")
Back to Journals » OncoTargets and Therapy » Volume 12
Targeting Trop-2 in solid tumors: future prospects
Authors Zaman S, Jadid H, Denson AC , Gray JE
Received 5 November 2018
Accepted for publication 29 January 2019
Published 1 March 2019 Volume 2019:12 Pages 1781—1790
DOI https://doi.org/10.2147/OTT.S162447
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Leo Jen-Liang Su
Saif Zaman,1 Hassan Jadid,2 Aaron C Denson,2 Jhanelle E Gray2
1Morsani College of Medicine, University of South Florida, Tampa, FL, USA; 2Department of Thoracic Oncology, H. Lee Moffitt Cancer Center and Research Institute, Tampa, FL, USA
Abstract: Trop-2 is a transmembrane glycoprotein that is upregulated in all cancer types independent of baseline levels of Trop-2 expression. Trop-2 is an ideal candidate for targeted therapeutics due to it being a transmembrane protein with an extracellular domain overexpressed on a wide variety of tumors as well as its upregulated expression relative to normal cells. As a result, several Trop-2-targeted therapeutics have recently been developed for clinical use, such as anti-Trop-2 antibodies and Trop-2-targeted antibody–drug conjugates (ADC). Subsequently, multiple early-phase clinical trials have demonstrated safety and clinical benefit of Trop-2-based ADCs across multiple tumor types. This includes clinical benefit and tolerability in tumor types with limited treatment options, such as triple-negative breast cancer, platinum-resistant urothelial cancer, and small-cell lung cancer. In this review, we elaborate on all clinical trials involving Trop-2.
Keywords: Trop-2, trophoblast antigen 2, targeted therapy, antibody–drug conjugates, oncotargets
Introduction
Trop-2, also known as epithelial glycoprotein-1, gastrointestinal antigen 733-1, membrane component surface marker-1, and tumor-associated calcium signal transducer-2, is the protein product of the TACSTD2 gene. This transmembrane glycoprotein functions in a variety of cell signaling pathways and was first elucidated as a transducer of an intracellular calcium signal.1 Trop-2 expression has been demonstrated to depend on a large variety of transcription factors. The transcription factor HNF4A has been shown to be the major hub for TACSTD2 (Trop-2) transcription. However, other transcription factors known to be associated with cancer development are involved in TACSTD2 (Trop-2) transcription, such as TP63/TP53L and Wilm’s tumor 1 (WT1). Other transcription factors demonstrated to be involved in TACSTD2 (Trop-2) transcription include ERG, HNF1A/TCF-1, autoimmune regulator, and FOXP3, among others.2 Trop-2 is involved in several cell signaling pathways, of which many are associated with tumorigenesis. For example, in thyroid cancer cell invasion, Trop-2 signal transduction has been seen as a downstream effect of the ERK and JNK pathways.3 Stoyanova et al demonstrated that Trop-2 signaling enhances stem cell-like properties of cancer cells, as Trop-2 regulates proliferation and self-renewal through b-catenin signaling.4 It has been speculated that phosphatidylinositol 4,5-bisphosphate (PIP2) may regulate Trop-2 phosphorylation and calcium signal transduction, as the cytoplasmic domain of Trop-2 contains a PIP2-binding sequence overlapping with a protein kinase C phosphorylation site.5
Trop-2 may play a role in tumor progression given the involvement in several molecular pathways traditionally associated with cancer development. High Trop-2 expression correlates with poor prognosis in pancreatic, hilar cholangiocarcinoma, cervical cancer, gastric cancer, and others.6–9 In a meta-analysis including 2,569 patients, increased Trop-2 expression was associated with poor overall and disease-free survival outcomes across several solid tumors.10 Given Trop-2’s expression pattern and associated poor prognostic outcomes, Trop-2 is a rational prognostic marker and therapeutic target.
Basic science background
Background/controversy
Lipinski et al first discovered Trop-1, Trop-2, Trop-3, and Trop-4 expression on synctio- and cytotrophoblasts after generating monoclonal antibodies against the human choriocarcinoma cell line, BeWo.11 It is important to note that as with Lipinski’s initial discovery of Trop-2 in noncancerous trophoblast cells, Trop-2 has been demonstrated to show variable levels of expressivity in other noncancer cell types, for example it is normally differentiating human keratinocytes.12,13 This has raised the question of whether Trop-2-targeted therapeutic approaches may result in toxicity. However, in support of therapeutic targeting of Trop-2, Trerotola et al demonstrated that Trop-2 is upregulated in all cancer types assayed independent of baseline levels of Trop-2 expression in normal cell counterparts.14 Trop-2 expression in normal cell seems to be cell-type dependent; Zhang et al conducted a gene expression pattern analysis comparing gastrointestinal tumors to their normal counterpart and demonstrated that Trop-2 was not overexpressed in normal tissue.15 As such, it may be pertinent to characterize Trop-2 expression in normal tissue on a cell type by cell type basis in order to postulate potential targets for Trop-2-dependent therapy.
The reason why Trop-2 is overexpressed in many cancers compared to noncancer cells is not completely understood. One reason may be that transcription factors known to be involved in cancer cell progression, such as WT1, also regulate Trop-2 transcription. Others have suggested that due to Trop-2’s intrinsic regulatory effects on cancer cell growth, invasion, and proliferation, the overexpression of Trop-2 naturally leads to tumor progression, suggesting a selective advantage.16 Additionally, Trerotola et al have suggested that Trop-2 expression is a key driver of cancer growth. This group has demonstrated that Trop-2 upregulation quantitatively stimulates tumor growth, while Trop-2 knockdown inhibits tumor cell growth.14 As such, possibly in a tumor’s evolution, clones that overexpress Trop-2 would be more represented among the population of cells due to increased proliferation. However, Wang et al have demonstrated that Trop-2-null mice are susceptible to increased incidence of sarcomatoid carcinomas.17 This study demonstrated how Trop-2 loss of function may also play a role in tumorigenesis, contrasting from previous studies, which demonstrated how Trop-2 gain of function increases cancer cell proliferation. As such it has been suggested that Trop-2 plays a context-dependent role in cancer development.18
Trop-2 function in tumorigenesis seems to be evolutionarily conserved. Murine Trop-2 shares 87.4% sequence similarity with its human homolog. Similar to its human homolog, murine Trop-2 mRNA is detected in normal murine tissue, including normal keratinocytes, kidney, lung, ovary, and testis.5 Moreover, Cubas et al demonstrated that murine Trop-2 expression increased cancer cell proliferation in vitro and in orthotopic pancreatic cancer mouse models. Cubas et al also demonstrated that human and murine Trop-2 share similar downstream targeting pathways, as ERK activation following Trop-2 overexpression was observed in both murine models and human pancreatic and colorectal cancer cells.19
It has long been established that matrix metallopeptidases and other extracellular proteases play a key role in extracellular matrix remodeling and cancer cell invasion and metastasis across several cell types.20–22 The ERK pathway is involved in several processes, including cancer cell invasion by regulating MMP2 expression.23,24 Antineoplastic therapies involving the suppression of MMP2 by inhibiting the ERK pathway are being developed.25 Given Trop-2’s initial discovery in trophoblast cells, which are invasive, it seems a logical step forward to investigate Trop-2’s function on cancer cell invasion. Li et al demonstrated that Trop-2 overexpression increases invasion in the A459 non-small-cell lung cancer (NSCLC) cell line.26 Similarly, it has been demonstrated that Trop-2 overexpression increases invasion of cervical, ovarian, colon, and thyroid cancers.3,8,27,28 Consistent with this, several groups have demonstrated that knockdown of Trop-2 decreases cancer cell invasion.26,29 The invasion resultant from Trop-2 overexpression is dependent on the ERK pathway and has been demonstrated to induce MMP2 in thyroid cancer.3,8 Similarly, Zhao et al have demonstrated that TNF-a enhanced colon cancer cell invasion via Trop-2 upregulation through the ERK1/2 pathway.28 This data supports the idea that Trop-2 plays a role in cancer cell invasion through downstream regulation of MMP2 via ERK.
Consistent with Trop-2’s role in cancer cell invasion, recent findings have demonstrated that Trop-2 regulates integrin-dependent signaling for cell–substrate adhesion and cell migration. For example, Trerotola et al have demonstrated that Trop-2 modulates β1 integrin functions to promote prostate cancer metastasis. The group has shown that Trop-2 and the α5β1 integrin associate through their extracellular domains. Moreover, they have also demonstrated that Trop-2 modulates Rac1 GTPase activity and induces activation of PAK4, thereby enhancing prostate cancer cell migration.30 Furthermore, regulating the linkage between the cytoskeleton and integrins is critical for cell migration and metastasis. Trerotola et al have demonstrated that Trop-2 regulates prostate cancer cell adhesion to fibronectin through the β1 integrin–RACK1–Src–FAK signaling axis.31
Trop-2 expression and clinical correlations across several cancer types
The differential expression of Trop-2 on cancer and normal epithelial cells continues to be investigated. Nakashima et al identified serum anti-Trop-2 antibodies in patients with esophageal squamous cell carcinoma (SCC) and demonstrated that Trop-2 expression was significantly greater in esophageal squamous cell cancer cell lines than both normal esophageal mucosa and immortalized esophageal mucosal cells. Moreover, through immunohistochemistry (IHC) analysis this group demonstrated that expression of Trop-2 protein in esophageal SCC was higher than expression of Trop-2 in hyperplastic esophageal mucosa.32
Eventually, clinical correlations regarding Trop-2 expression were investigated. Ohmachi et al demonstrated that Trop-2 expression correlates with poor prognosis and aggressive presentation of colorectal cancer and suggested Trop-2 as a possible candidate gene for targeted therapy of colorectal cancer.33 Fong et al demonstrated that Trop-2 expression is associated with poor survival outcomes and increased lymph node metastasis in pancreatic cancer.6
Wang et al first identified Trop-2 as an oncogene and demonstrated that knockdown of Trop-2 expression inhibits both tumorigenesis and invasion of colon cancer cells and that ectopic expression of Trop-2 can promote cancer cell growth.34 Results from Trerotola et al were consistent and demonstrate that upregulation of Trop-2 drives cancer cell growth across cell type and species.14 Trerotola et al also suggested a model where upregulation of wild-type Trop-2 is a key driver of cancer cell growth.
More recently, Trop-2 activity has been correlated to additional, classical tumor drivers. Kong et al correlated Trop-2 IHC to the presence of BRAF mutations, which are the most common genetic alteration in papillary thyroid cancer. The group concluded that Trop-2 expression is associated with BRAF mutations and that Trop-2 IHC could be used to predict BRAF mutations and alternatively diagnosing papillary thyroid cancer.35 Moreover, Zhao et al have correlated Trop-2 mRNA and protein expression levels to several breast cancer clinical prognostic indicators, including lymph node status, metastasis, stage, and estrogen receptor/progesterone receptor/HER2 expression. The group demonstrates that Trop-2 is a potential biomarker for the promotion of EMT in breast cancer.36 Gu et al have demonstrated that Trop-2 expression promotes proliferation and migration of osteosarcoma cell lines through the PI3K/AKT pathway. Interestingly, the group also demonstrated that Trop-2 knockdown decreases cell migration and proliferation, hinting toward therapeutic benefit of Trop-2 inhibition.37
Eventually it was discovered that Trop-2 may modulate the effects of certain chemotherapeutic regimens. For example, Jordheim et al have suggested that Trop-2 is potentially involved in a paradoxical growth-promoting effect of oxaliplatin, a platinum-based chemotherapeutic, when administered to a xenograft model of HCT-116 human colon cancer cells.38 Moreover, Zhang et al found that curcumin treatment decreases Trop-2 expression and that downregulation of Trop-2 expression suppressed cell proliferation and mobility and increased apoptosis in bladder cancer cells.39
Preclinical therapeutic development
Because Trop-2 is a transmembrane protein with an extracellular domain overexpressed on a wide variety of tumors, with upregulated expression relative to normal cells it is a natural candidate for targeted therapeutic development. Early studies on the therapeutic potential for Trop-2 targeting focused on native Trop-2 tumor antigen presentation on major histocompatability complex Class I. Mangino et al demonstrated that Trop-2-specific cytotoxic T lymphocytes were able to lyse MCF7 human breast cancer cells expressing endogenous HLA-A*02:01.40 Moreover, Nakashima et al have demonstrated that in a sample of 75 patients, 31% of patients were positive for serum anti-Trop-2 antibodies.32 Together, these studies suggest a future role that Trop-2 could play in immunotherapy.
Lin et al have demonstrated that Trop-2-targeted antigen-binding fragments (Fab) have inhibitory effects on breast cancer development in xenograft mouse models of MDA-MB-231 breast cancer cells. Moreover, the group demonstrated that the Trop-2 Fab was able to induce apoptosis in vivo, as evidenced by increased Caspase-3 expression and decreased bcl2 expression.41 Although several anti-Trop-2 antibodies have been developed, none were amenable to therapeutic use as naked antibodies until Ikeda et al developed Pr1E11. This is an anti-Trop-2 antibody with low internalization activity isolated from Balb/c mice immunized with Pc1 primary human prostate cancer cells, which the group suggested for further investigation in epithelial cancers.42
Following the development of anti-Trop-2 antibodies, Trop-2-targeted antibody–drug conjugates (ADC) were later developed. For example, anti-Trop-2 antibodies conjugated to doxorubicin-loaded nanoparticles demonstrated higher toxicity toward MDA-MB-231 triple-negative breast cancer cells when compared to doxorubicin-loaded control nanoparticles without the anti-Trop-2 antibody conjugate.43 The utility of anti-Trop-2 antibodies conjugated to additional chemotherapeutic agents has been demonstrated in a variety of preclinical studies. One Trop-2 ADC linked to a proprietary microtubule inhibitor displayed subnanomolar half maximal inhibitory concentration (IC50) concentrations in a panel of Trop-2-expressing cancer cell lines.44 Goldenberg et al presented IMMU-132, a conjugate of SN-38, a metabolite of irinotecan, to an anti-Trop-2 antibody. Initial preclinical studies demonstrated that IMMU-132 was able to deliver up to 136-fold more SN-38 to a xenograft mouse model than irinotecan with lower toxicity frequency, including lower frequency of severe diarrhea than irinotecan.45 These studies demonstrated superior SN-38 delivery by IMMU-132.46 There remains the question of whether the conjugated SN-38 is able to interact with the effects of the anti-Trop-2-antibody in signaling the cells to lead to either an additive or synergistic effect. Goldenberg et al have demonstrated that there are increased DNA double-strand breaks in vitro when Trop-2-expressing cells are treated with IMMU-132, when compared to another SN-38 ADC, hA20-SN-38, which is targeted to CD20.45 As such, synergy between anti-Trop-2 antibodies and their respective payloads in ADCs warrants further investigation.
Moreover, other humanized anti-Trop-2 IgG-SN-38 conjugates, such as the anti-Trop-2 hRS7-CL2A-SN-38 ADC, have been demonstrated to provide significant and specific anticancer effects in xenograft models of several tumor cell lines (Calu-3, Capan-1, BxPC-3, and COLO 205). Additionally, this same anti-Trop-2 ADC has been demonstrated to be well tolerated in monkeys, paving the way for future human clinical trials.47 Within the clinics, Han et al have presented a case of a 74-year-old woman that demonstrated an excellent response to IMMU-132 with 66% reduction of target lesions by RECIST criteria with over 10 months duration of response. The patient had widespread therapy-resistant uterine serous carcinoma and had previously failed multiple immunotherapy and chemotherapy regimens.48 These results across various Trop-2 antibodies and payloads, from cell lines to humans, suggest Trop-2’s potential for use in synergistic conjugation with more classical DNA-damaging chemotherapeutic options.
Moreover, Trop-2-targeted therapy in combination with other targeted therapy has also been demonstrated. Cardillo et al have demonstrated synergy between IMMU-132 and three different PARP inhibitors (olaparib, rucaparib, and talazoparib) in a mouse xenograft model of HCC1806 triple-negative breast cancer tumors, independent of BRCA1/2 status.49 Additionally, as ATP-binding cassette (ABC) transporters are associated with chemotherapeutic resistance through chemotherapy efflux from cancer cells, Chang et al investigated the combination of IMMU-132 and ABCG2 inhibitors in SN-38-resistant xenograft models and demonstrated that the combination increased median survival of mice.50 The synergistic behavior of Trop-2-targeted antibodies in these vastly different therapeutic strategies demonstrates potential diversity in Trop-2-targeted therapy.
Although the majority of Trop-2-targeted therapeutics have focused on ADCs, there have been additional delivery methods of Trop-2-targeted therapeutics. Farivar et al demonstrated that liposomes loaded with apoptosis activator 2 targeting Trop-2 cell surface antigen is able to induce apoptosis in a gastric adenocarcinoma cell line.51 ImmunoRNases have been generated as targeted cancer therapeutics; Trop-2 targeting tetrakis–ranpirnase is an example. Liu et al have demonstrated significant survival benefit in a xenograft mouse model of triple-negative breast cancer.52
Development for use in clinical imaging
Immunopositron emission tomography involves a targeted approach for assessing tumor distribution.53 TF12 is an anti-Trop-2 × antihapten bispecific antibody that has been demonstrated to be well retained on the cell surface of several epithelial tumors, suggesting that it can be used for pretargeted imaging.54 Trop-2, via TF12, has been demonstrated to effective for pretargeted therapy with radioactively labeled hapten peptides (68Ga, 177Lu-IMP288) and for specific imaging of prostate cancer in mouse xenograft models of human prostate cancer cells.55,56 Furthermore, the efficacy of Trop-2 targeting in radioimmunotherapy has been demonstrated. van Rij et al have demonstrated that mice treated with the trivalent bispecific antibody TF12 (anti-Trop-2 × anti-HSG [histamine-succinyl-glycine]) conjugated to (177)Lu-labeled diHSG-peptide (IMP288) allowed rapid accumulation of the radioimmunotherapeutic and inhibited growth of prostate cancer tumors in a mouse model.57 As such, there exists a use case for Trop-2 targeting in both imaging and radioimmunotherapy. Specifically, imaging approaches may provide a potential method to target metastatic cancer distribution and act as a prognostic indicator for response to Trop-2-targeted therapeutic approaches.
Clinical trials
There are currently several examples of Trop-2 in human clinical trials. These studies are summarized in Table 1.
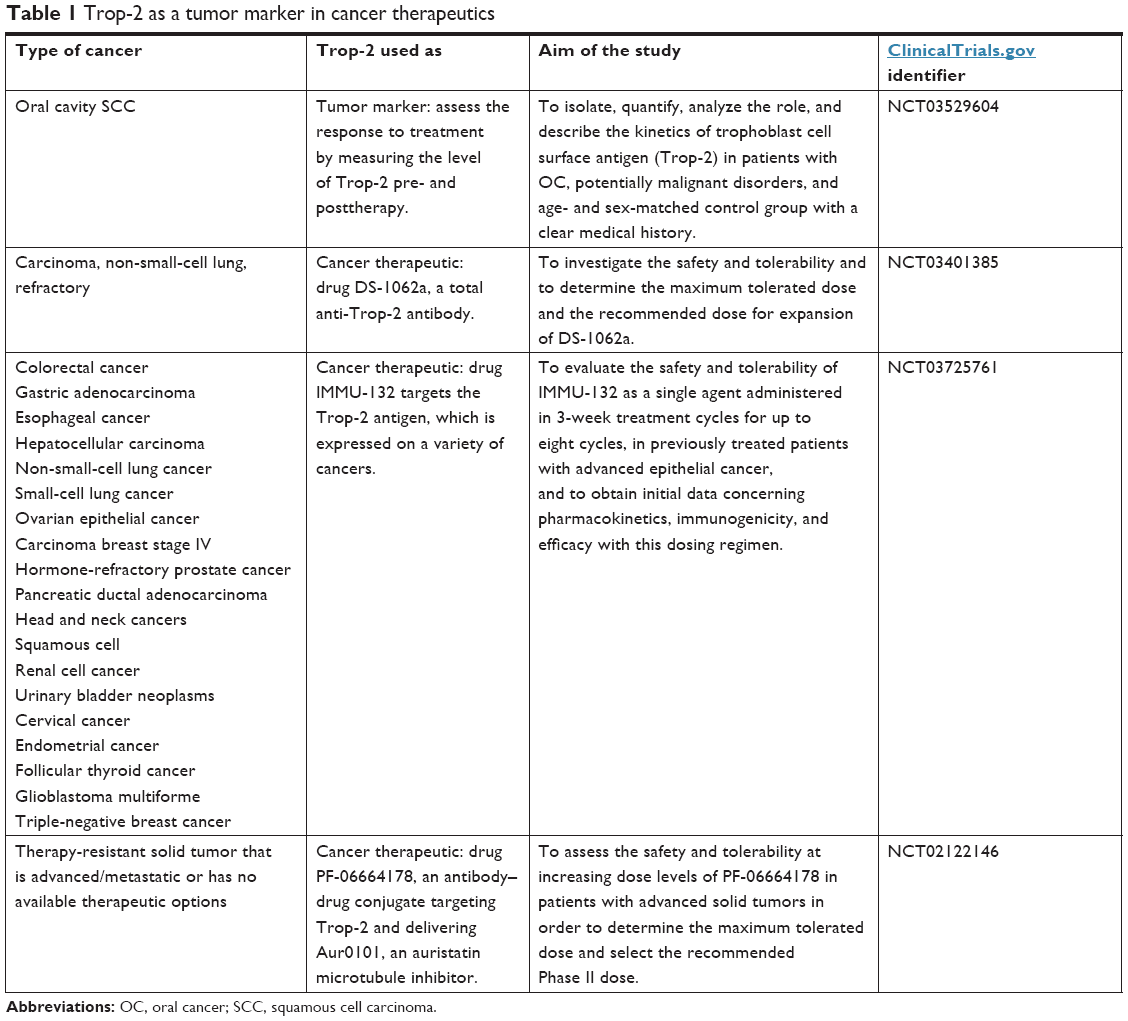 | Table 1 Trop-2 as a tumor marker in cancer therapeutics |
Dose-limiting toxicities for most ADCs undergoing clinical development are well associated with the payload and seem to be unrelated to the antigen that is targeted.58 Toxicity is a specific concern with anti-Trop-2 ADCs due to the expression of Trop-2 in normal tissues. Cardillo et al evaluated the efficacy of an SN-38-anti-Trop-2 ADC against human tumor types in cynomolgus monkeys, which express Trop-2 across several different normal tissue types similar to humans. Histological assessment of monkey liver tissue posttreatment demonstrated that normal tissues that express Trop-2 had no evidence of toxicity-related damage.47 As such, it appears that toxicity of Trop-2 ADCs follows the trend of other ADCs, whereby toxicities are more associated with the payload, rather than the targeted antigen. Of course, additional investigation of Trop-2 ADCs and on-target nonspecific toxicity will need to be conducted in humans.
A study of PF-06664178 in patients with advanced solid tumors (NCT02122146)
A Phase I study was conducted on PF-06664178, an ADC targeting Trop-2 and delivering Aur0101, an auristatin microtubule inhibitor.59 Inclusion criteria include the diagnosis of a therapy-resistant solid tumor that is advanced/metastatic or has no available therapeutic options. Of note, there was no requirement of Trop-2 overexpression to be enrolled in this study. The primary objective of this study was to determine maximum tolerated dose and doses for Phase II studies. Thirty-one patients with metastatic solid tumors were treated with escalating doses of PF-0664178. Of those 31 patients, 11 patients resulted in stable disease; there were no complete or partial responses. In February 2016, this clinical trial was prematurely terminated due to a business-related decision based on results from the study.
In this study, 31 patients with advanced or metastatic solid tumors were treated with PF-06664178 in escalating doses between 0.15 and 4.8 mg/kg. At the 3.6 mg/kg dose, two out of six (33%) patients experienced a dose-limiting toxicity; the toxicities were grade 4 neutropenia and grade 3 mucosal inflammation. At the 4.2 mg/kg dose, one out of one (100%) patients experienced a dose-limiting toxicity; this toxicity was a grade 3 maculopapular rash. At the 4.8 mg/kg dose, four out of eight patients (50%) experienced a dose-limiting toxicity; these toxicities included grade 4 febrile neutropenia, grade 4 toxic epidermal necrolysis, grade 4 dehydration, and grade 3 rash.59
Phase I/II study of IMMU-132 in patients with epithelial cancers (NCT01631552)
Sacituzumab govitecan (IMMU-132) is an ADC targeting Trop-2 and delivering SN-38.60 The Phase I data have been published, describing the first-in-human trial of IMMU-32 evaluated across 25 patients with 13 different epithelial tumors.61 Inclusion criteria included the diagnosis of a therapy-resistant solid tumor that is advanced/metastatic or has no available therapeutic options. The primary objective of this study was to obtain initial data on safety and tolerability of IMMU-132 in patients with advanced epithelial cancer that have been previously treated. This study included patients with various tumor types including breast, colorectal, endometrial, and both SCLC and NSCLC, among others. At the study end point, two patients achieved partial responses; specifically, these two patients had triple-negative breast cancer and colon cancer. Additionally, 16 other patients had stable disease while 12 patients maintained disease control with continued treatment for 16–26 weeks; of these patients, 6 survived for 15–20+ months. In this study, there were no treatment-related grade 4 toxicities, and the grade 3 toxicities were limited to diarrhea (n=1), fatigue (n=3), and neutropenia (n=2).61 As such, IMMU-132 is overall a well-tolerated therapeutic.
Phase II results have been published within several tumor types including SCLC, NSCLC, triple-negative breast cancer, and urothelial cancer. The results of these studies are detailed below.
Phase II study of IMMU-132 in patients with breast cancer
Given the encouraging outcome for the triple-negative breast cancer patient in the Phase I dose-finding trial for IMMU-132, Bardia et al have reported the efficacy and safety of IMMU-132 in heavily pretreated patients with metastatic triple-negative breast cancer.62 Results indicate that, of 69 patients, 2 patients had a complete response, and 19 had a partial response. The group demonstrated that the clinical benefit rate (in this study defined as complete response + partial response + stable disease ≥6 months) was 46%. The median response duration was 8.9 months, and the median overall survival was 16.6 months while the median progression-free survival was 6.0 months. As for toxicities, the patients experienced grade ≥3 adverse events, which included anemia (14%), diarrhea (13%), leukopenia (16%), and neutropenia (39%).62 These results are encouraging given the association between Trop-2 expression and unfavorable survival outcomes in breast cancer patients,63 high mortality associated with triple-negative breast cancer, and relative lack of effective therapeutic options for patients with metastatic triple-negative breast cancer.
Phase I/II study of IMMU-132 in patients with urothelial cancer
Faltas et al reported six patients with metastatic, platinum-resistant urothelial carcinoma (PRUC) who received sacituzumab govitecan (IMMU-132). Trop-2 is widely expressed in ≤83% of urothelial carcinomas. The median number of previous therapies was 3. Three of six patients had a clinically significant response (progression-free survival, 6.7–8.2 months; overall survival, 7.5+ to 11.4+ months). Sacituzumab govitecan was well tolerated. Two of six patients experienced grade 3 toxicity (flank pain and bacteremia) but no grade 4 nonhematologic toxicity was observed.64 In light of these promising results, a multicenter Phase II cohort has been expanded to enroll 50 patients with PRUC.
Phase II study of IMMU-132 in patients with NSCLC
Heist et al evaluated IMMU-132 in 54 pretreated metastatic NSCLC patients. The group demonstrated that the clinical benefit rate (in this study defined as complete response + partial response + stable disease ≥4 months) was 43%. Moreover, the median response duration to therapy was 6 months. The median overall survival was 9.5 months and the median progression-free survival was 5.2 months. Regarding toxicities, the patients experienced grade ≥3 adverse events, which included diarrhea (7%), fatigue (6%), febrile neutropenia (4%), nausea (7%), and neutropenia (28%). These results are encouraging, considering responses from patients who did not respond well to immune checkpoint inhibitor therapy.65
Phase II study of IMMU-132 in patients with SCLC
Another study evaluated IMMU-32 in heavily pretreated metastatic SCLC patients, including patients who were chemoresistant or chemosensitive to first-line chemotherapy. In this study 60% of patients demonstrated tumor shrinkage when compared to baseline CTs, with a 34% clinical benefit rate (in this study defined as complete response + partial response + stable disease ≥4 months). The median overall survival was 7.5 months, and the median progression-free survival was 3.7 months. Regarding toxicities, the patients experienced grade ≥3 adverse events, which included anemia (6%), diarrhea (9%), fatigue (13%), and neutropenia (34%). Results indicated that there was no difference between first-line chemosensitive vs chemoresistant patients; however, there was a suggested improvement in clinical benefit rate in second-line patients who were sensitive to the first-line therapy.66 There have been other studies that have demonstrated clinical benefit of ADCs to SCLC patients. For example, in a Phase I open-label study, rovalpituzumab tesirine, a DLL3-targeted ADC, demonstrated confirmed objective response in 18% of 60 assessable patients.67
Salivary Ap4A, SCCA, Trop-2 in oral cancer patients (NCT03529604)
This study aims to characterize the role and kinetics of salivary levels of Trop-2, among other proteins, as a salivary tumor marker for patients with oral cancer using liquid chromatography-ion trap-mass spectrometry, multiple reaction monitoring method. The study further aims to develop a highly sensitive and highly specific test in order to measure levels of Trop-2 and other tumor markers before and after therapy and during regular patient follow-up. This study is ongoing, although not actively recruiting and has an estimated study completion date of April 30, 2020.
First-in-human study of DS-1062a for advanced solid tumors (NCT03401385)
DS-1062a is an ADC targeting Trop-2 and delivering DXd, a topoisomerase I inhibitor and derivative of exatecan.74 Inclusion criteria include the diagnosis of an unresectable advanced NSCLC not amenable to curative intent therapeutic approaches. The primary objective of this study is to determine the safety and tolerability of DS-1062a. This study is ongoing and currently actively recruiting; it has an estimated study completion date of June 20, 2021. Prior to this clinical trial, one group demonstrated that DS-1062 markedly reduced in vitro cancer cell growth with IC50 dosing in the nanomolar range in Trop-2+ cell lines (CFPAC1, BxPC-3); to contrast the group also demonstrated that in Trop-2-negative tumor cells (Calu-6), 100-fold greater dosing was needed to achieve IC50.74
Discussion
The search for an ideal cancer-specific protein for targeted therapy has been ongoing for decades now. The Trop-2 oncogene is an ideal target for therapeutic development and ADCs, as it is ubiquitously overexpressed across solid tumors and is membrane bound with an extracellular domain. Moreover, in vitro testing of Trop-2-targeted therapeutics has been favorable, with certain Trop-2 ADCs demonstrating subnanomolar IC50s.44 Several animal models and human clinical trials have demonstrated tumor suppression and favorable therapeutic indexes with Trop-2-targeted therapy. As such, the future of Trop-2 as a therapeutic target appears promising.
In particular, the role of Trop-2-directed therapy as part of combination therapy seems particularly promising. Synergistic combination therapy has been used as a means to administer lower nontoxic dosages to decrease patient burden with drastic anticancer effects.68 Trop-2-targeted therapeutics have already been demonstrated to be potent and synergistic in combination with a number of other therapeutics in a preclinical setting. However, given the multitude of cell signaling pathways Trop-2 is involved with, it seems Trop-2-targeted therapeutics are amenable to rational combination therapy targets with several anticancer agents. For example, given Trop-2’s downstream activation of ERK pathway, it may be interesting to investigate combination therapy with Trop-2-targeted therapies and anticancer agents targeting the MEK/ERK pathway, such as selumetinib.69 Additionally, Trop-2 is a strong predictor of cancer response to AKT inhibitors, and as such AKT inhibitors seem to be a rational target for further investigation along with Trop-2-targeted therapy.70 Moreover, Wang et al have demonstrated that increased Trop-2 expression is associated with cisplatin resistance in lung cancer cells.71 Given the benefit of IMMU-132 (sacituzumab govitecan) therapy, which provides additional benefit at lower dosages than using irinotecan, this study demonstrates additional potential benefits of combining Trop-2-targeted therapeutics and traditional DNA-damaging chemotherapy, as we theorize Trop-2 inhibition may sensitize cells to DNA-damaging chemotherapy.
One interesting use case for Trop-2-targeted therapeutics may be further investigation in tumors where traditional targets do not currently exist, such as in triple-negative breast cancer.62 Trop-2 is overexpressed in 80% of triple-negative breast cancer patients.43 Moreover, as our understanding of tumor heterogeneity has evolved, it is apparent that different clonal populations of disease, whether at primary or metastatic sites, may respond differently to therapy. Nevertheless, rational exists that many potential uses for Trop-2-targeted therapeutics exist for metastatic disease. Mojica et al reported a case of metastatic invasive lobular carcinoma that presented with overexpression of Trop-2, suggesting Trop-2-targeted therapeutics may be useful for treating systemic disease.72 Given Trop-2’s ubiquitous overexpression on a wide variety of cancer cell types and amenability for combination therapy, the problems that arise with tumor heterogeneity in response to monotherapy comes into question, and whether or not Trop-2 combination therapy can overcome chemotherapeutic resistance is a question that remains mostly unexplored.
Although there have been reports of Trop-2 therapeutic efficacy across distinct cancer types, there has been little direct comparisons of Trop-2-targeted agents to other therapeutic approaches. Govindan et al have demonstrated through in vitro cytotoxicity and in vivo studies that anti-Trop-2 conjugates were superior to milatuzumab-SN-38 (an anti-CD74 antibody conjugated to SN-38) in gastric cancer. However, milatuzumab-SN-38 was more effective in lymphoma, melanoma, and pancreatic cancer models.73 The role of Trop-2-targeted agents in clinical practice is not clear at this time, and there is a need for further head-to-head comparisons across different tumor types. There is strong evidence that Trop-2 has potential as a therapeutic target, although much is unknown regarding the preferred disease type, preferred therapeutic combinations, drug-delivery methods, and side effect profiles. Further studies on Trop-2-targeted therapeutics are necessary to further elucidate these possibilities.
Disclosure
The authors report no conflicts of interest in this work.
References
Ripani E, Sacchetti A, Corda D, Alberti S. Human Trop-2 is a tumor-associated calcium signal transducer. Int J Cancer. 1998;76(5):671–676. | ||
Guerra E, Trerotola M, Aloisi AL, et al. The Trop-2 signalling network in cancer growth. Oncogene. 2013;32(12):1594–1600. | ||
Guan H, Guo Z, Liang W, et al. Trop2 enhances invasion of thyroid cancer by inducing MMP2 through ERK and JNK pathways. BMC Cancer. 2017;17(1):486. | ||
Stoyanova T, Goldstein AS, Cai H, et al. Regulated proteolysis of Trop2 drives epithelial hyperplasia and stem cell self-renewal via β-catenin signaling. Genes Dev. 2012;26(20):2271–2285. | ||
El Sewedy T, Fornaro M, Alberti S. Cloning of the murine Trop2 gene: conservation of a PIP2-binding sequence in the cytoplasmic domain of Trop-2. Int J Cancer. 1998;75(2):324–330. | ||
Fong D, Moser P, Krammel C, et al. High expression of Trop2 correlates with poor prognosis in pancreatic cancer. Br J Cancer. 2008;99(8):1290–1295. | ||
Ning S, Guo S, Xie J, et al. Trop2 correlates with microvessel density and poor prognosis in hilar cholangiocarcinoma. J Gastrointest Surg. 2013;17(2):360–368. | ||
Liu T, Liu Y, Bao X, et al. Overexpression of Trop2 predicts poor prognosis of patients with cervical cancer and promotes the proliferation and invasion of cervical cancer cells by regulating ERK signaling pathway. PLoS One. 2013;8(9):e75864. | ||
Zhao W, Zhu H, Zhang S, et al. Trop2 is overexpressed in gastric cancer and predicts poor prognosis. Oncotarget. 2016;7(5):6136–6145. | ||
Zeng P, Chen MB, Zhou LN, et al. Impact of Trop2 expression on prognosis in solid tumors: a systematic review and meta-analysis. Sci Rep. 2016;6:33658. | ||
Lipinski M, Parks DR, Rouse RV, Herzenberg LA. Human trophoblast cell-surface antigens defined by monoclonal antibodies. Proc Natl Acad Sci U S A. 1981;78(8):5147–5150. | ||
Alberti S, Miotti S, Stella M, et al. Biochemical characterization of Trop-2, a cell surface molecule expressed by human carcinomas: formal proof that the monoclonal antibodies T16 and MOv-16 recognize Trop-2. Hybridoma. 1992;11(5):539–545. | ||
Klein CE, Cordon-Cardo C, Soehnchen R, et al. Changes in cell surface glycoprotein expression during differentiation of human keratinocytes. J Invest Dermatol. 1987;89(5):500–506. | ||
Trerotola M, Cantanelli P, Guerra E, et al. Upregulation of Trop-2 quantitatively stimulates human cancer growth. Oncogene. 2013;32(2):222–233. | ||
Zhang L, Zhou W, Velculescu VE, et al. Gene expression profiles in normal and cancer cells. Science. 1997;276(5316):1268–1272. | ||
Shvartsur A, Bonavida B. Trop2 and its overexpression in cancers: regulation and clinical/therapeutic implications. Gene Canc. 2015;6(3–4):84–105. | ||
Wang J, Zhang K, Grabowska D, et al. Loss of Trop2 promotes carcinogenesis and features of epithelial to mesenchymal transition in squamous cell carcinoma. Mol Canc Res. 2011;9(12):1686–1695. | ||
McDougall AR, Tolcos M, Hooper SB, Cole TJ, Wallace MJ. Trop2: from development to disease. Dev Dyn. 2015;244(2):99–109. | ||
Cubas R, Zhang S, Li M, Chen C, Yao Q. Trop2 expression contributes to tumor pathogenesis by activating the ERK MAPK pathway. Mol Cancer. 2010;9:253. | ||
Stamenkovic I. Matrix metalloproteinases in tumor invasion and metastasis. Semin Cancer Biol. 2000;10(6):415–433. | ||
Zaman S, Chobrutskiy BI, Patel JS, et al. Mutant cytoskeletal and ECM peptides sensitive to the ST14 protease are associated with a worse outcome for glioblastoma multiforme. Biochem Biophys Res Commun. 2018;503(4):2218–2225. | ||
Zaman S, Chobrutskiy BI, Patel JS, et al. MMP7 sensitivity of mutant ECM proteins: an indicator of melanoma survival rates and T-cell infiltration. Clin Biochem. 2019;63:85–91. | ||
Wang D, Wang D, Wang N, Long Z, Ren X. Long non-coding RNA BANCR promotes endometrial cancer cell proliferation and invasion by regulating MMP2 and MMP1 via ERK/MAPK signaling pathway. Cell Physiol Biochem. 2016;40(3–4):644–656. | ||
Poudel B, Lee YM, Kim DK. DDR2 inhibition reduces migration and invasion of murine metastatic melanoma cells by suppressing MMP2/9 expression through ERK/NF-κB pathway. Acta Biochim Biophys Sin. 2015;47(4):292–298. | ||
Liu Y, Zhu P, Wang Y, et al. Antimetastatic therapies of the polysulfide diallyl trisulfide against triple-negative breast cancer (TNBC) via suppressing MMP2/9 by blocking NF-κB and ERK/MAPK signaling pathways. PLoS One. 2015;10(4):e0123781. | ||
Li Z, Jiang X, Zhang W. Trop2 overexpression promotes proliferation and invasion of lung adenocarcinoma cells. Biochem Biophys Res Commun. 2016;470(1):197–204. | ||
Wu B, Yu C, Zhou B, et al. Overexpression of Trop2 promotes proliferation and invasion of ovarian cancer cells. Exp Ther Med. 2017;14(3):1947–1952. | ||
Zhao P, Zhang Z. TNF-α promotes colon cancer cell migration and invasion by upregulating Trop-2. Oncol Lett. 2018;15(3):3820–3827. | ||
Gao XY, Zhu YH, Zhang LX, Lu HY, Jiang AG. siRNA targeting of Trop2 suppresses the proliferation and invasion of lung adenocarcinoma H460 cells. Exp Ther Med. 2015;10(2):429–434. | ||
Trerotola M, Jernigan DL, Liu Q, et al. Trop-2 promotes prostate cancer metastasis by modulating β(1) integrin functions. Cancer Res. 2013;73(10):3155–3167. | ||
Trerotola M, Li J, Alberti S, Languino LR. Trop-2 inhibits prostate cancer cell adhesion to fibronectin through the β1 integrin-RACK1 axis. J Cell Physiol. 2012;227(11):3670–3677. | ||
Nakashima K, Shimada H, Ochiai T, et al. Serological identification of Trop2 by recombinant cDNA expression cloning using sera of patients with esophageal squamous cell carcinoma. Int J Cancer. 2004;112(6):1029–1035. | ||
Ohmachi T, Tanaka F, Mimori K, et al. Clinical significance of Trop2 expression in colorectal cancer. Clin Cancer Res. 2006;12(10):3057–3063. | ||
Wang J, Day R, Dong Y, Weintraub SJ, Michel L. Identification of Trop-2 as an oncogene and an attractive therapeutic target in colon cancers. Mol Cancer Ther. 2008;7(2):280–285. | ||
Kong JS, Kim HJ, Kim MJ, et al. The significance of Trop2 expression in predicting BRAF mutations in papillary thyroid carcinoma. J Pathol Transl Med. 2018;52(1):14–20. | ||
Zhao W, Kuai X, Zhou X, et al. Trop2 is a potential biomarker for the promotion of EMT in human breast cancer. Oncol Rep. 2018;40(2):759–766. | ||
Gu QZ, Nijiati A, Gao X, et al. Trop2 promotes cell proliferation and migration in osteosarcoma through PI3K/AKT signaling. Mol Med Rep. 2018;18(2):1782–1788. | ||
Jordheim LP, Chettab K, Cros-Perrial E, Matera EL, Dumontet C. Unexpected growth-promoting effect of oxaliplatin in excision repair cross-complementation group 1 transfected human colon cancer cells. Pharmacology. 2018;102(3–4):161–168. | ||
Zhang L, Yang G, Zhang R, et al. Curcumin inhibits cell proliferation and motility via suppression of Trop2 in bladder cancer cells. Int J Oncol. 2018;53(2):515–526. | ||
Mangino G, Grazia Capri M, Barnaba V, Alberti S. Presentation of native Trop-2 tumor antigens to human cytotoxic T lymphocytes by engineered antigen-presenting cells. Int J Cancer. 2002;101(4):353–359. | ||
Lin H, Zhang H, Wang J, et al. A novel human Fab antibody for Trop2 inhibits breast cancer growth in vitro and in vivo. Int J Cancer. 2014;134(5):1239–1249. | ||
Ikeda M, Yamaguchi M, Kato K, et al. Pr1E11, a novel anti-Trop-2 antibody isolated by adenovirus-based antibody screening, recognizes a unique epitope. Biochem Biophys Res Commun. 2015;458(4):877–882. | ||
Son S, Shin S, Rao NV, et al. Anti-Trop2 antibody-conjugated bioreducible nanoparticles for targeted triple negative breast cancer therapy. Int J Biol Macromol. 2018;110:406–415. | ||
Strop P, Tran TT, Dorywalska M, et al. RN927C, a site-specific Trop-2 antibody-drug conjugate (ADC) with enhanced stability, is highly efficacious in preclinical solid tumor models. Mol Cancer Ther. 2016;15(11):2698–2708. | ||
Goldenberg DM, Cardillo TM, Govindan SV, Rossi EA, Sharkey RM. Trop-2 is a novel target for solid cancer therapy with sacituzumab govitecan (IMMU-132), an antibody-drug conjugate (ADC). Oncotarget. 2015;6(26):22496–22512. | ||
Sharkey RM, McBride WJ, Cardillo TM, et al. Enhanced delivery of SN-38 to human tumor xenografts with an anti-Trop-2-SN-38 antibody conjugate (sacituzumab govitecan). Clin Cancer Res. 2015;21(22):5131–5138. | ||
Cardillo TM, Govindan SV, Sharkey RM, Trisal P, Goldenberg DM. Humanized anti-Trop-2 IgG-SN-38 conjugate for effective treatment of diverse epithelial cancers: preclinical studies in human cancer xenograft models and monkeys. Clin Cancer Res. 2011;17(10):3157–3169. | ||
Han C, Bellone S, Schwartz PE, et al. Sacituzumab Govitecan (IMMU-132) in treatment-resistant uterine serous carcinoma: a case report. Gynecol Oncol Rep. 2018;25:37–40. | ||
Cardillo TM, Sharkey RM, Rossi DL, et al. Synthetic lethality exploitation by an anti-Trop-2-SN-38 antibody-drug conjugate, IMMU-132, plus PARP inhibitors in BRCA1/2-wild-type triple-negative breast cancer. Clin Cancer Res. 2017;23(13):3405–3415. | ||
Chang CH, Wang Y, Zalath M, et al. Combining ABCG2 inhibitors with IMMU-132, an anti-Trop-2 antibody conjugate of SN-38, overcomes resistance to SN-38 in breast and gastric cancers. Mol Cancer Ther. 2016;15(8):1910–1919. | ||
Farivar TN, Najafipour R, Johari P. Nano – drug delivery of apoptosis activator 2 to AGS cells by liposomes conjugated with anti-Trop2 antibody. N Am J Med Sci. 2012;4(11):582–585. | ||
Liu D, Cardillo TM, Wang Y, et al. Trop-2-targeting tetrakis-ranpirnase has potent antitumor activity against triple-negative breast cancer. Mol Cancer. 2014;13:53. | ||
Bailly C, Cléry PF, Faivre-Chauvet A, et al. Immuno-PET for clinical theranostic approaches. Int J Mol Sci. 2016;18(1):57. | ||
Sharkey RM, van Rij CM, Karacay H, et al. A new Tri-Fab bispecific antibody for pretargeting Trop-2-expressing epithelial cancers. J Nucl Med. 2012;53(10):1625–1632. | ||
van Rij CM, Lütje S, Frielink C, et al. Pretargeted immuno-PET and radioimmunotherapy of prostate cancer with an anti-Trop-2 X anti-HSG bispecific antibody. Eur J Nucl Med Mol Imaging. 2013;40(9):1377–1383. | ||
van Rij CM, Frielink C, Goldenberg DM, et al. Pretargeted immunoPET of prostate cancer with an anti-Trop-2 X anti-HSG bispecific antibody in mice with PC3 xenografts. Mol Imaging Biol. 2015;17(1):94–101. | ||
van Rij CM, Frielink C, Goldenberg DM, et al. Pretargeted radioimmunotherapy of prostate cancer with an anti-Trop-2×anti-HSG bispecific antibody and a (177)Lu-labeled peptide. Canc Biother Radiopharm. 2014;29(8):323–329. | ||
Campos MP, Konecny GE. The target invites a foe: antibody-drug conjugates in gynecologic oncology. Curr Opin Obstet Gynecol. 2018;30(1):44–50. | ||
King GT, Eaton KD, Beagle BR, et al. A phase 1, dose-escalation study of PF-06664178, an anti-Trop-2/Aur0101 antibody-drug conjugate in patients with advanced or metastatic solid tumors. Invest New Drugs. 2018;36(5):836–847. | ||
Cardillo TM, Govindan SV, Sharkey RM, et al. Sacituzumab govitecan (IMMU-132), an anti-Trop-2/SN-38 antibody-drug conjugate: characterization and efficacy in pancreatic, gastric, and other cancers. Bioconjug Chem. 2015;26(5):919–931. | ||
Starodub AN, Ocean AJ, Shah MA, et al. First-in-human trial of a novel anti-Trop-2 Antibody-SN-38 conjugate, sacituzumab govitecan, for the treatment of diverse metastatic solid tumors. Clin Cancer Res. 2015;21(17):3870–3878. | ||
Bardia A, Mayer IA, Diamond JR, et al. Efficacy and safety of anti-Trop-2 antibody drug conjugate sacituzumab govitecan (IMMU-132) in heavily pretreated patients with metastatic triple-negative breast cancer. J Clin Oncol. 2017;35(19):2141–2148. | ||
Ambrogi F, Fornili M, Boracchi P, et al. Trop-2 is a determinant of breast cancer survival. PLoS One. 2014;9(5):e96993. | ||
Faltas B, Goldenberg DM, Ocean AJ, et al. Sacituzumab govitecan, a novel antibody–drug conjugate, in patients with metastatic platinum-resistant urothelial carcinoma. Clin Genitourin Canc. 2016;14(1):e75–e79. | ||
Heist RS, Guarino MJ, Masters G, et al. Therapy of advanced non-small-cell lung cancer with an SN-38-anti-Trop-2 drug conjugate, sacituzumab govitecan. J Clin Oncol. 2017;35(24):2790–2797. | ||
Gray JE, Heist RS, Starodub AN, et al. Therapy of small cell lung cancer (SCLC) with a topoisomerase-I-inhibiting antibody-drug conjugate (ADC) targeting Trop-2, sacituzumab govitecan. Clin Cancer Res. 2017;23(19):5711–5719. | ||
Rudin CM, Pietanza MC, Bauer TM, et al. Rovalpituzumab tesirine, a DLL3-targeted antibody-drug conjugate, in recurrent small-cell lung cancer: a first-in-human, first-in-class, open-label, phase 1 study. Lancet Oncol. 2017;18(1):42–51. | ||
Janku F, Hong DS, Fu S, et al. Assessing PIK3CA and PTEN in early-phase trials with PI3K/AKT/mTOR inhibitors. Cell Rep. 2014;6(2):377–387. | ||
Troiani T, Vecchione L, Martinelli E, et al. Intrinsic resistance to selumetinib, a selective inhibitor of MEK1/2, by cAMP-dependent protein kinase A activation in human lung and colorectal cancer cells. Br J Cancer. 2012;106(10):1648–1659. | ||
Guerra E, Trerotola M, Tripaldi R, et al. Trop-2 induces tumor growth through Akt and determines sensitivity to Akt inhibitors. Clin Cancer Res. 2016;22(16):4197–4205. | ||
Wang X, Long M, Dong K, et al. Chemotherapy agents-induced immunoresistance in lung cancer cells could be reversed by Trop-2 inhibition in vitro and in vivo by interaction with MAPK signaling pathway. Cancer Biol Ther. 2013;14(12):1123–1132. | ||
Mojica WD, Brandwein-Weber M, Korangy EA. A case of metastatic lobular carcinoma with overexpression of trop-2: implications for the consideration of novel therapeutics. Breast J. 2017;23(6):757–759. | ||
Govindan SV, Cardillo TM, Sharkey RM, et al. Milatuzumab-SN-38 conjugates for the treatment of CD74+ cancers. Mol Cancer Ther. 2013;12(6):968–978. | ||
Okajima D, Yasuda S, Yokouchi Y, et al. Preclinical efficacy studies of DS-1062a, a novel TROP2-targeting antibody-drug conjugate with a novel DNA topoisomerase I inhibitor DXd. J Clin Oncol. 2008;36 (Suppl 15; abstract e24206). |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.