")
Back to Journals » International Journal of Nanomedicine » Volume 13
The impact of liposomal linolenic acid on gastrointestinal microbiota in mice
Authors Li XX, Shi S, Rong L, Feng MQ, Zhong L
Received 16 September 2017
Accepted for publication 7 December 2017
Published 9 March 2018 Volume 2018:13 Pages 1399—1409
DOI https://doi.org/10.2147/IJN.S151825
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Linlin Sun
Xuan-xuan Li,1 Si Shi,2 Lan Rong,1 Mei-qing Feng,2 Liang Zhong1
1Department of Digestive Diseases, Huashan Hospital Affiliated to Fudan University, Shanghai, China; 2School of Pharmacy, Fudan University, Shanghai, China
Background: The prevalence of Helicobacter pylori has long been a global health issue. Triple therapy, being the first-line treatment, has caused dysbiosis of the gastrointestinal tract that led to various complications. A novel nanomedicine – liposomal linolenic acid (LipoLLA) – has been proven to have great potential in eradicating H. pylori. However, the possible side effects of LipoLLA due to alteration of the gastrointestinal microbiota remain unknown.
Aim: This study focused on the impact of LipoLLA on gastrointestinal microbiota in mice in comparison with triple therapy in order to assess the safety profile.
Methods: Mice were divided into five groups: blank control group; H. pylori control group; triple therapy group; low-dose LipoLLA group (25 mg/kg); and high-dose LipoLLA group (50 mg/kg). Fecal samples were collected before and after the intake of corresponding formulas. Gastric tissues were obtained after mice dissection. These samples were analyzed with high-throughput sequencing.
Results: The analysis revealed that LipoLLA resulted in minor gut microbiota alteration at different levels. The altered proportions in the high-dose group were higher than that of the low-dose group. On the other hand, the triple therapy group showed dramatic shifts in the major community composition. It displayed a notable boost in the relative abundance of Proteobacteria and Firmicutes along with a decrease in that of Verrucomicrobia and Bacteroidetes. All of them belonged to the major phyla in the microbiome. Triple therapy also led to the growth of the family Enterobacteriaceae, Enterococcaceae, and Clostridiaceae_1 that may be associated with clinical illnesses. Gastric microbiota analysis reached similar conclusions.
Conclusion: Our findings indicated that LipoLLA causes minor gastrointestinal microbiota alterations while the triple therapy triggered dramatic changes. Thus, LipoLLA is not only promising but also a safe therapeutic medication to eradicate H. pylori infection.
Keywords: nanomedicine, linolenic acid, Helicobacter pylori, gastric microbiota, gut microbiota
Introduction
Helicobacter pylori (H. pylori) is one of the most common chronic bacterial infections in humans that may lead to chronic gastritis, peptic ulcer disease, or gastric malignancy. It is found in around half of the world population, thus forming a global issue.1–3
The 1-week triple therapy that consists of a proton-pump inhibitor, clarithromycin, and amoxicillin or metronidazole proposed at the first Maastricht conference remains the standardized first-line treatment of H. pylori infection.4,5 However, eradication rates have declined to 60%–75% owing to its high cost, poor compliance among patients, and the development of resistant H. pylori strains.6,7 Among all the side effects, some of the conditions result from the impact of antibiotics on the gastrointestinal microbiota.8,9 Gastrointestinal microbiota are a large community of symbiotic microorganisms that the gastrointestinal tract harbors. The host provides a physical niche and nutrition to the microbiota that in turn benefits the host by enhancing the resistance to infection. Furthermore, they also facilitate the absorption of ingested food and contribute to the development of the immune system of the host. Changes in the microbiota have been linked to a variety of diseases including infections, obesity, diabetes mellitus, arthritis, and inflammatory bowel diseases.10–13
A novel antibacterial nanomedicine, the liposomal nanoformulation of linolenic acid (LipoLLA), has been introduced and its bactericidal activity against the resistant strains of H. pylori has been proved.14 It is viewed as a potential replacement of triple therapy for the following reasons. Firstly, in the in vitro experiment, LipoLLA was found to be capable of killing both spiral and coccoid forms of the H. pylori.14 The coccoid form (also known as the dormant form) plays a critical role in infection transmission and relapses after inappropriate treatment; amoxicillin is known as only capable of inhibiting the spiral form of H. pylori.15 Secondly, the antibacterial mechanism of LipoLLA was proved to be associated with higher permeability of bacterial cell membrane, which is highly destructive.16 Amoxicillin works by inhibiting bacterial cell wall biosynthesis, and this can explain its lack of efficacy in inhibiting the coccoid H. pylori due to the “dormant” nature. Thirdly, LipoLLA killed H. pylori regardless of their resistance status to metronidazole, and did not arouse resistance afterwards.14 What is more, the in vivo test showed that the therapy also reduced levels of proinflammatory cytokines including interleukin 1β, interleukin 6, and tumor necrosis factor α.17
However, a few studies suggested that nanodrugs may cause adverse health effects mediated by microbiome through either directly eliminating the gastrointestinal microbiota or by alternating their functions. For example, silver nanoparticles could selectively damage the gut microbiota despite the fact that silver is generally considered to have low toxicity. This might be due to their small size, and unique physical and chemical properties.12,18,19
Currently, it is unclear whether LipoLLA poses harmful effects by affecting the gastrointestinal microbiota. Thus, we investigated the impact of LipoLLA intake on the gastrointestinal microbiota in mice groups.
High-throughput sequencing has wide applications in research in regard to the development of next-generation sequencing (NGS) technologies. The Illumina MiSeq genome analyzer, a representative NGS system, is used in this experiment.20,21
Materials and methods
This study was approved by the Experimental Animal Ethics Committee of the School of Pharmacy, Fudan University (ethical approval number: 2016-10-WS-FMQ-01). The Experimental Animal Welfare Ethical Review Guide for the care and use of animals was followed.
Preparation of LipoLLA
LipoLLA was prepared with a standard vesicle extrusion method following the protocols from existing articles.16,17 Briefly, L-α-phosphatidylcholine (EggPC), cholesterol (Avanti Polar Lipids, Inc, Alabaster, AL, USA), and linolenic acid (LLA; Ultra Scientific, North Kingstown, RI, USA) were blended in a weight ratio of 6:1:3 (16 mg in total) and dissolved in 4 mL chloroform. The chloroform was then evaporated for 1 hour, rehydrated by adding 2 mL phosphate buffered saline, vortexed for 1 minute, and sonicated for 3 minutes. Then a Ti-probe on a Branson 450 sonifier was used to sonicate the solution at 20 W for 2 minutes to form the lipid vesicles, which were then extruded through a 100 nm pore-sized polycarbonate membrane 11 times to form the final LipoLLA. Afterwards, a Sephadex G75 column (Thermo Fisher Scientific, Waltham, MA, USA) was used to remove the unloaded LLA from the suspension. Then, the solution containing LipoLLA was sterilized by filtration via a 0.22 μm filter unit (Merck Millipore, Billerica, MA, USA). To quantify the loading efficiency, LLA or LipoLLA was dried on a rotavapor (Buchi Corporation, New Castle, DE, USA; model R-124), dissolved in methanol, and derivatized with phenacylester. The final solution with LLA phenacylester derivatives was analyzed using liquid chromatography with a C18 column (PerkinElmer Inc, Waltham, MA, USA). The hydrodynamic size (diameter, nm) and surface ζ-potential (mV) of LipoLLA were measured by dynamic light scattering with a Malvern Zetasizer ZS (Malvern Instruments, Malvern, UK). All characterization measurements were repeated three times at 25°C.
H. pylori culture
H. pylori Sydney strain 1 was used in this study. The bacteria were routinely maintained on Columbia agar supplemented with 5% laked horse blood under microaerobic conditions (10% CO2, 85% N2, and 5% O2) at 37°C, according to a previous description.22
Sample collection
Thirty male C57BL/6 mice at 7 weeks weighing about 22 g were obtained and allowed to adapt to the Fudan University Zhangjiang Campus animal laboratory for 1 week. They were then randomly divided into five groups (n=6): (group A) the blank control group, (group B) the H. pylori control group, (group C) the triple therapy group, (group D) the low-dose LipoLLA group, and (group E) the high-dose LipoLLA group. All four groups other than group A were inoculated with H. pylori to establish the infection model. Each mouse received 0.3 mL of 1×109 colony forming units (CFU)/mL H. pylori in brain heart infusion broth administered via oral gavage every 48 hours for three times. The infection was able to develop 2 weeks after inoculation.
All mice were then treated with the assigned regimens once daily for 5 consecutive days.
Omeprazole (400 μmol/kg) was given to all mice 30 minutes before the treatment in order to neutralize gastric acid and to prevent potential drug degradation.
Group C was then treated with clarithromycin (14.3 mg/kg) and amoxicillin (28.5 mg/kg). Group D and E were given low-dose LipoLLA (25 mg/kg) and high-dose LipoLLA (50 mg/kg) for 1 week. Group A and B were treated with same volume of phosphate buffered saline (0.4 mL).
Fecal samples were collected twice in all five groups. They were collected before (A1, B1, C1, D1, E1) and after (A2, B2, C2, D2, E2) 5 days administration of different formulas, making 10 specimen groups in total. Samples were all immediately stored in sterile containers and frozen at −80°C until DNA extraction.
We also obtained gastric tissue of the mice from three groups – groups B, C, and E – to observe the influence of different regimens on the composition of gastric microbiota. The mice were killed using cervical dislocation and dissected to retrieve the gastric tissues. The stomachs were opened along the greater curvature, washed with normal saline (NS), and then homogenized and placed in 1 mL NS.
DNA extraction and PCR amplification
The E.Z.N.A.® Soil DNA Kit (Omega Bio-Tek Inc, Norcross, GA, USA) was used to extract the microbial DNA from fecal samples. PCR was used to amplify the V3–V4 regions of the bacteria 16S rDNA (95°C for 3 minutes, followed by 27 cycles at 95°C for 30 seconds, 55°C for 30 seconds, 72°C for 45 seconds, and a final extension at 72°C for 5 minutes). Primers 338F 5′-ACTCCTACGGGAGGCAGCAG-3′ and 806R 5′-GGACTACHVGGGTWTCTAAT-3′ were used. PCR reactions were performed in the triplicate of 20 μL mixture that contained 2 μL of 2.5 mM dNTPs, 4 μL of 5× FastPfu Buffer, 0.4 μL of FastPfu Polymerase, 0.8 μL of each primer (5 μM), and 10 ng of template DNA.
Illumina MiSeq sequencing
Amplicons were extracted from 2% agarose gels and were purified with the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA) following the manufacturer’s instructions. They were also quantified with QuantiFluor™-ST (Promega Corporation, Fitchburg, WI, USA). Purified amplicons were combined in equimolars and paired-end sequenced (2×250) on the Illumina MiSeq platform according to the standard protocol.
Processing of sequencing data
QIIME (version 1.17; Knight Lab, Evanston, IL, USA) was used to demultiplex and quality-filter the raw fastq files with the following criteria: 1) the 300 bp reads were shortened at the sites where the average quality score was <20 over a sliding window of 50 bp while disposing and the truncated reads were shorter than 50 bp; 2) exact barcode matching: two nucleotide mismatch in primer matching and readings that contained ambiguous characters were removed; 3) the only sequences overlapping longer than 10 bp were assembled accordingly. Reads that were unable to be assembled were discarded.
Operational taxonomic units (OTUs) were clustered with 97% similarity cutoff with UPARSE (version 7.1).37 The chimeric sequences were identified and removed using UCHIME.38 The taxonomy of every 16S rRNA gene sequence was analyzed by Ribosomal Database Project Classifier against the SILVA 16S rRNA database (SSU115) using the confidence threshold of 70%.
Results
Characteristics of LipoLLA
Due to the amphiphilic nature, LLA was readily loaded into liposomes and could subsequently merge with bacteria for bactericidal action. Here, EggPC, cholesterol, and LLA were mixed in a weight ratio of 6:1:3 and then extruded to form LipoLLA. The formulated LipoLLA had a hydrodynamic diameter of 109.7±0.2 nm, a polydispersity index of 0.18±0.01, and a surface ζ-potential of −53.9±1.0 mV.
Gut microbiota analysis
Richness and diversity index (Table 1)
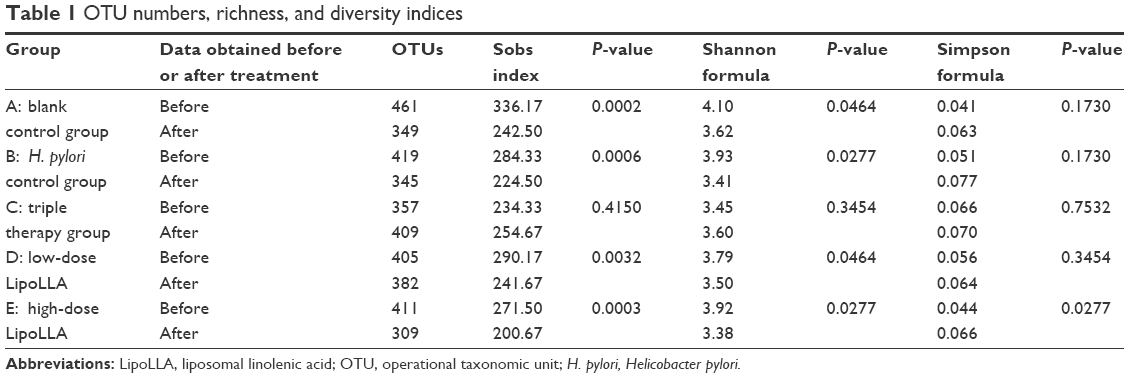 | Table 1 OTU numbers, richness, and diversity indices |
We generated a database consisting of 2,233,113 total sequences, with a mean of 37,219±3,838 (mean ± SD) reads per sample. The range of the number of the reads for the samples was from 30,049 to 44,905. There were up to 520 OTUs among all subjects. The numbers of OTUs in each group are shown in Table 1. The OTU richness in each group was roughly at the same level. We observed an insignificant increase in the OTU number in group C after treatment (P>0.05), while the other groups showed a decline. Group D had a minimal reduction, while group E decreased more, but no more than group A.
Richness and biodiversity indices were obtained with the Mothur software package.39 Richness, which is related to the observed OTUs number, was estimated with the Sobs index. The Shannon and Simpson formula was used for diversity estimation. It depended on how uniformly the sequences were spread in the different observed OTUs. All indices were evaluated with an OTU distance unit cutoff of 0.03 in order to test selectivity in the definition of OTUs. Paired t-test or Wilcoxon signed-rank test was used to test whether the mean differences between the indices before and after treatment were significant. Group C caused a slight increase in all three indices, but none of the results was significant (P>0.05). All other groups displayed similar changes: decrease in the Sobs and Shannon index, and increase in the Simpson estimator, suggesting declined richness and diversity. The differences were mostly significant except in the Simpson index, where only group E showed significance (P<0.05).
β Diversity analysis
To demonstrate intergroup differences or similarities, we assessed the β diversity using hierarchical clustering and principal component analysis (PCA) at different levels.
Hierarchical clustering
Hierarchical clustering was performed by using the unweighted pair-group method with arithmetic mean and dendrograms were constructed at the species level (Figure 1A and B). The distances between branches represented the differences of species composition between samples. It showed that, at the species level, there were no significant intergroup differences before giving the formulations. Afterwards, group C showed more similarity within the group than with any of the other groups. The LipoLLA groups, both group D and E, had no notable differences compared with group A and B.
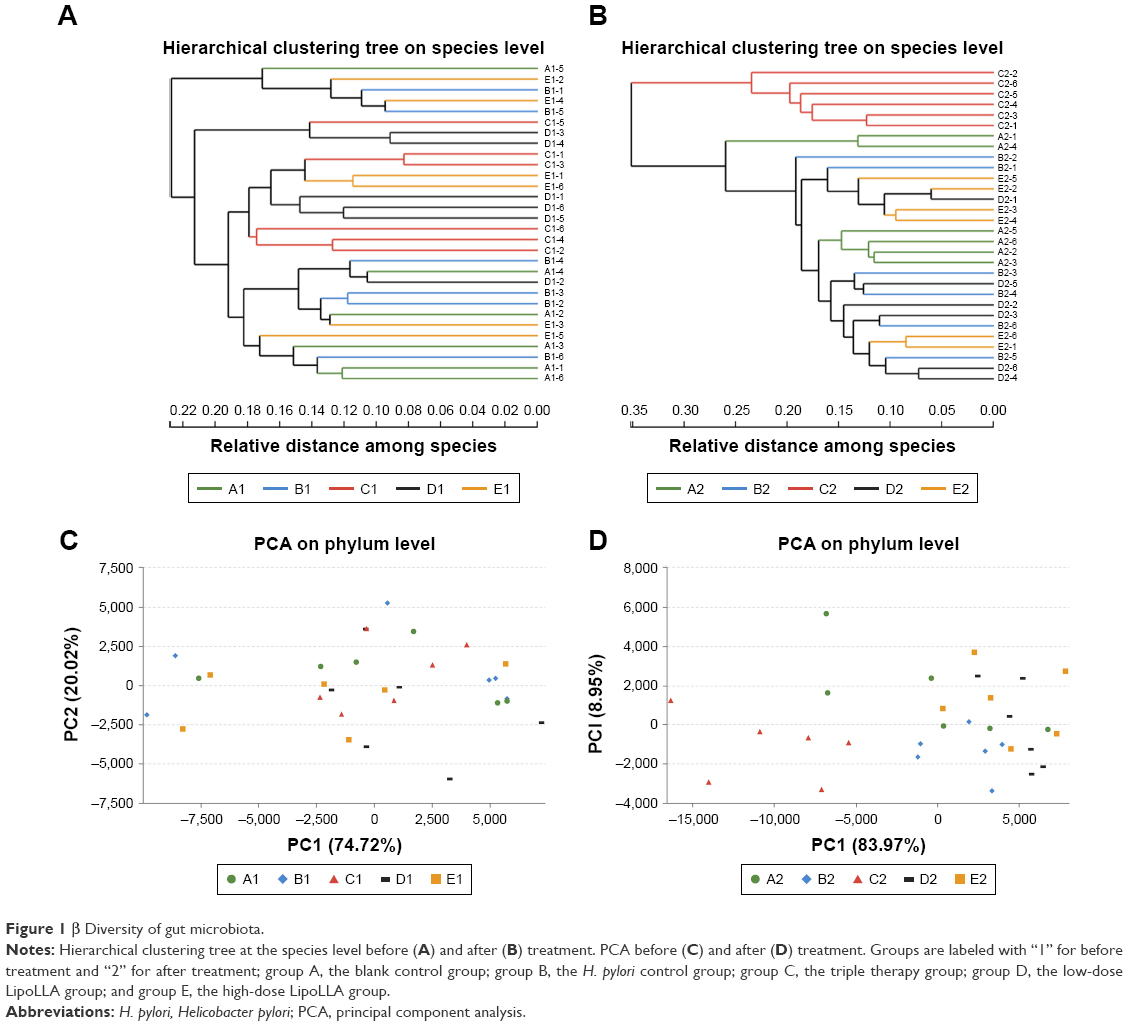 | Figure 1 β Diversity of gut microbiota. |
PCA
PCA also provided visualization of relative distances between the microbiota of each sample. At the phylum level (Figure 1C and D), group C clustered away from other groups after treatment, while group D and E were overall close to group A and B, suggesting that these bacterial communities were more similar to each other than to those of group C.
The β diversity analysis revealed that the compositions of microbiome remained similar among the other groups after administration of corresponding formulas, whereas those in group C were distinctively changed.
Microbiota composition
Eleven phyla were found in total among all subjects. Two of the phyla, Bacteroidetes and Firmicutes, were predominant in the gut microbiota of all subjects. Smaller populations included Verrucomicrobia, Cyanobacteria, Proteobacteria etc.
The Circos graph is effective in visualizing relationships between the microbiota and samples.23,24 These graphs not only reflect the proportion of the dominant phyla in each group, but also reflect the distribution of each dominant phylum in the different groups with different width of each side of the connecting bands between the sample groups and microbiota compositions. As shown in Figure 2A and B, the composition of group C was notably altered with a boost in the relative abundance of Proteobacteria with a relative decrease in that of Verrucomicrobia. There was also a decline in Bacteroidetes and an increase in Firmicutes. The changes in the rest of the groups were unnoticeable.
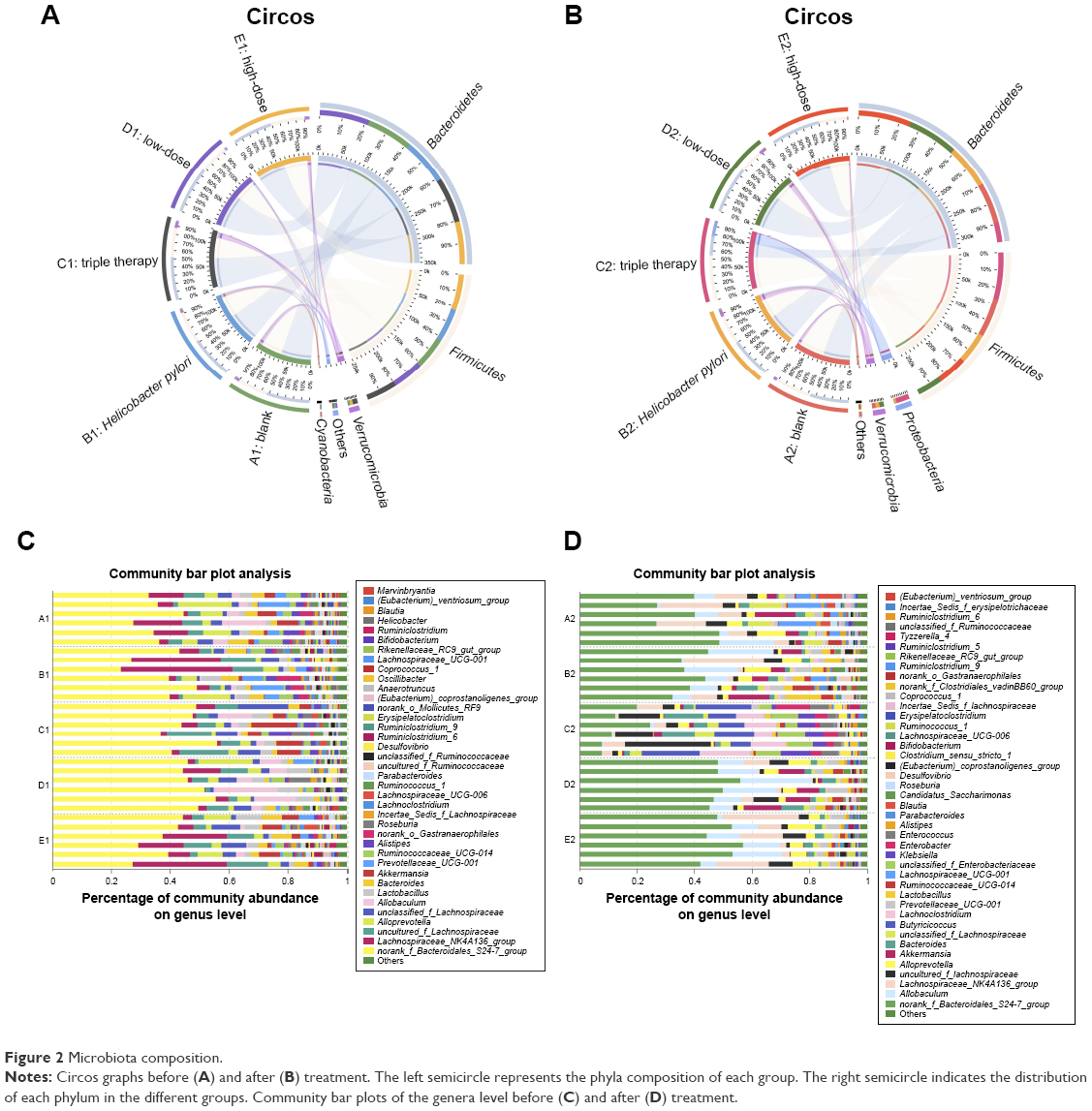 | Figure 2 Microbiota composition. |
Bar plot analysis at the genus level (Figure 2C and D) roughly showed that the abundance of different genera varied among the samples. The norank_f_Bacteroidales_S24-7_group predominated in the majority of all samples. Group C exhibited the most remarkable change in the composition after treatment. The percentage of the previously dominating genus was diminished, but a few minor communities that were hardly seen in other groups grew. These included Enterobacter, Klebsiella etc. However, both groups D and E were generally stable in the composition.
Analysis of community differences – comparison of multiple groups
One-way analysis of variance (ANOVA) bar plot at the phylum level
At the phylum level, we noticed significant alterations in several specific phyla (P<0.05) after treatment on the one-way ANOVA bar plot (Figure 3A and B). Bacteroidetes, Firmicutes, and Proteobacteria showed the most noteworthy changes, while the other altered phyla were either minor in abundance or had similar percentages before and after treatment (P>0.05).
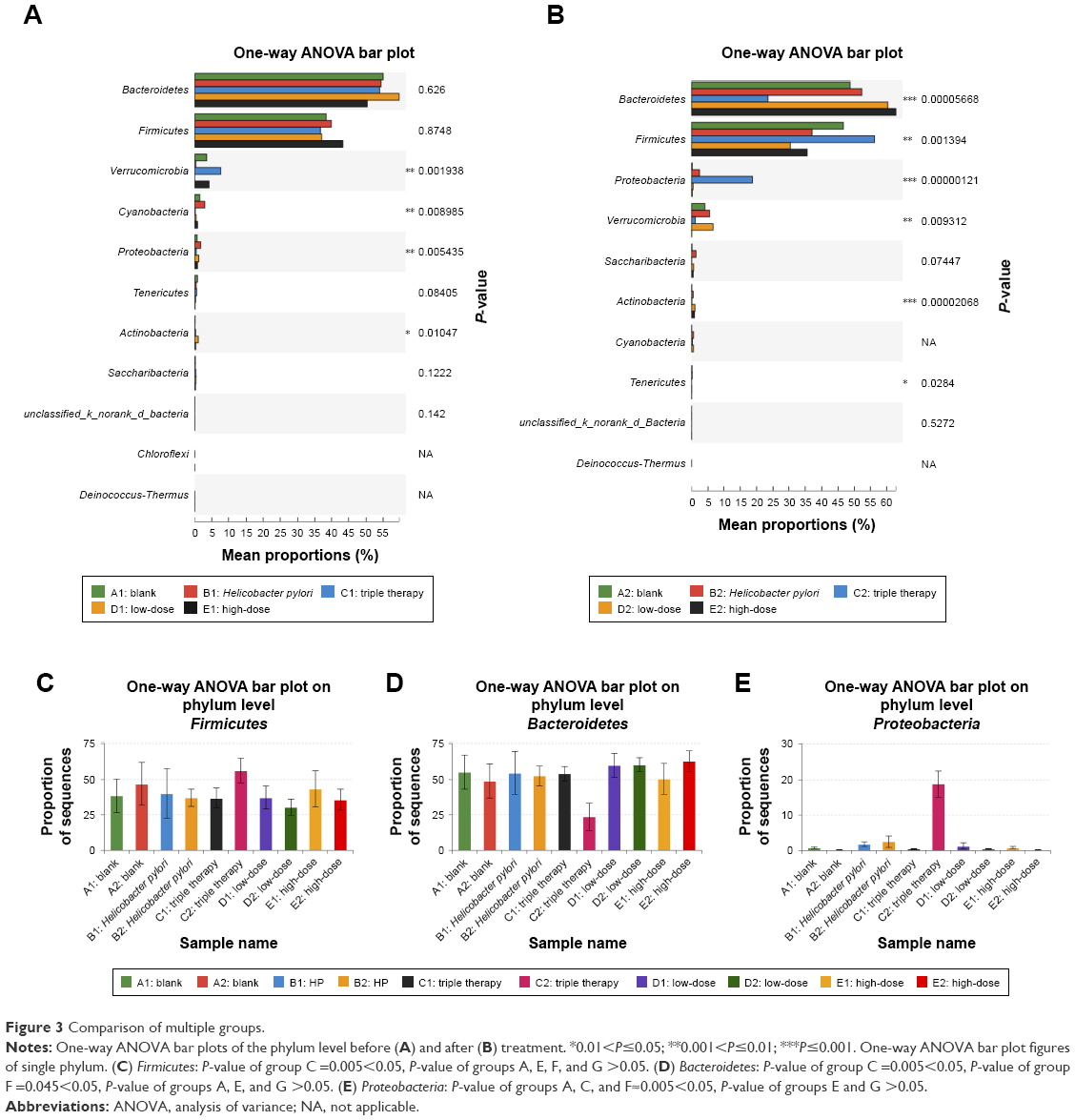 | Figure 3 Comparison of multiple groups. |
The one-way ANOVA bar plot figures of single phylum were made to demonstrate which groups contributed to the changes of three major phyla (Figure 3C–E). The level of Firmicutes rose in group C (P<0.05). Other groups displayed an insignificant drop except for group A that increased slightly (P>0.05) (Figure 3C). Group C also exhibited an evident decline in the level of Bacteroidetes and a remarkable increase in the Proteobacteria level (Figure 3D and E). Changes in the other groups were less significant. Group E had more significant changes than group D, but these two were more close to the control groups comparing with group C.
Unique communities in triple therapy group after treatment (Table 2)
 | Table 2 Unique communities in triple therapy group after treatment (C2) |
We found some communities that were barely noticeable in all the other groups either before or after treatment. They were only notable in group C after treatment (C2). At the family level, they were Enterobacteriaceae, Enterococcaceae, and Clostridiaceae_1. At the genus level, they were Butyricicoccus, Lachnoclostridium, Klebsiella, Clostridium_sensu_stricto_1, and Tyzzerella_4.
Gastric microbiota
The microbiota composition in the stomachs after treatment was compared.
On the one-way ANOVA bar plot at the genus level (Figure 4A), the level of Helicobacter had a significant drop in group C and group E after administration in comparison with group B. Group E showed a stronger effect.
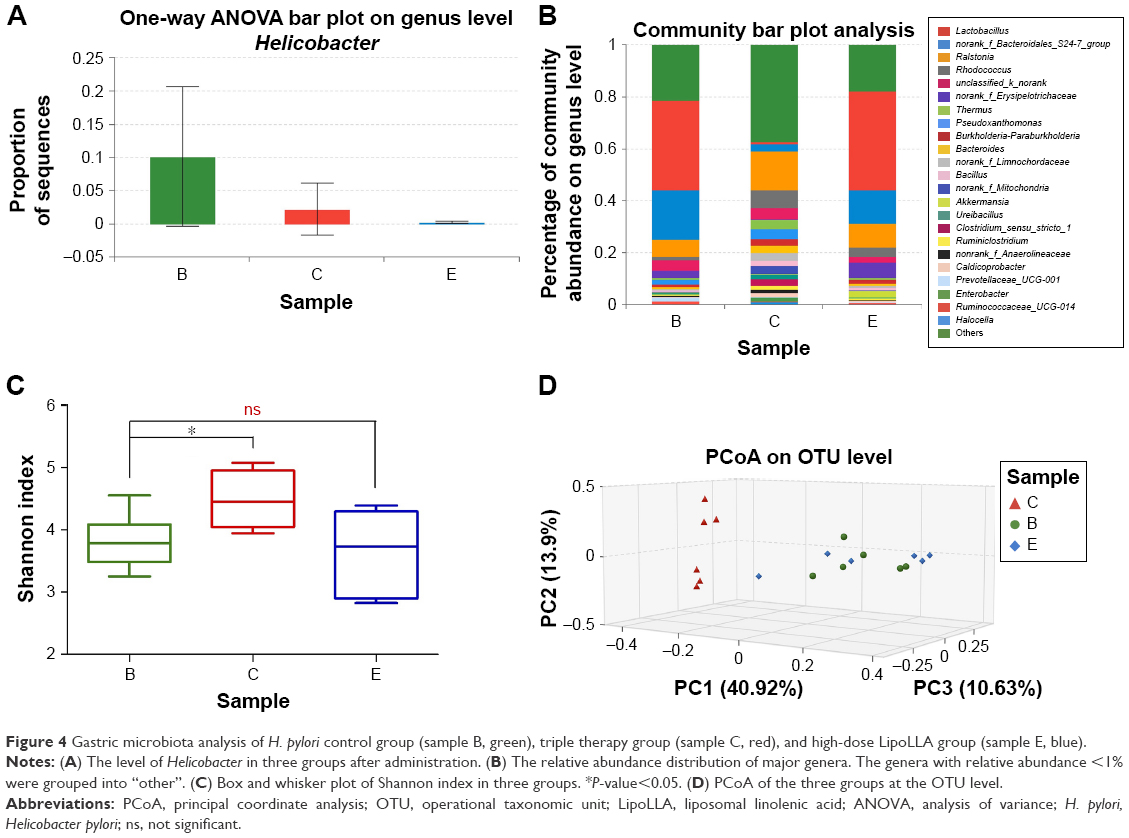 | Figure 4 Gastric microbiota analysis of H. pylori control group (sample B, green), triple therapy group (sample C, red), and high-dose LipoLLA group (sample E, blue). |
The community bar plot analysis (Figure 4B) presented the relative abundance distribution of major genera in groups B, C, and E. The genera with relative abundance <1% were grouped into “others”. Group B and group E displayed a similar composition, with Lactobacillales, norank_f_Bacteroidales_S24-7_group, and Ralstonia as the predominant genera. Notwithstanding, group C showed prominent difference, with only Ralstonia and Rhodococcus accounting for over 5%.
An obvious difference could be noted in the diversity of the gastric commensal bacteria with the Shannon index. A significant increase in the diversity of the gastric bacterial community was distinctly observed in group C, while the other two groups had a similar average level of diversity (P>0.05) (Figure 4C). Principal coordinate analysis also revealed that group C clustered away from the other two groups (Figure 4D). There were no significant intergroup differences for bacterial composition between group B and group E at the genus level displayed by the Wilcoxon rank-sum test bar plot with the exception of the proportions of Rhodococcus, Akkermansia, Prevotellaceae, and Pseudoxanthomonas (Figure 5A). However, the relative abundance of over half of the major genera in group C had a significant shift, compared to group B (Figure 5B). It is not hard to conclude from these data that the bacterial composition of group C exhibited considerable differences from that of groups B and E.
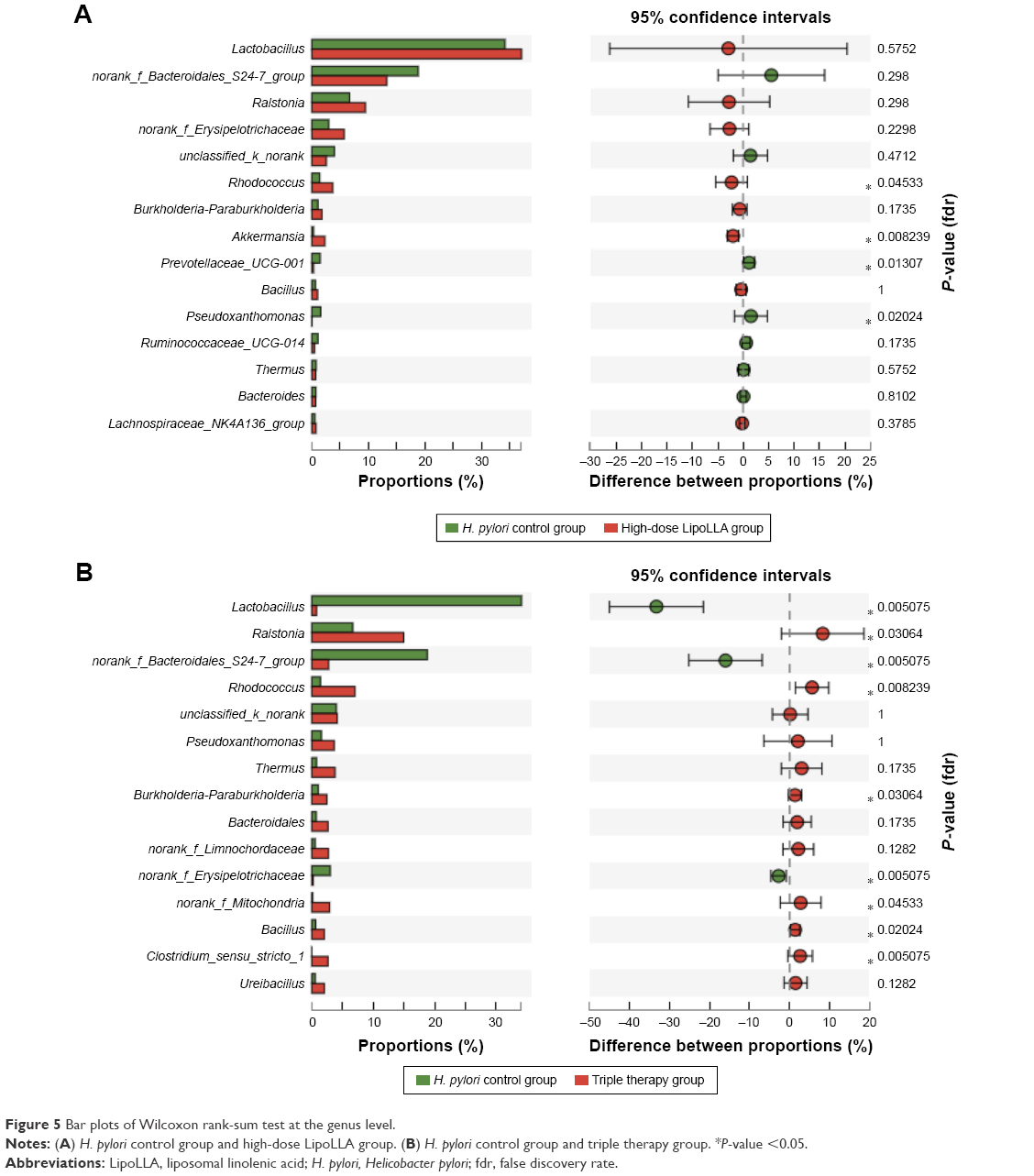 | Figure 5 Bar plots of Wilcoxon rank-sum test at the genus level. |
Discussion
The prevalence of H. pylori has long been a health issue worldwide. However, the current first-line treatment, triple therapy, was reported to have drawbacks especially the dysbiosis of gastrointestinal microbiota. Therefore, new therapeutical agents are being widely investigated. A new nanomedicine – LipoLLA – is proved to have great potential. This in vivo study left out the bactericidal act of LipoLLA toward H. pylori, but focused on its impact on gastrointestinal microbiota for the first time, and compared the results to the standard triple therapy regimen.
In this study, we adopted the high-throughput sequencing technique of the V3–V4 regions of bacteria 16S rRNA. This so-called massively parallel sequencing technique is able to obtain a tremendous load of sequence data in a short period of time.
The reported adverse effects on the microbiota by the triple therapy are mainly due to antibiotics, which are essential elements of the treatment. The antibiotic treatment-induced microbiota alterations in quantity and quality were observed back in 1999.25 It was reported that even short-term antibiotic use was able to shift the microbiome to long-term dysbiotic states that were characterized by a loss of taxonomy, and shifts in functional diversity and metabolic capacity as well as reducing the effects of the immune defensive system against pathogens.8,9,26
From either the α or β diversity analysis, the composition of both the gut and gastric microbiota in the triple therapy group presented substantial shifts at various levels after the administration compared to the rest of the groups. Conversely, the composition of the gastrointestinal microbiota in LipoLLA groups (groups D and E) was very close to the control groups (groups A and B). These findings strongly indicated that LipoLLA is generally safe without arousing tremendous changes in the microbiota, especially in comparison with triple therapy. Besides, the result of lowering gut microbial diversity of the two control groups implied that simply giving the mice drugs through oral gavage could result in stress and altered the gut microbiota.
Consistent with earlier reports, Bacteroidetes and Firmicutes were two predominant phyla in the gut microbiota of all the mice, followed by smaller populations such as Verrucomicrobia, Cyanobacteria, and Proteobacteria.5,11,27,28 Dysbiosis caused by antibiotics was commonly reported to be characterized by a reduced proportion of the phyla Firmicutes and Bacteroidetes, with an increased abundance of Proteobacteria, and accompanied by an overgrowth of the genera Enterococcus spp., Clostridium spp., and Enterobacteriaceae and the family Enterobacteriaceae.5,25,29,30 In our study, we observed an evident rise in Firmicutes and Proteobacteria with a concomitant decline in Bacteroidetes and Verrucomicrobia in the triple therapy group after treatment. The difference in influence on the phylum Firmicutes in the present study from other previous studies may be due to the different animal model used in the experiments and related to particular antibiotics. In the LipoLLA groups, however, we found minor changes in these important bacterial communities.
In addition, we observed a few overgrown unique populations only in the triple therapy group listed in Table 2, among which the family Enterobacteriaceae, Enterococcaceae, and Clostridiaceae_1 were noteworthy since they could be clinically important opportunistic bacteria and are major causes of hospital-acquired infections, and thus may play an important part in antibiotic-induced diseases.9,25,30 In addition, the growth of these communities could be an explanation for the nonsignificant rise of the OTU numbers in the triple therapy (Table 1).
In LipoLLA groups, the changed proportions of the gut microbiota in the high-dose group (50 mg/kg) were slightly higher than the low-dose group (25 mg/kg) (Table 1 and Figure 3). It could be deduced that low-dose LipoLLA is safer.
Traditionally, the human stomach has been viewed as an inhospitable environment for microorganisms because of the acidic conditions (median pH 1.4) and other antimicrobial factors. With the discovery of H. pylori and subsequent insight into the mechanisms of how these organisms adapt, the existence of a bacterial community in this harsh habitat became known. Consequently, however, the bacterial load is much lower in the stomach (102–104 CFU/mL) than in the colon (1010–1012 CFU/mL).31–33 Hence, gastric microbiota is less significant in human health than the gut microbiota, and is less tested in drug safety assessment. In addition, gastric tissue was unattainable until the mice were sacrificed; thus, only data after treatment were obtained. Nonetheless, the findings from the groups we chose in gastric microbiota analysis confirmed the aforesaid conclusion that the triple therapy group ended up with remarkable bacterial alterations.
Furthermore, the gastric microbiota analysis was able to roughly suggest the effect of the drugs on H. pylori. The species H. pylori was hardly detectable due to the limitation of 16S rRNA gene sequence on the accuracy at the species level. Therefore, we paid attention to the changes of the abundance of the genus Helicobacter, to which H. pylori belongs, as shown in Figure 4A. It was shown that the triple therapy and LipoLLA both succeeded in reducing the proportion of Helicobacter while high-dose LipoLLA showed a somewhat superior efficacy. Since bactericidal efficacy was not the objective of this study, it was not further investigated with different doses of drugs or quantified.
H. pylori infection itself was reported to alter the diversity of the gastric microbiota,28,34,35 in spite of reports stating otherwise.36 Either way, in comparison with triple therapy, LipoLLA was shown to have a better bactericidal activity against H. pylori in earlier studies, while it demonstrated better microbiota stability than the triple therapy in the present study.
Conclusion
In summary, our study investigated the impact of LipoLLA, the newly invented anti-H. pylori nanomedicine, on gastrointestinal microbiota in mice for the first time using the high-throughput sequencing technique and compared it with that of the standard triple therapy. The results showed that LipoLLA poses much less effect on the composition and diversity of gastrointestinal microbiota than triple therapy and therefore has less side effects. Much attention has been paid to the alteration of microbiota via antibiotics since it is related to a variety of diseases. The findings in this study could shed light on a new effective way to eliminate H. pylori. Further investigations with larger sample size and high-dose gradient are warranted.
Disclosure
The authors report no conflicts of interest in this work.
References
Nilsson HO, Pietroiusti A, Gabrielli M, Zocco MA, Gasbarrini G, Gasbarrini A. Helicobacter pylori and extragastric diseases – other Helicobacters. Helicobacter. 2005;10(Suppl 1):54–65. | ||
World Gastroenterology Organisation. World Gastroenterology Organisation global guideline: Helicobacter pylori in developing countries. J Clin Gastroenterol. 2011;45(5):383–388. | ||
Thamphiwatana S, Gao W, Pornpattananangkul D, et al. Phospholipase A2-responsive antibiotic delivery via nanoparticle-stabilized liposomes for the treatment of bacterial infection. J Mater Chem B. 2014;2(46):8201–8207. | ||
[No authors listed]. Current European concepts in the management of Helicobacter pylori infection. The Maastricht consensus report. Gut. 1997;41(1):8–13. | ||
Oh B, Kim BS, Kim JW, et al. The effect of probiotics on gut microbiota during the Helicobacter pylori eradication: randomized controlled trial. Helicobacter. 2016;21(3):165–174. | ||
Urgesi R, Cianci R, Riccioni ME. Update on triple therapy for eradication of Helicobacter pylori: current status of the art. Clin Exp Gastroenterol. 2012;5:151–157. | ||
Camargo MC, Garcia A, Riquelme A, et al. The problem of Helicobacter pylori resistance to antibiotics: a systematic review in Latin America. Am J Gastroenterol. 2014;109(4):485–495. | ||
Ubeda C, Pamer EG. Antibiotics, microbiota, and immune defense. Trends Immunol. 2012;33(9):459–466. | ||
Lange K, Buerger M, Stallmach A, Bruns T. Effects of antibiotics on gut microbiota. Dig Dis. 2016;34(3):260–268. | ||
Young VB. The intestinal microbiota in health and disease. Curr Opin Gastroenterol. 2012;28(1):63–69. | ||
D’Argenio V, Salvatore F. The role of the gut microbiome in the healthy adult status. Clin Chim Acta. 2015;451(Pt A):97–102. | ||
Frohlich EE, Frohlich E. Cytotoxicity of nanoparticles contained in food on intestinal cells and the gut microbiota. Int J Mol Sci. 2016;17(4):509. | ||
Marchesi JR, Adams DH, Fava F, et al. The gut microbiota and host health: a new clinical frontier. Gut. 2016;65(2):330–339. | ||
Obonyo M, Zhang L, Thamphiwatana S, Pornpattananangkul D, Fu V, Zhang L. Antibacterial activities of liposomal linolenic acids against antibiotic-resistant Helicobacter pylori. Mol Pharm. 2012;9(9):2677–2685. | ||
Faghri J, Poursina F, Moghim S, et al. Morphological and bactericidal effects of different antibiotics on Helicobacter pylori. Jundishapur J Microbiol. 2014;7(1):e8704. | ||
Jung SW, Thamphiwatana S, Zhang L, Obonyo M. Mechanism of antibacterial activity of liposomal linolenic acid against Helicobacter pylori. PLoS One. 2015;10(3):e0116519. | ||
Thamphiwatana S, Gao W, Obonyo M, Zhang L. In vivo treatment of Helicobacter pylori infection with liposomal linolenic acid reduces colonization and ameliorates inflammation. Proc Natl Acad Sci U S A. 2014;111(49):17600–17605. | ||
Williams K, Milner J, Boudreau MD, Gokulan K, Cerniglia CE, Khare S. Effects of subchronic exposure of silver nanoparticles on intestinal microbiota and gut-associated immune responses in the ileum of Sprague-Dawley rats. Nanotoxicology. 2015;9(3):279–289. | ||
Pietroiusti A, Magrini A, Campagnolo L. New frontiers in nanotoxicology: gut microbiota/microbiome-mediated effects of engineered nanomaterials. Toxicol Appl Pharmacol. 2016;299:90–95. | ||
Liu L, Li Y, Li S, et al. Comparison of next-generation sequencing systems. J Biomed Biotechnol. 2012;2012:251364. | ||
Wu X, Zhang H, Chen J, et al. Comparison of the fecal microbiota of dholes high-throughput Illumina sequencing of the V3–V4 region of the 16S rRNA gene. Appl Microbiol Biotechnol. 2016;100(8):3577–3586. | ||
Obonyo M, Sabet M, Cole SP, et al. Deficiencies of myeloid differentiation factor 88, Toll-like receptor 2 (TLR2), or TLR4 produce specific defects in macrophage cytokine secretion induced by Helicobacter pylori. Infect Immun. 2007;75(5):2408–2414. | ||
Krzywinski M, Schein J, Birol I, et al. Circos: an information aesthetic for comparative genomics. Genome Res. 2009;19(9):1639–1645. | ||
An J, Lai J, Sajjanhar A, Batra J, Wang C, Nelson CC. J-Circos: an interactive Circos plotter. Bioinformatics. 2015;31(9):1463–1465. | ||
Adamsson I, Nord CE, Lundquist P, Sjöstedt S, Edlund C. Comparative effects of omeprazole, amoxycillin plus metronidazole versus omeprazole, clarithromycin plus metronidazole on the oral, gastric and intestinal microflora in Helicobacter pylori-infected patients. J Antimicrob Chemother. 1999;44(5):629–640. | ||
Jernberg C, Lofmark S, Edlund C, Jansson JK. Long-term impacts of antibiotic exposure on the human intestinal microbiota. Microbiology. 2010;156(Pt 11):3216–3223. | ||
Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature. 2012;486(7402):207–214. | ||
Ianiro G, Molina-Infante J, Gasbarrini A. Gastric microbiota. Helicobacter. 2015;20(Suppl 1):68–71. | ||
Jakobsson HE, Jernberg C, Andersson AF, Sjolund-Karlsson M, Jansson JK, Engstrand L. Short-term antibiotic treatment has differing long-term impacts on the human throat and gut microbiome. PLoS One. 2010;5(3):e9836. | ||
Pallav K, Dowd SE, Villafuerte J, et al. Effects of polysaccharopeptide from Trametes versicolor and amoxicillin on the gut microbiome of healthy volunteers: a randomized clinical trial. Gut Microbes. 2014;5(4):458–467. | ||
Bik E, Eckburg P, Gill S, et al. Molecular analysis of the bacterial microbiota in the human stomach. Proc Natl Acad Sci U S A. 2006;103(3):732–737. | ||
Dicksved J, Lindberg M, Rosenquist M, Enroth H, Jansson JK, Engstrand L. Molecular characterization of the stomach microbiota in patients with gastric cancer and in controls. J Med Microbiol. 2009;58(Pt 4):509–516. | ||
Delgado S, Cabrera-Rubio R, Mira A, Suarez A, Mayo B. Microbiological survey of the human gastric ecosystem using culturing and pyrosequencing methods. Microb Ecol. 2013;65(3):763–772. | ||
Andersson AF, Lindberg M, Jakobsson H, Backhed F, Nyren P, Engstrand L. Comparative analysis of human gut microbiota by barcoded pyrosequencing. PLoS One. 2008;3(7):e2836. | ||
Li TH, Qin Y, Sham PC, Lau KS, Chu KM, Leung WK. Alterations in gastric microbiota after H. pylori eradication and in different histological stages of gastric carcinogenesis. Sci Rep. 2017;7:44935. | ||
Khosravi Y, Dieye Y, Poh BH, et al. Culturable bacterial microbiota of the stomach of Helicobacter pylori positive and negative gastric disease patients. Sci World J. 2014;2014:610421. | ||
Edgar RC. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat Methods. 2013;10(10):996–998. | ||
Edgar RC, Haas BJ, Clemente JC, Quince C, Knight R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics. 2011;27(16):2194–2200. | ||
Schloss PD, Westcott SL, Ryabin T, et al. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol. 2009;75(23):7537–7541. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.