhang C, Cox E, et al. Biological intratumoral therapy for the high-grade glioma part I: intratumoral delivery and immunotoxins. CNS Oncol. 2019;8(3):CNS38. doi:10.2217/cns-2019-0001
207. Mohammed S, Shamseddine AA, Newcomb B, et al. Sublethal doxorubicin promotes migration and invasion of breast cancer cells: role of Src Family non-receptor tyrosine kinases. Breast Cancer Res. 2021;23(1):1–20. doi:10.1186/s13058-021-01452-5
208. Wang T, Tang J, Yang H, et al. Effect of apatinib plus pegylated liposomal doxorubicin vs pegylated liposomal doxorubicin alone on platinum-resistant recurrent ovarian cancer: the APPROVE randomized clinical trial. JAMA Oncol. 2022;8(8):1169–1176. doi:10.1001/jamaoncol.2022.2253
209. Riley RS, June CH, Langer R, et al. Delivery technologies for cancer immunotherapy. Nat Rev Drug Discov. 2019;18(3):175–196. doi:10.1038/s41573-018-0006-z
210. Feng X, Xu W, Liu J, et al. Polypeptide nanoformulation-induced immunogenic cell death and remission of immunosuppression for enhanced chemoimmunotherapy. Sci Bull. 2021;66(4):362–373. doi:10.1016/j.scib.2020.07.013
211. Ding Y, Xu Y, Yang W, et al. Investigating the EPR effect of nanomedicines in human renal tumors via ex vivo perfusion strategy. Nano Today. 2020;35:100970. doi:10.1016/j.nantod.2020.100970
212. Singh PK, Srivastava AK, Dev A, et al. 1, 3β-Glucan anchored, paclitaxel loaded chitosan nanocarrier endows enhanced hemocompatibility with efficient anti-glioblastoma stem cells therapy. Carbohydr Polym. 2018;180:365–375. doi:10.1016/j.carbpol.2017.10.030
213. Dorababu A. Recent Advances in Nanoformulated Chemotherapeutic Drug Delivery (2015‐2019). ChemistrySelect. 2019;4(29):8731–8744. doi:10.1002/slct.201901064
214. Joshi HA, Patwardhan RS, Sharma D, et al. Pre-clinical evaluation of an innovative oral nano-formulation of baicalein for modulation of radiation responses. Int J Pharm. 2021;595:120181. doi:10.1016/j.ijpharm.2020.120181
215. Frankel BM, Cachia D, Patel SJ, et al. Targeting subventricular zone progenitor cells with intraventricular liposomal encapsulated cytarabine in patients with secondary glioblastoma: A report of two cases. SN Compr Clin Med. 2020;2(6):836–843. doi:10.1007/s42399-020-00322-z
216. Ahmadzada T, Reid G, McKenzie DR. Fundamentals of siRNA and miRNA therapeutics and a review of targeted nanoparticle delivery systems in breast cancer. Biophys Rev. 2018;10(1):69–86. doi:10.1007/s12551-017-0392-1
217. Rahman M, Alharbi KS, Alruwaili NK, et al. Nucleic acid-loaded lipid-polymer nanohybrids as novel nanotherapeutics in anticancer therapy. Expert Opin Drug Deliv. 2020;17(6):805–816. doi:10.1080/17425247.2020.1757645
218. Ferreira D, Fontinha D, Martins C, et al. Gold nanoparticles for vectorization of nucleic acids for cancer therapeutics. Molecules. 2020;25(15):3489. doi:10.3390/molecules25153489
219. Galli M, Guerrini A, Cauteruccio S, et al. Superparamagnetic iron oxide nanoparticles functionalized by peptide nucleic acids. RSC Adv. 2017;7(25):15500–15512. doi:10.1039/C7RA00519A
220. Li C, Dou Y, Chen Y, et al. Site‐specific microRNA‐33 antagonism by pH‐responsive nanotherapies for treatment of atherosclerosis via regulating cholesterol efflux and adaptive immunity. Adv Funct Mater. 2020;30(42):2002131. doi:10.1002/adfm.202002131
221. Liu Y, Zhang D, An Y, et al. Non-invasive PTEN mRNA brain delivery effectively mitigates growth of orthotopic glioblastoma. Nano Today. 2023;49:101790. doi:10.1016/j.nantod.2023.101790
222. Wang Y, Sun Y, Geng N, et al. A Biomimetic Nanomedicine Targets Orthotopic Glioblastoma by Combinatorial Co‐Delivery of Temozolomide and a Methylguanine‐DNA Methyltransferase Inhibitor. Adv Ther. 2022;5(12):2200095. doi:10.1002/adtp.202200095
223. Kubiatowicz LJ, Mohapatra A, Krishnan N, Fang RH, Zhang L. mRNA nanomedicine: Design and recent applications. In: Exploration. Wiley Online Library; 2022.
224. Shah U, Joshi G, Sawant K. Improvement in antihypertensive and antianginal effects of felodipine by enhanced absorption from PLGA nanoparticles optimized by factorial design. Mater Sci Eng C. 2014;35:153–163. doi:10.1016/j.msec.2013.10.038
225. Binda A, Murano C, Rivolta I. Innovative therapies and nanomedicine applications for the treatment of Alzheimer’s disease: a state-of-the-art (2017–2020). Int J Nanomedicine. 2020;15:6113. doi:10.2147/IJN.S231480
226. Tóth OM, Menyhárt Á, Varga VÉ, et al. Chitosan nanoparticles release nimodipine in response to tissue acidosis to attenuate spreading depolarization evoked during forebrain ischemia. Neuropharmacology. 2020;162:107850. doi:10.1016/j.neuropharm.2019.107850
227. Remya P, Damodharan N. Formulation, development and characterisation of nimodipine loaded solid lipid nanoparticles. Int J App Pharm. 2020;2020:265–271.
228. Sha’at F, Pavaloiu RD, Salceanu DC, et al. Formulation of Polymeric Multicomponent Systems Containing Cardiovascular APIs. Mater Plast. 2018;55(1):121. doi:10.37358/MP.18.1.4976
229. Alawdi SH, Eidi H, Safar MM, et al. Loading amlodipine on diamond nanoparticles: A novel drug delivery system. Nanotechnol Sci Appl. 2020;12:47. doi:10.2147/NSA.S232517
230. Fathi-Achachelouei M, Knopf-Marques H, Ribeiro da Silva CE, et al. Use of nanoparticles in tissue engineering and regenerative medicine. Front Bioeng Biotechnol. 2019;7:113. doi:10.3389/fbioe.2019.00113
231. Yuan M, Wang Y, Qin Y-X. Engineered nanomedicine for neuroregeneration: light emitting diode-mediated superparamagnetic iron oxide-gold core-shell nanoparticles functionalized by nerve growth factor. Nanomedicine. 2019;21:102052. doi:10.1016/j.nano.2019.102052
232. Xiao Y, Zhang E, Fu A. Promotion of SH-SY5Y cell growth by gold nanoparticles modified with 6-mercaptopurine and a neuron-penetrating peptide. Nanoscale Res Lett. 2017;12(1):1–9. doi:10.1186/s11671-017-2417-x
233. Katebi S, Esmaeili A, Ghaedi K, et al. Superparamagnetic iron oxide nanoparticles combined with NGF and quercetin promote neuronal branching morphogenesis of PC12 cells. Int J Nanomedicine. 2019;14:2157. doi:10.2147/IJN.S191878
234. Wójtowicz S, Strosznajder AK, Jeżyna M, et al. The novel role of PPAR alpha in the brain: promising target in therapy of Alzheimer’s disease and other neurodegenerative disorders. Neurochem Res. 2020;45(5):972–988. doi:10.1007/s11064-020-02993-5
235. Strosznajder AK, Wójtowicz S, Jeżyna MJ, et al. Recent insights on the role of PPAR-β/δ in neuroinflammation and neurodegeneration, and its potential target for therapy. Neuromolecular Med. 2020:1–13. doi:10.1007/s12017-019-08556-4
236. Wagner N, Wagner K-D. PPAR beta/delta and the hallmarks of cancer. Cells. 2020;9(5):1133. doi:10.3390/cells9051133
237. Silva-Abreu M, Calpena AC, Andrés-Benito P, et al. PPARγ agonist-loaded PLGA-PEG nanocarriers as a potential treatment for Alzheimer’s disease: in vitro and in vivo studies. Int J Nanomedicine. 2018;13:5577. doi:10.2147/IJN.S171490
238. Carradori D, Balducci C, Re F, et al. Antibody-functionalized polymer nanoparticle leading to memory recovery in Alzheimer’s disease-like transgenic mouse model. Nanomedicine. 2018;14(2):609–618. doi:10.1016/j.nano.2017.12.006
239. Sun J, Xie W, Zhu X, et al. Sulfur nanoparticles with novel morphologies coupled with brain-targeting peptides RVG as a new type of inhibitor against metal-induced aβ aggregation. ACS Chem Neurosci. 2017;9(4):749–761. doi:10.1021/acschemneuro.7b00312
240. Ji D, Wu X, Li D, et al. Protective effects of chondroitin sulphate nano-selenium on a mouse model of Alzheimer’s disease. Int J Biol Macromol. 2020;154:233–245. doi:10.1016/j.ijbiomac.2020.03.079
241. Sun T, Li Y, Huang Y, et al. Targeting glioma stem cells enhances anti-tumor effect of boron neutron capture therapy. Oncotarget. 2016;7(28):43095. doi:10.18632/oncotarget.9355
242. Qin L, Wang C-Z, Fan H-J, et al. A dual‑targeting liposome conjugated with transferrin and arginine‑glycine‑aspartic acid peptide for glioma‑targeting therapy. Oncol Lett. 2014;8(5):2000–2006. doi:10.3892/ol.2014.2449
243. Caruso G, Caffo M, Raudino G, et al. Antisense oligonucleotides as an innovative therapeutic strategy in the treatment of high-grade gliomas. Recent Pat CNS Drug Discov. 2010;5(1):53–69. doi:10.2174/157488910789753503
244. Chou S-T, Patil R, Galstyan A, et al. Simultaneous blockade of interacting CK2 and EGFR pathways by tumor-targeting nanobioconjugates increases therapeutic efficacy against glioblastoma multiforme. J Control Release. 2016;244:14–23. doi:10.1016/j.jconrel.2016.11.001
245. Kaluzova M, Bouras A, Machaidze R, et al. Targeted therapy of glioblastoma stem-like cells and tumor non-stem cells using cetuximab-conjugated iron-oxide nanoparticles. Oncotarget. 2015;6(11):8788. doi:10.18632/oncotarget.3554
246. Wang X, Zhao Y, Dong S, et al. Cell-penetrating peptide and transferrin co-modified liposomes for targeted therapy of glioma. Molecules. 2019;24(19):3540. doi:10.3390/molecules24193540
247. Kakinen A, Javed I, Davis TP, et al. In vitro and in vivo models for anti-amyloidosis nanomedicines. Nanoscale Horiz. 2021;6(2):95–119. doi:10.1039/D0NH00548G
248. Uddin M, Kabir MT, Niaz K, et al. Molecular insight into the therapeutic promise of flavonoids against Alzheimer’s disease. Molecules. 2020;25(6):1267. doi:10.3390/molecules25061267
249. Rajput AP, Butani SB. Resveratrol anchored nanostructured lipid carrier loaded in situ gel via nasal route: Formulation, optimization and in vivo characterization. J Drug Deliv Sci Technol. 2019;51:214–223. doi:10.1016/j.jddst.2019.01.040
250. Savla SR, Laddha AP, Kulkarni YA. Pharmacology of apocynin: a natural acetophenone. Drug Metab Rev. 2021;53(4):1–21.
251. Brenza TM, Ghaisas S, Ramirez JEV, et al. Neuronal protection against oxidative insult by polyanhydride nanoparticle-based mitochondria-targeted antioxidant therapy. Nanomedicine. 2017;13(3):809–820. doi:10.1016/j.nano.2016.10.004
252. Vaz GR, Hädrich G, Bidone J, et al. Development of nasal lipid nanocarriers containing curcumin for brain targeting. J Alzheimer's Dis. 2017;59(3):961–974. doi:10.3233/JAD-160355
253. Djiokeng Paka G, Doggui S, Zaghmi A, et al. Neuronal Uptake and Neuroprotective Properties of Curcumin-Loaded Nanoparticles on SK-N-SH Cell Line: Role of Poly(lactide- co -glycolide) Polymeric Matrix Composition. Mol Pharm. 2016;13(2):391–403. doi:10.1021/acs.molpharmaceut.5b00611
254. Sokolik V, Berchenko O, Shulga S. Comparative analysis of nasal therapy with soluble and liposomal forms of curcumin on rats with Alzheimer’s disease model. J Alzheimer's Dis Parkinsonism. 2017;7(357):2161–0460.1000357.
255. Ishak RA, Mostafa NM, Kamel AO. Stealth lipid polymer hybrid nanoparticles loaded with rutin for effective brain delivery – comparative study with the gold standard (Tween 80): optimization, characterization and biodistribution. Drug Deliv. 2017;24(1):1874–1890. doi:10.1080/10717544.2017.1410263
256. Lohan S, Raza K, Mehta SK, et al. Anti-Alzheimer’s potential of berberine using surface decorated multi-walled carbon nanotubes: a preclinical evidence. Int J Pharm. 2017;530(1–2):263–278. doi:10.1016/j.ijpharm.2017.07.080
257. Su X, Zhang D, Zhang H, et al. Preparation and characterization of angiopep-2 functionalized ginsenoside-Rg3 loaded nanoparticles and the effect on C6 glioma cells. Pharm Dev Technol. 2020;25(3):385–395. doi:10.1080/10837450.2018.1551901
258. Lv L, Yang F, Li H, et al. Brain-targeted co-delivery of β-amyloid converting enzyme 1 shRNA and epigallocatechin-3-gallate by multifunctional nanocarriers for Alzheimer’s disease treatment. IUBMB life. 2020;72(8):1819–1829. doi:10.1002/iub.2330
259. Fan S, Zheng Y, Liu X, et al. Curcumin-loaded PLGA-PEG nanoparticles conjugated with B6 peptide for potential use in Alzheimer’s disease. Drug Deliv. 2018;25(1):1091–1102. doi:10.1080/10717544.2018.1461955
260. Ali T, Kim MJ, Rehman SU, et al. Anthocyanin-loaded PEG-gold nanoparticles enhanced the neuroprotection of anthocyanins in an Aβ1–42 mouse model of Alzheimer’s disease. Mol Neurobiol. 2017;54(8):6490–6506. doi:10.1007/s12035-016-0136-4
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.