")
Back to Journals » Journal of Inflammation Research » Volume 15
The Contrasting Seasonality Patterns of Some Cancer-Types and Herpes Zoster Can Be Explained by a Binary Classification of Immunological Reactions
Authors Elkoshi Z
Received 3 October 2022
Accepted for publication 1 December 2022
Published 15 December 2022 Volume 2022:15 Pages 6761—6771
DOI https://doi.org/10.2147/JIR.S392082
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Zeev Elkoshi
Research and Development Department, Taro Pharmaceutical Industries Ltd, Haifa, Israel
Correspondence: Zeev Elkoshi, Email [email protected]
Abstract: A binary classification of the pathogenic immune reactions as anti-inflammatory high-Treg reactions or pro-inflammatory low-Treg reactions explains both the relatively low incidence rate of several types of cancer, and the relatively high incidence rate of herpes zoster cases diagnosed in the summer compared to cases diagnosed in the winter (in regions with temperate climate). This binary model also elucidates the longer survival of cancer patients diagnosed during the summer compared to these diagnosed in the winter. The three key elements of this explanation are: (a) the effect of sunlight on Treg production; (b) the evolvement of cancer from a low-Treg condition at early stage, to a high-Treg condition at advanced stage, and (c) the evolvement of herpes zoster from a high-Treg condition at pre-exudative stage to a low-Treg condition at acute exudative stage. A significant proportion of indolent tumors at the time of diagnosis (> 20%) is a prerequisite for a beneficial effect of sunlight on cancer incidence rate and prognosis. This prerequisite restricts the beneficial effect of diagnosis during summer to certain types of cancer. Clinical implication: the prognosis of early stage tumors may be improved by a course of corticosteroid (or other immunosuppressant) treatment.
Keywords: cancer, herpes zoster, Treg cells, vitamin D, UV radiation, incidence rate, cancer prognosis
Introduction
A binary model for the classification of chronic diseases was presented in an earlier paper.1 The model was based on the division of chronic diseases into two classes: high-Treg class versus low-Treg class, representing diseases with high or low regulatory T cell (Treg) activities, as drivers of pathogenicity. This division was found useful in explaining the association between certain pathogens and certain chronic diseases and in predicting the efficacy or inefficacy of immune-modulating drugs (such as corticosteroids) in the treatment of different chronic diseases. It also explained why certain specific pathogens are involved in coinfections, and why some pathogens may alleviate cancer, while others may promote it.1 In addition, the model elucidated why lymphomas and autoimmune diseases may be treated by the same immune modulating drugs.2 The model was also found helpful in assessing the efficacy or inefficacy of several immune-modulating drugs in treating COVID-19.3,4 From the mechanistic point of view, it was demonstrated that high-Treg diseases are driven by a group of anti-inflammatory kinases while low-Treg diseases are driven by a separate group of pro-inflammatory kinases.5
Several studies conducted in Scandinavia, Northern England, and North America reported lower incidence rates (subsequently referred to simply as, incidence) and better prognosis of some types of cancer when diagnosed during the summer as compared to the winter months.6–9 These observations are commonly explained by the effect of Vitamin D (VitD), which is more extensively produced by skin keratinocytes during summer due to a higher exposure to ultraviolet (UV) radiation. Vitamin D exerts direct anti-cancer effect,10 and immunosuppressive effect.11 While the anti-cancer activity of VitD may explain the aforementioned seasonal effects of the period of diagnosis, the role of the immunosuppressive effect of VitD in it is not clear. It is unclear: (a) why immunosuppression lowers the incidence of cancer (intuitively, it should raise it) and (b) how immunosuppression during the period of diagnosis (but not later) improves survival. Moreover, it is unclear why certain cancer-types are affected by higher exposure to UV radiation while others are not. The binary classification model of immune responses elucidates these puzzles. In contrast to cancer, a summer peak of new herpes zoster (HZ) cases was reported in a US study.12 This peak is also explained by the binary model. A clinical implication regarding early cancer treatment is drawn.
The Seasonal Variation in Cancer Incidence Rate and Prognosis
Several works describe seasonal variations in the incidence rate of some types of cancer. Most of these observations were reported in the Nordic countries, where the difference in the number of daylight hours between summer and winter is significant. Lambe et al6 used data from the Swedish Cancer Register 1987–1996 to evaluate seasonal trends in the diagnosis of cancer. Prostate cancer, breast cancer, thyroid cancer and melanoma were the only types of cancer for which a deviation from a uniform distribution throughout the year could be observed. The incidence of melanoma cases was minimal in winter while the incidence of prostate, breast and thyroid cancer was minimal in July. McNally et al8 analyzed data on all cancer cases diagnosed in children in Northern England during the period 1968–2005. In girls, lymphoma incidence peaked in March and Hodgkin lymphoma in January. In boys, a peak in osteosarcoma was observed in October.
Many studies have demonstrated a relationship between the season of cancer diagnosis and patient survival. For example, analysis of 5-year survival data of 19,204 female breast cancer, 6740 colorectal cancer, and 1644 prostate cancer cases, extracted from the US National Cancer Institute data, revealed that, at all latitudes, female breast cancer patients exhibited improved survival when diagnosed in the summer as compared to the winter. Colorectal cancer and prostate cancer demonstrated a similar seasonal pattern, when the analysis was restricted to northern regions of the US.9 Similarly, an analysis of 115,096 cases of breast, colon, or prostate cancer, diagnosed between 1964 and 1992 in Norway, revealed that diagnosis during summer and fall was associated with the lowest risk of cancer death.7 Analysis of data on all new cases of lung cancer diagnosed in Norway between 1964 and 2000 demonstrated approximately 15% lower cases of fatality for young male patients diagnosed during autumn versus winter. Young females diagnosed during the summer exhibited a trough in the relative risk of death, but this result was not statistically significant (p = 0.2).13 In addition, Norwegian patients residing in regions with higher UV radiation showed a better prognosis than those residing in regions with lower UV radiation.13 In a recent analysis of breast, prostate, colorectal and lung cancer data from the Cancer Registry of Zurich (Switzerland), a diagnosis in summer and/or autumn was associated with improved survival in all the sites combined for both sexes, and in colorectal and breast cancers, in women.14 An exception to these results was reported in one Swedish study of breast and prostate cancers.15 The lower incidence and better survival of patients diagnosed in summer are commonly explained by the higher production rate of vitamin D during summer, due to the higher exposure to UV radiation.14,16
Several direct anti-cancer effects of VitD are reported in the literature.10 Additionally, the suppressive effect of VitD on the immune system is well documented.17 UV-induced immunosuppression by VitD-independent pathways was also demonstrated.11 Nevertheless, none of these reports explains why the immunosuppressive effect of UV radiation lowers the incidence of cancer, and why it pertains specifically to the period of diagnosis. Likewise, none of these reports clarifies why only certain types of cancers are affected by UV radiation. The binary model of chronic diseases1 elucidates these points.
Cancer starts as a low-Treg pro-inflammatory disease and progresses over the time into a high-Treg anti-inflammatory disease.18–21 Figure 1 describes schematically the time dependence of Tregs, active natural killer (NK) cells, and active CD8+ T cells levels within the tumor microenvironment (TME) of hepatocellular carcinoma tumor induced in mice.22 UV radiation promotes a high-Treg response.23 UV light (mainly UVB) induces immunosuppression by several modes of action: (a) by the production of vitamin D, which promotes Treg generation;16 (b) by the production of nitric oxide and cis-urocanic acid, which enhances Treg generation;11 (c) by inducing the migration of dendritic cells (DCs) from irradiated skin to draining lymph nodes where DCs activate Tregs.11 The resulting high-Treg anti-inflammatory response ameliorates the low-Treg pro-inflammatory TME that drives early cancers. At an early stage, internal cancers are indolent, replicate slowly and are sensitive to the direct anti-cancer effect of TGFβ.2 Importantly, the probability of detecting indolent tumors at the time of diagnosis is relatively high in all the cancer types reported to display a better prognosis when diagnosed in summer. At the time of diagnosis, indolence comprises 33–40% of prostate cancer tumors (indolence = Gleason Score 1–2),24 20% of non-squamous cell lung cancer (NSCLC) tumors (NSCLC stages I, II)25 and about 90% of thyroid cancer tumors.26 In situ or localized colon cancer comprises about 40% of colon cancer tumors diagnosed in patients 30–60 years old.27 It is also reported that at the time of diagnosis, indolence comprises 2–35% of breast cancer tumors (at 80% test sensitivity),28 however, the number is probably closer to 30% since about 15% of overdiagnosis in breast cancer was reported in a recent large US study.29
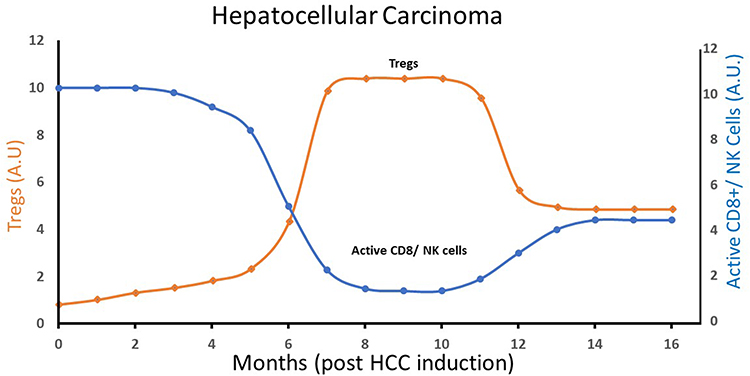 |
Figure 1 A schematic plot of Tregs and inflammatory cells (active CD8+ T cells/active NK cells) levels in hepatocellular carcinoma (HCC) tumor microenvironment (TME), as a function of time (months post HCC induction in a mouse model). Based on data from Nguyen PHD, Wasser M, Tan CT, et al. Trajectory of immune evasion and cancer progression in hepatocellular carcinoma. Nat Commun. 2022;13(1):1441. doi:10.1038/s41467-022-29122-w.22 The Treg level attains a maximum during the intermediate period (around 8 months post HCC induction), while active cellular immunity is at its lowest level during this period. Abbreviations: Tregs, regulatory T cells; NK cells, natural killer cells; HCC, hepatocellular carcinoma. |
Due to the relatively high fraction of indolent low-Treg tumors at the time of diagnosis, the UV induced high-Treg reaction exerts an anti-cancer effect in these cancer types. The binary model of chronic diseases therefore explains the seasonal variation in the incidence rate and prognosis, of cancer types characterized by a significant fraction of early tumors at workup. By the same rationale, sunlight is not expected to benefit advanced cancers, since the TME of advanced cancer is a high-Treg anti-inflammatory milieu.
The beneficial effect of VitD on lung cancer prognosis further supports the hypothesis that this effect is related to early cancer only: more than 85% of NSCLC tumors can be classified histologically as squamous cell carcinoma (SCC) or adenocarcinoma.30 Adenocarcinoma has a significantly higher stage-specific 5-year overall survival rate compared to SCC.31 An analysis of a lung cancer screening study conducted in the US revealed that about 95% of indolent cancer tumors detected were adenocarcinomas by histology.32 Lung adenocarcinoma is more prevalent in young patients compared to elderly patients while the opposite is true for SCC.30 If UV radiation affects mainly less aggressive cancers (adenocarcinoma in this case), the beneficial effect of lung cancer diagnosis during the summer on the prognosis must be more pronounced in young compared to elderly patients. Indeed, as mentioned above, a Norwegian lung cancer study demonstrated a lower mortality of young male patients diagnosed in the autumn (compare to the winter), but not of elderly patients.13
In contrast to internal cancers, skin cancers are triggered by direct UV radiation, as a consequence of UV-induced DNA damage. These DNA lesions may turn mutagenic if damaged cells are not disposed by apoptosis or if repair process fails, leading to the activation of proto-oncogenes or the inactivation of anti-oncogenes.33 However, skin cancers are also promoted by UV-induced immunosuppression, as was shown in the pioneering work of Kripke and Fisher.34 This may explain the correlation between skin cancer incidence and annual solar UV exposure dose.35 Congruently, the use of corticosteroids (CS), and other immunosuppressive drugs, increases the risk of melanoma36 and non-melanoma skin cancers.37,38 Another study showed a minor increase in the risk of non-melanoma skin cancers following CS use, but the effect was statistically non-significant.39 Several studies demonstrated an improved melanoma prognosis when diagnosed in the summer [Ref. 14 and references therein]; a large recent Australian study revealed about 60% in situ melanoma tumors (low-Treg tumors) at diagnosis.40
The Seasonal Variation in Herpes Zoster Incidence Rate
Several studies have shown that both herpes simplex virus (HSV) infection and anti-HSV vaccination induce a Treg reaction that suppresses virus-specific CD8+ T cell activation.41–43 Using an ocular murine model, Yu et al44 were able to demonstrate that this induced suppression is critical for the expression of latency genes which drive dormancy of the HSV-1 virus. At the same time, the authors showed that stress-induced corticosteroids trigger Treg reaction which reactivates ocular HSV-1 virus from its latency. The last observation is in agreement with the well-known correlation between stress and the recurrence of HSV-1 infection.45
Varicella zoster virus (VZV), also known as human herpesvirus 3, triggers chickenpox (varicella) following a primary infection by the virus. Chickenpox is an acute infection characterized by skin rash that forms small, itchy blisters that usually fade away after 5–7 days. Chickenpox commonly affects children and young adults. The virus invades sensory neurons, where it may stay dormant for years, and reactivates in later life, resulting in herpes zoster (shingles). Shingles are manifested by a painful skin rash with blisters usually localized in 1–2 adjacent dermatomes. In most cases, the rash heals within two to four weeks, however, some people develop postherpetic neuralgia which can last for months or years.46 It is useful to define three stages in the development of HZ: a pre-eruptive stage, an acute exudative stage, and a chronic stage.47
Primary VZV infection, chickenpox, is controlled by the immune response, just like primary HSV-1 infection. VZV-specific B and T cells suppress the primary infection and secure the latency of the virus. Humoral immunity has a special role in ameliorating or even preventing primary varicella. The involvement of the innate immunity in the pathogenesis of VZV infection, however, is not clear cut, but the severity of chickenpox is aggravated in individuals with compromised invariant natural killer T (iNKT) cell responses.48 Blood levels of inflammation-markers in unvaccinated children with active chickenpox correlated inversely with chickenpox severity.49 Since an anti-inflammatory reaction drives the disease, primary VZV infection (chickenpox) may be considered a high-Treg condition during the initial phase of the disease (the term high-Treg, which was reserved to chronic conditions in earlier publications, is used here to describe the early stage of an acute condition as well).
The present work suggests that the secondary VZV infection, herpes zoster, progresses from an anti-inflammatory high-Treg condition in the pre-eruptive stage, to a pro-inflammatory low-Treg condition in the acute exudative stage. This is presented schematically by Figure 2.
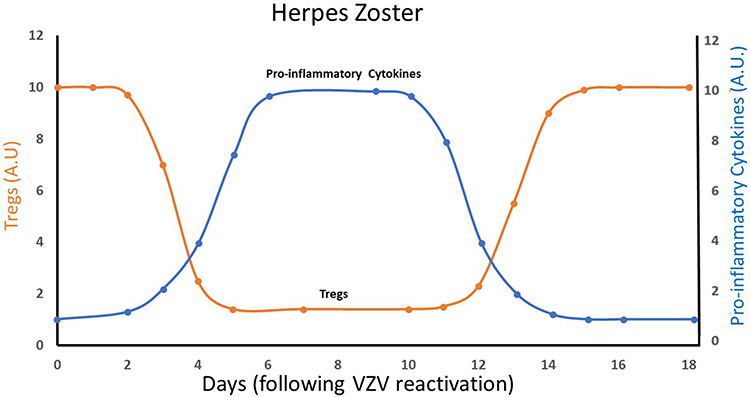 |
Figure 2 A schematic plot of circulating Tregs and pro-inflammatory cytokines levels in an acute herpes zoster infection, as a function of time following VZV reactivation. The first phase (days 1–2) correlates with the pre-eruptive stage, the second phase (days 3–12) correlates with the acute exudative stage and the last phase (starts about day 13) correlates with the resolution stage. Some patients, however, develop chronic postherpetic neuralgia, which may last for years. Abbreviations: Tregs, regulatory T cells; VZV, varicella zoster virus. |
In the pre-eruptive stage, Treg cells reactivate the virus by a suppression of the specific immune response that controls the virus latency (similar to their role in reactivating HSV-1). In the acute exudative stage, the replication of VZV in the dorsal root ganglia results in inflammatory neural damage.50 This inflammatory low-Treg process is mediated by inflammatory cytokines via toll-like receptor 251 and by the formation of the NLRP3 inflammasome, which catalyzes the processing of IL-1β by caspase-1 enzyme activity in innate immune cells.52 Indeed, serum levels of homocysteine (Hcy) and C-reactive protein (CRP), two markers of inflammation, were higher in HZ patients with active disease compared to healthy controls.53
The high-Treg character of initial chickenpox and of the pre-eruptive stage of herpes zoster is reflected by the effect of CS on the initiation of these diseases. Corticosteroids promote a high- Treg reaction of the immune system.54 As a high-Treg trigger, CS are expected to propel the onset of VZV primary infection (chickenpox) and secondary infection (HZ), since both are high-Treg conditions at their initiation. Indeed, CS use increases the risk of chickenpox55,56 and HZ.57 In addition, the use of CS prior to live attenuated varicella vaccination and live attenuated zoster vaccination contributes to serious adverse events.58
On the other hand, “paradoxically”, CS are helpful in ameliorating the acute exudative stage of HZ. German guidelines for the diagnosis and treatment of herpes zoster and postherpetic neuralgia,59 recommend the use of CS for the treatment of HZ-related ailments such as: vasculitis, retinal necrosis, herpes zoster oticus, and facial nerve involvement, complications which typically develop during the acute exudative stage. Several studies have found CS effective in the treatment of adult varicella pneumonia, another inflammatory complication of HZ, when used as an adjuvant to anti-viral therapy.60–62
In fact, recognizing that exudative HZ is a low-Treg condition, the beneficial effect of CS on exudative HZ is predicted by the binary model.
The effect of CS on pain associated with postherpetic neuralgia is however controversial.63 In line with this, it has been proposed that inflammation is not the main driver of pain in postherpetic neuralgia.64
Chickenpox is a self-resolved acute inflammation with a short duration of the inflammatory phase. Corticosteroid intervention may interfere with the resolution process, and CS therapy is not recommended in chickenpox.
Due to the intensified UV-exposure during summer, and the ensuing high-Treg reaction, the risk of HZ in countries with a temperate climate is expected to rise in summer, since HZ starts as a high-Treg disease. In agreement with this expectation, a summer peak in new HZ cases was observed in a large dataset analysis of 513,911 patients from across the US.12 On the other hand, herpes zoster ophthalmicus, an inflammatory low-Treg condition that involves the eye or the surrounding area, showed a peak in October, but this result was not statistically significant.12
Discussion
It is important to understand that the terms high-Treg and low-Treg do not refer to the inflammatory condition per se. Instead, they describe the pathogenic effect of inflammation on the state of disease: inflammation can either control the disease or promote it. In the first case, higher levels of Tregs are pathogenic (establishing a high-Treg condition), in the second case lower levels of Tregs are pathogenic (establishing a low-Treg condition).
It should also be realized that Tregs regulate the function of a myriad of immune cells: CD4+ T cells, CD8+ T cells, B cells, NK cells, NKT cells, neutrophils, macrophages and dendritic cells.65 Hence, a change in Treg activity leads to a change in the function of many immune cells. A high-Treg condition therefore results in a wide-range immune suppression, while a low-Treg condition results in a wide-range of immune activation.
Next, the pathophysiology of cancer and HZ infection in relation to UV exposure is discussed. The possible involvement of other environmental drivers that may affect seasonality patterns is examined as well.
Cancer
Cancer starts as an inflammatory low-Treg condition. Macrophages and neutrophils are potent producers of reactive oxygen and nitrogen species (ROS and RNS). These inflammatory species may induce mutations in cancer promoting genes (oncogenes) which trigger cancer. For example, chronic inflammation associated with intestinal bowel disease results in an accumulation of mutations in the tumor suppressor Tp53 gene (and in other cancer-related genes) in intestinal epithelial cells. Moreover, inflammatory cytokines such as IL-6, TNFα, and IL-1β can induce epigenetic changes by inactivating mutations in tumor suppressors (anti-oncogenes) and by activating mutations in oncogenes, with the net result of tumor initiation. In addition to initiating cancer, inflammation also promotes cancer cell proliferation, for example, by inactivation of nuclear factor-kappa B (NFκB) in immune cells (the nuclear factor NFκB is a suppressor of cancer cell survival and proliferation).66
As cancer advances, the immune reaction gradually changes from a low-Treg into a high-Treg condition. High levels of regulatory cells suppress the immune system and further drive the disease.18–21
For a discernable beneficial effect of sunlight on cancer, the proportion of low-grade tumors at the time of diagnosis must not be too low. In fact, all cancer types that reported to have a lower incidence or a better prognosis when diagnosed during the summer (compared to the winter), exhibit at least 20% indolent tumors at diagnosis, as delineated above.
The seasonality of cancer may be related to the seasonality of the pathogens that induce this cancer. For example, the authors of a recent acute myeloid leukemia (AML) epidemiological study raised the possibility that the monthly variation in AML incidence follows the variation in the rates of infectious agents capable of precipitating or accelerating the course of AML.67 This study analyzed data obtained in Spain, where the differences in daylight hours between summer and winter are (relatively) moderate, ensuing a moderate seasonal variation in UV exposure. This renders the effect of infections more conspicuous. However, in regions with a high seasonal variation in the number of daylight hours (for example, the number of daylight hours in Oslo, Norway, varies from 19 in June to 6 in December), the seasonal effect of UV exposure at diagnosis on the course of cancer becomes more prominent. Except for the effect of UV radiation, other causes for the lower incidence of cancer diagnosis in summer are possible. For example, the tradition of vacationing from mid-June to mid-August in some countries may result in a postponement of clinic visits of symptomatic patients during these months. In addition, many elective routine procedures in health care (such as mammography) are less available during the summer in some countries, for example in Sweden.15 Even though other causes cannot be excluded, seasonal variations in UV radiation probably contribute to the observed seasonal changes in cancer incidence and to the effect of the period of diagnosis on cancer prognosis. This hypothesis is supported by the following evidence: (a) UV radiation induces the production of VitD by skin keratinocytes;13 (b) in the Nordic countries serum VitD status exhibits a strong seasonal variation that parallels the seasonal change in UVB radiation that reaches the earth;13 (c) vitamin D can promote the development and function of Tregs;17 (d) Treg blood levels are significantly higher in summer compared to winter in temperate climate regions;68 (e) Tregs suppress early cancer;69 (f) low VitD levels are associated with increased cancer incidence and mortality70 (g) vitamin D intake reduces cancer risk;71 (h) colorectal cancer and prostate cancer seasonal variation in the US depend on the latitude;9 (i) UV radiation may induce immunosuppression by VitD-independent pathways.11
As a clinical implication, this work suggests that early stage cancer prognosis may be improved by a course of CS (or other immunosuppressive drugs) treatment. Corticosteroids may be administered during the period between cancer diagnosis and tumor resection, during a “wait and watch” period, or as an adjuvant to chemotherapy or radiation therapy of early stage tumors.
Herpes Zoster Infection
Some infections start as low-Treg inflammatory conditions. It is pertinent to ask the question: in what way can intensified inflammation (a low-Treg reaction) aggravate infection?
It seems that some infections involve intensified non-specific response (innate immunity) accompanied by an impaired specific response (adaptive immunity) against the pathogen. Highly activated innate immunity increases the level of pro-inflammatory cytokines (and generates inflammation or a low-Treg state) while defective specific response aggravates the infection.
For example, using a mouse model, Takahashi et al72 have shown that cigarette smoke induces an inflammation characterized by increased levels of TNFα and IL-6. The inflammation was mediated by IKKβ and JNK1enzymes expressed on myeloid cells (macrophages and neutrophils). Importantly, exposure of mice to Non-typeable Haemophilus influenzae (NTHi) bacteria, following a chronic exposure to tobacco smoke, elicited a mixed reaction of high proinflammatory myeloid reaction, side-by-side with an impaired pathogen-specific lymphoid reaction.73 The impaired specific response hampered NTHi clearance rate in spite of an enhanced non-specific immune response (increased levels of IL-1β, TNFα, IL-6, and an increased number of neutrophils).73
Another example: investigations confirmed the ability of 1,25-dihydroxy vitamin D to induce expression of cathelicidin (an anti-microbial peptide) in myeloid cell lines.74,75 Other studies demonstrated the role of VitD in activating cathelicidin expressed by macrophages, in this way enhancing the human antimicrobial activity against Mycobacterium tuberculosis.76,77 At the same time, 1,25-dihydroxy vitamin D suppresses the activity and production of immunoglobulins by mononuclear cells78,79 and T cells.80–84 In addition, activated VitD suppresses the secretion of IL-12 and IL-23 by dendritic cells, which affects the differentiation of T cells into Th1 and Th17 cell, respectively.85 This subject is thoroughly reviewed by Ao et al.86
It is the belief of the author of this article, that a combination of a highly activated innate response and an impaired adaptive response is shared by other low-Treg infections.
Some infections are initiated by an anti-inflammatory high-Treg reaction. A secondary VZV infection (HZ) is an example. Contrary to cancer, HZ is elicited by a high-Treg response: an enhanced specific immune response is needed to control the dormancy of the virus in sensory neurons.48 Regulatory T cells induced by endogenous or exogenous CS, or by exposure to UV radiation, reactivate the virus and trigger the disease by the suppression of the specific immune response that maintains its dormancy57,58 just as in the case with HSV-1.44 Later on, a pro-inflammatory low-Treg response develops within the branches of the sensory trigeminal nerve where the virus multiplies, leading to sore inflamed lesions. These lesions may be treated with CS.59
The seasonal variation of infections depends on multiple environmental drivers in addition to UV radiation. Among these are: vector (usually an insect) seasonality, seasonality in nonhuman animal host (livestock), seasonal flare-up/symptoms and/or remission/latency, seasonal coinfection, seasonal exposure and/or behavior and/or contact rate.87 Secondary VZV infection (HZ) is not transmitted by a vector or carried by nonhuman animal host, and is only modestly contagious. A dependence of VZV seasonality on coinfection cannot be excluded, however, and coinfections of VZV and either Epstein–Barr virus (EBV), Monkeypox virus, or SARS-CoV-2 are documented.88–90 However, the presence of Monkeypox virus and SARS-CoV-2 was not reported in the USA when the demographic data used by Berlinberg et al12 was collected (2006–2017), and coinfection of VZV and EBV is rare (only 2 out of 16 patients with EBV coinfection were found positive to VZV).88 Therefore, in the study by Berlinberg et al,12 the effect of sunlight exposure was not disguised by other environmental drivers.
It should be added that chickenpox and HZ differ in their modes of transmission. While chickenpox spreads by both, direct contact with infected skin lesions and inhalation of airborne respiratory droplets,91,92 HZ spreads almost exclusively by direct contact with infected vesicular fluid.93 Even though chickenpox starts as a high-Treg disease, new chickenpox cases in northern European countries attain their peak in winter.94 Winter is a season when people in the Nordic areas, especially children, tend to gather within closed, poorly ventilated spaces (schools for example), resulting in the accelerated transmission of the virus. On the other hand, herpes zoster affects mainly adults, and airborne spread of HZ is (at best) limited. Therefore, in contrast to chickenpox, HZ summer peak arising from a higher exposure to UV radiation is not shifted to the winter season.
Summary
A lower incidence and better prognosis of some types of cancer when diagnosed in summer, as compared to winter, have been documented in temperate zones. These beneficial effects have been commonly attributed to an increased VitD production as a result of higher exposure to UV radiation during summer months, and to an anti-cancer and immunosuppressive effect of vitamin D. However, no answers were provided for the questions: Why does immunosuppression induce a decrease in the incidence of cancer? How immunosuppression at the time of diagnosis (but not later) improves prognosis? Why only certain cancer-types are affected? The binary model of immune reaction suggests an explanation for these puzzles. Cancer develops from a low-Treg condition at an early stage, to a high-Treg condition at advanced stages. It seems that VitD-induced immunosuppression slows down cancer progression during the early stage of cancer by counteracting this pro-inflammatory low-Treg condition. A relatively high percentage of early stage tumors at diagnosis is required for the beneficial effect of UV on tumor initiation and prognosis, and a high percentage of early stage tumors is common to cancer types that are affected by UV radiation. In contrast to cancer, herpes zoster starts as a high-Treg condition and develops into a low-Treg condition. The immunosuppressive effect of UV supports the anti inflammatory conditions that are necessary for HZ initiation, and therefore a peak in HZ new cases is observed during the summer period.
Clinical Implication
A course of corticosteroid treatment (or another immunosuppressive drug) during the early stage of cancer may improve survival. This applies only to indolent localized tumors. This drug treatment can take place in the period between cancer diagnosis and tumor resection, or during a “wait and watch” period. It can also be added as an adjuvant to chemotherapy or radiation therapy of early-stage tumors.
Disclaimer
The views and opinions expressed, and/or conclusions drawn, in this article are those of the author and do not necessarily reflect those of Taro Pharmaceutical Industries Ltd., its affiliates, directors or employees.
Acknowledgment
The author would like to thank Dr. Elad Lavee-laviad for his help in preparing the figures for this article.
Disclosure
Zeev Elkoshi is employed by Taro Pharmaceutical Industries Ltd.
References
1. Elkoshi Z. The binary classification of chronic diseases. J Inflamm Res. 2019;12:319333. doi:10.2147/JIR.S227279
2. Elkoshi Z. ”High Treg” inflammations promote (most) non-hematologic cancers while “Low Treg” inflammations promote lymphoid cancers. J Inflamm Res. 2020;13:209221. doi:10.2147/JIR.S249384
3. Elkoshi Z. The binary model of chronic diseases applied to COVID-19. Front Immunol. 2021;12:716084. doi:10.3389/fimmu.2021.716084
4. Elkoshi Z. SARS-CoV-2 omicron (B.1.1.529) variant: corticosteroids treatment/respiratory coinfection. Front Immunol. 2022;13:856072. doi:10.3389/fimmu.2022.856072
5. Elkoshi Z. The binary classification of protein kinases. J Inflamm Res. 2021;14:929–947. doi:10.2147/JIR.S303750
6. Lambe M, Blomqvist P, Bellocco R. Seasonal variation in the diagnosis of cancer: a study based on national cancer registration in Sweden. Br J Cancer. 2003;88(9):135860. doi:10.1038/sj.bjc.6600901
7. Robsahm TE, Tretli S, Dahlback A, Moan J. Vitamin D3 from sunlight may improve the prognosis of breast-, colon- and prostate cancer (Norway). Cancer Causes Control. 2004;15(2):149–158. doi:10.1023/B:CACO.0000019494.34403.09
8. McNally R, Basta N, James P, Craft A. Seasonal variation in birth and diagnosis of cancer in children and young people in Northern England, 1968–2005. Epidemiology. 2009;20:S18. doi:10.1097/01.ede.0000362231.71672.43
9. Ho A, Gabriel A, Bhatnagar A, Etienne D, Loukas M. Seasonality pattern of breast, colorectal, and prostate cancer is dependent on latitude. Med Sci Monit. 2014;20:818–824. doi:10.12659/MSM.890062
10. Trump DL, Aragon-Ching JB. Vitamin D in prostate cancer. Asian J Androl. 2018;20(3):244–252. doi:10.4103/aja.aja_14_18
11. Hart PH, Norval M, Byrne SN, Rhodes LE. Exposure to ultraviolet radiation in the modulation of human diseases. Annu Rev Pathol. 2019;14:55–81. doi:10.1146/annurevpathmechdis-012418-012809
12. Berlinberg EJ, Kim E, Deiner MS, Patterson C, Porco TC, Acharya NR. Seasonality of herpes zoster and herpes zoster ophthalmicus. J Clin Virol. 2020;126:104306. doi:10.1016/j.jcv.2020.104306
13. Porojnicu AC, Robsahm TE, Dahlback A, et al. Seasonal and geographical variations in lung cancer prognosis in Norway. Does vitamin D from the sun play a role? Lung Cancer. 2007;55(3):263–270. doi:10.1016/j.lungcan.2006.11.013
14. Hysaj O, Karavasiloglou N, Limam M, Wanner M, Korol D, Rohrmann S. Is season of diagnosis a predictor of cancer survival? Results from the Zurich Cancer Registry. Nutrients. 2022;14(20):4291. doi:10.3390/nu14204291
15. Holmberg L, Adolfsson J, Mucci L, et al. Season of diagnosis and prognosis in breast and prostate cancer. Cancer Causes Control. 2009;20(5):663–670. doi:10.1007/s10552-008-9279-6
16. Moan J, Lagunova Z, Bruland O, Juzeniene A. Seasonal variations of cancer incidence and prognosis. Dermatoendocrinol. 2010;2(2):55–57. doi:10.4161/derm.2.2.12664
17. Athanassiou L, Mavragani CP, Koutsilieris M. The immunomodulatory properties of vitamin D. Mediterr J Rheumatol. 2022;33(1):7–13. doi:10.31138/mjr.33.1.7
18. Gonzalez H, Hagerling C, Werb Z. Roles of the immune system in cancer: from tumor initiation to metastatic progression. Genes Dev. 2018;32(19–20):1267–1284. doi:10.1101/gad.314617.118
19. Domagala-Kulawik J, Osinska I, Hoser G. Mechanisms of immune response regulation in lung cancer. Transl Lung Cancer Res. 2014;3(1):15–22. doi:10.3978/j.issn.22186751.2013.11.03
20. Lim RJ, Liu B, Krysan K, Dubinett SM. Lung cancer and immunity markers. Cancer Epidemiol Biomarkers Prev. 2020;29(12):2423–2430. doi:10.1158/1055-9965.EPI200716
21. Anichini A, Perotti VE, Sgambelluri F, Mortarini R. Immune escape mechanisms in non small cell lung cancer. Cancers. 2020;12(12):3605. doi:10.3390/cancers12123605
22. Nguyen PHD, Wasser M, Tan CT, et al. Trajectory of immune evasion and cancer progression in hepatocellular carcinoma. Nat Commun. 2022;13(1):1441. doi:10.1038/s41467-022-29122-w
23. Schwarz T. 25 years of UV-induced immunosuppression mediated by T cells-from disregarded T suppressor cells to highly respected regulatory T cells. Photochem Photobiol. 2008;84(1):10–18. doi:10.1111/j.1751-1097.2007.00223.x
24. Ji G, Huang C, Song G, et al. Are the pathological characteristics of prostate cancer more aggressive or more indolent depending upon the patient age? Biomed Res Int. 2017;2017:1438027. doi:10.1155/2017/1438027
25. Walters S, Maringe C, Coleman MP, et al.; ICBP Module 1 Working Group. Lung cancer survival and stage at diagnosis in Australia, Canada, Denmark, Norway, Sweden and the UK: a population-based study, 2004–2007. Thorax. 2013;68(6):551–564. doi:10.1136/thoraxjnl-2012-202297
26. Hahn LD, Kunder CA, Chen MM, Orloff LA, Desser TS. Indolent thyroid cancer: knowns and unknowns. Cancers Head Neck. 2017;2:1. doi:10.1186/s411990160021-x
27. Abualkhair WH, Zhou M, Ahnen D, Yu Q, Wu XC, Karlitz JJ. Trends in Incidence of Early-Onset Colorectal Cancer in the United States Among Those Approaching Screening Age. JAMA Netw Open. 2020;3(1):e1920407. doi:10.1001/jamanetworkopen.2019.20407
28. Shen Y, Dong W, Gulati R, Ryser MD, Etzioni R. Estimating the frequency of indolent breast cancer in screening trials. Stat Methods Med Res. 2019;28(4):1261–1271. doi:10.1177/0962280217754232
29. Tanne JH. Breast cancer is overdiagnosed in one in six or seven cases, finds large US study. BMJ. 2022;376:o581. doi:10.1136/bmj.o581
30. Zhong W, Zhao J, Huang K, Zhang J, Chen Z. Comparison of clinicopathological and molecular features between young and old patients with lung cancer. Int J Clin Exp Pathol. 2018;11(2):1031–1035.
31. Van Herck Y, FeyaertsAlibhai S, Papamichael D, et al. Is cancer biology different in older patients? Lancet Heal Longev. 2021;2:e663–e677. doi:10.1016/S2666-7568(21)00179-3
32. Thalanayar PM, Altintas N, Weissfeld JL, Fuhrman CR, Wilson DO. Indolent, potentially inconsequential lung cancers in the Pittsburgh lung screening study. Ann Am Thorac Soc. 2015;12(8):1193–1196. doi:10.1513/AnnalsATS.201412-577OC
33. Cadet J, Douki T. Formation of UV-induced DNA damage contributing to skin cancer development. Photochem Photobiol Sci. 2018;17(12):1816–1841. doi:10.1039/c7pp00395a
34. Kripke ML, Fisher MS. Immunologic aspects of tumor induction by ultraviolet radiation. Natl Cancer Inst Monogr. 1978;50:179–183. PMID: 753974.
35. Moan J, Grigalavicius M, Baturaite Z, Juzeniene A, Dahlback A. North-South gradients of melanomas and non-melanomas: a role of vitamin D? Dermatoendocrinol. 2013;5(1):186–191. doi:10.4161/derm.23791
36. Berge LAM, Andreassen BK, Stenehjem JS, et al. Use of immunomodulating drugs and risk of cutaneous melanoma: a nationwide nested case-control study. Clin Epidemiol. 2020;12:1389–1401. doi:10.2147/CLEP.S269446
37. Jensen AØ, Thomsen HF, Engebjerg MC, et al. Use of oral glucocorticoids and risk of skin cancer and non-Hodgkin’s lymphoma: a population based case-control study. Br J Cancer. 2009;100(1):200–205. doi:10.1038/sj.bjc.6604796
38. Sørensen HT, Mellemkjaer L, Nielsen GL, Baron JA, Olsen JH, Karagas MR. Skin cancers and non-Hodgkin lymphoma among users of systemic glucocorticoids: a population-based cohort study. J Natl Cancer Inst. 2004;96(9):709–711. doi:10.1093/jnci/djh118
39. Baibergenova AT, Weinstock MA; VATTC Trial Group. Oral prednisone use and risk of keratinocyte carcinoma in non-transplant population. The VATTC trial. J Eur Acad Dermatol Venereol. 2012;26(9):1109–1115. doi:10.1111/j.1468-3083.2011.04226.x
40. Whiteman DC, Olsen CM, MacGregor S, et al. QSkin Study. The effect of screening on melanoma incidence and biopsy rates. Br J Dermatol. 2022;187(4):515–522. doi:10.1111/bjd.21649
41. Milman N, Zhu J, Johnston C, et al. In situ detection of regulatory T cells in human genital herpes simplex virus type 2 (HSV-2) reactivation and their influence on spontaneous HSV-2 reactivation. J Infect Dis. 2016;214(1):23–31. doi:10.1093/infdis/jiw091
42. Rouse BT, Sarangi PP, Suvas S. Regulatory T cells in virus infections. Immunol Rev. 2006;212:272–286. doi:10.1111/j.0105-2896.2006.00412.x
43. Toka FN, Suvas S, Rouse BT. CD4+ CD25+ T cells regulate vaccine-generated primary and memory CD8+ T-cell responses against herpes simplex virus type 1. J Virol. 2004;78(23):13082–13089. doi:10.1128/JVI.78.23.13082-13089.2004
44. Yu W, Geng S, Suo Y, et al. Critical role of regulatory T cells in the latency and stress-induced reactivation of HSV-1. Cell Rep. 2018;25(9):2379–2389.e3. doi:10.1016/j.celrep.2018.10.105
45. Padgett DA, Sheridan JF, Dorne J, Berntson GG, Candelora J, Glaser R. Social stress and the reactivation of latent herpes simplex virus type 1. Proc Natl Acad Sci U S A. 1998;95(12):7231–7235. doi:10.1073/pnas.95.12.7231
46. Cohen JI, Solomon CG. Clinical practice: herpes zoster. N Engl J Med. 2013;369(3):255–263. doi:10.1056/NEJMcp1302674
47. Patil A, Goldust M, Wollina U. Herpes zoster: a review of clinical manifestations and management. Viruses. 2022;14(2):192. doi:10.3390/v14020192
48. Laing KJ, Ouwendijk WJD, Koelle DM, Verjans GMGM. Immunobiology of varicellazoster virus infection. J Infect Dis. 2018;218(suppl_2):S68–S74. doi:10.1093/infdis/jiy403
49. Karadag Oncel E, Kara A, Celik M, Karahan S, Cengiz AB, Ceyhan M. Determination and clinical correlation of markers of inflammation in unvaccinated patients with varicella-zoster infection. Eur Rev Med Pharmacol Sci. 2013;17(15):2032–2039.
50. Jeon YH. Herpes zoster and postherpetic neuralgia: practical consideration for prevention and treatment. Korean J Pain. 2015;28(3):177–184. doi:10.3344/kjp.2015.28.3.177
51. Wang JP, Kurt-Jones EA, Shin OS, Manchak MD, Levin MJ, Finberg RW. Varicellazoster virus activates inflammatory cytokines in human monocytes and macrophages via Toll-like receptor 2. J Virol. 2005;79(20):12658–12666. doi:10.1128/JVI.79.20.1265812666.2005
52. Nour AM, Reichelt M, Ku CC, Ho MY, Heineman TC, Arvin AM. Varicella-zoster virus infection triggers formation of an interleukin-1β (IL-1β)-processing inflammasome complex. J Biol Chem. 2011;286(20):17921–17933. doi:10.1074/jbc.M110.210575
53. Oskay T, Keskin C, Özen M. Antioxidant and inflammatory biomarkers in herpes zoster. J Med Virol. 2022;94(8):3924–3929. doi:10.1002/jmv.27781
54. Bereshchenko O, Coppo M, Bruscoli S, et al. GILZ promotes production of peripherally induced Treg cells and mediates the crosstalk between glucocorticoids and TGF-β signaling. Cell Rep. 2014;7(2):464–475. doi:10.1016/j.celrep.2014.03.004
55. Dowell SF, Bresee JS. Severe varicella associated with steroid use. Pediatrics. 1993;92(2):223–228. doi:10.1542/peds.92.2.223
56. Hill G, Chauvenet AR, Lovato J, McLean TW. Recent steroid therapy increases severity of varicella infections in children with acute lymphoblastic leukemia. Pediatrics. 2005;116(4):e525–9. doi:10.1542/peds.2005-0219
57. Qian J, Banks E, Macartney K, Heywood AE, Lassere MN, Liu B. Corticosteroid use and risk of herpes zoster in a population-based cohort. Mayo Clin Proc. 2021;96(11):2843–2853. doi:10.1016/j.mayocp.2021.05.029
58. Price NB, Grose C. Corticosteroids contribute to serious adverse events following live attenuated varicella vaccination and live attenuated zoster vaccination. Vaccines. 2021;9(1):23. doi:10.3390/vaccines9010023
59. Gross GE, Eisert L, Doerr HW, et al. S2k guidelines for the diagnosis and treatment of herpes zoster and postherpetic neuralgia. J Dtsch Dermatol Ges. 2020;18(1):55–78. doi:10.1111/ddg.14013
60. Adhami N, Arabi Y, Raees A, Al-Shimemeri A, Ur-Rahman M, Memish ZA. Effect of corticosteroids on adult varicella pneumonia: cohort study and literature review. Respirology. 2006;11(4):437–441. doi:10.1111/j.1440-1843.2006.00870.x
61. Anwar SK, Masoodi I, Alfaifi A, Hussain S, Sirwal IA. Combining corticosteroids and Acyclovir in the management of varicella pneumonia: a prospective study. Antivir Ther. 2014;19(2):221–224. doi:10.3851/IMP2751
62. Mer M, Richards GA. Corticosteroids in life-threatening varicella pneumonia. Chest. 1998;114(2):426–431. doi:10.1378/chest.114.2.426
63. Kowalsky DS, Wolfson AB, Zehtabchi S. Corticosteroids for preventing postherpetic neuralgia after herpes zoster infection. Acad Emerg Med. 2019;26(6):686–687. doi:10.1111/acem.13661
64. Devor M. Rethinking the causes of pain in herpes zoster and postherpetic neuralgia: the ectopic pacemaker hypothesis. Pain Rep. 2018;3(6):e702. doi:10.1097/PR9.0000000000000702
65. Grover P, Goel PN, Greene MI. Regulatory T Cells: regulation of Identity and Function. Front Immunol. 2021;12:750542. doi:10.3389/fimmu.2021.750542
66. Greten FR, Grivennikov SI. Inflammation and cancer: triggers, mechanisms, and consequences. Immunity. 2019;51(1):27–41. doi:10.1016/j.immuni.2019.06.025
67. Sánchez-Vizcaíno F, Tamayo C, Ramos F, et al. Identification of seasonal variation in the diagnosis of acute myeloid leukaemia: a population-based study. Br J Haematol. 2022;198(3):545–555. doi:10.1111/bjh.18279
68. Lamikanra AA, Tsang HP, Elsiddig S, et al. TheMigratory properties and numbers of T regulatory cell subsets in circulation are differentially Influenced by season and are associated with vitamin D status. Front Immunol. 2020;11:685. doi:10.3389/fimmu.2020.00685
69. Elkoshi Z. Cancer and autoimmune diseases: a tale of two immunological opposites? Front Immunol. 2022;13:821598. doi:10.3389/fimmu.2022.821598
70. Giovannucci E, Liu Y, Rimm EB, et al. Prospective study of predictors of vitamin D status and cancer incidence and mortality in men. J Natl Cancer Inst. 2006;98(7):451–459. doi:10.1093/jnci/djj101
71. Chen P, Hu P, Xie D, Qin Y, Wang F, Wang H. Meta-analysis of vitamin D, calcium and the prevention of breast cancer. Breast Cancer Res Treat. 2010;121(2):469–477. doi:10.1007/s10549-009-0593-9
72. Takahashi H, Ogata H, Nishigaki R, Broide DH, Karin M. Tobacco smoke promotes lung tumorigenesis by triggering IKKbeta- and JNK1-dependent inflammation. Cancer Cell. 2010;17(1):89–97. doi:10.1016/j.ccr.2009.12.008
73. Lugade AA, Bogner PN, Thatcher TH, Sime PJ, Phipps RP, Thanavala Y. Cigarette smoke exposure exacerbates lung inflammation and compromises immunity to bacterial infection. J Immunol. 2014;192(11):5226–5235. doi:10.4049/jimmunol.1302584
74. Gombart AF, Borregaard N, Koeffler HP. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxy vitamin D3. FASEB J. 2005;19(9):1067–1077. doi:10.1096/fj.04-3284com
75. Yuk JM, Shin DM, Lee HM, et al. Vitamin D3 induces autophagy in human monocytes/macrophages via cathelicidin. Cell Host Microbe. 2009;6(3):231–243. doi:10.1016/j.chom.2009.08.004
76. Liu PT, Stenger S, Li H, et al. Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science. 2006;311(5768):1770–1773. doi:10.1126/science.1123933
77. Martineau AR, Wilkinson KA, Newton SM, et al. IFN-gamma- and TNF-independent vitamin D-inducible human suppression of mycobacteria: the role of cathelicidin LL-37. J Immunol. 2007;178(11):7190–7198. doi:10.4049/jimmunol.178.11.7190
78. Lemire JM, Adams JS, Sakai R, Jordan SC. 1 alpha,25-dihydroxy vitamin D3 suppresses proliferation and immunoglobulin production by normal human peripheral blood mononuclear cells. J Clin Invest. 1984;74(2):657–661. doi:10.1172/JCI111465
79. Linker-Israeli M, Elstner E, Klinenberg JR, Wallace DJ, Koeffler HP. Vitamin D(3) and its synthetic analogs inhibit the spontaneous in vitro immunoglobulin production by SLEderived PBMC. Clin Immunol. 2001;99(1):82–93. doi:10.1006/clim.2000.4998
80. Rigby WF, Stacy T, Fanger MW. Inhibition of T lymphocyte mitogenesis by 1,25 dihydroxy vitamin D3 (calcitriol). J Clin Invest. 1984;74(4):1451–1455. doi:10.1172/JCI111557
81. Lemire JM, Adams JS, Kermani-Arab V, Bakke AC, Sakai R, Jordan SC. 1,25 Dihydroxy vitamin D3 suppresses human T helper/inducer lymphocyte activity in vitro. J Immunol. 1985;134(5):3032–3035.
82. Tsoukas CD, Watry D, Escobar SS, et al. Inhibition of interleukin-1 production by 1,25-dihydroxy vitamin D3. J Clin Endocrinol Metab. 1989;69(1):127–133. doi:10.1210/jcem-69-1-127
83. Ikeda U, Wakita D, Ohkuri T, et al. 1α,25 Dihydroxy vitamin D3 and all-trans retinoic acid synergistically inhibit the differentiation and expansion of Th17 cells. Immunol Lett. 2010;134(1):7–16. doi:10.1016/j.imlet.2010.07.002
84. Joshi S, Pantalena LC, Liu XK, et al. 1,25 dihydroxy vitamin D(3) ameliorates Th17 autoimmunity via transcriptional modulation of interleukin-17A. Mol Cell Biol. 2011;31(17):3653–3669. doi:10.1128/MCB.05020-11
85. Berer A, Stöckl J, Majdic O, et al. 1,25-Dihydroxy vitamin D(3) inhibits dendritic cell differentiation and maturation in vitro. Exp Hematol. 2000;28(5):575–583. doi:10.1016/s0301-472x(00)00143-0
86. Ao T, Kikuta J, Ishii M. The effects of vitamin d on immune system and inflammatory diseases. Biomolecules. 2021;11(11):1624. doi:10.3390/biom11111624
87. Martinez ME, Kline KA. The calendar of epidemics: seasonal cycles of infectious diseases. PLoS Pathog. 2018;14(11):e1007327. doi:10.1371/journal.ppat.1007327
88. Weinberg A, Bloch KC, Li S, Tang YW, Palmer M, Tyler KL. Dual infections of the central nervous system with Epstein-Barr virus. J Infect Dis. 2005;191(2):234–237. doi:10.1086/426402
89. Hughes CM, Liu L, Davidson WB, et al. A tale of two viruses: coinfections of monkeypox and varicella zoster virus in the democratic Republic of Congo. Am J Trop Med Hyg. 2020;104(2):604–611. doi:10.4269/ajtmh.200589
90. Loh J, Tham SM, Tambyah PA, Yan G, Lee CK, Chai LYA. Range of varicella zoster co-infections with COVID-19, Singapore. Infect Chemother. 2021;53(2):391–394. doi:10.3947/ic.2020.0154
91. The centers for disease control and prevention. Chickenpox (varicella). Available from: https://www.cdc.gov/chickenpox/hcp/index.html.
92. Tellier R, Li Y, Cowling BJ, Tang JW. Recognition of aerosol transmission of infectious agents: a commentary. BMC Infect Dis. 2019;19(1):101. doi:10.1186/s12879-019-3707y
93. The centers for disease control and prevention. Shingles (herpes zoster). Available from: https://www.cdc.gov/shingles/hcp/index.html.
94. Sumi A. Role of temperature in reported chickenpox cases in Northern European countries: Denmark and Finland. BMC Res Notes. 2018;11(1):377. doi:10.1186/s13104018-3497-0
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.