")
Back to Journals » Drug Design, Development and Therapy » Volume 13
The Osteoprotective Effects Of Kaempferol: The Evidence From In Vivo And In Vitro Studies
Authors Wong SK , Chin KY , Ima-Nirwana S
Received 18 August 2019
Accepted for publication 12 September 2019
Published 7 October 2019 Volume 2019:13 Pages 3497—3514
DOI https://doi.org/10.2147/DDDT.S227738
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Georgios Panos
Sok Kuan Wong, Kok-Yong Chin, Soelaiman Ima-Nirwana
Department of Pharmacology, Faculty of Medicine, Universiti Kebangsaan Malaysia, Kuala Lumpur, Malaysia
Correspondence: Soelaiman Ima-Nirwana
Department of Pharmacology, Faculty of Medicine, Universiti Kebangsaan Malaysia, Jalan Yaakob Latif, Bandar Tun Razak, Cheras, Kuala Lumpur 56000, Malaysia
Tel +60 3 9145 9579
Fax +60 3 9145 9547
Email [email protected]
Abstract: Kaempferol is a dietary bioflavonoid ubiquitously found in various types of plant. It possesses a wide range of medicinal properties suggesting its potential clinical utility that requires further investigation. The present review intends to highlight the efficacy of kaempferol and its molecular mechanisms of action in regulating bone metabolism. Many reports have acknowledged the bone-protecting property of kaempferol and kaempferol-containing plants using in vitro and in vivo experimental models. Kaempferol supplementation showed bone-sparing effects in newborn rats, glucocorticoid-induced and ovariectomy-induced osteoporotic models as well as bone fracture models. It achieves the bone-protective effects by inhibiting adipogenesis, inflammation, oxidative stress, osteoclastic autophagy and osteoblastic apoptosis while activating osteoblastic autophagy. The anti-osteoporotic effects of kaempferol are mediated through regulation of estrogen receptor, bone morphogenetic protein-2 (BMP-2), nuclear factor-kappa B (NF-κB), mitogen-activated protein kinase (MAPK) and mammalian target of rapamycin (mTOR) signaling pathways. In summary, kaempferol exhibits beneficial effects on skeleton, thus is potentially effective for the prophylaxis and treatment of osteoporosis.
Keywords: bone, flavonoid, fracture, osteoblast, osteoclast, osteoporosis
Introduction
Osteoporosis is a degenerative bone disorder characterized by deterioration of bone microstructure resulting from excessive bone resorption and declining bone formation. It is a multifactorial disease, whereby aging,1 sex hormone deficiency,2 medication use,3,4 alcoholism,5 nicotine6 and medical conditions (such as diabetes, hypertension, dyslipidemia and metabolic syndrome)7 can initiate and exacerbate progression of bone loss. The choices of medications for osteoporosis include bisphosphonates, teriparatide, receptor activator of nuclear factor-kappa-Β ligand (RANKL) inhibitors (denosumab) and selective estrogen receptor modulators (SERMs). These therapies are effective to prevent further bone loss and fractures but they are accompanied by undesirable side effects and cost issues.8 On the other hand, prophylactic strategies for osteoporosis are limited to behavior interventions, calcium and vitamin D supplementation.9
The discovery of natural products with potential bone-protecting effects might offer alternative treatment agents to overcome the drawbacks of conventional therapies.10–17 Kaempferol (3,5,7-trihydroxy-2-(4-hydroxyphenyl)-4H-1-benzopyran-4-one) and its derivatives are natural flavonoids contributing to the nutritional qualities of fruits and vegetables. It is also present in botanic plants commonly used as traditional medicines, such as Ginkgo biloba, Moringa oleifera and propolis.18 Kaempferol is referred to as a nutraceutical due to its various health benefits previously proven scientifically, which include cardioprotective, neuroprotective, anxiolytic, analgesic, anti-allergic, anti-platelet aggregation, anti-cancer, anti-microbial, anti-obesity, anti-hyperglycemic, anti-hypertensive, anti-hyperlipidemic, anti-aging, anti-oxidative, anti-inflammatory and anti-osteoporotic effects [reviewed by Calderon-Montano et al18 and Imran et al19]. Some of the medicinal properties of kaempferol are directly associated with its bone-sparing effects, which will be reviewed in the following discourse.
This review article aims to provide the in vivo and in vitro experimental evidence surrounding the efficacy of kaempferol in preventing bone loss. This review also discusses the mechanisms of action of kaempferol and its potential as a therapeutic agent for the treatment of osteoporosis.
Literature Search
Literature search was performed using PubMed and Medline databases from June 1, 2019 to June 30, 2019, using the string “kaempferol AND (bone OR osteoporosis OR fracture OR osteoblast OR osteoclast)”. Only original research articles written in English, published since the inception of the databases until June 30, 2019, were included. The titles and abstracts were screened, and full texts of the relevant articles were retrieved.
In Vivo Studies On The Effects Of Kaempferol On Bone
The bone-sparing action of kaempferol in animals has been identified (Table 1). Using newborn Sprague–Dawley rats as an animal model, 5 μM kaempferol was injected into the top of the periosteum of the parietal bones for 12 days. Histological analysis of parietal bones showed that calcification at the area of new bone formation was increased. Immunostaining with bone sialoprotein (BSP), osterix (OSX) and Runt-related transcription factor 2 (Runx-2) antibodies showed that the expression of these proteins was enhanced by kaempferol treatment. The bone area and number of osteoblasts were significantly increased in the kaempferol-treated group compared to the vehicle-treated control group. Osteoblasts have angular-shaped cytoplasm with nuclear polarity, reiterating that the osteoblasts were in the state of active osteoblast differentiation.20 Several other studies investigated the anti-osteoporotic effects of kaempferol using an animal model of bone loss caused by estrogen deficiency, whereby the animals were bilaterally ovariectomized (OVX) to mimic postmenopausal osteoporosis in elderly women. Trivedi and co-authors found that kaempferol at 5 mg/kg prevented trabecular bone loss in the whole femur, femoral neck of the femur, proximal tibia, the whole vertebra and L3 vertebra. The compression test indicated that L3 vertebra of the kaempferol-treated animals required more compressive energy than the negative controls. Kaempferol also inhibited bone turnover by lowering the serum alkaline phosphatase (ALP) in the OVX rats. The bone marrow cells derived from the kaempferol-supplemented OVX group had higher mineralized nodules but lower adipocytes compared to the vehicle-supplemented OVX group.21 A recent study demonstrated that oral administration of kaempferol (5 mg/kg) for 8 weeks increased femoral bone mineral density (BMD) and Young’s modulus of elasticity but decreased osteocalcin (OCN) and RANKL in OVX rats. Histologically, the OVX rats receiving kaempferol had higher bone volume/total volume (BV/TV), trabecular bone area (B.Ar), trabecular bone perimeter (B.Pm) and bone surface/total volume (BS/TV) relative to the negative control animals.22 Kaempferitrin, another name for kaempferol-3,7-dirhamnoside, at the dose of 8 or 16 mg/kg had been shown to increase BMD, bone mineral content (BMC), tissue mineral content, tissue mineral density, bone volume fraction, trabecular number (Tb.N), connectivity density (Conn.D) and decrease trabecular separation (Tb.Sp) in the OVX rats. Kaempferitrin also influenced the levels of bone formation and resorption markers, whereby a higher level of ALP, as well as lower levels of cathepsin K and tartrate-resistant acid phosphatase (TRAP), were observed after the treatment.23
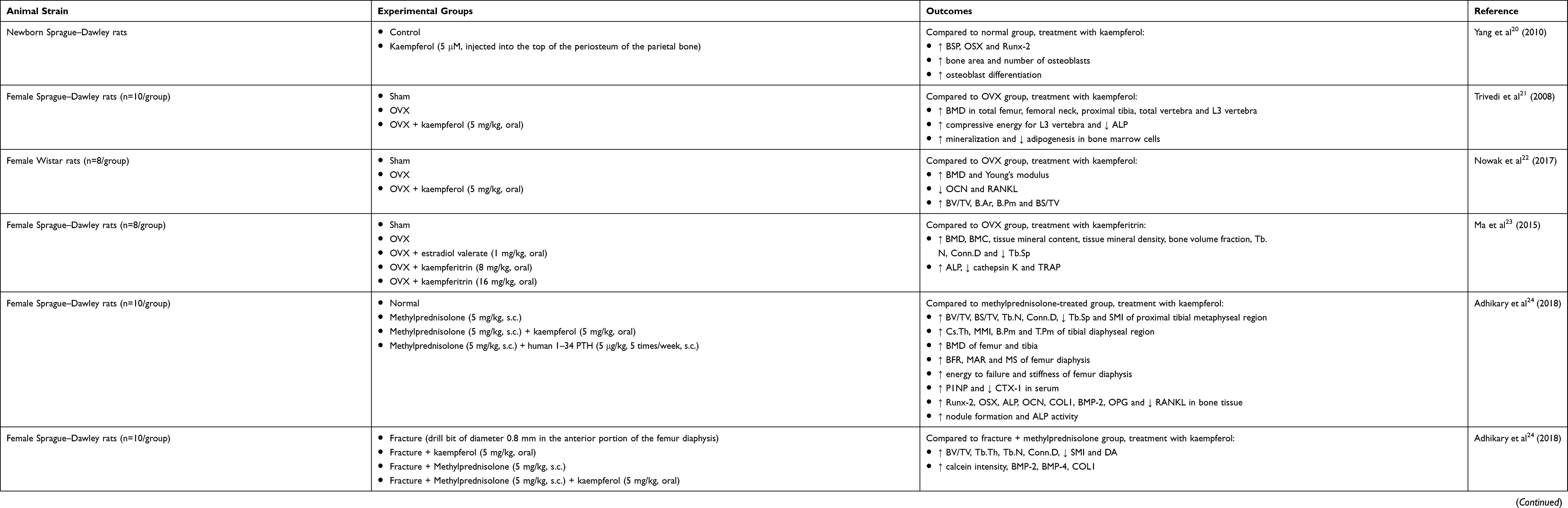 |
Table 1 In Vivo Studies On The Bone-Protecting Properties Of Kaempferol |
Apart from the surgical-castrated animal model, Adhikary et al used corticosteroid-induced osteoporotic and fractured animals to explore the osteoprotective and bone healing properties of kaempferol, respectively. Methylprednisolone (a corticosteroid hormone) was injected subcutaneously (s.c.) into female Sprague–Dawley rats to induce bone loss. The animals receiving kaempferol (5 mg/kg) for four weeks had higher BV/TV, BS/TV, Tb.N, Conn.D as well as lower Tb.Sp and structure model index (SMI) at the proximal tibial metaphyseal region than the animals without treatment of kaempferol. Examination of the cortical bones revealed that higher cross-sectional thickness (Cs.Th), mean polar moment of inertia (MMI), B.Pm and tissue perimeter (T.Pm) was detected at tibial diaphyseal region in the kaempferol-treated group. Oral administration of kaempferol also increased BMD, bone strength, bone formation-related genes [Runx-2, OSX, ALP, OCN, collagen type I (COL1), bone morphogenetic protein-2 (BMP-2) and osteoprotegerin (OPG)], as well as decreasing bone resorption-related gene (RANKL) in methylprednisolone-induced osteoporotic animals. In another animal model, a fracture was created by inserting a drill bit (diameter = 0.8 mm) in the anterior portion of femur diaphysis. Micro-computed tomography analysis showed that kaempferol (5 mg/kg) increased BV/TV, trabecular thickness (Tb.Th), Tb.N, Conn.D, reduced SMI and degree of anisotropy (DA). Kaempferol also caused acceleration of bone regeneration at the fractured site (indicated by increased calcein intensity) and increment in osteogenic markers [indicated by increased COL1, BMP-2 and bone morphogenetic protein-4 (BMP-4)].24 Moreover, kaempferol (0.2, 0.5 and 5 mg/kg for three weeks) enhanced the healing process of fracture by increasing new bone formation, endochondral ossification, callus diameter, size and density in fractured male mice.25 Similarly, Kim and co-researchers found that kaempferol (0.2, 1.0 and 5 mg/kg for 21 days) raised ultimate force and fracture energy at the fracture site in male mice.26
Kaempferol exhibits low oral bioavailability due to its poor absorption and rapid elimination after oral administration, thus hindering its utility as a promising therapeutic agent and leaving room for improvements. These limitations were improved by the fabrication of layer-by-layer thin-film assembly to encapsulate drug biomolecules. The entrapment in multilayer thin film ensures the biological stability and sustained release of kaempferol. Gupta et al explored the effects of kaempferol and formulated kaempferol in layer-by-layer matrix on bone in female OVX rats. The findings of this study pinpointed that 1 mg/kg kaempferol increased BMD (at whole femur, femur diaphysis, tibia head and total vertebra), femoral stiffness and mineralization of osteoblasts. The formulated kaempferol at the same dose further elevated all these measured parameters in the OVX rats.27 In line with these findings, 5 mg/kg of kaempferol was found to improve bone formation [evidenced by higher mineral apposition rate (MAR) and bone formation rate (BFR)] and trabecular microarchitecture (evidenced by higher Tb.Th, BV/TV, Tb.N but lower SMI and Tb.Sp) in osteopenic rats. An engineered layer-by-layer matrix of kaempferol was more superior in improving the bone indices as compared to free kaempferol.28 Kaempferol has been incorporated with titanium dioxide (TiO2) frequently utilized as a biomaterial for medical implants, particularly in dental and orthopedic implants in a study to examine the bone-regenerating effects. The in vivo study by Tsuchiya et al (2018) revealed that the implantation of kaempferol-immobilized TiO2 in tibias showed no signs of osteonecrosis, inflammation, loosening and dislocation, demonstrating that the implants were well connected to the bones. The results from histological detection were further confirmed by quantitative analysis, whereby the bone-implant contact rate in the tibia of kaempferol-immobilized TiO2 implant group was significantly higher than the group implanted with TiO2 without kaempferol. These findings reiterated the fact that implantation resulted in better cell proliferation and osteogenic differentiation, thus enhancing bone formation around the implants.29
In Vitro Studies And Mechanism Of Action Of Kaempferol As A Bone-Protecting Agent
A wide array of laboratory studies investigated the direct effects of kaempferol on osteoblastic cells or osteoblastic precursor cells (Table 2). Kaempferol did not show any cytotoxic effect on bone marrow-derived mesenchymal stem cells (MSCs) at the concentration up to 100 μM.24,30 Kaempferol, extracted from Polygonum tinctorium, stimulated the differentiation and mineralization of murine pre-osteoblastic MC3T3-E1 cells, as seen by augmentation of ALP activity and calcification.31 Osteoblast-like UMR-106 cells treated with kaempferol had significantly higher expression of Runx-2, OSX and BSP.20 The presence of kaempferol also resulted in increased calcium content in the rat femoral diaphyseal and metaphyseal tissue culture.32 Apart from that, stimulation of kaempferol-immobilized TiO2 in rat bone marrow stromal cell culture promoted ALP activity, calcium deposition and osteoblast differentiation-related genes such as Runx-2, OCN, osteonectin and osteopontin (OPN).29
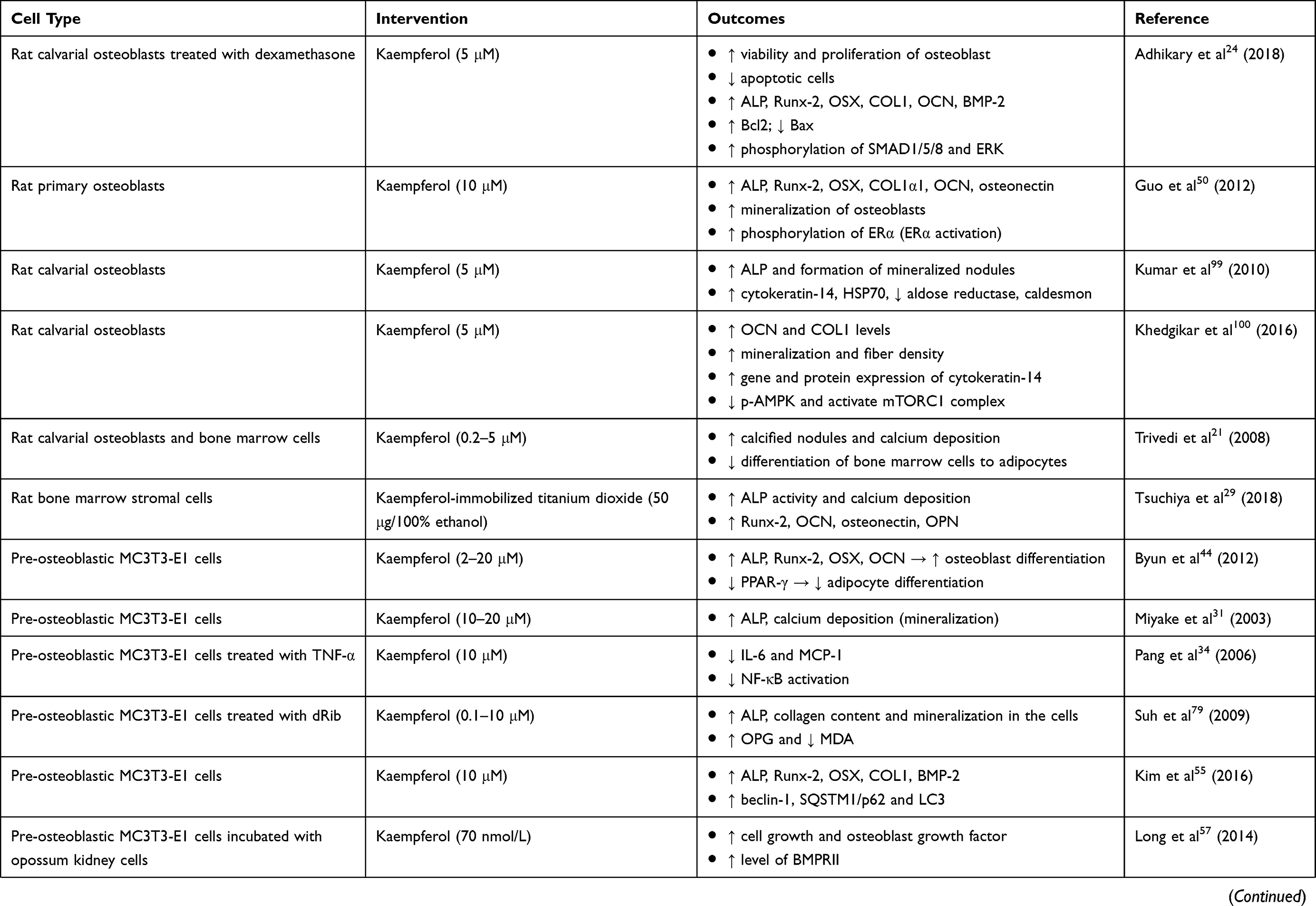 | 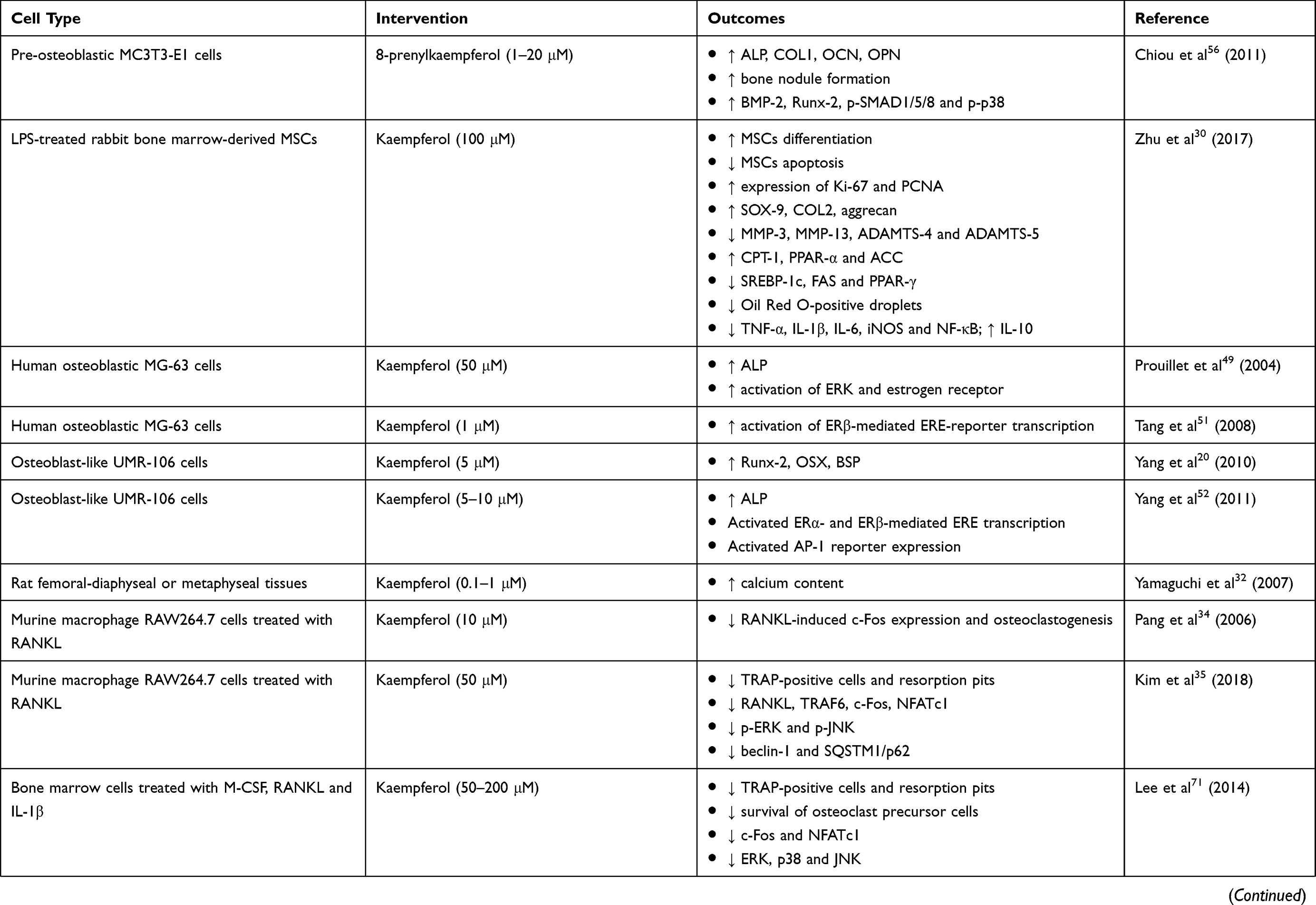 | 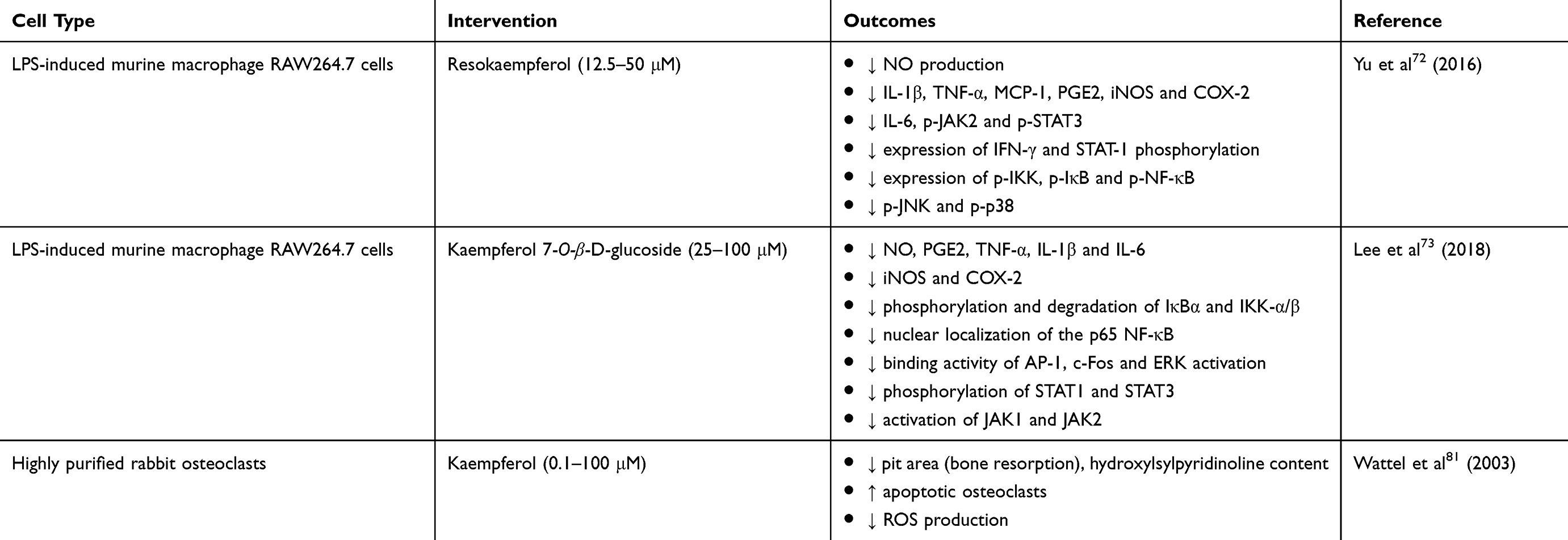 |
Table 2 In Vitro Studies On The Bone-Protecting Properties Of Kaempferol |
In an in vitro study, mature osteoclasts were generated from osteoclast precursors (such as monocyte/macrophage cells and bone marrow cells) in the presence of macrophage-colony stimulating factor (M-CSF) and RANKL.33 Several in vitro studies showed the anti-osteoclastogenic effects of kaempferol (Table 2). In murine macrophage RAW264.7 cells treated with RANKL, kaempferol was shown to abrogate RANKL-induced formation of TRAP-positive multinucleated cells and resorption pits, which are the indicator for osteoclast differentiation. The downregulation of osteoclastogenic factors including RANKL, Fos proto-oncogene (c-Fos), nuclear factor of activated T-cells cytoplasmic 1 (NFATc1) and tumor necrosis factor receptor-associated factor 6 (TRAF6) were also observed in kaempferol-treated cells.34,35 Other in vitro studies were performed to illustrate the mechanisms of action of kaempferol as a potential anti-osteoporotic agent.
Effects Of Kaempferol On The Fate Of Mesenchymal Stem Cells
Bone is formed through the endochondral (for long, short and irregular bones) and intramembranous ossification (for flat bones). During endochondral ossification, MSCs differentiate into chondrocytes forming the cartilage matrix and followed by gradual substitution by the bone. Meanwhile, intramembranous ossification is a process of direct differentiation of MSCs into osteoblasts.36 Matrix metalloproteinases (MMPs) are highly expressed in bone and cartilage and appear to have a role in endochondral ossification during bone modeling and remodeling.37 MMPs are proteolytic enzymes driving extracellular matrix (ECM) remodeling, chondrocyte proliferation and differentiation, osteoblast, osteoclast and osteocyte viability and function.38 Matrix metalloproteinases can be categorized into several subgroups according to their substrate preference. Collagenases (MMP-1, MMP-8, MMP-13 and MMP-18) degrade COL1, collagen type 2 (COL2) and collagen type 3 (COL3). Gelatinases (MMP-2, MMP-9) degrade gelatin and other ECM proteins such as laminin and aggrecan. Stromelysins (MMP-3, MMP-10 and MMP-11) cleave non-collagen molecules in the ECM.39 In addition, a disintegrin and metalloproteinase with thrombospondin motifs (ADAMTS) are also aggrecanases and proteoglycanases that control the structure and function of ECM.40 Hence, the alteration of MMP and ADAMTS level determines the quality and quantity of bone.
Mesenchymal stem cells are progenitor cells capable of equally differentiating into multiple cell lineages, including adipocytes, osteoblasts and chondrocytes under physiological condition. Hence, they are reciprocally regulated during pathological conditions. The tendency of MSCs differentiation into adipocytes rather than osteoblasts and chondrocytes results in the progression of osteoporosis, thereby suggesting the possible association between obesity-related conditions and bone loss.7,41,42 In a state of over-nutrition, the underlying mechanisms include: (1) the downregulation of peroxisome proliferator-activated receptor-alpha (PPAR-α) suppresses the expression of carnitine palmitoyl transferase-1 (CPT-1, an enzyme that facilitates in the β-oxidation of long-chain fatty acids); (2) the upregulation of sterol regulatory element-binding proteins-1c (SREBP-1c) increases fatty acid synthase (FAS) and acetyl coenzyme A carboxylase (ACC), resulting in fatty acid synthesis and reduced capacity in fatty acid oxidation; (3) the upregulation of peroxisome proliferator-activated receptor-gamma (PPAR-γ) increases the production of lipoprotein lipase (LPL) to hydrolyze triglycerides to two free fatty acids and one monoglycerol molecule.43
Kaempferol has a role in modulating these transcription factors that guide MSCs to commit to the osteoblastic and chondrogenic lineage. Increased osteoblast mineralization and calcium deposition but decreased adipocyte differentiation and lipid accumulation were detected in bone marrow cells after kaempferol stimulation.21 Kaempferol promoted the differentiation of MSCs into osteoblasts by enhancing the expression of ALP, Runx-2, OSX and OCN but inhibited the differentiation of MSCs into adipocyte by downregulating PPAR-γ.44 In another in vitro study using lipopolysaccharides (LPS)-treated rabbit bone marrow-derived MSCs, kaempferol promoted cell viability and decreased apoptosis.30 The expression of Ki-67 protein (a mitosis-associated nuclear antigen) and proliferating cell nuclear antigen (PCNA) was found underexpressed in LPS-treated cells and returned to normal after kaempferol treatment. Concomitantly, kaempferol promoted osteogenesis through suppressing matrix degradation. The increase in levels of chondrogenic markers [SRY-Box 9 (SOX-9), COL2 and aggrecan] and the decrease in levels of matrix-degrading enzymes (MMP-3, MMP-13, ADAMTS-4 and ADAMTS-5) were observed.30 Kaempferol also stimulated the expression of lipid catabolism-related genes (CPT-1 and PPAR-α) while suppressed the expression of lipid anabolism-related genes (SREBP-1c, FAS, ACC and PPAR-γ)].30
Effects Of Kaempferol In Eestrogen Receptor Signaling
Estrogen deficiency is a major cause of osteoporosis in women after menopause. Estrogen receptors exist in two types, namely estrogen receptor-alpha (ERα) and estrogen receptor-beta (ERβ). The expression of both ERα and ERβ on osteoblasts, osteocytes, osteoclasts and bone marrow stromal cells suggests that bone cells are direct targets for estrogen.45 The protective effects of estrogen on bone through genomic and non-genomic pathways have historically been established. In the traditional genomic pathway, the binding of estrogen to its receptor causes conformational changes on estrogen receptor with the dissociation of heat shock protein 90 (HSP90) and heat shock protein 70 (HSP70) to form estrogen receptor dimer. Subsequently, the dimer translocates into the nucleus and binds directly to a specific estrogen-responsive elements (ERE) sequence to enhance its transcriptional response.46 Apart from that, the non-genomic action of estrogen-estrogen receptor complex may be mediated through the activation of downstream signaling molecules including phosphoinositide 3-kinases (PI3K)/protein kinase B (Akt), mitogen-activated protein kinase (MAPK) and protein kinase C (PKC).47 The activation of estrogen receptor signaling, whether through classical or non-classical pathways, leads to the suppression of osteoclast differentiation, inhibition of osteoblastic apoptosis, repression of inflammatory cytokines and induction of osteoclastic apoptosis.48
In vivo studies also showed that kaempferol exerted potential in counteracting the adverse effects of estrogen depletion on the skeleton. An earlier in vitro study demonstrated that kaempferol increased ALP activity in MG-63 osteoblasts, but this activity was reduced after treatment with extracellular-regulated kinase (ERK) pathway inhibitor and estrogen receptor antagonist. These findings suggested that the increase of ALP activity by kaempferol was due to activation of ERK, which was the downstream target of estrogen receptor activation.49 Several in vitro studies were performed to examine the effects of kaempferol on ERα or ERβ activation of ERE gene transcription. Kaempferol was shown to induce luciferase activity in osteoblasts expressing ERE, ERα phosphorylation, ALP activity, bone differentiation markers (Runx-2, OSX, COL1, OCN and osteonectin) and osteoblastic mineralization.50 Another study by Tang et al confirmed that kaempferol activated ERβ-mediated ERE-reporter transcription in MG-63 osteoblasts.51 In accordance with former results, kaempferol isolated from Cuscuta chinensis raised ALP activity in osteoblast-like UMR cells as well as activating both ERα- and ERβ-mediated ERE gene transcription.52
Effects Of Kaempferol In BMP-2 Signaling Pathway
Bone morphogenetic protein-2 is a member of transforming growth factor-beta (TGF-β) superfamily. It functions to regulate the differentiation of MSCs during skeletal development, bone formation and bone homeostasis by binding to its tetrameric receptor complex. The binding of BMP-2 to its receptor transduces signal via the suppressor of mothers against decapentaplegic (SMAD)-dependent and non-canonical SMAD-independent (p38 MAPK) signaling.53 The SMAD-dependent signaling involves the formation of SMAD1/5/8-SMAD4 complex which induces the expression of Runx-2. In addition, Runx-2 can be phosphorylated and activated by BMP-2-induced p38 MAPK activation. Thus, Runx-2 is a transcription factor that acts as the center of convergence for the SMAD and p38 MAPK signaling pathways to orchestrate the amplification of its own gene activation and the expression of other osteogenic factors leading to osteoblastogenesis.54
Adhikary et al reported that the osteogenic action of kaempferol was attributable to the upregulation of BMP-2 and sustained phosphorylation of SMAD1/5/8.24 The activation of SMAD1/5/8 via BMP-2 signaling caused by kaempferol treatment resulted in stimulated osteoblast proliferation, exhibited by the high expression levels of ALP, Runx-2, OSX, COL1 and OCN in dexamethasone-induced rat calvarial osteoblasts.24 Similarly, kaempferol upregulated the expression of BMP-2 along with other osteoblast-activated factors (ALP, Runx-2, OSX and COL1) in pre-osteoblastic MC3T3-E1 cells.55 8-prenylkaempferol, isolated from Sophora flavescens, accelerated ALP activity, osteogenic markers (OCN, OPN, COL1 and Runx-2) expression as well as bone nodules formation. This improvement was associated with increased BMP-2 expression that subsequently caused phosphorylation of SMAD1/5/8 and p38 MAPK. Treatment with noggin (a BMP-2 antagonist) blocked BMP-2-induced ALP activity, phosphorylation of SMAD1/5/8 and p38, thus suggesting that BMP-2 signaling was implicated in the osteogenic action of 8-prenylkaempferol.56 Apart from the protecting action of BMP-2 produced in osteoblasts, bone morphogenetic protein receptor II (BMPR II) was identified in opossum kidney cells.57 The authors found that kaempferol increased cell growth, secretion of osteoblast growth factor and level of BMPR II in opossum kidney cells. Findings from this study implied that kaempferol stimulated kidney repair which indirectly stimulates bone formation.57
Effects Of Kaempferol In Inflammation
Inflammation is closely relevant to bone loss as numerous inflammatory cytokines are recognized to inhibit osteoblastogenesis58 and stimulate osteoclastogenesis.59 Tumor necrosis factor-alpha (TNF-α) and interleukin-1 (IL-1), apart from their direct role in promoting osteoclastogenesis and bone resorption, also stimulate osteoblasts to release other bone-resorbing cytokines such as interleukin-6 (IL-6) and monocyte chemoattractant protein (MCP-1).60,61 The amplification of inflammatory response increases the inhibitory effect on osteoblast activities and stimulatory effect on osteoclast activities.62 Specifically, these cytokines suppressed ALP activity, gene expression for ALP, Runx-2, OSX, osteonectin and OPN.58 The underlying mechanisms of inflammatory bone loss are mediated partly through (a) the upregulation of sclerostin (SOST) and Dickkopf-related protein 1 (DKK-1) that inhibits the canonical Wnt/Frizzled/β-catenin pathway, (b) the activation of nuclear factor-kappa B (NF-κB), as well as (c) the activation of signal transducers and activators of transcription (STAT), which in turn inhibits MAPK activities.54 These events subsequently contribute to reduced expression of osteogenic factors (such as Runx-2, OSX, COL1α1, ALP, OCN, OPG and BMP-2) and elevated expression of osteoclastogenic factors (such as RANKL) in osteoblasts.10
The ratio of OPG and RANKL determines the fate of osteoclast precursors, whether osteoclastogenesis is promoted or inhibited. The expression of OPG and RANKL is sensitive to inflammatory cytokines. The imbalance in inflammatory response, characterized by the increase in pro-inflammatory cytokines and the decrease in anti-inflammatory cytokines, directs the inhibition of OPG and release of RANKL by osteoblasts to stimulate osteoclast differentiation and bone resorption.63,64 The binding of RANKL to RANK on the surface of osteoclast recruits the key downstream adaptor protein TRAF6. Multiple essential signaling pathways are further activated including the MAPK, NF-κB, NFATc1 and PI3K/Akt which ultimately lead to osteoclastogenesis and bone resorption.10,65 On the other hand, OPG competitively binds to RANKL, thus blocking the interaction with RANK and preventing excessive bone resorption.66
Nitric oxide (NO) is an important multifunctional signaling molecule regulating both physiological and pathological conditions in bone. It appears to exert biphasic effects by affecting the bone formation and resorption processes in osteoblasts and osteoclasts. Bone formation is enhanced while bone resorption is suppressed in low concentration of NO and vice versa.67 Inducible nitric oxide synthase (iNOS) is one of the three isoforms of NO synthase catalyzing the production of NO from L-arginine. iNOS can be expressed to produce large quantities of NO in response to lipopolysaccharide and pro-inflammatory cytokines.68 Using murine macrophage RAW264.7 cells, RANKL triggers NO production and iNOS expression through NF-κB activation thereby enhancing osteoclast formation.69 In vivo studies showed that wild-type C57BL/6 mice with iNOS genotype resulted in bone depletion (decreased bone volume and bone formation) after ovariectomy. Conversely, bone loss did not occur in iNOS-knockout mice and wild-type mice treated with iNOS inhibitor.70
Kaempferol potentially protects the bone through its anti-inflammatory property on osteoblastic cells. The inhibitory effect of kaempferol on LPS-induced inflammation in rabbit bone marrow-derived MSCs was reported, whereby the levels of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6, iNOS and MCP-1) declined and level of anti-inflammatory cytokine [interleukin-10 (IL-10)] elevated via inhibition of NF-κB nuclear translocation.30 Congruent with these findings, kaempferol also inhibited the secretion of IL-6, MCP-1 and activation of NF-κB in TNF-α-induced MC3T3-E1 cells.34 Inflammatory markers have been shown to further stimulate osteoclastogenesis. Addition of IL-1β into bone marrow cells treated with M-CSF and RANKL significantly increased TRAP-positive cells, resorption pit area, survival of osteoclast precursor cells, expression of c-Fos and NFATc1. Treatment of kaempferol reversed all these parameters in IL-1β-stimulated, RANKL-mediated bone marrow cells. The postulated mechanisms include the inhibition of intracellular MAPK, evidenced by decreased phosphorylation of ERK, p38 and c-Jun N-terminal kinase (JNK).71 In another study, more comprehensive mechanisms of the anti-inflammatory effects of kaempferol have been depicted. LPS triggered inflammation in murine macrophage RAW264.7 cells, whereby the production of cyclooxygenase-2 (COX-2), prostaglandin E2 (PGE2), NO, iNOS, IL-1β, TNF-α, MCP-1 and IL-6 were elevated. Treatment with resokaempferol exerted inhibitory effects on all these parameters, indicating its anti-inflammatory capacity. The molecular mechanisms involved were the activation (phosphorylation) of Janus kinase 2 (JAK2)/signal transducer and activator of transcription 3 (STAT3), signal transducer and activator of transcription 1 (STAT1), NF-κB and MAPK (including JNK and p38) following LPS stimulation, whereby the production of inflammatory mediators ensued. Resokaempferol alleviated inflammation by blocking the activation of these signaling pathways by LPS.72 In agreement with previous findings, kaempferol 7-O-β-D-glucoside (extracted from Cudrania tricuspidata) was capable of suppressing the expression of inflammatory mediators through inactivation (reduced phosphorylation) of NF-κB, AP-1 and JAK/STAT signaling pathways in LPS-treated RAW264.7 macrophages.73
Effects Of Kaempferol In Oxidative Stress
Oxidative stress occurs as a result of excessive production of reactive oxygen species (ROS) not counteracted by the antioxidant capacity in the body.74 High levels of ROS have been recognized as the pathogenic culprit in bone loss. The increased production of ROS elicits a spectrum of events that inhibit osteoblast differentiation and osteoclast apoptosis but induce osteoclast activity and osteoblast apoptosis. Hydrogen peroxide-induced oxidative stress causes a reduction in osteogenic markers ALP, COL1, BSP and Runx-2; thus inhibiting osteoblastic differentiation.75,76 Apart from that, ROS affects osteoblast lifespan by inducing apoptosis.77,78 The anti-oxidative effect of kaempferol was examined by Suh et al using MC3T3-E1 cells treated with 2-deoxy-D-ribose (dRib), a reducing sugar that produces ROS through the process of auto-oxidation and protein glycosylation resulting in osteoblast dysfunction.79 Kaempferol resulted in increased MC3T3-E1 cells growth, ALP activity, collagen content, mineralization and OPG secretion in the cells. Kaempferol also alleviated oxidative stress by reducing the malondialdehyde (MDA) contents in the dRib-treated MC3T3-E1 cells.79
Despite its influence in the inhibition of osteoblast activity and function, ROS promotes osteoclastogenesis and inhibits osteoclast apoptosis through elevation of RANKL production in the osteoblastic lineage.80 The ROS-stimulated osteoclastogenesis has been shown to be mediated through multiple mechanisms, including the NF-κB and ERK signaling pathways.80 Kaempferol exerted potent inhibitory effects on in vitro bone resorption through alleviation of oxidative stress and induction of osteoclast apoptosis. Kaempferol inhibited bone resorption (evaluated by reduced resorption pit formation and hydroxylsylpyridinoline content) as well as inducing osteoclastic programmed cell death (evaluated by an increased number of apoptotic osteoclasts) in highly purified rabbit osteoclasts. Furthermore, kaempferol showed its anti-oxidative property by lowering the intracellular ROS production.81
Effects Of Kaempferol In Autophagy
Autophagy, also known as autophagocytosis, is a major intracellular protein degradation pathway responsible for the removal of damaged cells in order to regenerate newer and healthier cells.82 There are four major steps involved in the process of autophagy, namely initiation/nucleation, elongation, maturation and degradation.83 In the initiation step, phagophore is formed facilitated by class III PI3K complex (consists of beclin-1, Vps34, Vps15, Ambra1 and UVRAG) as well as ULK1/ULK2. Herein, beclin-1 is the key autophagic regulator as it interacts with other co-factors to form the multiprotein complex.84 In the elongation step, phagophore is elongated forming an autophagosome facilitated by LC3II protein (converted from LC3I via lipidation process) and several autophagy-related genes (Atg). LC3II is recruited to the autophagosome’s membrane assisting the elongation and closure; therefore, it is fundamental in the formation of autophagosome. Typically, cytoplasmic components selected for degradation are tagged by p62/SQSTM1 (an autophagy receptor) and recognized by LC3II, which leads them to the interior of the autophagosome.85 In the final step, the fusion of autophagosomes with endosomes and lysosomes occurs, which is followed by the degradation of the autophagosome content.83 Thus, analyses of beclin-1, LC3II and p62/SQSTM1 are commonly used as the markers of autophagy.
Autophagy has been demonstrated to have a role in osteoblasts based on several considerations. Firstly, autophagy regulates bone cell survival and apoptosis which is important for the balance between bone formation and bone resorption.86 Secondly, autophagy potentially alleviates oxidative stress, a key factor for the progression of osteoporosis.87 Thirdly, the autophagy modulators such as calcium, vitamin D, resveratrol, estradiol and bisphosphonates have been reported to be beneficial to bone health.88 Hence, autophagy in osteoblasts is implicated in the process of osteoblast differentiation and mineralization to maintain bone homeostasis. A study by Kim et al verified that kaempferol at concentrations up to 10 μM increased ALP activity and expression of osteoblast-activated factors (including Runx-2, OSX, BMP-2 and COL1).55 Along with the enhancement of proliferation, differentiation and mineralization of osteoblastic MC3T3-E1 cells, kaempferol induced autophagosome formation by increasing the expression of the autophagy-related factors, beclin-1, sequestosome-1 (SQSTM1/p62) and the conversion of LC3-II from LC3-I.55
Autophagy is also involved in modulating osteoclast formation and function. The increase in autophagy is associated with an increase in the viability of osteoclasts and bone resorption. Using a mouse model of systemic bone loss induced by glucocorticoids and ovariectomy, pharmacological and genetic inactivation of autophagy prevented bone loss by suppressing osteoclastogenesis and bone resorption in the animals.89 Mechanistically, it has been postulated that inhibition of autophagy lowers the secretion of cathepsin K thus disabling the differentiation and functional activity of osteoclasts.90 In a recent study, Kim et al revealed that kaempferol exhibited inhibitory effects on osteoclastogenesis (as shown by reduced TRAP-positive multinucleated cells, resorption pit formation, and expression of NFATc1, TRAF6 and c-Fos) as well as autophagy-related factors (as shown by decreased expression of beclin-1 and SQSTM1/p62) in RAW264.7 cells.35 In addition, the authors suggested that the inactivation of ERK and JNK was involved in the mechanism of action for kaempferol-inhibited autophagy in osteoclasts.35
It is evident that kaempferol exerts differential effects on autophagy in osteoblast and osteoclast. Kaempferol activates autophagy in the bone-forming osteoblasts but inhibits autophagy in the bone-resorbing osteoclasts. Both actions favor the maintenance of bone homeostasis and prevention of bone loss.
Effects Of Kaempferol In Apoptosis
Apoptosis is a process of cell death mediated by caspase and B-cell lymphoma 2 (Bcl-2) family proteins through two distinct mechanisms, the extrinsic and intrinsic signaling pathways. The extrinsic pathway is activated by the binding of extracellular ligands to cell surface death receptors, including the interactions between Fas ligand (FasL)/Fas receptor (FasR), TNF-α/tumor necrosis factor receptor 1 (TNFR1) and TNF-related apoptosis-inducing ligand (TRAIL)/TRAIL receptor (TRAIL-R). The engagement between these best characterized ligands and receptors leads to the formation of death-inducing signaling complex (DISC), recruitment of Fas-associated death domain (FADD) adaptor and activation of initiator caspase-8. Subsequently, executioner caspase-3 is activated causing the dismantling of cellular components.91 The intracellular stress signals such as oxidative stress, endoplasmic reticulum stress, DNA damage and cytosolic calcium overload trigger the intrinsic pathway. The presence of internal stimuli causes disruption of mitochondrial outer membrane permeabilization (MOMP), governed by increased pro-apoptotic genes [Bcl-2-associated X protein (Bax)] and decreased anti-apoptotic gene (Bcl-2), resulting in the release of cytochrome c. Cytochrome c binds to apoptosis protease activating factor-1 (Apaf-1) and deoxyadenosine triphosphate (dATP) to form apoptosome complex. The apoptosome further activates caspase-9 and downstream caspase-3/6/7 to trigger apoptosis.92
The anti-apoptotic effects of kaempferol have been previously explored in osteoblasts. Using dexamethasone-induced rat calvarial osteoblasts, kaempferol caused a reduction in osteoblast apoptosis indicated by the stimulation of Bcl-2 expression and suppression of Bax expression. In this study, the postulated mechanism of action for the anti-apoptotic effects of kaempferol was elicited through the ERK signaling pathway. Western analysis results indicated that kaempferol amplified the phosphorylation of ERK.24
Effects Of Kaempferol On Other Proteins
There are several other proteins that have been shown to implicate in the modulation of bone metabolism. Cytokeratin-14 or keratin-14 belongs to the cytoplasmic intermediate filament family and functions to self-assemble forming large bundles. Keratins are a group of tough and fibrous proteins providing structural framework of strength and resiliency from mechanical and non-mechanical stress.93 HSP70 is a stress-inducible protein released in response to cellular stressors, such as heat shock, inflammation, fractures and infections.94 The osteogenic effects of HSP70 are accomplished through the enhancement of ALP activity, calcium deposition and upregulation of osteo-specific genes (Runx-2 and OSX) by activating the ERK signaling pathway.95,96 Aldose reductase is a key enzyme that catalyzes the conversion of glucose to sorbitol. A negative correlation between AR level and bone health has been reported. Galactose-induced diabetic rats had enhanced bone resorption and lower bone volume and osteoblast numbers, which were restored by the administration of epalrestat (an aldose reductase inhibitor).97 Caldesmon is known to participate in the regulation of actin cytoskeletal remodeling and alteration of cell surface adherence force, thereby facilitating the fusion of osteoclasts into multinucleated osteoclasts during osteoclastogenesis.98 Therefore, high levels of cytokeratin-14 and HSP-70 as well as low levels of AR and caldesmon might have a role in promoting osteoblast differentiation and inhibiting osteoclast differentiation.
In the presence of kaempferol, the rat calvarial osteoblasts had cuboidal morphology, developed intercellular networks, high ALP activity and increased mineralized nodules. Proteomic data revealed elevation of cytokeratin-14 and HSP70 but reduction of aldose reductase and caldesmon in rat calvarial osteoblasts treated with kaempferol.99 Another recent study further confirmed the role of cytokeratin-14 in osteoblast differentiation. The findings showed a positive relationship between cytokeratin-14 and matrix mineralization, which was corroborated by increased expression of cytokeratin-14, COL1 level, OCN level and collagen fiber density. It was surmised that the augmentation of cytokeratin-14 was involved in the regulation of osteoblast mineralization through reduction of phosphorylated adenosine monophosphate-activated protein kinase (AMPK) to activate the mammalian target of rapamycin complex 1 (mTORC1).100
Conclusion And Future Directions
Various doses of kaempferol have been tested in previous preclinical studies. Kaempferol may act as osteogenic30,49,56 and anti-osteoclastogenic32,71,81 agent by acting on both osteoblasts and osteoclasts in a dose-dependent manner. In vivo studies also showed that higher dose of kaempferol increased callus diameter in the fractured mice, but this effect was not seen in the group treated with lower dose of kaempferol.25 Thus, it is postulated that higher dose of kaempferol conferred better protection on bone. However, the safety profile of kaempferol remains a major concern as researchers revealed signs of mutagenicity in the Ames test using Salmonella typhimurium.101 Based on the documented evidence, it is hypothesized that the possible underlying mechanisms for the anti-osteoporotic effects of kaempferol are in part mediated through (a) suppression of adipogenesis thus favoring osteoblastogenesis and chondrogenesis, (b) activation of estrogen receptor signaling pathway, (c) increase in Runx-2 expression that acts as the master of osteogenic transcription factor in BMP-2 signaling, (d) inhibition of inflammatory response through inhibition of NF-κB, (e) reduction of intracellular ROS production and lipid peroxidation, (f) differential regulation of autophagy in osteoblast and osteoclast, (g) suppression of osteoblast apoptosis as well as (h) regulation of proteins involved in osteoblast mineralization. All these molecular mechanisms play a relevant role in the maintenance of tightly coupled bone remodeling processes. The summary of the mechanisms of action that underlie the osteogenic property of kaempferol is illustrated in Figure 1.
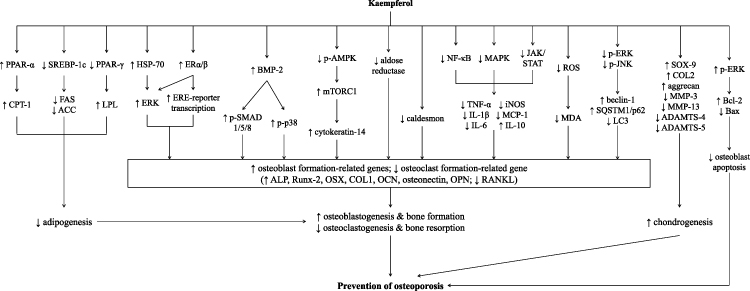 |
Figure 1 Summary of the mechanism of action involved in the osteoprotective effects of kaempferol. Abbreviations: ACC, acetyl coenzyme A carboxylase; ADAMTS, a disintegrin and metalloproteinase with thrombospondin motifs; ALP, alkaline phosphatase; AMPK, adenosine monophosphate-activated protein kinase; Bax, bcl-2-like protein 4; Bcl2, B-cell lymphoma 2; BMP-2, bone morphogenetic protein-2; COL1, collagen type I; COL2, collagen type 2; COX-2, cyclooxygenase-2; CPT-1, carnitine palmitoyl transferase-1; ERα, estrogen receptor-alpha; ERβ, estrogen receptor-beta; ERE, estrogen responsive elements; ERK, extracellular-regulated kinase; FAS, fatty acid synthase; HSP70, heat shock protein 70; IL, interleukin; iNOS, inducible nitric oxide synthase; JAK, Janus kinase; JNK, c-Jun N-terminal kinase; LPL, lipoprotein lipase; MAPK, mitogen-activated protein kinase; MCP-1, monocyte chemoattractant protein; MDA, malondialdehyde; MMP, matrix metalloproteinase; mTORC1, mammalian target of rapamycin complex 1; NF-κB, nuclear factor-kappa B; OCN, osteocalcin; OPN, osteopontin; OSX, osterix; PPAR-α, peroxisome proliferator-activated receptor-alpha; PPAR-γ, peroxisome proliferator-activated receptor-gamma; RANKL, receptor activator of nuclear factor kappa-Β ligand; ROS, reactive oxygen species; Runx-2, Runt-related transcription factor 2; SMAD, suppressor of mothers against decapentaplegic; SOX-9, SRY-Box 9; SQSTM1, sequestosome-1; SREBP-1c, sterol regulatory element-binding proteins-1c; STAT, signal transducer and activator of transcription; TNF-α, tumor necrosis factor-alpha. |
Several limitations need to be addressed. Firstly, research on the effects of kaempferol on bone in humans is scarce but the results from in vivo and in vitro researches seem promising. Secondly, there is only a paucity of data indicating the anti-oxidative capacity of kaempferol using osteoblastic and osteoclastic cells. Thirdly, investigation on the effect of kaempferol on the antioxidant system is limited; particularly the levels of superoxide dismutase (SOD), glutathione peroxidase (GPx), catalase (CAT) and glutathione (GSH) were not evaluated. Fourthly, there is also a lack of scientific studies on the apoptotic activity of kaempferol in bone cells. By affecting both cell types (osteoblasts and osteoclasts) as well as bone cell communication, a multitude of redundant pathways (such as estrogen, BMP-2, MAPK and mTOR signaling mechanisms) and regulators (including cytokines and ROS) seem to be of major importance in bone cell function. This review reiterates that the design of potential agents targeting these major signaling cascades should be explored as the treatment for bone diseases. The growing evidence on the skeletal-protecting effects of kaempferol supports its development as a potent adjunct for maintaining bone mass, skeletal integrity and preventing fractures. More importantly, the studies included in this review were performed in the in vivo and in vitro models. Further studies are warranted particularly in validating the clinical uses of kaempferol.
Acknowledgment
We thank Universiti Kebangsaan Malaysia for supporting this work via the MI-2019-006 and the FF-2019-036/1 grants.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Duque G, Rivas D, Li W, et al. Age-related bone loss in the LOU/c rat model of healthy ageing. Exp Gerontol. 2009;44:183–189. doi:10.1016/j.exger.2008.10.004
2. Chin KY, Ima-Nirwana S. The effects of orchidectomy and supraphysiological testosterone administration on trabecular bone structure and gene expression in rats. Aging Male. 2015;18:60–66. doi:10.3109/13685538.2014.954995
3. Chen Z, Xue J, Shen T, Mu S, Fu Q. Curcumin alleviates glucocorticoid-induced osteoporosis through the regulation of the Wnt signaling pathway. Int J Mol Med. 2016;37:329–338. doi:10.3892/ijmm.2015.2432
4. Mohamad N-V, Che Zulkepli MAA, May Theseira K, et al. Establishing an animal model of secondary osteoporosis by using a gonadotropin-releasing hormone agonist. Int J Med Sci. 2018;15:300–308. doi:10.7150/ijms.22732
5. Broulik PD, Vondrova J, Ruzicka P, et al. The effect of chronic alcohol administration on bone mineral content and bone strength in male rats. Physiol Res. 2010;59:599–604.
6. Hapidin H, Othman F, Soelaiman IN, Shuid AN, Mohamed N. Effects of nicotine administration and nicotine cessation on bone histomorphometry and bone biomarkers in Sprague-Dawley male rats. Calcif Tissue Int. 2011;88:41–47. doi:10.1007/s00223-010-9426-4
7. Wong SK, Chin KY, Suhaimi FH, et al. The relationship between metabolic syndrome and osteoporosis: a review. Nutrients. 2016;8:347. doi:10.3390/nu8060347
8. Tu KN, Lie JD, Wan CKV, et al. Osteoporosis: a review of treatment options. P & T. 2018;43:92–104.
9. Kling JM, Clarke BL, Sandhu NP. Osteoporosis prevention, screening, and treatment: a review. J Womens Health. 2014;23:563–572. doi:10.1089/jwh.2013.4611
10. Wong SK, Mohamad NV, Ibrahim N, et al. The molecular mechanism of vitamin E as a bone-protecting agent: a review on current evidence. Int J Mol Sci. 2019;20. doi:10.3390/ijms20061453
11. Wong SK, Chin KY, Ima-Nirwana S. Berberine and musculoskeletal disorders: the therapeutic potential and underlying molecular mechanisms. Phytomedicine. 2019;152892. doi:10.1016/j.phymed.2019.152892
12. Chin KY, Ima-Nirwana S. Olives and bone: a green osteoporosis prevention option. Int J Environ Res Public Health. 2016;13. doi:10.3390/ijerph13121252
13. Chin K-Y, Ima-Nirwana S. The biological effects of tocotrienol on bone: a review on evidence from rodent models. Drug Des Devel Ther. 2015;9:2049–2061. doi:10.2147/DDDT.S79660
14. Wong SK, Chin KY, Suhaimi FH, Ahmad F, Ima-Nirwana S. Exploring the potential of tocotrienol from Bixa orellana as a single agent targeting metabolic syndrome and bone loss. Bone. 2018;116:8–21. doi:10.1016/j.bone.2018.07.003
15. Wong SK, Chin KY, Suhaimi FH, et al. The effects of vitamin E from elaeis guineensis (oil palm) in a rat model of bone loss due to metabolic syndrome. Int J Environ Res Public Health. 2018;15:1828.
16. Wong SK, Chin KY, Suhaimi FH, Ahmad F, Ima-Nirwana S. The effects of palm tocotrienol on metabolic syndrome and bone loss in male rats induced by high-carbohydrate high-fat diet. J Funct Foods. 2018;44:246–254. doi:10.1016/j.jff.2018.03.022
17. Wong SK, Chin K-Y, Ima-Nirwana S. The effects of tocotrienol on bone peptides in a rat model of osteoporosis induced by metabolic syndrome: the possible communication between bone cells. Int J Environ Res Public Health. 2019;16:3313. doi:10.3390/ijerph16183313
18. Calderon-Montano JM, Burgos-Moron E, Perez-Guerrero C, et al. A review on the dietary flavonoid kaempferol. Mini Rev Med Chem. 2011;11:298–344.
19. Imran M, Rauf A, Shah ZA, et al. Chemo-preventive and therapeutic effect of the dietary flavonoid kaempferol: A comprehensive review. Phytother Res. 2019;33:263–275. doi:10.1002/ptr.6227
20. Yang L, Takai H, Utsunomiya T, et al. Kaempferol stimulates bone sialoprotein gene transcription and new bone formation. J Cell Biochem. 2010;110:1342–1355. doi:10.1002/jcb.22649
21. Trivedi R, Kumar S, Kumar A, et al. Kaempferol has osteogenic effect in ovariectomized adult Sprague-Dawley rats. Mol Cell Endocrinol. 2008;289:85–93. doi:10.1016/j.mce.2008.02.027
22. Nowak B, Matuszewska A, Nikodem A, et al. Oral administration of kaempferol inhibits bone loss in rat model of ovariectomy-induced osteopenia. Pharmacol Rep. 2017;69:1113–1119. doi:10.1016/j.pharep.2017.05.002
23. Ma X-Q, Han T, Zhang X, et al. Kaempferitrin prevents bone lost in ovariectomized rats. Phytomedicine. 2015;22:1159–1162. doi:10.1016/j.phymed.2015.09.003
24. Adhikary S, Choudhary D, Ahmad N, et al. Dietary flavonoid kaempferol inhibits glucocorticoid-induced bone loss by promoting osteoblast survival. Nutrition. 2018;53:64–76. doi:10.1016/j.nut.2017.12.003
25. Nguyen LT, Maidar O, Sharma AR, et al. Enhanced healing process of fracture by treatment of Kaempferol. Biomed Res. 2016;27.
26. Kim H, Han C, Maidar O, et al. Biomechanical effects of kaempferol treatments on the bone healing process of murine tibia.
27. Gupta G, Kumar A, Khedgikar V, et al. Osteogenic efficacy enhancement of kaempferol through an engineered layer-by-layer matrix: a study in ovariectomized rats. Nanomedicine. 2013;8:757–771. doi:10.2217/nnm.12.171
28. Kumar A, Gupta G, Khedgikar V, et al. In vivo efficacy studies of layer-by-layer nano-matrix bearing kaempferol for the conditions of osteoporosis: a study in ovariectomized rat model. Eur J Pharm Biopharm. 2012;82:508–517. doi:10.1016/j.ejpb.2012.08.001
29. Tsuchiya S, Sugimoto K, Kamio H, et al. Kaempferol-immobilized titanium dioxide promotes formation of new bone: effects of loading methods on bone marrow stromal cell differentiation in vivo and in vitro. Int J Nanomedicine. 2018;13:1665–1676. doi:10.2147/IJN.S150786
30. Zhu J, Tang H, Zhang Z, et al. Kaempferol slows intervertebral disc degeneration by modifying LPS-induced osteogenesis/adipogenesis imbalance and inflammation response in BMSCs. Int Immunopharmacol. 2017;43:236–242. doi:10.1016/j.intimp.2016.12.020
31. Miyake M, Arai N, Ushio S, Iwaki K, Ikeda M, Kurimoto M. Promoting effect of kaempferol on the differentiation and mineralization of murine pre-osteoblastic cell line MC3T3-E1. Biosci Biotechnol Biochem. 2003;67:1199–1205. doi:10.1271/bbb.67.1199
32. Yamaguchi M, Hamamoto R, Uchiyama S, Ishiyama K. Effects of flavonoid on calcium content in femoral tissue culture and parathyroid hormone-stimulated osteoclastogenesis in bone marrow culture in vitro. Mol Cell Biochem. 2007;303:83–88. doi:10.1007/s11010-007-9458-x
33. Marino S, Logan JG, Mellis D, Capulli M. Generation and culture of osteoclasts. Bonekey Rep. 2014;3:570. doi:10.1038/bonekey.2014.65
34. Pang J, Ricupero D, Huang S, et al. Differential activity of kaempferol and quercetin in attenuating tumor necrosis factor receptor family signaling in bone cells. Biochem Pharmacol. 2006;71:818–826. doi:10.1016/j.bcp.2005.12.023
35. Kim CJ, Shin SH, Kim BJ, et al. The effects of kaempferol-inhibited autophagy on osteoclast formation. Int J Mol Sci. 2018;19:125.
36. Shahi M, Peymani A, Sahmani M. Regulation of bone metabolism. Rep Biochem Mol Biol. 2017;5:73–82.
37. Krane SM, Inada M. Matrix metalloproteinases and bone. Bone. 2008;43:7–18. doi:10.1016/j.bone.2008.03.020
38. Paiva KB, Granjeiro JM. Bone tissue remodeling and development: focus on matrix metalloproteinase functions. Arch Biochem Biophys. 2014;561:74–87. doi:10.1016/j.abb.2014.07.034
39. Almalki SG, Agrawal DK. Effects of matrix metalloproteinases on the fate of mesenchymal stem cells. Stem Cell Res Ther. 2016;7:129. doi:10.1186/s13287-016-0393-1
40. Kelwick R, Desanlis I, Wheeler GN, et al. The ADAMTS (A Disintegrin and Metalloproteinase with Thrombospondin motifs) family. Genome Biol. 2015;16:113. doi:10.1186/s13059-015-0667-4
41. Wong SK, Chin KY, Suhaimi FH, Ahmad F, Jamil NA, Ima-Nirwana S. Osteoporosis is associated with metabolic syndrome induced by high-carbohydrate high-fat diet in a rat model. Biomed Pharmacother. 2018;98:191–200. doi:10.1016/j.biopha.2017.12.042
42. Wong SK, Chin KY, Suhaimi FH, Ahmad F, Ima-Nirwana S, Bader M. Effects of metabolic syndrome on bone mineral density, histomorphometry and remodelling markers in male rats. PLoS One. 2018;13:e0192416. doi:10.1371/journal.pone.0192416
43. Valenzuela R, Videla LA. The importance of the long-chain polyunsaturated fatty acid n-6/n-3 ratio in development of non-alcoholic fatty liver associated with obesity. Food Funct. 2011;2:644–648. doi:10.1039/c1fo10133a
44. Byun MR, Jeong H, Bae SJ, Kim AR, Hwang ES, Hong J-H. TAZ is required for the osteogenic and anti-adipogenic activities of kaempferol. Bone. 2012;50:364–372. doi:10.1016/j.bone.2011.10.035
45. Weitzmann MN, Pacifici R. Estrogen deficiency and bone loss: an inflammatory tale. J Clin Invest. 2006;116:1186–1194. doi:10.1172/JCI28550
46. Lee H-R, Kim T-H, Choi K-C. Functions and physiological roles of two types of estrogen receptors, ERα and ERβ, identified by estrogen receptor knockout mouse. Lab Anim Res. 2012;28:71–76. doi:10.5625/lar.2012.28.2.71
47. Gupte AA, Pownall HJ, Hamilton DJ. Estrogen: an emerging regulator of insulin action and mitochondrial function. J Diabetes Res. 2015;2015:9. doi:10.1155/2015/815839
48. Khalid AB, Krum SA. Estrogen receptors alpha and beta in bone. Bone. 2016;87:130–135. doi:10.1016/j.bone.2016.03.016
49. Prouillet C, Maziere JC, Maziere C, et al. Stimulatory effect of naturally occurring flavonols quercetin and kaempferol on alkaline phosphatase activity in MG-63 human osteoblasts through ERK and estrogen receptor pathway. Biochem Pharmacol. 2004;67:1307–1313. doi:10.1016/j.bcp.2003.11.009
50. Guo AJ, Choi RC, Zheng KY, et al. Kaempferol as a flavonoid induces osteoblastic differentiation via estrogen receptor signaling. Chin Med. 2012;7:10. doi:10.1186/1749-8546-7-10
51. Tang X, Zhu X, Liu S, Nicholson RC, Ni X. Phytoestrogens induce differential estrogen receptor beta-mediated responses in transfected MG-63 cells. Endocrine. 2008;34:29–35. doi:10.1007/s12020-008-9099-1
52. Yang L, Chen Q, Wang F, Zhang G. Antiosteoporotic compounds from seeds of Cuscuta chinensis. J Ethnopharmacol. 2011;135:553–560. doi:10.1016/j.jep.2011.03.056
53. Wu M, Chen G, Li YP. TGF-beta and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016;4:16009. doi:10.1038/boneres.2016.9
54. Redlich K, Smolen JS. Inflammatory bone loss: pathogenesis and therapeutic intervention. Nat Rev Drug Discov. 2012;11:234. doi:10.1038/nrd3669
55. Kim IR, Kim SE, Baek HS, et al. The role of kaempferol-induced autophagy on differentiation and mineralization of osteoblastic MC3T3-E1 cells. BMC Complement Altern Med. 2016;16:333. doi:10.1186/s12906-016-1320-9
56. Chiou W-F, Lee C-H, Liao J-F, Chen CC. 8-Prenylkaempferol accelerates osteoblast maturation through bone morphogenetic protein-2/p38 pathway to activate Runx2 transcription. Life Sci. 2011;88:335–342. doi:10.1016/j.lfs.2010.12.009
57. Long M, Li SX, Xiao JF, et al. Kidney tubular-cell secretion of osteoblast growth factor is increased by kaempferol: a scientific basis for “the kidney controlling the bone” theory of Chinese medicine. Chin J Integr Med. 2014;20:675–681. doi:10.1007/s11655-014-1336-z
58. Lacey DC, Simmons PJ, Graves SE, Hamilton JA. Proinflammatory cytokines inhibit osteogenic differentiation from stem cells: implications for bone repair during inflammation. Osteoarthritis Cartilage. 2009;17:735–742. doi:10.1016/j.joca.2008.11.011
59. Seong S, Kim JH, Kim N. Pro-inflammatory cytokines modulating osteoclast differentiation and function. J Rheum Dis. 2016;23:148–153. doi:10.4078/jrd.2016.23.3.148
60. Confalone E, D’Alessio G, Furia A. IL-6 induction by TNFα and IL-1β in an osteoblast-like cell line. Int J Biomed Sci. 2010;6:135–140.
61. Zhu JF, Valente AJ, Lorenzo JA, Carnes D, Graves DT. Expression of monocyte chemoattractant protein 1 in human osteoblastic cells stimulated by proinflammatory mediators. J Bone Miner Res. 1994;9:1123–1130. doi:10.1002/jbmr.5650090721
62. Pietschmann P, Mechtcheriakova D, Meshcheryakova A, Föger-Samwald U, Ellinger I. Immunology of osteoporosis: a mini-review. Gerontology. 2016;62:128–137. doi:10.1159/000431091
63. Khosla S. Minireview: the OPG/RANKL/RANK system. Endocrinology. 2001;142:5050–5055. doi:10.1210/endo.142.12.8536
64. Hardy R, Cooper MS. Bone loss in inflammatory disorders. J Endocrinol. 2009;201:309–320. doi:10.1677/JOE-08-0568
65. Boyce BF, Xing L. Biology of RANK, RANKL, and osteoprotegerin. Arthritis Res Ther. 2007;9:S1. doi:10.1186/ar2172
66. Boyce BF, Xing L. The RANKL/RANK/OPG pathway. Curr Osteoporos Rep. 2007;5:98–104.
67. Wimalawansa SJ. Nitric oxide and bone. Ann N Y Acad Sci. 2010;1192:391–403. doi:10.1111/j.1749-6632.2009.05230.x
68. Förstermann U, Sessa WC. Nitric oxide synthases: regulation and function. Eur Heart J. 2012;33:829d–837d. doi:10.1093/eurheartj/ehr304
69. Zheng H, Yu X, Collin-Osdoby P, et al. RANKL stimulates inducible nitric-oxide synthase expression and nitric oxide production in developing osteoclasts. An autocrine negative feedback mechanism triggered by RANKL-induced interferon-beta via NF-kappaB that restrains osteoclastogenesis and bone resorption. J Biol Chem. 2006;281:15809–15820. doi:10.1074/jbc.M513225200
70. Cuzzocrea S, Mazzon E, Dugo L, et al. Inducible nitric oxide synthase mediates bone loss in ovariectomized mice. Endocrinology. 2003;144:1098–1107. doi:10.1210/en.2002-220597
71. Lee WS, Lee EG, Sung MS, et al. Kaempferol inhibits IL-1beta-stimulated, RANKL-mediated osteoclastogenesis via downregulation of MAPKs, c-Fos, and NFATc1. Inflammation. 2014;37:1221–1230. doi:10.1007/s10753-014-9849-6
72. Yu Q, Zeng K, Ma X, et al. Resokaempferol-mediated anti-inflammatory effects on activated macrophages via the inhibition of JAK2/STAT3, NF-kappaB and JNK/p38 MAPK signaling pathways. Int Immunopharmacol. 2016;38:104–114. doi:10.1016/j.intimp.2016.05.010
73. Lee SB, Shin JS, Han HS, et al. Kaempferol 7-O-beta-D-glucoside isolated from the leaves of Cudrania tricuspidata inhibits LPS-induced expression of pro-inflammatory mediators through inactivation of NF-kappaB, AP-1, and JAK-STAT in RAW 264.7 macrophages. Chem Biol Interact. 2018;284:101–111. doi:10.1016/j.cbi.2018.02.022
74. Domazetovic V, Marcucci G, Iantomasi T, Brandi ML, Vincenzini MT. Oxidative stress in bone remodeling: role of antioxidants. Clin Cases Miner Bone Metab. 2017;14:209–216. doi:10.11138/ccmbm/2017.14.1.209
75. Liu AL, Zhang ZM, Zhu BF, Liao Z-H, Liu Z. Metallothionein protects bone marrow stromal cells against hydrogen peroxide-induced inhibition of osteoblastic differentiation. Cell Biol Int. 2004;28:905–911. doi:10.1016/j.cellbi.2004.09.004
76. Arai M, Shibata Y, Pugdee K, Abiko Y, Ogata Y. Effects of reactive oxygen species (ROS) on antioxidant system and osteoblastic differentiation in MC3T3-E1 cells. IUBMB Life. 2007;59:27–33. doi:10.1080/15216540601156188
77. Wauquier F, Leotoing L, Coxam V, Guicheux J, Wittrant Y. Oxidative stress in bone remodelling and disease. Trends Mol Med. 2009;15:468–477. doi:10.1016/j.molmed.2009.08.004
78. Deng S, Dai G, Chen S, et al. Dexamethasone induces osteoblast apoptosis through ROS-PI3K/AKT/GSK3β signaling pathway. Biomed Pharmacother. 2019;110:602–608. doi:10.1016/j.biopha.2018.11.103
79. Suh KS, Choi EM, Kwon M, et al. Kaempferol attenuates 2-deoxy-d-ribose-induced oxidative cell damage in MC3T3-E1 osteoblastic cells. Biol Pharm Bull. 2009;32:746–749. doi:10.1248/bpb.32.746
80. Almeida M, Han L, Martin-Millan M, et al. Skeletal involution by age-associated oxidative stress and its acceleration by loss of sex steroids. J Biol Chem. 2007;282:27285–27297. doi:10.1074/jbc.M702810200
81. Wattel A, Kamel S, Mentaverri R, et al. Potent inhibitory effect of naturally occurring flavonoids quercetin and kaempferol on in vitro osteoclastic bone resorption. Biochem Pharmacol. 2003;65:35–42. doi:10.1016/s0006-2952(02)01445-4
82. Hocking LJ, Whitehouse C, Helfrich MH. Autophagy: a new player in skeletal maintenance? J Bone Miner Res. 2012;27:1439–1447. doi:10.1002/jbmr.1668
83. Florencio-Silva R, Sasso GR, Simoes MJ, et al. Osteoporosis and autophagy: what is the relationship? Rev Assoc Med Bras. 2017;63:173–179. doi:10.1590/1806-9282.63.02.173
84. Fu LL, Cheng Y, Liu B. Beclin-1: autophagic regulator and therapeutic target in cancer. Int J Biochem Cell Biol. 2013;45:921–924. doi:10.1016/j.biocel.2013.02.007
85. Lamark T, Svenning S, Johansen T. Regulation of selective autophagy: the p62/SQSTM1 paradigm. Essays Biochem. 2017;61:609–624. doi:10.1042/EBC20170035
86. Nollet M, Santucci-Darmanin S, Breuil V, et al. Autophagy in osteoblasts is involved in mineralization and bone homeostasis. Autophagy. 2014;10:1965–1977. doi:10.4161/auto.36182
87. Dutta D, Xu J, Kim JS, Dunn WA, Leeuwenburgh C. Upregulated autophagy protects cardiomyocytes from oxidative stress-induced toxicity. Autophagy. 2013;9:328–344. doi:10.4161/auto.22971
88. Pierrefite-Carle V, Santucci-Darmanin S, Breuil V, Camuzard O, Carle GF. Autophagy in bone: self-eating to stay in balance. Ageing Res Rev. 2015;24:206–217. doi:10.1016/j.arr.2015.08.004
89. Lin NY, Chen CW, Kagwiria R, et al. Inactivation of autophagy ameliorates glucocorticoid-induced and ovariectomy-induced bone loss. Ann Rheum Dis. 2016;75:1203–1210. doi:10.1136/annrheumdis-2015-207240
90. Dawodu D, Patecki M, Hegermann J, Dumler I, Haller H, Kiyan Y. oxLDL inhibits differentiation and functional activity of osteoclasts via scavenger receptor-a mediated autophagy and cathepsin K secretion. Sci Rep. 2018;8:11604. doi:10.1038/s41598-018-29963-w
91. Charlier E, Relic B, Deroyer C, et al. Insights on molecular mechanisms of chondrocytes death in osteoarthritis. Int J Mol Sci. 2016;17:2146. doi:10.3390/ijms17122146
92. Singh R, Letai A, Sarosiek K. Regulation of apoptosis in health and disease: the balancing act of BCL-2 family proteins. Nat Rev Mol Cell Biol. 2019;20:175–193. doi:10.1038/s41580-018-0089-8
93. Nolting J-F, Möbius W, Köster S. Mechanics of individual keratin bundles in living cells. Biophys J. 2014;107:2693–2699. doi:10.1016/j.bpj.2014.10.039
94. Goldstein MG, Li Z. Heat-shock proteins in infection-mediated inflammation-induced tumorigenesis. J Hematol Oncol. 2009;2:5. doi:10.1186/1756-8722-2-5
95. Chen E, Xue D, Zhang W, Lin F, Pan Z. Extracellular heat shock protein 70 promotes osteogenesis of human mesenchymal stem cells through activation of the ERK signaling pathway. FEBS Lett. 2015;589:4088–4096. doi:10.1016/j.febslet.2015.11.021
96. Li C, Sunderic K, Nicoll SB, Wang S. Downregulation of heat shock protein 70 impairs osteogenic and chondrogenic differentiation in human mesenchymal stem cells. Sci Rep. 2018;8:553. doi:10.1038/s41598-017-18541-1
97. Inaba M, Terada M, Nishizawa Y, et al. Protective effect of an aldose reductase inhibitor against bone loss in galactose-fed rats: possible involvement of the polyol pathway in bone metabolism. Metabolism. 1999;48:904–909. doi:10.1016/s0026-0495(99)90227-5
98. Chan CL, Chen JY, Shih MC, et al. L-caldesmon alters cell spreading and adhesion force in RANKL-induced osteoclasts. J Biomed Sci. 2019;26:12. doi:10.1186/s12929-019-0555-4
99. Kumar A, Singh AK, Gautam AK, et al. Identification of kaempferol-regulated proteins in rat calvarial osteoblasts during mineralization by proteomics. Proteomics. 2010;10:1730–1739. doi:10.1002/pmic.200900666
100. Khedgikar V, Kushwaha P, Gautam J, et al. Kaempferol targets Krt-14 and induces cytoskeletal mineralization in osteoblasts: A mechanistic approach. Life Sci. 2016;151:207–217. doi:10.1016/j.lfs.2016.03.009
101. Resende FA, Vilegas W, Dos Santos LC, Varanda EA. Mutagenicity of flavonoids assayed by bacterial reverse mutation (Ames) test. Molecules. 2012;17:5255–5268. doi:10.3390/molecules17055255
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.