")
Back to Journals » Cancer Management and Research » Volume 11
The opposite role of alternatively spliced isoforms of LINC00477 in gastric cancer
Authors Zhao H, He Y, Li H, Zhu A, Ye Y, Liu G, Zhao C, Zhang X
Received 21 January 2019
Accepted for publication 9 April 2019
Published 16 May 2019 Volume 2019:11 Pages 4569—4576
DOI https://doi.org/10.2147/CMAR.S202430
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Beicheng Sun
Hongchao Zhao,* Yuanhang He,* Haohao Li, Ali Zhu, Yanwei Ye, Guanghui Liu, Chunlin Zhao, Xiefu Zhang
Department of Gastroenterology, The First Affiliated Hospital of Zhengzhou University, Zhengzhou, People’s Republic of China
*These authors contributed equally to this work
Background: Aberrant transcript alternative splicing is an important regulatory process closely connected with oncogenesis.
Purpose: The objective of this study was to determine the phenotype and function of a novel long noncoding RNA (lncRNA) LINC00477 in gastric cancer.
Patients and methods: The gastric cancer samples of 140 from Oncomine database and 17 from our own hospital, as well as three gastric cancer cell lines MKN-45, AGS and KATO III were used in this study. The expression of the spliced isoforms of LINC00477 were tested. The tumor effects of LINC00477 on gastric cancer were investigated in vitro and in vivo. The mechanism of LINC00477 interacted with aconitase 1 (ACO1) was further examined by RIP and pull down assay.
Results: The overall expression of LINC00477 was reduced in gastric cancers compared to normal gastric tissues. The isoform 1 of LINC00477 was down-regulated while the isoform 2 was up-regulated in gastric cancer cells. The opposite role of isoforms 1 and 2 in the proliferation and migration of cancer cells in vitro and in vivo was observed. Furthermore, isoform 1 of LINC00477 was determined to interact with ACO1 and suppress the conversion ability from citrate to isocitrate by ACO1.
Conclusion: we presented the important roles of the spliced isoforms of long noncoding RNA, LINC00477 in gastric carcinogenesis.
Keywords: LINC00477, gastric cancer, lncRNA, ACO1
Introduction
Gastric cancer (GC) is one of the most common gastrointestinal malignancy, ranking the second leading cause of cancer-related death worldwide.1 The five-year survival rate is only 20%.2 Most of the cases are diagnosed at a terminal stage, which is accompanied by malignant multiplication and extensive invasion in lymph node or distant metastasis.3,4 Even though the improved chemotherapy protocols reduced the five-year mortality rate, it is still an urgent need to clarify the metastatic mechanisms and identify new prognostic biomarkers or potential therapeutic target for GC.
Long non-coding RNAs (lncRNAs) are a series of non-protein coding transcripts in excess of 200 nt, which have been determined to play crucial roles in multiple biological processes, such as transcriptional regulation, alternative splicing, chromatin remodeling, X-chromosome imprinting and cell differentiation, as well as metastasis and drug resistance in cancer.5–7 Alternative splicing is a posttranscriptional regulation via generating multiple spliced isoforms with a tissue- and cell-specific manner to increase the diversity of the transcriptome. Alternative splicing of lncRNAs further expands their regulatory and functional complexity in cancerogenesis and cancer development. Therefore, illustration of the differential profiles of lncRNA variants in GC can be beneficial to identify gastric cancer-specific biomarkers, provide the potential therapeutic targets, and figure out the underlying mechanisms of lncRNAs in stomach tumorigenesis.
In this study, we investigated the lncRNA profiles in GC patients of oncomine database and found that LINC00477, a novel lncRNA with no detailed research currently, was downregulated in GC cancers compared to adjacent tissues. Interestingly, we further found that LINC00477 had two spliced isoforms, whose transcriptional levels cannot be reflected from oncomine data. Therefore, we focused on LINC00477 and explored the roles of different variants played in GC.
Materials and methods
Samples, cells, vectors, RNA oligos, and antibodies
Tissues including 7 gastric mucosal epithelium from ulcer patients, 5 squamous carcinoma, and 5 adenocarcinoma of stomach and their corresponding para-carcinoma tissues were harvested from Department of Gastroenterology in the First Affiliated Hospital of Zhengzhou University. The clinical information of the patients are listed in Table 1. Signed informed consent and ethics committee documents of Ethics Committee of The First Affiliated Hospital of Zhengzhou University were all provided to approve this study. Gastric cancer cell lines MKN-45, AGS and KATO III and gastric epithelial cells GES-1 were purchased from American Type Culture Collection (ATCC). LINC00477 isoform 1 and 2 abbreviated as L1 and L2 were cloned the full length from GES-1 cells and generated into pcDNA3.1. The RNAi oligos of L1 and L2 were designed from (Thermo, USA) and inserted into pLKO.1-GFP vector. All the sequence of primers and oligos are listed in Table 2. The lncRNA stably expressing or blocking cell lines were prepared via vectors transfection and screening using 600 μg/mL G418 or 1.5 μg/mL puromycin. The second and third exons of LINC00477 were synthetic with biotin labeled (Thermo Fisher Scientific, USA) respectively, for RNA pull down assay. GAPDH and streptavidin primary antibodies were obtained from CST. IP grade Aconitase 1 (ACO1) primary antibody was obtained from Abcam.
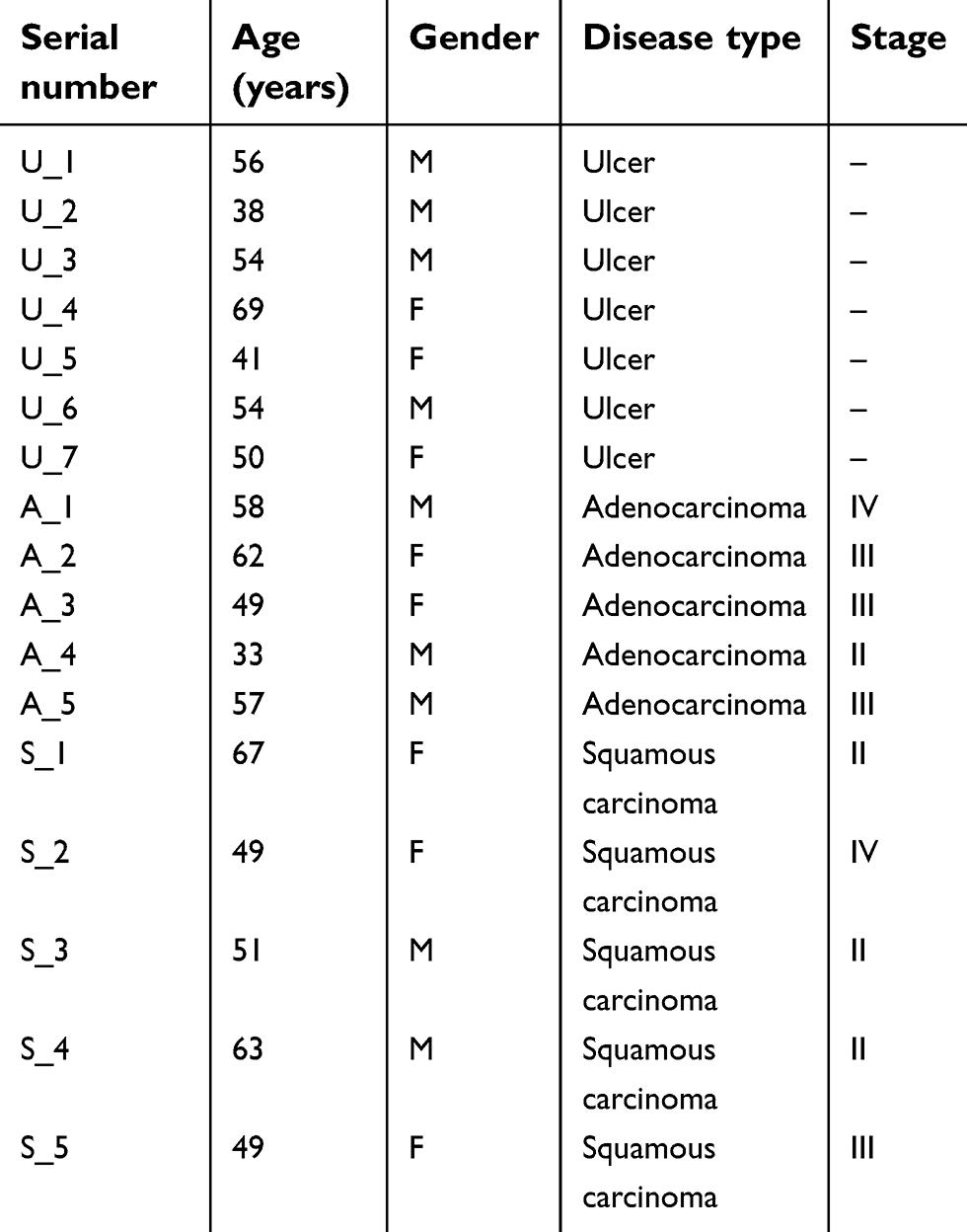 | Table 1 The clinical information of the patients used in this study |
 | Table 2 All the primers and RNA oligos used in this study were listed |
RNA extraction
Freshly prepared cells or tissues were washed by PBS and added into 1 mL RNAiso plus (Takara, Japan). Tissues needed to be ground within liquid nitrogen. Samples were added 200 μL chloroform, vortex to mix and centrifuged in highest speed at 4°C. The supernatant was transferred to new tubes, added isopropanol with the same volume, and centrifuged in highest speed at 4°C, then washed by cold ethanol and 75% ethanol, and dissolved in DEPC water. A total of 100 ng RNA measured by Nanodrop was conducted reverse transcription using PrimeScriptTM RT-PCR Kit (Takara, Japan). Real-time PCR was performed using FastStart Universal SYBR Green Master (Roche, USA). All primers used in this study are listed in Table 2.
CCK-8 assay
A total of 1×104 cells within 100 µL volume were transplanted into 96 well plates, changed the medium with 10% CCK8 (Solarbio, China) for culture 4 hrs after cell adherence, and examined the absorption within the wavelength of 450–490 nm using MK3 well reader (Thermo Fisher Scientific, USA) to evaluate the cell growth.
Scratch wound healing assay
A total of 2×105 cells were resuspended and subcultured into 6 well plates overnight to spread out a monolayer. The cells were scratched by sterile tips every 0.5 cm on dish and washed away the floated cells by PBS twice, cultured within non-FBS medium and captured images of the trace every 4 hrs. The edge distance and the scratch area were harvested using ImageJ to access the cell migration ability. The migration rate was calculated as (AreaT0-AreaTt)/AreaT0×100%.
Xenograft mouse model
This animals feeding and experiments were approved by the Institutional Animal Care and Use Committee of Zhengzhou University. Briefly, 6–8-week-old nude female mice (SLACCAS, China) were prepared. And 1×107 AGS cells with stably L1 overexpression were injected into the armpits of nude mice. Twelve individual mice were used in each group for this study. The tumors were isolated and accessed the tumor volume (V= length × width2×0.5)8 after one and two weeks.
RNA pull down assay
One microgram biotinylated ribonucleic exons of LINC00477 (L1: 4,567–4,695; L2: 15,390–15,993; L1&2: 16,800–17,202) (Figure 1B) were added with structure buffer (10 mM Tris pH 7.4, 0.1 M KCl, 10 mM MgCl2) to form the RNA secondary structure and incubated at 95°C for 2 mins, on ice for 3 mins, and room temperature for 30 mins in order to stabilize RNA. The denatured RNA was suspended by 100 μL streptavidin magnetic beads (Thermo Fisher Scientific, USA) and rotated overnight at 4°C. The next day, mixture were centrifuged at 3,000 rpm for 1 min and washed by wash buffer (150 mM KCl, 25 mM Tris pH 7.4, 5 mM EDTA, 0.5 mM DTT, 0.5% NP40, 100 U/mL RNAase inhibitor, 1× Protease inhibitors cocktail) three times, followed by the incubation and rotation with 1 mL AGS cell lysate at 37°C for 1 hr. Beads-RNA-proteins mixture was centrifuged at 3,000 rpm, washed by wash buffer additional three times, added 5×SDS for denaturation at 95°C for 10 mins and detected ACO1 and streptavidin level by SDS-PAGE.
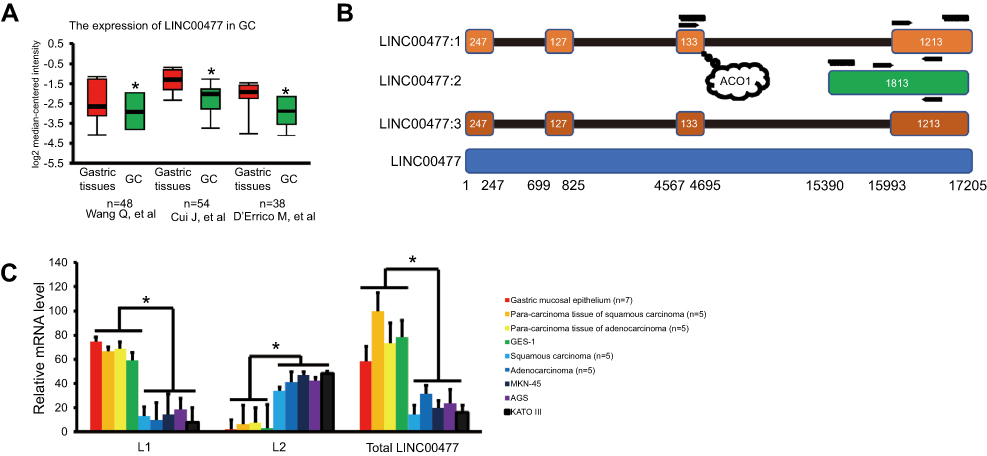 | Figure 1 The transcripts of LINC00477 in GC. (A) The total expression of LINC00477 in the individual GC studies from Oncomine database. Data from these studies.9–11(B) The scheme of LINC00477. LINC00477:1–3 represent isoform 1–3. The bottom means the genomic location of this lncRNA. Arrows mean the location of primers used in this study. The curves mean the synthetic target RNAs labeled by biotin. The bubble means the binding site of ACO1. (C) The mRNA level of L1, L2 and total LINC00477 in GC tissues, cells, and stomach cells. Each experiment was performed at least three duplications. “*” and “**” represents p-value < 0.05 and 0.01, respectively.Abbreviation: GC, gastric cancer. |
RNA immunoprecipitation (RIP)
A total of 1×107 AGS cells were harvested, resuspended in nuclear isolation buffer (1.28 M sucrose, 40 mM Tris pH 7.5, 20 mM MgCl2, 4% Triton X-100) and kept on ice for at least 30 mins with frequent mixing. The pellet nuclei were centrifuged by highest speed for 15 mins, resuspended by wash buffer (same with RNA pull down) and shear the chromatin through sonication by 30% power, 3 s on, 6 s off for 9 mins. After that, 90% nuclei were incubated with 1 μg ACO1 antibody overnight and 40 μl Protein A/G beads 2 hrs by gentle rotation at 4°C while the rest of 10% were harvested as input. The pellet beads were centrifuged by 3,000 rpm for 3 mins, washed three times. Both the input and pellet beads were purified by RNAiso plus and conducted reverse transcription which was described in RNA extraction.
Biochemical assay
The levels of intracellular citrate and isocitrate were measured by Citrate/Isocitrate Assay Kit (Biovision, USA). The cellular ferritin protein level was measured by ELISA (Abcam, UK). The experimental procedure was followed by the instructions provided.
Statistical analysis
All statistical analysis was performed in SPSS 20 (IBM Corporation, Armonk, NY, USA). Delta–delta method was used to normalize the real-time PCR data. One-way ANOVA was used to analyze the difference among the groups. p-Value < 0.05 was considered as statistical significance.
Results
The aberrant transcriptional pattern of LINC00477 in GCs
The presence of LINC00477 transcript profiles in three individual datasets of GC patients (the cases of 48,9 54,10 and 3811 paired gastric cancers) from Oncomine database (
The opposite function of spliced isoforms of LINC00477 in GC cells
Next, we further investigated the role of LINC00477 played in GC. Because we have found the presence of downregulation of L1 and upregulation of L2 in GC, we determined to change inversely but not intensify their transcriptional levels. Therefore, the overexpression of L1 and silencing of L2, respectively, in MKN-45, AGS, and KATO III cells were conducted and studied the impact on cancer proliferation and migration ability. We observed that the overexpression of L1 could robustly repress the cancer cell growth using CCK-8 assay (Figure 2A) and migration using wound healing assay (Figure 2B), while the silencing of L2 displayed more slight effect compared to L1. Furthermore, the stably L1 expression AGS cells and the xenograft mouse models were prepared to verify the tumor suppression of L1 in vitro. Consistently, we observed the inhibitory effect of AGS cells growth in vivo (Figure 2C). Collectively, our results demonstrated that the isoform 1 of LINC00477 acted as a tumor suppressor in GC cells.
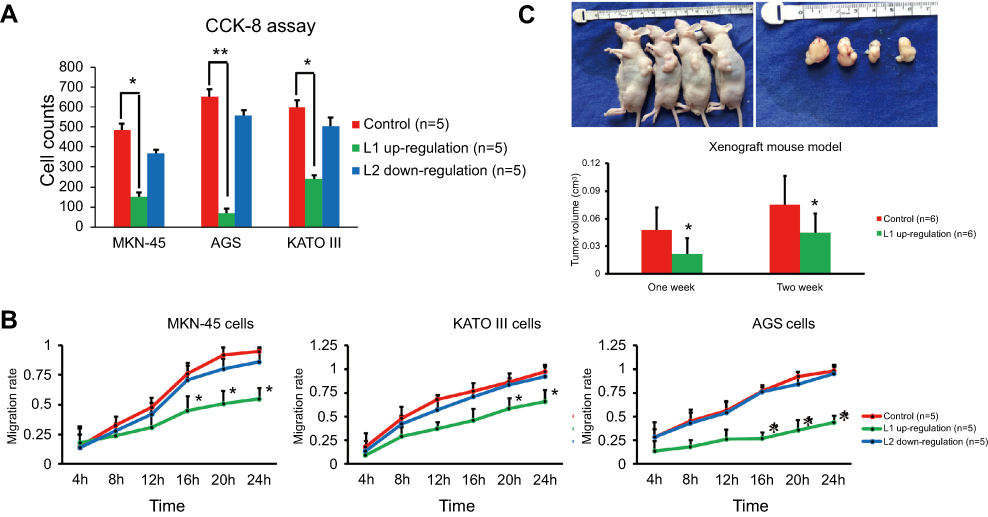 | Figure 2 The function of LINC00477 in GC cells. (A) The cell growth of MKN-45, AGS, and KATO III with L1 overexpression or L2 silence assessed by CCK-8 assay. (B) The migration ability of MKN-45, AGS, and KATO III with L1 overexpression or L2 silence assessed by wound healing assay. (C) The cell growth of AGS with L1 overexpression assessed by xenograft mouse model. Each experiment was performed at least three duplications. “*” represents p-value < 0.05.Abbreviation: GC, gastric cancer. |
The unique interaction between variant 1 of LINC00477 and ACO1 in GC cells
Next, we investigated the underlying mechanism of LINC00477 in GC. RBPDB database (
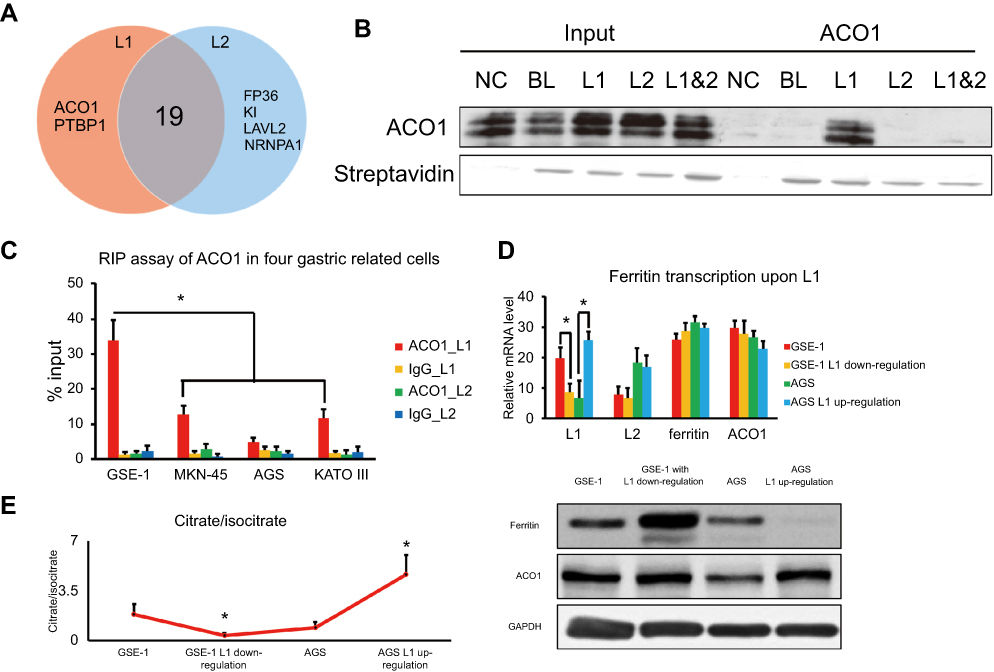 | Figure 3 The tumor suppressive role of L1 in GC. (A) The Venn diagram of RNA binding proteins of L1 and L2 predicted from RBPDB database. (B) RNA pull down assay for the interaction between L1 and ACO1 in AGS cells. NC, BL, L1, L2, and L1&2 represent negative control, non-LINC00477 RNA oligos, 4,567–4,695 of L1; 15,390–15,993 of L2; 16,800–17,202 of both L1 and L2, respectively. (C) RIP assay for the interaction between L1 and ACO1 in four cells. ACO1_L1, IgG_L1, ACO1_L2, and IgG_L2 mean the transcriptome of the four cells pulled down by ACO1 or IgG antibody and detected the enrichment of L1 or L2. (D) The RNA and protein level of ferritin upon L1 up-/downregulation in GSE-1 and AGS cells. (E) The proportion of citrate and isocitrate upon L1 up/downregulation in GSE-1 and AGS cells. Each experiment was performed at least three duplications. “*” represents p-value < 0.05. Abbreviation: GC, gastric cancer. |
Discussion
Recently, with the development of next-generation sequencing, an increasing number of lncRNAs which were previously considered as the junk DNA have been characterized and caused widespread attraction on the fields on epigenetics of histone modification and DNA methylation,14,15 transcription regulation via enhancer activation or insulator blocking16,17 as well as post-transcriptional regulation including alternative splicing,18,19 RNA stability,20 and subcellular localization21 in various biological events and diseases.
In our study, we found that LINC00477 was downregulated in GC patients compared to control stomach tissues or cells (Figure 1A). Unlike normal coding RNAs, a number of lncRNAs presented the low abundance transcripts because of an expression-dependent bias22 and the enormous isoforms due to the alternative splicing in RNA-seq.23 Therefore, RNA-seq might not reflect the true level of the multiple isoforms of LINC00477. We validated the expression pattern of two given LINC00477 isoforms in various stomach associated tissues or cells using real-time PCR, observed that the overall level of LINC00477 was downregulated in cancer although L2 was upregulated, and confirmed that LINC00477 was a potential biomarker that downregulated in GC compared to normal stomach control (Figure 1C) which was consistent with RNA-seq data. Here, we speculated that those multiple spliced isoforms might represent less biological significance, but the universally alternatively splicing specifically happened in noncoding exons.24
Moreover, we focused on L1 and L2 of LINC00477, and figured out that L1 played an important role of tumor suppressor in GC cells growth and migration (Figure 2). Compared to the robust tumor suppression presence of L1, L2 displayed a relatively slight effect on GC development. We speculated that L2 might be an alternative splicing product with less significant function. We did not further investigate the RNA secondary structure of L1 and L2, but assumed that the different exons of L1 distinct from L2 might contribute to the tumor suppressor function of GC. Based on this assumption, we next predicted the potential RNA binding proteins using RBPDB database. Most of the proteins included in the pattern of L1 and L2 binding proteins were mainly involved in pre-mRNA processing, alternative splicing, and RNA metabolism and transport (Figure 3A). In this study, we were more interested in the biological function of L1 in GC but not the underlying mechanism of LINC00477 splicing isoforms generation. Thus, we focused on ACO1 which was a unique cytosolic enzyme for regulating transferrin translation to control the level of intracellular iron and citrate/isocitrate conversion in tricarboxylic acid cycle, as well as aberrantly upregulated in leukemia and rectal carcinoma in previous studies.25,26 Interestingly, we determined that LINC00477 could repress the activation through the interaction with ACO1 by RNA pull down and RIP assay (Figure 3B and C). And an unknown reason resulted in L1 of LINC00477 reduction in GC lost the compromising activation of ACO1 and further gave rise to the disorder of glycometabolism (Figure 3D and E) for tumorigenesis.
Abbreviation list
GC, gastric cancer; LncRNA, long non-coding RNA; ACO1, aconitase 1; RIP, RNA immunoprecipitation.
Ethicals approval and informed consent
Guidelines for the care and use of animals of Institutional Animal Care and Use Committee of Zhengzhou University were followed.
Informed consent was obtained from all individual participants included in the study.
Acknowledgment
This study was supported by grants from National Natural Science Foundation of China (NSFC, Grant No. 31602600).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Luo G, Hu Y, Zhang Z, et al. Clinicopathologic significance and prognostic value of Ki-67 expression in patients with gastric cancer: a meta-analysis. Oncotarget. 2017;8(30):50273–50283. doi:10.18632/oncotarget.17305
2. Armero VES, Tremblay MP, Allaire A, et al. Transcriptome-wide analysis of alternative RNA splicing events in Epstein-Barr virus-associated gastric carcinomas. PLoS One. 2017;12(5):e0176880. doi:10.1371/journal.pone.0176880
3. Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA Cancer J Clin. 2013;63(1):11–30. doi:10.3322/caac.21166
4. Zhou X, Zhu W, Li H, et al. Diagnostic value of a plasma microRNA signature in gastric cancer: a microRNA expression analysis. Sci Rep. 2015;5:11251. doi:10.1038/srep11251
5. Malek E, Jagannathan S, Driscoll JJ. Correlation of long non-coding RNA expression with metastasis, drug resistance and clinical outcome in cancer. Oncotarget. 2014;5(18):8027–8038. doi:10.18632/oncotarget.2469
6. DiStefano JK. The emerging role of long noncoding RNAs in human disease. Methods Mol Biol. 2018;1706:91–110. doi:10.1007/978-1-4939-7471-9_6
7. Kwok ZH, Tay Y. Long noncoding RNAs: lincs between human health and disease. Biochem Soc Trans. 2017;45(3):805–812. doi:10.1042/BST20160376
8. Naito S, von Eschenbach AC, Giavazzi R, Fidler IJ. Growth and metastasis of tumor cells isolated from a human renal cell carcinoma implanted into different organs of nude mice. Cancer Res. 1986;46(8):4109–4115.
9. Wang Q, Wen YG, Li DP, et al. Upregulated INHBA expression is associated with poor survival in gastric cancer. Med Oncol. 2012;29(1):77–83. doi:10.1007/s12032-010-9766-y
10. Cui J, Li F, Wang G, Fang X, Puett JD, Xu Y. Gene-expression signatures can distinguish gastric cancer grades and stages. PLoS One. 2011;6(3):e17819. doi:10.1371/journal.pone.0017819
11. D’Errico M, de Rinaldis E, Blasi MF, et al. Genome-wide expression profile of sporadic gastric cancers with microsatellite instability. Eur J Cancer. 2009;45(3):461–469. doi:10.1016/j.ejca.2008.10.032
12. Volders PJ, Helsens K, Wang X, et al. LNCipedia: a database for annotated human lncRNA transcript sequences and structures. Nucleic Acids Res. 2013;41(Databaseissue):D246–D251. doi:10.1093/nar/gks915
13. Cook KB, Kazan H, Zuberi K, Morris Q, Hughes TR. RBPDB: a database of RNA-binding specificities. Nucleic Acids Res. 2011;39(Databaseissue):D301–D308. doi:10.1093/nar/gkq1069
14. Inoue A, Jiang L, Lu F, Zhang Y. Genomic imprinting of Xist by maternal H3K27me3. Genes Dev. 2017;31(19):1927–1932. doi:10.1101/gad.304113.117
15. Wang L, Zhao Y, Bao X, et al. LncRNA Dum interacts with Dnmts to regulate Dppa2 expression during myogenic differentiation and muscle regeneration. Cell Res. 2015;25(3):335–350. doi:10.1038/cr.2015.21
16. Stavropoulos N, Rowntree RK, Lee JT. Identification of developmentally specific enhancers for Tsix in the regulation of X chromosome inactivation. Mol Cell Biol. 2005;25(7):2757–2769. doi:10.1128/MCB.25.7.2757-2769.2005
17. Lefevre P, Witham J, Lacroix CE, Cockerill PN, Bonifer C. The LPS-induced transcriptional upregulation of the chicken lysozyme locus involves CTCF eviction and noncoding RNA transcription. Mol Cell. 2008;32(1):129–139. doi:10.1016/j.molcel.2008.07.023
18. Tripathi V, Ellis JD, Shen Z, et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol Cell. 2010;39(6):925–938. doi:10.1016/j.molcel.2010.08.011
19. Ciarlo E, Massone S, Penna I, et al. An intronic ncRNA-dependent regulation of SORL1 expression affecting Abeta formation is upregulated in post-mortem Alzheimer’s disease brain samples. Dis Model Mech. 2013;6(2):424–433. doi:10.1242/dmm.009761
20. Zhao HY, Wu HJ, He JL, et al. Chronic sleep restriction induces cognitive deficits and cortical beta-amyloid deposition in mice via BACE1-antisense activation. CNS Neurosci Ther. 2017;23(3):233–240. doi:10.1111/cns.12667
21. Johnsson P, Ackley A, Vidarsdottir L, et al. A pseudogene long-noncoding-RNA network regulates PTEN transcription and translation in human cells. Nat Struct Mol Biol. 2013;20(4):440–446. doi:10.1038/nsmb.2516
22. Hardwick SA, Chen WY, Wong T, et al. Spliced synthetic genes as internal controls in RNA sequencing experiments. Nat Methods. 2016;13(9):792–798. doi:10.1038/nmeth.3958
23. Conesa A, Madrigal P, Tarazona S, et al. A survey of best practices for RNA-seq data analysis. Genome Biol. 2016;17:13. doi:10.1186/s13059-016-0881-8
24. Deveson IW, Brunck ME, Blackburn J, et al. Universal alternative splicing of noncoding exons. Cell Systems. 2018;6(2):245–255.e245. doi:10.1016/j.cels.2018.02.008
25. Martin-Lorenzo A, Auer F, Chan LN, et al. Loss of Pax5 exploits Sca1-BCR-ABL(p190) susceptibility to confer the metabolic shift essential for pB-ALL. Cancer Res. 2018;78(10):2669–2679. doi:10.1158/0008-5472.CAN-17-3262
26. Choi SY, Jang JH, Kim KR. Analysis of differentially expressed genes in human rectal carcinoma using suppression subtractive hybridization. Clin Exp Med. 2011;11(4):219–226. doi:10.1007/s10238-010-0130-5
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.