")
Back to Journals » Cancer Management and Research » Volume 11
The role of visfatin in cancer proliferation, angiogenesis, metastasis, drug resistance and clinical prognosis
Authors Lin TC
Received 26 December 2018
Accepted for publication 11 March 2019
Published 23 April 2019 Volume 2019:11 Pages 3481—3491
DOI https://doi.org/10.2147/CMAR.S199597
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Lu-Zhe Sun
Tsung-Chieh Lin
Genomic Medicine Core Laboratory, Chang Gung Memorial Hospital, Linkou, Taiwan
Abstract: Visfatin, also known as nicotinamide phosphoribosyltransferase or pre-B-cell colony-enhancing factor (PBEF), is an adipocytokine secreted by adipocytes, macrophages and inflamed endothelial tissue. Related reports have indicated a positive correlation between the visfatin level and obesity and cancer risk. In addition to its original function, visfatin is multifunctional and plays critical roles in the promotion of several processes relevant to cancer progression including cancer cell proliferation, angiogenesis, metastasis and drug resistance. The relative expression of visfatin and the potential visfatin receptor on a pan-cancer scale was determined based on the transcriptome analysis data in The Cancer Genome Atlas. We further show the clinical association of its signaling axis with the survival of cancer patients, which reveals its prognostic power for specific cancer types. This review illustrates visfatin’s biological functions related to cancer progression and demonstrates its clinical significance in predicting outcomes of cancer patients.
Keywords: visfatin, cancer progression, clinical outcome
Introduction
The adipocytokine visfatin is also known as pre-B-cell colony-enhancing factor (PBEF), and is secreted by human peripheral blood lymphocytes.1 Its intracellular form has been recognized for its enzyme function in the nicotinamide adenine dinucleotide NAD+ salvage pathway where it is designated as nicotinamide phosphoribosyltransferase (NAMPT).2,3 Recently, increases in both the NAD(H) pool size and the NAD+/NADH ratio caused by NAMPT were shown to promote colorectal cancer (CRC) progression.4 Importantly, the abnormal expression of visfatin in many types of cancers and a significant correlation between high circulating visfatin levels and an increase in cancer risk were observed.5 Furthermore, emerging evidence has demonstrated visfatin expression in a broad range of cancer types along with visfatin-mediated effects on the regulation of several critical factors in processes related to cancer progression including tumor proliferation, angiogenesis, metastasis and drug resistance. In this review, we focus on the biological function of visfatin in cancer and further illustrate its clinical significance including its relative expression level on a pan-cancer scale and its specific correlation with patient survival in various cancer types.
Expression of visfatin and its potential receptor in cancer
Visfatin was detected in tumors or plasma of patients with variant types of cancers, which indicates its clinical importance in cancer progression. According to the results of a tissue array analysis in oral squamous cell carcinoma, visfatin overexpression was observed.6 In addition, higher levels of visfatin were detected in tumor tissues from 8 matched pairs of human pancreatic ductal adenocarcinoma (PDAC), and similar findings were obtained in PDAC cells.7 Breast cancer MDA-MB-231, MDA-MB-468 and MCF-7 cell lines displayed elevated visfatin levels compared with the non-transformed MCF-10A cell line, and an inverse relationship between the visfatin and p73 levels was also proposed.8 In clear cell renal cell carcinoma (RCC), visfatin expression was higher in tumors as compared to that in the adjacent normal tissues (GSE6344).9 Patients with thyroid malignancy also exhibited a higher visfatin expression level, and visfatin appeared to correlate with advanced tumor stage and metastasis.10 Moreover, an increase in visfatin expression was observed in experimental myelomatous bones compared with non-myelomatous bones.11 A comprehensive analysis using the Xena browser to analyze pan-cancer transcriptome data and matched clinical information was launched by the University of California Santa Cruz.12 The omics data were mainly generated by microarray experiments and RNA sequencing in combination with cancer patients’ follow-up data in The Cancer Genome Atlas (TCGA). The results demonstrated the relative expression levels of visfatin (NAMPT) and its potential receptor (INSR) after normalization to the pan-cancer scale (Figure 1). A relatively high visfatin level was found in lung squamous cell carcinoma, stomach adenocarcinoma, glioblastoma multiforme, liver hepatocellular carcinoma, lung adenocarcinoma, kidney clear cell carcinoma, esophageal carcinoma, head and neck squamous carcinoma, prostate carcinoma and skin cutaneous carcinoma. In addition, the potential visfatin receptor was relatively highly expressed in adrenocortical cancer, kidney chromophobe, stomach adenocarcinoma, liver hepatocellular carcinoma, kidney clear cell carcinoma, kidney papillary cell carcinoma, breast invasive carcinoma, pancreatic adenocarcinoma, pheochromocytoma and paraganglioma, prostate adenocarcinoma and thyroid carcinoma, which indicates the potential pathological role in cancer.
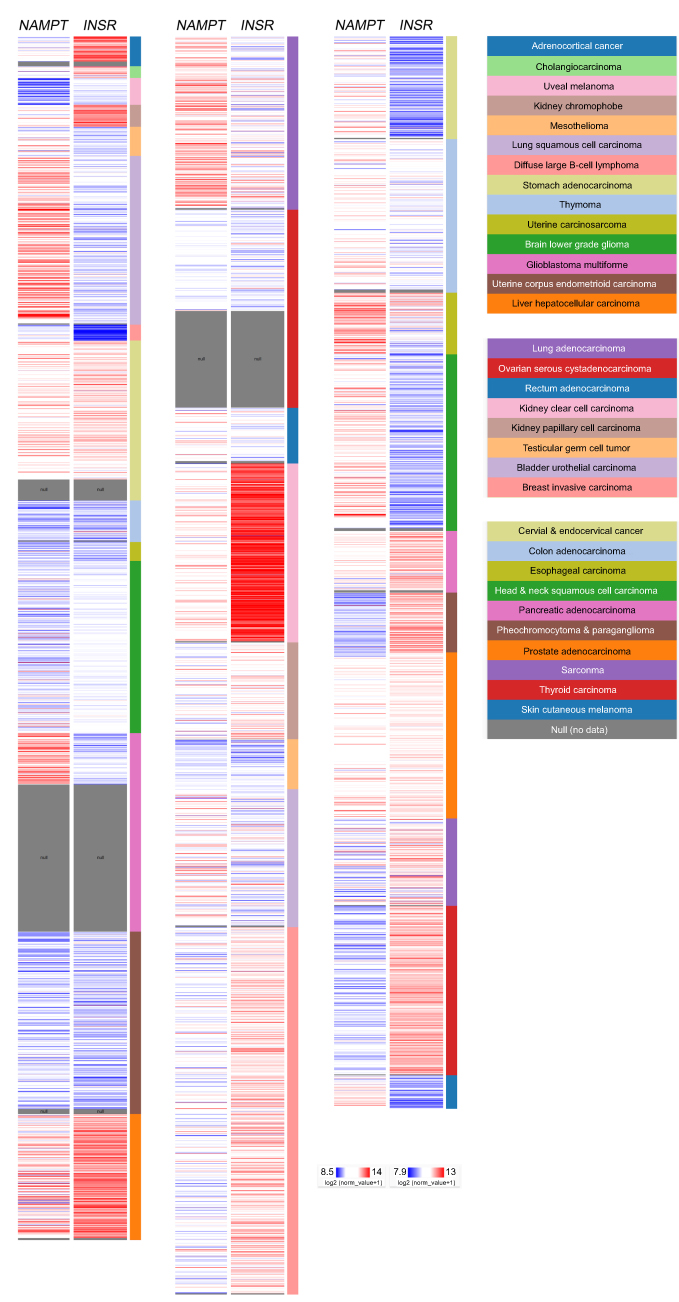 | Figure 1 Relative expression of visfatin and potential visfatin receptor in the pan-cancer panel. In The Cancer Genome Atlas pan-cancer dataset, the relative visfatin (NAMPT) and potential visfatin receptor (INSR) expression levels were presented on a pan-cancer scale. Red color in heat map represents genes with high expression. Blue color in heat map represents genes with low expression. |
Correlation with clinical outcome
Evaluation of the association of the visfatin level with clinical outcome in various types of cancers has been reported. Visfatin expression was shown to correlate with poor overall survival in melanoma patients.13 In addition, hepatocellular carcinoma patients with high serum visfatin levels had shorter overall survival times compared with those with low serum visfatin levels (p<0.001).14 In colorectal carcinoma cases, the results of a Kaplan–Meier analysis of 87 patients indicated a statistically negative correlation between visfatin levels and overall survival probability (p<0.001).15 A study of 176 breast cancer biopsy tissues demonstrated that visfatin is a prognosis marker for its association with poor patient survival.8 Another cohort study in breast cancer further suggested that the combination of high serum visfatin and estrogen receptor-negative status appeared to correlate with the worst disease-free survival (p<0.001).16 The concurrent status of poor survival outcome and high visfatin level was verified in patients with upper tract urothelial carcinoma,17 gastric cancer,18 gastric cancer with diabetes,19 breast cancer20,21 and bortezomib-resistant myeloma.22 Furthermore, the gene expression profile, as determined by microarray and RNA sequencing technology, together with clinical follow-up data were retrieved and analyzed through SurvExpress,23 TCGA12 and the Kaplan–Meier plotter database,24 which illustrates the prognostic impact of visfatin (NAMPT) and its potential receptor (INSR) in specific cancer types (Tables 1 and 2).
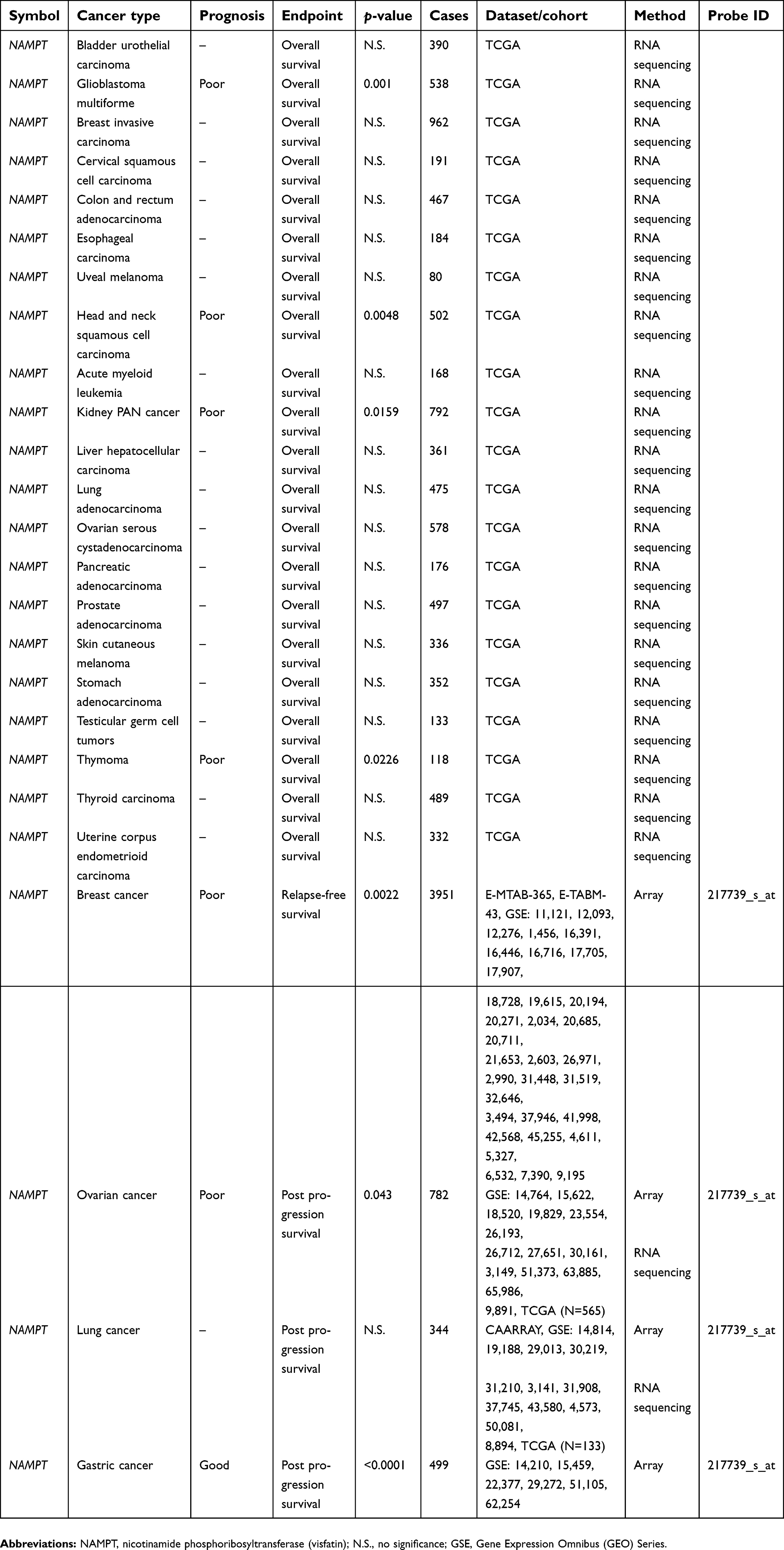 | Table 1 The correlation of visfatin (NAMPT) with cancer patient survival |
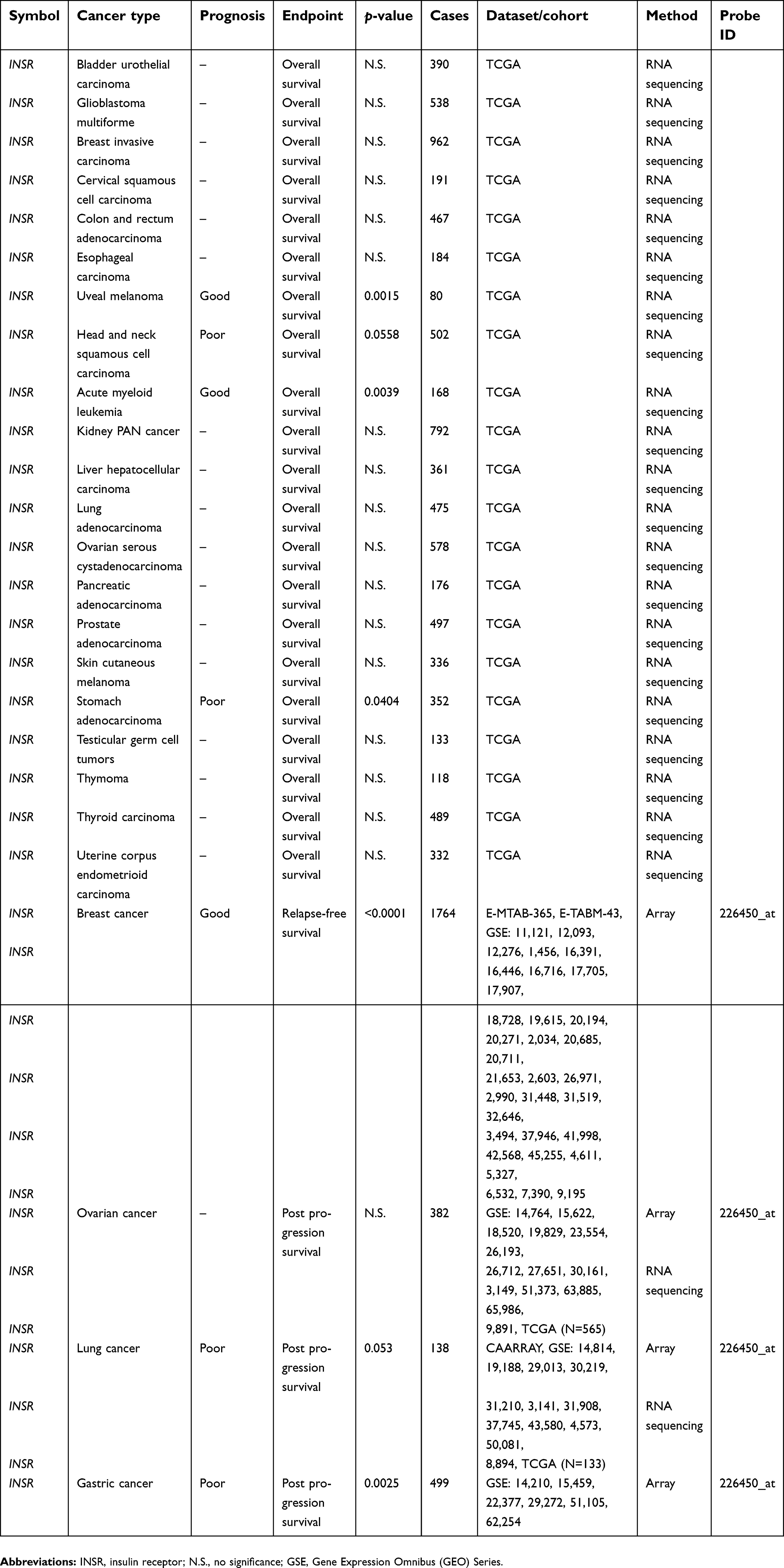 | Table 2 The correlation of visfatin receptor (INSR) with cancer patient survival |
Visfatin and proliferation
The role of visfatin in regulating cancer cell proliferation has also been reported. In melanoma cells, NAMPT promotes proliferation and inhibits p53-dependent apoptosis via the E2F2/SIRT1 axis, and this effect was more significant than the NAD+-driven transcriptional program.13 Visfatin regulated redox adaptative responses by increasing the activity of anti-oxidative enzyme including SOD, CAT and GSH-Px along with an increase in the proliferation rate via [3H]thymidine incorporation in Me45 human malignant melanoma cells.25 Visfatin was revealed to preferentially induce the proliferation of HepG2, Hep3B and HuH7 human hepatoma cells compared with normal hepatocyte Hc cells. This biological effect was reversed by PI3K/MEK1/GSK3β pathway inhibitors.26 In one study, visfatin also promoted breast tumor growth by upregulating Notch1, which contributed to activation of the NF- B pathway.27 Furthermore, visfatin induced proliferation of breast cancer MCF-7 and MDA-MB-231 cells, whereas the effect was further inhibited by treatment with AKT and ERK1/2 inhibitors.28 Induced MCF-7 cell proliferation as well as an increase in both extracellular and intracellular NAD concentrations using visfatin have also been reported by another group, which showed that the effect could be abolished by inhibition of visfatin enzymatic activity.29 In endometrial cancer, visfatin increased the proliferation of both Ishikawa and KLE cells via promoting G1/S phase progression through the PI3K/Akt and MAPK/ERK1/2 signaling pathways. The regulation of endometrial carcinoma tumor growth was also revealed in a BALB/c-nu mice model.30 Furthermore, the NAD biosynthetic pathway is essential for RCC growth. The visfatin inhibitor KPT-9274 was shown to interfere with the signaling pathway and resulted in a reduction of G2/M transit as well as an induction of apoptosis in several human RCC cell lines.31 In addition, NAMPT gene silencing by specific siRNA decreased cell proliferation and increased apoptosis in multiple myeloma RPMI 8226 cells.32 The pharmacological inhibition of visfatin by its specific inhibitor FK866 together with gemcitabine revealed the antitumor activity in PDAC cells, and in orthotopic xenograft mouse models.7 A similar inhibitory effect of FK866 was reported in a study of CRC progression. FK866 addition blocked the NAMPT-mediated upregulation of the NAD(H) pool which protected cancer cells against detrimental oxidative stress.4
Visfatin and angiogenesis
Visfatin appears to be involved in the regulation of cancer cell angiogenesis. In breast cancer, visfatin increased the expression of the matrix metalloproteinase (MMP)-2, MMP-9 and vascular endothelial growth factor genes, which demonstrates its potential function in angiogenesis.33 Visfatin induced thromboxane synthase-dependent IL-8 production leading to the activation of angiogenesis in endothelial cells.34 Visfatin exerted an angiogenic effect through activation of the mTOR pathway, thereby increasing the expression of vascular endothelial growth factor (VEGF) and hypoxia inducible factor 1α in human endothelial cells.35 Furthermore, visfatin stimulated fibroblast growth factor 2 gene expression in a Notch1-dependent manner, which triggered tube formation of endothelial cells.36 Visfatin promoted endothelial cell angiogenesis via IL-6, and its functions demonstrated by the tube formation, the rat aortic ring assay, and the mouse Matrigel plug assay were further eliminated by the inactivation of STAT3 signaling or the neutralization of IL-6 activity.37 In addition, visfatin showed the dosage and time-dependent effects in terms of the increase in tube formation capacity via the PI3K/DDAH2/VEGF pathway in human umbilical vein endothelial cells, whereas the angiogenic effect was inhibited by dimethylarginine dimethylaminohydrolase 2 (DDAH2) siRNA and PI3K inhibitors.38 The results from another group further indicated that 24-h treatment with visfatin (400 ng/mL) significantly increased capillary tube formation, the mechanism of which was shown to be via activation of the MCP-1/CCR2 receptor axis in endothelial cells.39 Another group also suggested that visfatin increased gene expression and protein production of VEGF and MMP-2/9, and suppressed expression of tissue inhibitors of MMPs (TIMP-1 and TIMP-2) in a dose-dependent manner, which led to capillary-like tube formation.40 Importantly, visfatin significantly triggered in vivo neovascularization in chick chorioallantoic membrane and mouse Matrigel plug assays.41 The combination of GMX1777, which inhibits visfatin activity, with radiotherapy has been explored in head and neck cancer in vivo. Tumor microvessel density was estimated using the CD31 expression level, which was significantly reduced to 18% after GMX1777 treatment alone (p<0.01) and 4% when combined with radiotherapy (p<0.01) in FaDu tumors.42
Visfatin and metastasis
The visfatin level is clinically correlated with cancer metastasis in different cancer types. Patients with endometrial cancer have significantly higher visfatin levels than controls (p=0.011), and the association of visfatin with deep myometrial invasion (p=0.019) was also observed.43 In addition, elevated levels of visfatin in the serum of small cell lung cancer (SCLC) patients correlated with brain metastasis.44 Furthermore, the positive correlation of plasma visfatin with cancer invasion depth, lymph node metastasis, distant metastasis and tumor node metastasis stage was observed in gastric cancer patients.18 Visfatin also plays critical roles in the modulation of cancer cell metastasis. In non-SCLC, the visfatin plasma level was found to be associated with lymph node metastasis (p=0.015) and distant metastasis (p=0.003). Visfatin (100 ng/ml) triggered wound closure, migration and invasion as well as the upregulation of MMP-2 and MMP-9 in A549 and H358 cells, whereas those effects were blocked by BAY 11-7082 (NF- B inhibitor).45 Similar associations between serum visfatin and clinical pathologic variables were observed in breast cancer patients along with regulation of MDA-MB-231 cell migration and invasion capabilities via c-Abl and STAT3 activation.16 The induction of the epithelial–mesenchymal transition process by visfatin was investigated in CRC, and the results indicated that visfatin could upregulate Snail in CRC cells by activating Akt/GSK-3β/β-catenin signaling.15 Furthermore, visfatin was found to significantly increase the in vitro migration and invasion capability of osteosarcoma MG-63 and HOS cells, and to upregulate MMP-2 and fibronectin expression through the NF- B/IL-6 signaling axis.46 In addition, ascites-derived visfatin was investigated for its potential biological impact during ovarian cancer progression. Ascites-derived visfatin was found to augment migration of ovarian cancer cells via the Rho/ROCK axis, which triggered actin polymerization, actin stress fiber aggregation and formation of lamellipodia and filopodia.47 Importantly, visfatin appeared to promote CC chemokine ligand 2-dependent SCLC NCI-H446 cell migration through human brain microvascular endothelial cells using an in vitro blood–brain barrier model.44 On the contrary, visfatin knockdown in breast cancer cells revealed significant increase in metastatic activity as compared with the control shRNA-transduced cells. It was proposed that the reduction in NAD+ synthesis led to aggressiveness of MDA-MB-231 breast cancer cells.48
Visfatin and drug resistance
Emerging studies have suggested the involvement of visfatin in cancer drug resistance. In CRC, plasma visfatin might serve as a prognostic indicator for poor response upon 5-fluorouracil chemotherapy because a higher visfatin level was observed in patients who experienced disease progression compared with those in partial response and the stable disease group.49 In addition, visfatin appeared to decrease doxorubicin sensitivity in NSCLC A549 and H1793 cells via activation of the Akt/ABCC1 signaling axis. The visfatin protein and mRNA expression levels were found to be significantly increased in doxorubicin-resistant NSCLC cells, and visfatin appeared to increase the binding of Akt toward the ABCC1 promoter region, leading to its upregulation.50 Furthermore, the visfatin level was increased in established doxorubicin-resistant CRC HCT-116 and SW480 cells. Visfatin silencing resulted in the elevation of drug sensitivity in CRC-resistant cells through transcriptional activation of multidrug resistance 1 (MDR1) via p65 activation and nuclear localization.51 The visfatin inhibitor GMX1778 further showed an enhancement in the efficacy of 177Lu-DOTATATE treatment for neuroendocrine tumors.52 Moreover, pharmacological suppression of visfatin by the inhibitor FK866 decreased the NAD level and glycolytic activity, which led to an elevated antitumor capability of gemcitabine in orthotopic xenograft animal models and PDAC cells.7 Another study indicated that suppression of vistatin signaling by the inhibitors FK866 and CHS828 could sensitize glioblastoma to temozolomide treatment via activation of the ROS/JNK axis.53 In myeloma, the combination of NAD+ depletion by FK866 and bortezomib activated caspase 8, caspase 9, caspase 3 and poly(ADP-ribose) polymerase, leading to synergistic effects in triggering cell death and increasing bortezomib sensitivity. The results of that study also suggested the impact of visfatin in myeloma drug resistance.22
Discussion and conclusions
The results from publications and in silico analysis revealed the expression of visfatin and its potential receptor in various types of cancer. Moreover, the critical roles of visfatin in regulating processes relevant to cancer progression, including cancer cell proliferation, angiogenesis, metastasis and drug resistance, have also been illustrated. In this review, we demonstrate the relative levels of visfatin and potential visfatin receptor on a pan-cancer scale which indicates the consensus upregulation of the axis in stomach adenocarcinoma, liver hepatocellular carcinoma, kidney clear cell carcinoma and prostate adenocarcinoma. This points out the potentially critical role of this axis in tumor progression. In addition, the differential expression of visfatin and its potential receptor at the RNA level in specific cancer types suggests that the alterations in transcriptional activity and RNA stability might be of value for further studies on the mechanism of tumorigenesis and cancer progression. As regards exploration of the prognostic value of visfatin and its potential receptor, the signaling axis concurrently serves as a poor prognostic indicator in head and neck squamous cell carcinoma. On the contrary, the prognostic trend of this axis reveals a discrepancy in several types of cancers. Therefore, the impact of this axis in those tumor types remains to be determined based on additional evidence. Moreover, the differences raised by analytic platforms and endpoint designs of studies are also a concern. The differences in the number of cases enrolled in each cohort also limit the prognostic power. In addition to the variations caused by experimental design, another possibility is that the visfatin-dependent effects are alternatively induced by other receptors. It is noticed that the receptors mediating visfatin’s downstream signaling are still under debate.25,54,55
Acknowledgments
It is grateful for obtaining help from the Genomic Medicine Core Laboratory, Chang Gung Memorial Hospital, Linkou, Taiwan. This study was funded by Chang Gung Memorial Hospital, Linkou (CMRPG3G0612) and Ministry of Science and Technology (MOST 106-2314-B-182A-004 -MY2, 107-2314-B-182A-158 -MY3), Taiwan to Dr Tsung-Chieh Lin.
Author contributions
The author contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The author reports no conflict of interest in this work.
References
1. Samal B, Sun Y, Stearns G, Xie C, Suggs S, McNiece I. Cloning and characterization of the cDNA encoding a novel human pre-B-cell colony-enhancing factor. Mol Cell Biol. 1994;14(2):1431–1437.
2. Gerner RR, Klepsch V, Macheiner S, et al. NAD metabolism fuels human and mouse intestinal inflammation. Gut. 2018;67(10):1813–1823. doi:10.1136/gutjnl-2017-314241
3. Revollo JR, Grimm AA, Imai S. The NAD biosynthesis pathway mediated by nicotinamide phosphoribosyltransferase regulates Sir2 activity in mammalian cells. J Biol Chem. 2004;279(49):50754–50763. doi:10.1074/jbc.M408388200
4. Hong SM, Hwang SW, Wang T, et al. Increased NAD(H) pool promotes colon cancer progression by suppressing ROS level. Cancer Sci. 2018;110(2):629-638. doi: 10.1111/cas. 13886
5. Mohammadi M, Mianabadi F, Mehrad-Majd H. Circulating visfatin levels and cancers risk: a systematic review and meta-analysis. J Cell Physiol. 2018;234:5011–5022.
6. Wang XY, Wang JZ, Gao L, et al. Inhibition of nicotinamide phosphoribosyltransferase and depletion of nicotinamide adenine dinucleotide contribute to arsenic trioxide suppression of oral squamous cell carcinoma. Toxicol Appl Pharmacol. 2017;331:54–61. doi:10.1016/j.taap.2017.05.008
7. Ju HQ, Zhuang ZN, Li H, et al. Regulation of the Nampt-mediated NAD salvage pathway and its therapeutic implications in pancreatic cancer. Cancer Lett. 2016;379(1):1–11. doi:10.1016/j.canlet.2016.05.024
8. Sharif T, Ahn DG, Liu RZ, et al. The NAD(+) salvage pathway modulates cancer cell viability via p73. Cell Death Differ. 2016;23(4):669–680. doi:10.1038/cdd.2015.134
9. Zhang HP, Zou J, Xu ZQ, et al. Association of leptin, visfatin, apelin, resistin and adiponectin with clear cell renal cell carcinoma. Oncol Lett. 2017;13(1):463–468. doi:10.3892/ol.2016.5408
10. Sawicka-Gutaj N, Waligorska-Stachura J, Andrusiewicz M, et al. Nicotinamide phosphorybosiltransferase overexpression in thyroid malignancies and its correlation with tumor stage and with survivin/survivin DEx3 expression. Tumour Biol. 2015;36(10):7859–7863. doi:10.1007/s13277-015-3506-z
11. Venkateshaiah SU, Khan S, Ling W, et al. NAMPT/PBEF1 enzymatic activity is indispensable for myeloma cell growth and osteoclast activity. Exp Hematol. 2013;41(6):547–557 e542. doi:10.1016/j.exphem.2013.02.008
12. Zhu J, Sanborn JZ, Benz S, et al. The UCSC cancer genomics browser. Nat Methods. 2009;6(4):239–240. doi:10.1038/nmeth0409-239
13. Zhao H, Tang W, Chen X, et al. The NAMPT/E2F2/SIRT1 axis promotes proliferation and inhibits p53-dependent apoptosis in human melanoma cells. Biochem Biophys Res Commun. 2017;493(1):77–84. doi:10.1016/j.bbrc.2017.09.071
14. Sun Y, Zhu S, Wu Z, et al. Elevated serum visfatin levels are associated with poor prognosis of hepatocellular carcinoma. Oncotarget. 2017;8(14):23427–23435. doi:10.18632/oncotarget.15080
15. Yang J, Zhang K, Song H, et al. Visfatin is involved in promotion of colorectal carcinoma malignancy through an inducing EMT mechanism. Oncotarget. 2016;7(22):32306–32317. doi:10.18632/oncotarget.8615
16. Hung AC, Lo S, Hou MF, et al. Extracellular visfatin-promoted malignant behavior in breast cancer is mediated through c-Abl and STAT3 activation. Clin Cancer Res. 2016;22(17):4478–4490. doi:10.1158/1078-0432.CCR-15-2704
17. Ke HL, Lin HH, Li WM, et al. High visfatin expression predicts poor prognosis of upper tract urothelial carcinoma patients. Am J Cancer Res. 2015;5(8):2447–2454.
18. Lu GW, Wang QJ, Xia MM, Qian J. Elevated plasma visfatin levels correlate with poor prognosis of gastric cancer patients. Peptides. 2014;58:60–64. doi:10.1016/j.peptides.2014.05.016
19. Zhao W, Chen R, Zhao M, Li L, Fan L, Che XM. High glucose promotes gastric cancer chemoresistance in vivo and in vitro. Mol Med Rep. 2015;12(1):843–850. doi:10.3892/mmr.2015.3522
20. Li XY, Tang SH, Zhou XC, Ye YH, Xu XQ, Li RZ. Preoperative serum visfatin levels and prognosis of breast cancer among Chinese women. Peptides. 2014;51:86–90. doi:10.1016/j.peptides.2013.11.010
21. Zhou SJ, Bi TQ, Qin CX, Yang XQ, Pang K. Expression of NAMPT is associated with breast invasive ductal carcinoma development and prognosis. Oncol Lett. 2018;15(5):6648–6654. doi:10.3892/ol.2018.8164
22. Cagnetta A, Cea M, Calimeri T, et al. Intracellular NAD(+) depletion enhances bortezomib-induced anti-myeloma activity. Blood. 2013;122(7):1243–1255. doi:10.1182/blood-2013-02-483511
23. Aguirre-Gamboa R, Gomez-Rueda H, Martinez-Ledesma E, et al. SurvExpress: an online biomarker validation tool and database for cancer gene expression data using survival analysis. PLoS One. 2013;8(9):e74250. doi:10.1371/journal.pone.0074250
24. Gyorffy B, Schafer R. Meta-analysis of gene expression profiles related to relapse-free survival in 1,079 breast cancer patients. Breast Cancer Res Treat. 2009;118(3):433–441. doi:10.1007/s10549-008-0242-8
25. Buldak RJ, Buldak L, Polaniak R, et al. Visfatin affects redox adaptative responses and proliferation in Me45 human malignant melanoma cells: an in vitro study. Oncol Rep. 2013;29(2):771–778. doi:10.3892/or.2012.2175
26. Ninomiya S, Shimizu M, Imai K, et al. Possible role of visfatin in hepatoma progression and the effects of branched-chain amino acids on visfatin-induced proliferation in human hepatoma cells. Cancer Prev Res (Phila). 2011;4(12):2092–2100. doi:10.1158/1940-6207.CAPR-11-0340
27. Park HJ, Kim SR, Kim SS, et al. Visfatin promotes cell and tumor growth by upregulating Notch1 in breast cancer. Oncotarget. 2014;5(13):5087–5099. doi:10.18632/oncotarget.2086
28. Gholinejad Z, Kheiripour N, Nourbakhsh M, et al. Extracellular NAMPT/Visfatin induces proliferation through ERK1/2 and AKT and inhibits apoptosis in breast cancer cells. Peptides. 2017;92:9–15. doi:10.1016/j.peptides.2017.04.007
29. Behrouzfar K, Alaee M, Nourbakhsh M, Gholinejad Z, Golestani A. Extracellular NAMPT/visfatin causes p53 deacetylation via NAD production and SIRT1 activation in breast cancer cells. Cell Biochem Funct. 2017;35(6):327–333. doi:10.1002/cbf.3279
30. Wang Y, Gao C, Zhang Y, et al. Visfatin stimulates endometrial cancer cell proliferation via activation of PI3K/Akt and MAPK/ERK1/2 signalling pathways. Gynecol Oncol. 2016;143(1):168–178. doi:10.1016/j.ygyno.2016.07.109
31. Abu Aboud O, Chen CH, Senapedis W, Baloglu E, Argueta C, Weiss RH. Dual and specific inhibition of NAMPT and PAK4 by KPT-9274 decreases kidney cancer growth. Mol Cancer Ther. 2016;15(9):2119–2129. doi:10.1158/1535-7163.MCT-16-0197
32. Bong IP, Ng CC, Fakiruddin SK, Lim MN, Zakaria Z. Small interfering RNA-mediated silencing of nicotinamide phosphoribosyltransferase (NAMPT) and lysosomal trafficking regulator (LYST) induce growth inhibition and apoptosis in human multiple myeloma cells: a preliminary study. Bosn J Basic Med Sci. 2016;16(4):268–275. doi:10.17305/bjbms.2016.1568
33. Kim JG, Kim EO, Jeong BR, et al. Visfatin stimulates proliferation of MCF-7 human breast cancer cells. Mol Cells. 2010;30(4):341–345. doi:10.1007/s10059-010-0124-x
34. Kim SR, Jung YH, Park HJ, et al. Upregulation of thromboxane synthase mediates visfatin-induced interleukin-8 expression and angiogenic activity in endothelial cells. Biochem Biophys Res Commun. 2012;418(4):662–668. doi:10.1016/j.bbrc.2012.01.072
35. Park JW, Kim WH, Shin SH, et al. Visfatin exerts angiogenic effects on human umbilical vein endothelial cells through the mTOR signaling pathway. Biochim Biophys Acta. 2011;1813(5):763–771. doi:10.1016/j.bbamcr.2011.02.009
36. Bae YH, Park HJ, Kim SR, et al. Notch1 mediates visfatin-induced FGF-2 up-regulation and endothelial angiogenesis. Cardiovasc Res. 2011;89(2):436–445. doi:10.1093/cvr/cvq276
37. Kim JY, Bae YH, Bae MK, et al. Visfatin through STAT3 activation enhances IL-6 expression that promotes endothelial angiogenesis. Biochim Biophys Acta. 2009;1793(11):1759–1767. doi:10.1016/j.bbamcr.2009.09.006
38. Xiao J, Xiao ZJ, Liu ZG, et al. Involvement of dimethylarginine dimethylaminohydrolase-2 in visfatin-enhanced angiogenic function of endothelial cells. Diabetes Metab Res Rev. 2009;25(3):242–249. doi:10.1002/dmrr.939
39. Adya R, Tan BK, Chen J, Randeva HS. Pre-B cell colony enhancing factor (PBEF)/visfatin induces secretion of MCP-1 in human endothelial cells: role in visfatin-induced angiogenesis. Atherosclerosis. 2009;205(1):113–119. doi:10.1016/j.atherosclerosis.2008.11.024
40. Adya R, Tan BK, Punn A, Chen J, Randeva HS. Visfatin induces human endothelial VEGF and MMP-2/9 production via MAPK and PI3K/Akt signalling pathways: novel insights into visfatin-induced angiogenesis. Cardiovasc Res. 2008;78(2):356–365. doi:10.1093/cvr/cvm111
41. Kim SR, Bae SK, Choi KS, et al. Visfatin promotes angiogenesis by activation of extracellular signal-regulated kinase 1/2. Biochem Biophys Res Commun. 2007;357(1):150–156. doi:10.1016/j.bbrc.2007.03.105
42. Kato H, Ito E, Shi W, et al. Efficacy of combining GMX1777 with radiation therapy for human head and neck carcinoma. Clin Cancer Res. 2010;16(3):898–911. doi:10.1158/1078-0432.CCR-09-1945
43. Ilhan TT, Kebapcilar A, Yilmaz SA, et al. Relations of serum visfatin and resistin levels with endometrial cancer and factors associated with its prognosis. Asian Pac J Cancer Prev. 2015;16(11):4503–4508.
44. Liu T, Miao Z, Jiang J, et al. Visfatin mediates SCLC cells migration across brain endothelial cells through upregulation of CCL2. Int J Mol Sci. 2015;16(5):11439–11451. doi:10.3390/ijms160511439
45. Wang G, Tian W, Liu Y, et al. Visfatin triggers the cell motility of non-small cell lung cancer via up-regulation of matrix metalloproteinases. Basic Clin Pharmacol Toxicol. 2016;119(6):548–554. doi:10.1111/bcpt.12623
46. Wang GJ, Shen NJ, Cheng L, Yehan F, Huang H, Li KH. Visfatin triggers the in vitro migration of osteosarcoma cells via activation of NF-kappaB/IL-6 signals. Eur J Pharmacol. 2016;791:322–330. doi:10.1016/j.ejphar.2016.08.029
47. Li Y, Li X, Liu KR, Zhang JN, Liu Y, Zhu Y. Visfatin derived from ascites promotes ovarian cancer cell migration through Rho/ROCK signaling-mediated actin polymerization. Eur J Cancer Prev. 2015;24(3):231–239. doi:10.1097/CEJ.0000000000000064
48. Santidrian AF, LeBoeuf SE, Wold ED, Ritland M, Forsyth JS, Felding BH. Nicotinamide phosphoribosyltransferase can affect metastatic activity and cell adhesive functions by regulating integrins in breast cancer. DNA Repair (Amst). 2014;23:79–87. doi:10.1016/j.dnarep.2014.08.006
49. Slomian G, Swietochowska E, Nowak G, Pawlas K, Zelazko A, Nowak P. Chemotherapy and plasma adipokines level in patients with colorectal cancer. Postepy Hig Med Dosw (Online). 2017;71:281–290.
50. Cao Z, Liang N, Yang H, Li S. Visfatin mediates doxorubicin resistance in human non-small-cell lung cancer via Akt-mediated up-regulation of ABCC1. Cell Prolif. 2017;50(5). doi:10.1111/cpr.12368
51. Yan X, Zhao J, Zhang R. Visfatin mediates doxorubicin resistance in human colorectal cancer cells via up regulation of multidrug resistance 1 (MDR1). Cancer Chemother Pharmacol. 2017;80(2):395–403. doi:10.1007/s00280-017-3365-y
52. Elf AK, Bernhardt P, Hofving T, et al. NAMPT inhibitor GMX1778 enhances the efficacy of 177Lu-DOTATATE treatment of neuroendocrine tumors. J Nucl Med. 2017;58(2):288–292. doi:10.2967/jnumed.116.177584
53. Feng J, Yan PF, Zhao HY, Zhang FC, Zhao WH, Feng M. Inhibitor of nicotinamide phosphoribosyltransferase sensitizes glioblastoma cells to temozolomide via activating ROS/JNK signaling pathway. Biomed Res Int. 2016;2016:1450843. doi:10.1155/2016/1450843
54. Xie H, Tang SY, Luo XH, et al. Insulin-like effects of visfatin on human osteoblasts. Calcif Tissue Int. 2007;80(3):201–210. doi:10.1007/s00223-006-0155-7
55. Fukuhara A, Matsuda M, Nishizawa M, et al. Visfatin: a protein secreted by visceral fat that mimics the effects of insulin. Science. 2005;307(5708):426–430. doi:10.1126/science.1097243
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.