")
Back to Journals » Infection and Drug Resistance » Volume 13
Structural Genomics of repA, repB1-Carrying IncFIB Family pA1705-qnrS, P911021-tetA, and P1642-tetA, Multidrug-Resistant Plasmids from Klebsiella pneumoniae
Authors Nazir A, Zhao Y, Li M, Manzoor R, Tahir RA , Zhang X, Qing H , Tong Y
Received 16 September 2019
Accepted for publication 26 December 2019
Published 22 June 2020 Volume 2020:13 Pages 1889—1903
DOI https://doi.org/10.2147/IDR.S228704
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Amina Nazir,1,2,* Yachao Zhao,2,* Manli Li,2 Rakia Manzoor,3,4 Rana Adnan Tahir,1 Xianglilan Zhang,2 Hong Qing,1 Yigang Tong2
1Key Laboratory of Molecular Medicine and Biotherapy in the Ministry of Industry and Information Technology, Department of Biology, School of Life Sciences, Beijing Institute of Technology, Beijing, People’s Republic of China; 2State Key Laboratory of Pathogen and Biosecurity, Beijing Institute of Microbiology and Epidemiology, Beijing 100071, People’s Republic of China; 3State Key Laboratory of Molecular Development Biology, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, People’s Republic of China; 4School of Life Science, University of Chinese Academy of Sciences, Beijing 100101, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Hong Qing
Key Laboratory of Molecular Medicine and Biotherapy in the Ministry of Industry and Information Technology, Department of Biology, School of Life Sciences, Beijing Institute of Technology, Beijing, People’s Republic of China
Tel +8613264286266
Email [email protected]
Yigang Tong
State Key Laboratory of Pathogen and Biosecurity, Beijing Institute of Microbiology and Epidemiology, Beijing 100071, People’s Republic of China
Email [email protected]
Background: Multidrug-resistant plasmids carrying replication genes have been widely present in various strains of Klebsiella pneumoniae. RepA and repB1 were found in plasmids belong to the IncFIB, but their detailed structural and genomic characterization was not reported yet. This is the first study that delivers structural and functional insights of repA- and repB1-carrying IncFIB plasmids.
Methods: Klebsiella pneumoniae strains A1705, 911021, and 1642 were isolated from the human urine samples and bronchoalveolar fluids collected from different hospitals of China. Antibacterial susceptibility and plasmid transfer ability were tested to characterize the resistant phenotypes mediated by the pA1705-qnrS, p911021-tetA, and p1642-tetA. The complete nucleotide sequences of these plasmids were determined through high-throughput sequencing technology and comparative genomic analyses of plasmids belong to the same incompatibility group were executed to extract the genomic variations and features.
Results: The pA1705-qnrS, p911021-tetA, and p1642-tetA are defined as non-conjugative plasmids, having two replication genes, repA and repB1 associated with IncFIB family, and unknown incompatible group, respectively. Comparative genomic analysis revealed that relatively small backbones of IncFIB plasmids integrated massive accessory module at one “hotspot” that was located between orf312 and repB1. These IncFIB plasmids exhibited the distinct profiles of accessory modules including one or two multidrug-resistant regions, many complete and remnant mobile elements comprising integrons, transposons and insertion sequences. The clusters of resistant genes were recognized in this study against different classes of antibiotics including β-lactam, phenicol, aminoglycoside, tetracycline, quinolone, trimethoprim, sulfonamide, tunicamycin, and macrolide. It has been observed that all resistant genes were located in multidrug resistance regions.
Conclusion: It is concluded that multidrug-resistant repA and repB1-carrying IncFIB plasmids are a key source to mediate the resistance through mobile elements among Klebsiella pneumoniae. Current findings provide a deep understanding of horizontal gene transfer among plasmids of the IncFIB family via mobile elements that will be utilized in further in vitro studies.
Keywords: plasmids, repA, repB1, multidrug resistance, structural genomics, bioinformatics
Introduction
Klebsiella pneumoniae (K. pneumoniae) is a Gram-negative Enterobacteriaceae bacterium and the most concerning multidrug-resistant (MDR) pathogen in nosocomial infections.1 Due to MDR and limited treatment choices, K. pneumoniae has been associated with a high mortality rate up to 40–50%.2–6 K. pneumoniae, along with other highly important MDR pathogens, has been categorized as ESKAPE (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter aumannii, Pseudomonas aeruginosa, and Enterobacter species) organisms.7,8 Many antibiotics, including aminoglycosides, fluoroquinolones, and carbapenems, are used for the treatment of infectious diseases caused by MDR pathogens.9 However, these antibiotics are tremendously threatened by MDR strains of K. pneumoniae because they harbor plasmids encoding resistance genes that lead to the increased disease burden.10 The widespread occurrence of these resistance genes highlights the significance of the underlying mechanism that needs to be elucidated.
A bacterial strain can transfer its antibiotic-resistant traits either horizontally or vertically to its filial generations or act as a donor for mobile genetic elements (MGEs) for instance plasmids and transposons. MGEs are the main effective vehicles for antibiotic resistance horizontal transfer from one bacterial strain to other strains, species, or genera.11,12 Among all kinds of MGEs, plasmids play a key role in gene transfer processes.13,14 K. pneumoniae acquired resistance through the successful acquisition of MDR plasmids, including those classified into IncFII, IncFIIY, IncHI2, and IncX1, incompatibility groups.15–17 Plasmids harboring antimicrobial resistance markers in clinical strains are a severe threat to public health worldwide.
In the present study, three MDR plasmids pA1705-qnrS, p911021-tetA, and p1642-tetA isolated from K. pneumoniae strain A1705, 911021, and 1642, respectively, were sequenced. Each of pA1705-qnrS, p911021-tetA, and p1642-tetA harbored two different replication initiation genes repA (IncFIB-family) and repB1 of an unknown incompatibility group. Initially, repA and repB1 were found in pKPN-c22 (GenBank AC# CP009879.1) isolated from K. pneumoniae. Until now, total nine fully sequenced plasmids carrying repA and repB1 have been reported including pKPN-c22, pKPN3-307_typeA (GenBank AC# KY271404.1), pKPN3-307_TypeC (GenBank AC# KY271406.1), p6234-198.371kb (GenBank AC# CP010390.1) and pKPSH11 (GenBank AC# KT896504.1), pKPN3-307_typeD (GenBank AC# accession number: KY271407),18 pCN1_1 (GenBank AC#CP015383), pRJ119-NDM1 (GenBank AC# KX636095),19 and pKP301cro (GenBank AC# KY495890) isolated from K. pneumoniae (last accessed, 26 October 2017). However, among these plasmids, not a single one has been assigned to any incompatibility group. Although, the detailed structures of IncFIB plasmids carrying repA, repB1 have not been characterized and genomic comparison of this incompatibility group has not been performed.
The current study provides a deep understanding concerning structural genomics of pA1705-qnrS, p911021-tetA, and p1642-tetA and five other sequenced plasmids carrying repA, repB1 with the highest homology provide further insights into the incompatibility (FIB) group. The common features and differences of their backbone regions and accessory modules are extensively analyzed and elaborated. The detailed genomic characterization of MDR plasmids will contribute to improve the diagnostics and understand the epidemiological relevance of Klebsiella strains.
Materials and Methods
Bacterial Strains and Identification
K. pneumoniae A1705 and 911021 strains were isolated from the urine samples of patients attending teaching hospitals in Shenyang and Chongqing, respectively. K. pneumoniae 1642 was isolated from a bronchoalveolar lavage fluid of a patient in the 307th Hospital of the People’s Liberation Army in Beijing. Bacterial species were identified by the VITEK-2 automated system (BioMerieux Inc., Marcy-l’Etoile, France) and 16S rRNA gene sequencing.20 The multilocus sequence typing (MLST) scheme for K. pneumoniae was followed as mentioned on the homepage (http://bigsdb.pasteur.fr/klebsiella)3 by utilizing seven housekeeping genes (gapA, infB, mdh, pgi, phoE, rpoB, and tonB).21 PCR screening of ESBLs,22 carbapenem resistance,23 quinolone resistance,24 macrolide resistance25 and common tetracycline resistance genes26 was carried out for each strain. All PCR amplicons were sequenced on an ABI 3730 platform (Applied Biosystems, CA, USA).
Plasmid Transfer
Plasmid conjugal transfer was carried out by K. pneumoniae strains A1705, 911021, 1642 as donors and E.coli DH5α as the recipient. Overnight cultures of both strains (3mL), recipient and donor bacteria, were grown, mixed together and then harvested. The mixed culture was resuspended in 80ul of brain-heart infusion broth (BD Biosciences, CA, USA). The mixture was put onto a filter membrane of about 1 cm2, subsequently placed onto a plate containing brain-heart infusion agar (BD Biosciences) and subsequently incubated at 37°C for mating for 12–18 h. The bacterial culture washed from the filter membrane and spread onto Mueller-Hinton (MH) agar (BD Biosciences) plates which had 4 µg/mL ciprofloxacin and 4 µg/mL tetracycline for the screening of trans-conjugants.27 As successful conjugation was not achieved, so electroporation experiments were carried out using K. pneumonia A1705, 911021, 1642 as the donor and E.coli DH5α as the recipient. The 2µL of plasmid DNA solution was added in 50 µL of E.coli DH5α and the cell suspension was transferred into ice-cooled electroporation cuvette followed by electroporation using a single pulse at the highest setting, corresponding to, 25 µF, 200 Ω, and 2.5 kV. After an electric shock, the cell suspension was diluted immediately by adding 1 mL volume of Super Optimal Broth (SOB) liquid medium and incubated for 1 hour at 37°C in a shaker at 220 rpm/min. An appropriate amount of suspension was applied to the resistant plate and incubated overnight at 37°C. Bacterial growth was re-enriched on the new resistant plate by picking up single isolated colonies. The transformation was confirmed by selection in ciprofloxacin 4ug/mL; (A1705) or tetracycline 4ug/mL (911021 and 1642) containing medium and plasmids.28,29
Antimicrobial Susceptibility Test
Antimicrobial susceptibility to ampicillin, compound sulfamethoxazole, ciprofloxacin, levofloxacin, aztreonam, ceftazidime, piperacillin, cefazolin, cefuroxime, cefuroxime axetil, ceftriaxone, gentamicin, cefotetan, cefepime, imipenem, meropenem, nitrofurantoin, piperacillin/tazobactam, and amikacin was tested by broth dilution method and the minimum inhibitory concentration values were interpreted according to the Clinical and Laboratory Standards Institute (CLSI, 2015) guidelines.27,30
Sequencing and Sequence Assembly
Genomic DNA was extracted from each of the A1705, 911021, and 1642 isolates using a QIAGEN Blood & Cell Culture DNA Maxi Kit (Qiagen, Hilden, Germany). Genomic sequencing of A1705 and 911021 isolates was performed using a Single-MoleculeReal-Time technique on a PacBio RSII sequencer (Pacific Biosciences, CA, USA). Contigs were assembled using SMARTdenovo1.0 (https://github.com/ruanjue/smartdenovo).
Genomic DNA of the 1642 isolate was sequenced from a mate-pair library with an average insert size of 5000 bp, using a MiSeq sequencer (Illumina, CA, USA). Reads were trimmed to remove the poor quality sequences,31 afterwards the contigs were congregated using Newbler 3.0.32 Gaps among contigs were bridged by utilizing a combination of PCR and Sanger sequencing using an ABI 3730 Sequencer (Applied Biosystems).
Sequence Annotation and Comparison
Open reading frames (ORFs) and pseudogenes were predicted using RAST server33 combined with BLASTP/BLASTN34 results against UniProtKB/Swiss-Prot35 and RefSeq databases.36 Annotation of resistance genes, mobile elements, and other features were performed using online databases including ISfinder,37 ResFinder,38 and INTEGRALL.39 Gene organization diagrams were drawn in Inkscape 0.48.1 (https://inkscape.org/en/).
GenBank Accession Numbers
The complete sequences of pA1705-qnrS, p911021-tetA, and p1642-tetA were submitted to GenBank under the accession numbers MG764551, MG288679, and MF156696, respectively.
Results
Characterization of K. pneumoniae Strains
K. pneumoniae strains A1705 and 911021 were assigned to sequence type 449 and 11, respectively, while 1642 was given a new sequence type 2040, as determined by MLST. PCR screening confirmed the presence of blaKPC-2, blaNDM-1, blaOXA-1, qnrS1, oqxAB, tetA(A), tetA(D), blaCTX-M-14, blaCTX-M-15, blaTEM-1, and blaSHV-33 genes in K. pneumoniae strain A1705 while the presence of the blaKPC-2, qnrS1, oqxAB, mph(A), tetA(A), blaCTX-M-14, blaCTX-M-65, blaSHV-11, and blaTEM-1 genes in K. pneumoniae strain 911021 and the presence of blaKPC-2, qnrS1, mph(A), tetA(A), blaTEM-1, blaCTX-M-65, blaSHV-12, blaCTX-M-14 genes in K. pneumoniae strain 1642. Later, three common drug resistance genes qnrS1, tetA(A), and blaCTX-M-14 were co-transferred from K. pneumoniae strains A1705, 911021, 1642 into E.coli DH5α through electroporation, generating the transformants A1705-qnrS-DH5α, 911021-tetA-DH5α, and 1642-tetA-DH5α respectively. These results demonstrated the existence of these genes in all three plasmids of our study.
Antibiotic resistant K. pneumoniae strains (A1705, 911021, and 1642) and their respective transformants, were found to be resistant against ampicillin, ampicillin/sulbactam, compound sulfamethoxazole, piperacillin, cefazolin, cefuroxime, cefuroxime axetil, ceftriaxone, and gentamicin. Additionally, these strains also exhibited resistance against cefotetan, cefepime, nitrofurantoin, aztreonam, meropenem, imipenem, ceftazidime, piperacillin/tazobactam, levofloxacin, and ciprofloxacin. It also has been observed that 911021 and 1642 strains were resistant against amikacin, but strain A1705 was sensitive to amikacin (Table 1).
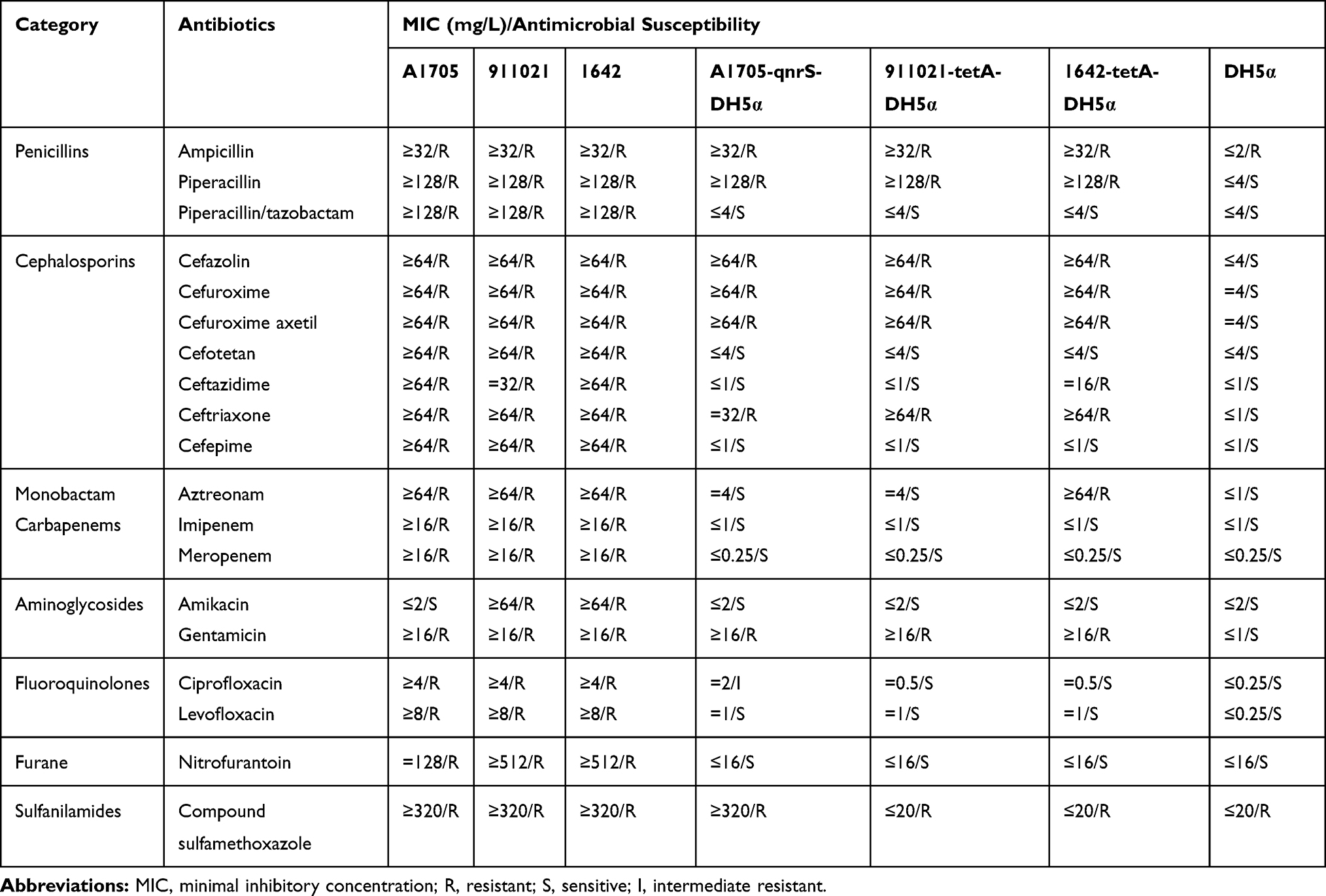 |
Table 1 Antimicrobial Drug Susceptibility Profiles |
Overview of Plasmids pA1705-qnrS, P911021-tetA, and P1642-tetA
Clinical isolates of K. pneumoniae strains A1705, 911021, 1642 were resistant to β-lactams (including carbapenems), quinolones and carried qnrS1, tetA(A), blaCTX-M-14 plasmid-borne resistance genes. Considering this, we have chosen these isolates for further genome sequencing and detailed plasmid sequence analysis.
Genome sequencing demonstrated that pA1705-qnrS, p911021-tetA, and p1642-tetA are circular DNA sequences of 271, 255bp, 238, 681bp, 237, 591bp and the average GC contents are 51.89%, 51.69%, 51.77%, annotated with 335, 301, 300 predicted ORFs, respectively (Supplementary Figure 1 and Table 2). The pA1705-qnrS, p911021-tetA, and p1642-tetA carry two replicons, an IncFIB-type of 1011-bp repA (replication initiator) gene and 1014-bp repB1 belong to unknown incompatibility group. These plasmids contain an IncFIB-type repA gene, so these are referred to as the IncFIB group.
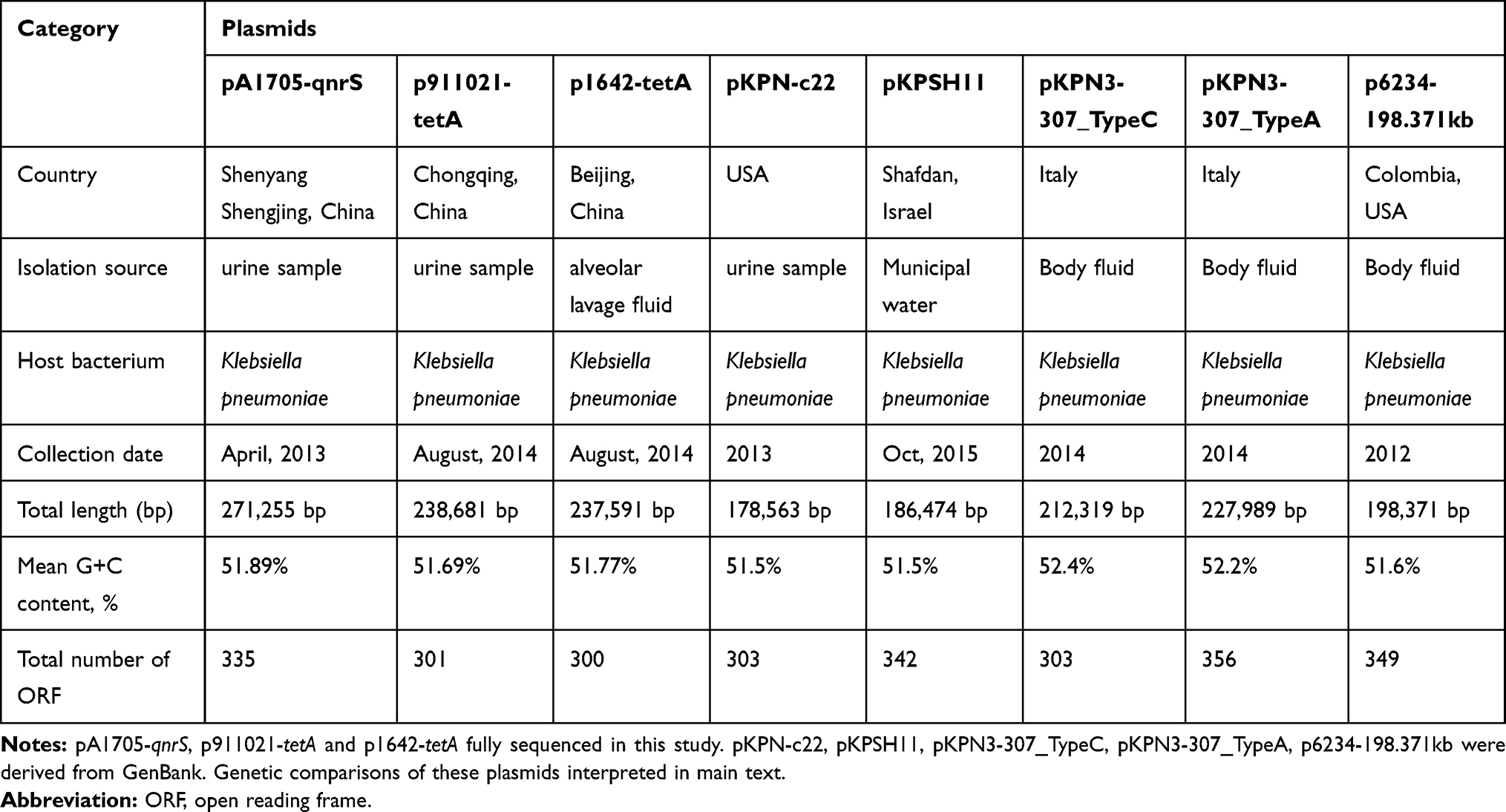 |
Table 2 Major Features of Plasmids Analyzed |
In current research, a linear genomic comparison was performed with eight plasmids including (pKPN-c22 [first discovered plasmid, harboring repA and repB1] as the reference plasmid, pA1705-qnrS, p911021-tetA, p1642-tetA [three plasmids of this study], pKPN3-307_typeA, pKPN3-307_TypeC, p6234-198.371kb, and pKPSH11) which showed the highest sequence homology to each of these three plasmids and shared replication genes (Supplementary Table S1). The molecular structure of each plasmid was divided into the conserved backbones, and a large number of separate accessory modules (Figure 1).
 |
Figure 1 Linear comparison of the sequenced plasmid. Notes: A linear comparison of the group of plasmids namely pKPN-c22, pKPSH11, p6234-198.371k, pKPN3-307_TypeC, pKPN3-307_typeA, pA1705-qnrS, p911021-tetA and p1642-tetA was performed containing the replication initiation genes repA and the repB1. Genes are indicated by arrows; genes, moving elements, and other regions are represented by different colors by function; shaded parts indicate regional nucleotide identity is greater than 95%. |
The conserved backbones comprising plasmid replication (repA and repB1), plasmid maintenance, and conjugal transfer regions. Plasmid maintenance region contained umuCD, parAB and the parB’s binding sites parC [copy number varied among plasmids (9, 9, 11, 4, 11, 9, 9, 4 for pKPN-c22, pKPSH11, p6234-198.371kb, pKPN3-307_TypeC, pKPN3-307_typeA, pA1705-qnrS, p911021-tetA, and p1642-tetA, respectively)] of the 43-bp tandem repeat (gcaGatAAcCTgAcCgAcGAtGCAGGTGGGACCGTGGTCcCAG) were found in these plasmids, a capital letter represents the base for all copies as the same and a lower letter differing bases between the copies. The conjugal transfer region composed of finO, a set of F-type conjugative DNA transfer genes including rlx, dtr, cpl, sfx, eex, tivF (tivF1 to tivF16, tivF18, and tivF19), traJQ, and trbEF (Supplementary Figure 1).
Backbone Regions of Plasmids
Pairwise comparison analyses showed that eight plasmids had >96% nucleotide identity across>77% of their backbone sequences (Supplementary Table S2). However, there were three major differences among the backbones of these eight plasmids: First, compared with pKPN-c22, the translocation of two separate regions (orf414-orf162 and orf2340-orf543) occurred in pKPSH11. Second the deletion of these two separate regions was found in pKPN3-307_TypeC, pKPN3-307_typeA, pA1705-qnrS, p911021-tetA, and p1642-tetA. Third, nine modular differences were found in the conjugal transfer region. These differences are i): the gene finO was truncated in the plasmid pKPN3-307_TypeC, pA1705-qnrS, p911021-tetA, and p1642-tetA; ii): the gene tivF3 was interrupted into two parts, namely ΔtivF3 −5ʹ and ΔtivF3-3ʹ in the plasmid pKPN-c22; iii): the gene tivF13 was truncated in the plasmid pKPN-c22; iv): the gene sfx was truncated in the plasmids pKPN3-307_typeA, pA1705-qnrS, p911021-tetA, and p1642-tetA; v): the gene rlx was interrupted into three fragments in the plasmid pKPN3-307_TypeC; vi): the gene traQ was truncated in the plasmid pKPN3-307_TypeC; vii): the gene tivF12 was interrupted in two parts, ΔtivF12-5ʹ and ΔtivF12-3ʹ in the plasmid pKPN3-307_TypeC; viii): the gene tivF18 was truncated in the plasmids pKPN3-307_TypeC, pA1705-qnrS, p911021-tetA, and p1642-tetA; ix): the gene tivF16 was truncated in plasmids pA1705-qnrS, p911021-tetA, and p1642-tetA (Figure 1).
Accessory Regions
Many different accessory modules were integrated at various sites within the backbones of these eight analyzed plasmids (Figure 1; Table 3). The IS903B, IS1X1–ISEcl1, ΔIS1X1–ΔISKpn25, CHASRI, IS1G–ISSen4, and MDR were found common in the all analyzed plasmids with slight differences. Notably, all these eight plasmids harbored resistance genes; all of which were located in the MDR regions (Table 4). MDR region from pKPN-c22, pKPSH11, p6234-198.371kb, pKPN3-307_TypeC, and pKPN3-307_typeA, and the MDR-1 region from pA1705-qnrS has a complex mosaic structure and is sequentially organized (Figure 2 and Supplementary Table S3) while the MDR regions from p1642-tetA and p911021-tetA, and the MDR-2 from pA1705-qnrS shared similar structure with slight variations (Figure 3 and Supplementary Table S4).
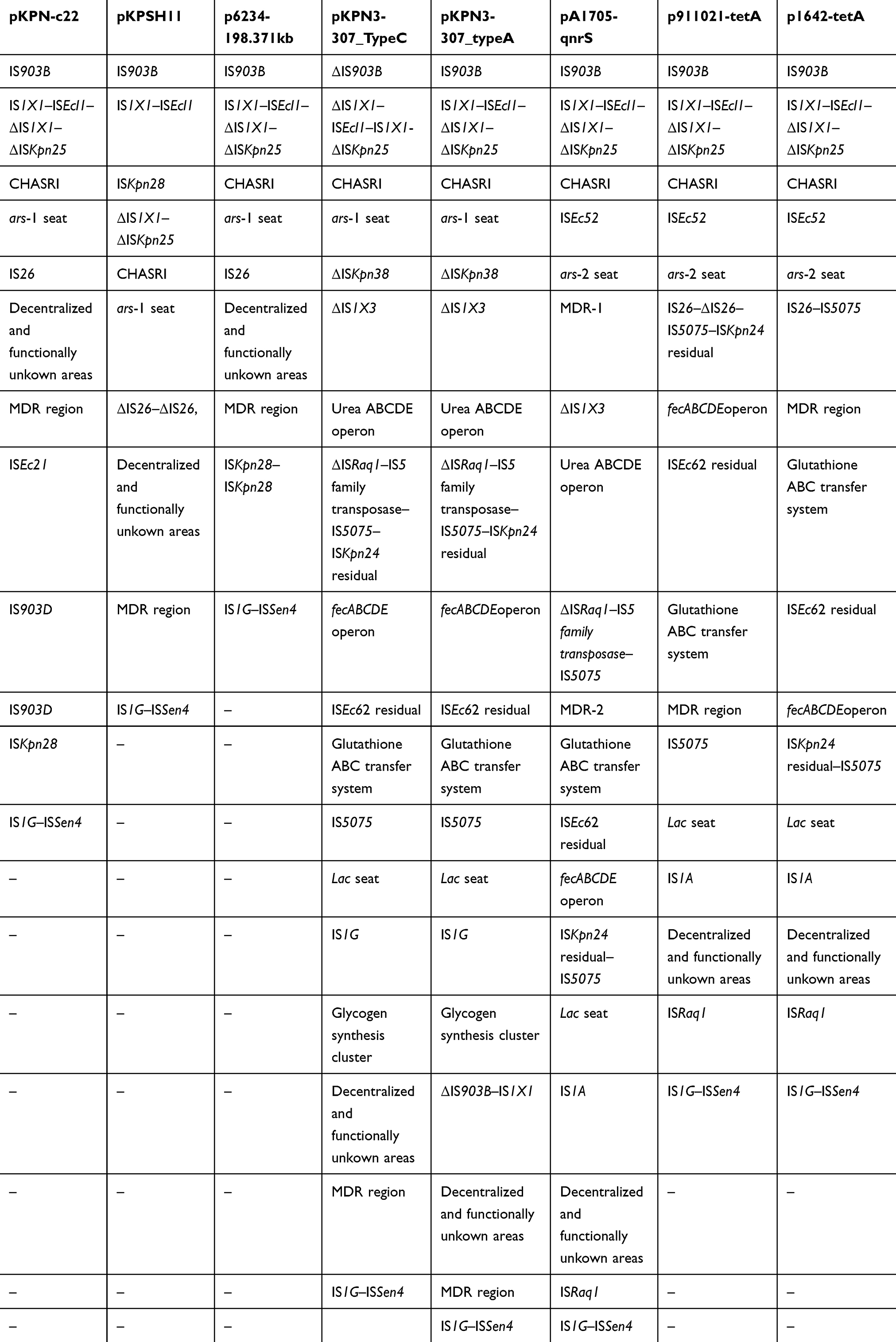 |
Table 3 Accessory Modules of Plasmids Analyzed |
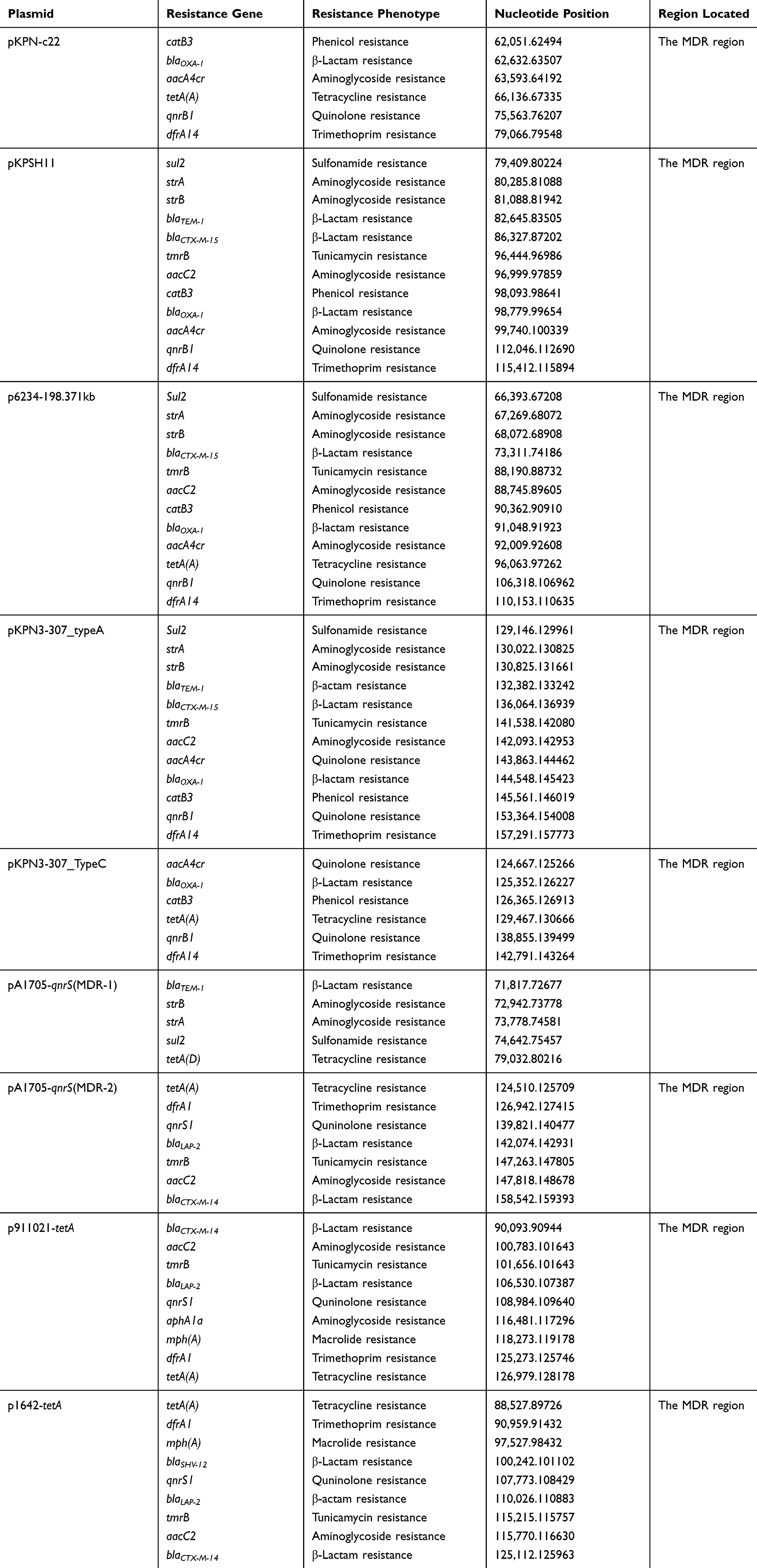 |
Table 4 Drug Resistance Genes in Plasmids Analyzed |
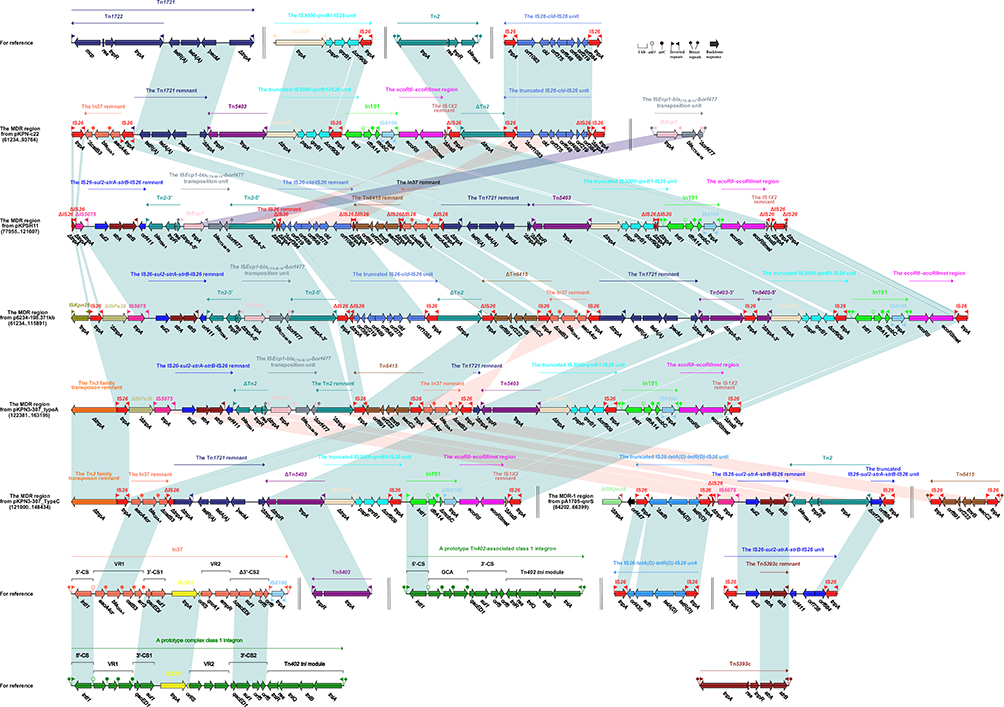 |
Figure 2 Accessory modules of the MDR region from pKPN-c22, pKPSH11, p6234-198.371kb, pKPN3-307_TypeC, and pKPN13-307_typeA and the MDR-1 region from pA1705-qnrS. Notes: Genes are indicated by arrows; genes, moving elements, and other regions are represented by different colors by function; shaded parts indicate regional nucleotide identity greater than 95%. |
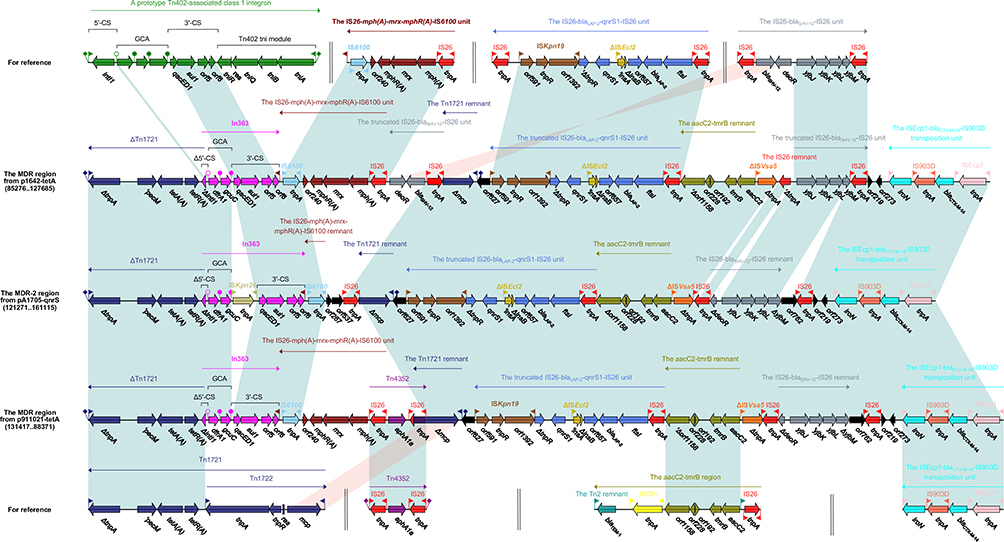 |
Figure 3 Accessory modules of the MDR region from p1642-tetA and p911021-tetA, and the MDR-2 region from pA1705-qnrS. Notes: Genes are indicated by arrows; genes, moving elements, and other regions are represented by different colors by function; shaded parts indicate regional nucleotide identity is greater than 95%. |
In191, In37 from pKPSH11/P6234-198.371kb/pKPN3-307_typeA/pKPN3-307_TypeC and pKPN-C22
The prototype Tn402-associated class 1 integron was typically divided sequentially into IRi (inverted repeat at the integrase end), the 5ʹ-conserved segment (5ʹ-CS: intI1[integrase]–attI1 [IntI1-recognizing recombination site]), the gene cassette array, the 3ʹ-conserved segment (3ʹ-CS: qacE1 [quaternary ammonium compound resistance]–sul1 [sulfonamide resistance]–orf5–orf6), the Tn402tni module (tniABQR) and IRt (inverted repeat at the tni end), bounded by 5ʹ-bp DRs. In191 from the MDR region of pKPSH11/p6234-198.371kb/pKPN3-307_typeA was derived from the prototype Tn402-associated class 1 integron. In191 had IRi, 5ʹ-CS, a single-gene (dfrA14) cassette (trimethoprim resistance), mobC–IRt–IS6100, IRt with the loss of 3ʹ-conserved segment (3ʹ-CS: ΔqacEΔ1–sul1–orf5–orf6) and tni, bounded by 5-bp DRs. In comparison with prototype Tn402-associated class 1 integron, the In191 in pKPN-c22 had undergone the loss of IRi, while the truncation at the 3ʹ-end of mobC, IS6100 and the loss IRt occurred within In191 from pKPN3-307_TypeC (Figure 2).
A complex class 1 integron, In37 was surrounded by terminal 5-bp DRs and included IRi, 5ʹ-CS, variable region 1 (VR1:aacA4cr [aminoglycoside resistance]–blaOXA-1 [β-lactam resistance]–catB3 [phenicol resistance]–arr3 [rifampin resistance]), the first copy of 3ʹ-conserved segment (3ʹ-CS1: ΔqacEΔ1[quaternary ammonium compound resistance]–sul1[sulfonamide resistance]), ISCR1, variable region 2 (VR2: qnrA1–ampR), the truncation of the second copy of 3ʹ-conserved segment (Δ3ʹ-CS2: ΔqacEΔ1–sul1–orf5–orf6), IRt (inverted repeat at the tni end), IS6100 replacing the Tn402tni module and IRt. In pKPN3-307_typeA/pKPN3-307_TypeC, In37 had undergone the segmentation into a remnant (aacA4cr–blaOXA-1–ΔcatB3), which was inverted compared with that in pKPN-c22/pKPSH11/p6234-198.371kb (Figure 2).
Tn2 and the Truncated IS26-tetA(D)-tetR(D)-IS26 Unit from MDR-1 Region of pA1705-qnrS
Tn2, a Tn3-family transposon was flanked by 5-bp DRs and showed the following modular structure: IRL–tnpA–res(resolution site)–tnpR(resolvase)–blaTEM-1 (penicillin resistance)–IRR.40 pKPN-c22 carried a 2645-bp Tn2 segment (IRL–ΔtnpA), that was also found in p6234-198.371kb, but at the opposite orientation. An intact Tn2 was identified in the MDR-1 region from pA1705-qnrS, while the Tn2 had undergone disintegration to form two parts, namely Tn2-5ʹ, Tn2-3ʹ in pKPSH11 and p6234-198.371kb. In pKPN3-307_typeA, ΔTn2 (identical to Tn2-3ʹ from pKPSH11/p6234-198.371kb) and the Tn2 remnant (ΔtnpA) were identified (Figure 2).
The IS26-tetA(D)-tetR(D)-IS26 unit [also designated Tntet(D)], derivative of the tetracycline resistance unit, was made up of two directly oriented IS26 elements flanking a central region that contained orf435, adh, tetA(D) (tetracycline efflux protein), and tetR(D) (tetracycline repressor protein).41 The truncated IS26-tetA(D)-tetR(D)-IS26 unit found in MDR-1 region of pA1705-qnrS comprised adh, tetA(D), and tetR(D) with IS26 on both sides (Figure 2).
Tn5403 from P6234-198.371kb and pKPN3-307_TypeC
Tn5403, a Tn3 family unit transposon with typical 38-bp IRs at both ends, was first found in a K. pneumoniae strain and displayed the structure tnpA–tnpR.42 Tn5403 was intact in pKPN-c22, pKPSH11, and pKPN3-307_typeA, but has been found in fragmented forms; Tn5403-5ʹ and Tn5403-3ʹ in p6234-198.371kb and had undergone the loss of IRR_Tn5403 and a 5-bp truncation of tnpR at the 3ʹ-end in pKPN3-307_TypeC (Figure 2).
Tn1721 Remnant and Unit IS3000–qnrB1–IS26 from pKPN-C22, pKPSH11, P6234-198.371kb, and pKPN3-307_TypeC, pKPN3-307_typeA
The tetracycline resistance gene of hybridization class A [tet(A)] is associated with non-conjugative transposon Tn1721. Gram-negative bacteria obtained mobilizable plasmids from different sources having complete or truncated variants of Tn1721.43 Tn1721 is a member of Tn21 subgroup of Tn3-family unit bacterial transposons44 having three 38 bp inverted repeats, with the following modular structure: IRR–mcp(methyl-accepting chemotaxis protein)–res (resolution site)–tnpR(resolvase)–tnpA(transposase)–IRL-1–tetR(A)–tetA(A)–pecM(PecM-like protein)–tnpA–IRL-2. Tn1721 consisted of a “basic transposon,” Tn1722, which was cryptic, with the modular structure IRR–tnpAR–mcp–IRL. The tetR(A)–tetA(A)–pecM module remained same in each of pKPN-c22, pKPSH11, p6234-198.371kb, and pKPN3-307_TypeC, while in pKPN3-307_typeA, only a small segment of ΔtnpA (transposase) was discovered (Figure 2).
The IS3000–qnrB1–IS26 unit, a qnrB1 transmission vehicle, was first found in pPKPN1 from PittNDM01.45 The pspF–qnrB1–Δorf909 was bracketed by two different ISs, namely IS3000 and IS26. In each of pKPN-c22, pKPSH11, p6234-198.371kb, pKPN3-307_typeA, and pKPN3-307_TypeC, the IS3000 within this unit was truncated at the 5ʹ-terminal due to the transposition of Tn5403. The Δorf909 was truncated in pKPN-c22/pKPSH11/p6234-198.371kb, while in pKPSH11, IS26 was truncated as well (Figure 2).
Tn6415 from pKPN3-307_typeA and pKPSH11
Tn6415, first appraised in plasmid unitig_2 (GenBank accession number CP021536) from Escherichia coli strain AR_0119, was an IS26-flanked composite transposon. It was bracketed by 8-bp DRs and arranged in the following order: IS26, aacC2 (aminoglycoside resistance), tmrB (tunicamycin resistance), orf222, orf891, and IS26. Tn6415 had undergone the deletion of DRs in pKPN3-307_typeA, the truncation upstream of right-hand IS26 in p6234-198.371kb and further truncation at the 5ʹ-end of the right-hand IS26, 3ʹ-end of left-hand IS26 in pKPSH11 (Figure 2).
IS26-Cld-IS26 Unit in pKPN-C22, P6234-198.371kb, and pKPSH11
In previous studies, genomic analyses suggested that chlorite dismutase originates in the perchlorate respiratory islands, from which it is transferred into transposons in the chlorate respirers. The cld gene behaves like a rogue and is predicted as a result of horizontal gene transfer.46 We found IS26-cld-IS26 unit carrying cld (chlorite dismutase) and some genes of unknown function, encircled by two directly orientated IS26 elements. In pKPN-c22, a 38-bp deletion was observed at the 5ʹ-end of orf1083. In p6234-198.371kb, the left-hand IS26 within this unit was truncated, while in pKPSH11, the left-hand IS26 was further truncated, and the truncation of IS26 in orf384 at the right-hand was discovered (Figure 2).
Tn5393c and IS26–Sul2–strA–strB–IS26 Unit from pA1705-qnrS, pKPSH11, P6234-198.371kb and pKPN3-307_typeA
Tn5393c, an active transposon, containing strA–strB was reported for the first time in the family of Tn5393.47 It showed the modular structure as follows; tnpA-res-tnpR-strA-strB with direct and inverted repeats at both ends. The IS26–sul2–strA–strB–IS26 unit was an IS26-flanked transposition unit and first discovered in pK245 from K. pneumoniae strain NK245.48 It was comprised of remnant (strA–strB–IRR_Tn5393c) of the Tn5393 family unit transposon Tn5393c, the upstream sul2, and the downstream orf411–orf738–orf684–IS26. In pA1705-qnrS, this unit had undergone segmentation into two parts of different lengths (1478 and 3152 bp) resulting from the insertion of an intact Tn2. Only a 3607-bp remnant (sul2–strA–strB–IRR_Tn5393c–orf411) was observed in pKPSH11/p6234-198.371kb/pKPN3-307_typeA (Figure 2).
In363, Tn1721 Remnant and IS26-blaSHV-12-IS26 Unit from P1642-tetA/P911021-tetA and pA1705-qnrS
In363, a dfrA1 gene and a gene array of undefined function was classified in class 1 integrons.49 We observed the In363 from the MDR region of p1642-tetA/p911021-tetA and found that it was similar to the prototype Tn402-associated class 1 integron. In363 had Δ5ʹ-CS (ΔintI1–attI1), GCA (dfrA1 [trimethoprim resistance]–gcuC [unknown function]), 3ʹ-CS and IRt. Compared with the prototype Tn402-associated class 1 integron, In363 has gone through the truncation of intI1, the loss of IRi and tni. The In363 in the MDR-2 region of pA1705-qnrS have been found to be inserted with ISKpn26 (Figure 3).
In plasmids pA1705-qnrS, p1642-tetA, and p911021-tetA, Tn1721 have been dislocated into two fragments, namely the Tn1721 remnant (IRL_Tn1721–Δmcp) and ΔTn1721[tetR(A)–tetA(A)–pecM–ΔtnpA–IRR-2_Tn1721]. The 5-bp DRs are locating upstream of the Tn1721 remnant and downstream of ΔTn1721 indicate that the disruption occurred after transposition. Remarkably, the lengths of the Tn1721 remnants varied among plasmids (1455-, 1601-, and 1707-bp for p1642-tetA, MDR-2 pA1705-qnrS, and p911021-tetA respectively) (Figure 3).
IS26-blaSHV-12-IS26 locus was found in different genetic environments, including class 1 integrons and transposons.50 These elements could play an important role in the spread of blaSHV-12 and are suggestive of multiple recombination events. IS26-blaSHV-12-IS26 carried blaSHV-12–deoR-yjbJ–yjbK–yjbL–yjbM genes, encircled by IS26 at both ends. Two fragments (IS26–blaSHV-12–deoR and yjbJ–yjbK–yjbL–yjbM–IS26) were identified in the opposite direction in p1642-tetA. Nonetheless, a single portion (ΔdeoR–yjbJ–yjbK–yjbL–ΔyjbM) of the IS26-blaSHV-12-IS26 unit was found in pA1705qnrS/p911021-tetA (Figure 3).
IS26-mph(A)–mrx–mphR(A)–IS6100, IS26-blaLAP-2–qnrS-IS26 and aacC2–tmrB Transposition Units from P1642-tetA/P911021-tetA and pA1705-qnrS
The IS6100 transposable unit is based on IS26, IS6100 and includes mph(A)–mrx–mphR(A) operon.51 The insertion sequences IS26 and IS6100 belong to the IS6 family, contain almost identical IRs of 14 bp in length. The complete IS26–mph(A)–mrx–mphR(A)–IS6100 transposable unit appears in plasmid p1642-tetA/p911021-tetA, but only a small residue was found in plasmid pA1705-qnrS (Figure 3).
The IS26-blaLAP-2–qnrS-IS26 transposition unit was first discovered in plasmid pE66An in E. coli E66An (AC # HF545433). The transposable unit structure is IS26, ftsI, blaLAP-2, orf657, ΔISEcl2, qnrS1, ΔtnpR, ISKpn19, and IS26. The unit lost its IS26 at its right-hand end in plasmids p1642-tetA, pA1705-qnrS, and p911021-tetA (Figure 3).
Furthermore, The complete aacC2–tmrB transposition unit was found in at least 13 plasmids, including plasmid pEl1573 in Enterobacter cloacae El1573.52 The structure of the aacC2-tmrB unit is IS26, Tn2 residual, aacC2, tmrB, orf192, orf228, orf1158, ISCfr1, and Tn2 residues. There is only a small portion (aacC2–tmrB–orf192–orf228–Δorf1158) of this unit that was found in each p1642-tetA, pA1705-qnrS, and p911021-tetA (Figure 3).
Discussion
Antibiotics such as aminoglycosides, β-lactams, quinolones and macrolides, consumption have been spread globally, but the emergence of MDR K. pneumoniae often lead to the failure of clinical antibiotics. It is important to distinguish and elucidate the drug resistance genes in plasmid and genetic environment, thereby illustrating the drug resistance mechanism mediated by MDR plasmids. There are many mechanisms involved in K. pneumoniae resistance to multiple antibiotics, among those, horizontal transfer of resistance genes is the most significant.
All of the eight analyzed plasmids from environmental and clinical isolates belonging to K. pneumoniae were obtained from different countries including China, USA, Italy, and Israel. The transferability of plasmids through conjugation was still unsuccessful due to the lack of some conjugal transfer genes even after the repetitive attempts of plasmid transformation. However, the mobility of plasmids was verified by the transformation experiments. The findings showed that pA1705-qnrS, p911021-tetA, and p1642-tetA could be transferred from K. pneumoniae strains A1705, 911021, 1642 into A1705-qnrS-DH5α, 911021-tetA-DH5α, and 1642-tetA-DH5α through electroporation. Although conjugation is necessary for plasmid maintenance, it has been found that compensatory adaptation plays a possible role in plasmid stability by eliminating the plasmid carriage cost together with positive selection for antibiotic resistance. Compensatory adaptation is sufficient to maintain the plasmid stability and possibly explain the reasons behind the existence of non-conjugative plasmids.53 Bacteria become resistant by picking up such MDR plasmids carrying resistance genes.
The comparison of pKPN-c22 (reference plasmid) backbone structure and other plasmids demonstrate genetic conservation in terms of gene contents and organization even omitting the deletion and inversion of some segments. Interestingly, backbones of plasmids could integrate a large number of accessory modules, mostly integrated at one “hotspot”, located between orf312 and repB1. It is interesting that the insertion of MDR regions occurs at the same site in the backbone of all plasmids, but occasionally with the insertion of two MDR regions. It is worth determining whether there are specific mechanisms associated with this incompatibility group plasmid that promote their involvement in the complex processes of acquisition of foreign genetic material.
The distribution of antibiotic resistance genes, particularly multidrug resistance genes, via transposable elements is an important concerning issue globally. In the current study, we found that pKPN-c22, pA1705-qnrS, p911021-tetA, p1642-tetA, pKPN3-307_typeA, pKPN3-307_TypeC, p6234-198.371kb, and pKPSH11 resistant plasmids encompass genes that are involved in resistance against seven different classes of antibiotics including aminoglycosides, quinolones, sulfonamides, tetracycline, trimethoprim, tunicamycin, phenicol, and macrolides. Notably, the presence of redundant resistance genes formed highly resistant strains A1705, 911021, and 1642 correspond to the classes of antibiotics, including aminoglycosides (aacC2), quinolones (qnrB1), sulfonamides (sul2), and macrolides (mph(A)). This resistance limits the selection of antibiotics for the therapies of infections caused by these bacterial strains.
As the acquisition of multiple-resistance genes is associated with a variety of mobile elements, such as insertion sequences, integrons (In363, In37, In191) and transposons (Tn4352, Tn1721, Tn6415, Tn2) which mediate the gene transfer events. The presence of mobile elements containing antibiotic resistance genes is a concern since it can promote the dissemination of resistance. This study also demonstrated the various truncated versions of mobile elements that carried the multiple resistance genes. These findings could explain the evolution of these eight plasmids through complex transposition and homologous recombination events.
Conclusion
Comparative genomic analysis of non-conjugative, repA, repB1-carrying plasmids pA1705-qnrS, p911021-tetA, p1642-tetA were carried out to determine the structural insights of these plasmids. The backbone regions of IncFIB plasmids are small as compared to the accessory regions, and the accessory regions are composed of a large number of mobile elements. Multidrug resistance mediated through these MGE’s, which contained the resistant genes, among K. pneumoniae strains. Stable inheritance and replication of these plasmids are promoted by the coordination of replicons with maintenance gene sets and the conjugation regions. This study provides a detailed genetic characterization of IncFIB plasmids, an important route for horizontal transfer of the resistance genes through mobile elements among IncFIB-family plasmids. Additionally, current findings provided a primary cause of Enterobacteriaceae epidemiology, especially MDR K. pneumoniae. The prevalence of resistant IncFIB plasmids carrying repA and repB1 at various geographic areas is still required to determined from clinical settings cultures especially those from immuno-compromised patients. Moreover, the identification and evaluation of specific factors and underlying mechanisms associated in the spread of these resistant plasmids also needs to be elucidated. There is still extensive experimental, clinical, and Bioinformatics techniques are required to reduce the dissemination of virulence and antimicrobial resistant plasmids in hospital settings. Furthermore, epidemiological studies and regular inspection of repA and repB1-carrying IncFIB plasmids will be of great importance.
Ethics Statement
Ethics approval and informed consent were not required. All the bacterial isolates involved in this study were part of the routine hospital laboratory procedure.
Acknowledgments
This research was supported by a grant from The National Key Research and Development Program of China (2015AA020108, 2016YFC1202705, SKLPBS1518, AWS16J020 and AWS15J006), the National Natural Science Foundation of China (81572045, 81672001, and 81621005), National Science and Technology Major Project (Grant No. 2018ZX10201001). The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
Disclosure
The authors state that they have no conflicts of interest.
References
1. Yan J, Pu S, Jia X, et al. Multidrug resistance mechanisms of carbapenem resistant Klebsiella pneumoniae strains isolated in Chongqing, China. Ann Lab Med. 2017;37(5):398–407. doi:10.3343/alm.2017.37.5.398
2. Gomez-Simmonds A, Uhlemann AC. Clinical implications of genomic adaptation and evolution of carbapenem-resistant Klebsiella pneumoniae. J Infect Dis. 2017;215(suppl_1):S18–s27. doi:10.1093/infdis/jiw378
3. Jelic M, Butic I, Plecko V, et al. KPC-producing Klebsiella pneumoniae isolates in Croatia: a nationwide survey. Microb Drug Res. 2016;22(8):662–667. doi:10.1089/mdr.2015.0150
4. Navon-Venezia S, Kondratyeva K, Carattoli A. Klebsiella pneumoniae: a major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol Rev. 2017;41(3):252–275. doi:10.1093/femsre/fux013
5. Hou XH, Song XY, Ma XB, Zhang SY, Zhang JQ. Molecular characterization of multidrug-resistant Klebsiella pneumoniae isolates. Braz J Microbiol. 2015;46(3):759–768. doi:10.1590/S1517-838246320140138
6. Pitout JD, Nordmann P, Poirel L. Carbapenemase-producing Klebsiella pneumoniae, a key pathogen set for global nosocomial dominance. Antimicrob Agents Chemother. 2015;59(10):5873–5884. doi:10.1128/AAC.01019-15
7. Boucher HW, Talbot GH, Bradley JS, et al. Bad bugs, no drugs: no ESKAPE! An update from the Infectious Diseases Society of America. Clin Infect Dis. 2009;48(1):1–12. doi:10.1086/595011
8. Karlowsky JA, Hoban DJ, Hackel MA, Lob SH, Sahm DF. Resistance among Gram-negative ESKAPE pathogens isolated from hospitalized patients with intra-abdominal and urinary tract infections in Latin American countries: SMART 2013–2015. Braz J Infect Dis. 2017;21(3):343–348. doi:10.1016/j.bjid.2017.03.006
9. Paterson DL, Ko W-C, Von Gottberg A, et al. Antibiotic therapy for Klebsiella pneumoniae bacteremia: implications of production of extended-spectrum β-lactamases. Clin Infect Dis. 2004;39(1):31–37. doi:10.1086/420816
10. Jiang Y, Yu D, Wei Z, Shen P, Zhou Z, Yu Y. Complete nucleotide sequence of Klebsiella pneumoniae multidrug resistance plasmid pKP048, carrying blaKPC-2, blaDHA-1, qnrB4, and armA. Antimicrob Agents Chemother. 2010;54(9):3967–3969. doi:10.1128/AAC.00137-10
11. Woodford N, Turton JF, Livermore DM. Multiresistant gram-negative bacteria: the role of high-risk clones in the dissemination of antibiotic resistance. FEMS Microbiol Rev. 2011;35(5):736–755. doi:10.1111/j.1574-6976.2011.00268.x
12. Frost LS, Leplae R, Summers AO, Toussaint A. Mobile genetic elements: the agents of open source evolution. Nat Rev Microbiol. 2005;3(9):722. doi:10.1038/nrmicro1235
13. Johnson TJ, Wannemuehler YM, Johnson SJ, et al. Plasmid replicon typing of commensal and pathogenic Escherichia coli isolates. Appl Environ Microbiol. 2007;73(6):1976–1983. doi:10.1128/AEM.02171-06
14. Gilmour MW, Thomson NR, Sanders M, Parkhill J, Taylor DE. The complete nucleotide sequence of the resistance plasmid R478: defining the backbone components of incompatibility group H conjugative plasmids through comparative genomics. Plasmid. 2004;52(3):182–202. doi:10.1016/j.plasmid.2004.06.006
15. Carattoli A. Plasmids and the spread of resistance. Int J Med Microbiol. 2013;303(6–7):298–304. doi:10.1016/j.ijmm.2013.02.001
16. Ying J, Wu S, Zhang K, et al. Comparative genomics analysis of pKF3-94 in Klebsiella pneumoniae reveals plasmid compatibility and horizontal gene transfer. Front Microbiol. 2015;6:831. doi:10.3389/fmicb.2015.00831
17. Li R, Xie M, Lv J, Wai-Chi Chan E, Chen S. Complete genetic analysis of plasmids carrying mcr-1 and other resistance genes in an Escherichia coli isolate of animal origin. J Antimicrob Chemother. 2016;72(3):696–699.
18. Villa L, Feudi C, Fortini D, et al. Diversity, virulence, and antimicrobial resistance of the KPC-producing Klebsiella pneumoniae ST307 clone. Microb Genom. 2017;3(4). doi:10.1099/mgen.0.000110.
19. Xie L, Dou Y, Zhou K, et al. Coexistence of blaOXA-48 and truncated blaNDM-1 on different plasmids in a Klebsiella pneumoniae isolate in China. Front Microbiol. 2017;8:133. doi:10.3389/fmicb.2017.00133
20. Frank JA, Reich CI, Sharma S, Weisbaum JS, Wilson BA, Olsen GJ. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl Environ Microbiol. 2008;74(8):2461–2470. doi:10.1128/AEM.02272-07
21. Diancourt L, Passet V, Verhoef J, Grimont PA, Brisse S. Multilocus sequence typing of Klebsiella pneumoniae nosocomial isolates. J Clin Microbiol. 2005;43(8):4178–4182. doi:10.1128/JCM.43.8.4178-4182.2005
22. Dallenne C, Da Costa A, Decré D, Favier C, Arlet G. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J Antimicrob Chemother. 2010;65(3):490–495. doi:10.1093/jac/dkp498
23. Bonnedahl J, Hernandez J, Stedt J, Waldenström J, Olsen B, Drobni M. Extended-spectrum β-lactamases in Escherichia coli and Klebsiella pneumoniae in gulls, Alaska, USA. Emerg Infect Dis. 2014;20(5):897. doi:10.3201/eid2005.130325
24. Ciesielczuk H, Hornsey M, Choi V, Woodford N, Wareham D. Development and evaluation of a multiplex PCR for eight plasmid-mediated quinolone-resistance determinants. J Med Microbiol. 2013;62(12):1823–1827. doi:10.1099/jmm.0.064428-0
25. Wang L, Liu L, Liu D, et al. The first report of a fully sequenced resistance plasmid from Shigella boydii. Front Microbiol. 2016;7:1579. doi:10.3389/fmicb.2016.01579
26. Maynard C, Fairbrother JM, Bekal S, et al. Antimicrobial resistance genes in enterotoxigenic Escherichia coli O149: K91 isolates obtained over a 23-year period from pigs. Antimicrob Agents Chemother. 2003;47(10):3214–3221. doi:10.1128/AAC.47.10.3214-3221.2003
27. Zhao Y, Wang L, Zhang Z, et al. Structural genomics of pNDM-BTR harboring In191 and Tn6360, and other bla NDM-carrying IncN1 plasmids. Future Microbiol. 2017;12:1271–1281. doi:10.2217/fmb-2017-0067
28. Papagianni M, Papamichael EM. Plasmid transformation of Weissella paramesenteroides DX by electroporation. Anaerobe. 2014;30:60–64. doi:10.1016/j.anaerobe.2014.08.013
29. Benachour A, Flahaut S, Frère J, Novel G. Plasmid transfer by electroporation and conjugation in Tetragenococcus and Pediococcus genera and evidence of plasmid-linked metabolic traits. Curr Microbiol. 1996;32(4):188–194. doi:10.1007/s002849900034
30. Clinical, Institute LS. M100-S25 performance standards for antimicrobial susceptibility testing; twenty-fifth informational supplement. CLSI. 2015;35:1–240.
31. Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. 2014;30(15):2114–2120. doi:10.1093/bioinformatics/btu170
32. Margulies M, Egholm M, Altman WE, et al. Corrigendum: genome sequencing in microfabricated high-density picolitre reactors. Nature. 2006;441(7089):120. doi:10.1038/nature04726
33. Aziz RK, Bartels D, Best AA, et al. The RAST server: rapid annotations using subsystems technology. BMC Genomics. 2008;9(1):75. doi:10.1186/1471-2164-9-75
34. Boratyn GM, Camacho C, Cooper PS, et al. BLAST: a more efficient report with usability improvements. Nucleic Acids Res. 2013;41(W1):W29–W33. doi:10.1093/nar/gkt282
35. Boutet E, Lieberherr D, Tognolli M, et al. UniProtKB/Swiss-Prot, the manually annotated section of the UniProt KnowledgeBase: how to use the entry view. In: Plant Bioinformatics. Springer; 2016:23–54.doi:10.1007/978-1-4939-3167-5_2
36. O’Leary NA, Wright MW, Brister JR, et al. Reference sequence (RefSeq) database at NCBI: current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016;44(D1):D733–D745. doi:10.1093/nar/gkv1189
37. Siguier P, Pérochon J, Lestrade L, Mahillon J, Chandler M. ISfinder: the reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006;34(90001):D32–D36. doi:10.1093/nar/gkj014
38. Kleinheinz KA, Joensen KG, Larsen MV. Applying the ResFinder and VirulenceFinder web-services for easy identification of acquired antibiotic resistance and E. coli virulence genes in bacteriophage and prophage nucleotide sequences. Bacteriophage. 2014;4(2):e27943. doi:10.4161/bact.27943
39. Moura A, Soares M, Pereira C, Leitão N, Henriques I, Correia A. INTEGRALL: a database and search engine for integrons, integrases and gene cassettes. Bioinformatics. 2009;25(8):1096–1098. doi:10.1093/bioinformatics/btp105
40. Partridge SR, Zong Z, Iredell JR. Recombination in IS26 and Tn2 in the evolution of multiresistance regions carrying blaCTX-M-15 on conjugative IncF plasmids from Escherichia coli. Antimicrob Agents Chemother. 2011;55(11):4971–4978. doi:10.1128/AAC.00025-11
41. Anantham S, Harmer CJ, Hall RM. p39R861-4, a type 2 A/C2 plasmid carrying a segment from the A/C1 plasmid RA1. Microb Drug Res. 2015;21(6):571–576. doi:10.1089/mdr.2015.0133
42. Rinkel M, Hubert J-C, Roux B, Lett M-C. Transposon Tn 5403, a mobilization-helper element: complete nucleotide sequence and distribution in aquatic strains. FEMS Microbiol Ecol. 1994;15(1–2):89–95. doi:10.1111/j.1574-6941.1994.tb00233.x
43. Pasquali F, Kehrenberg C, Manfreda G, Schwarz S. Physical linkage of Tn 3 and part of Tn 1721 in a tetracycline and ampicillin resistance plasmid from Salmonella Typhimurium. J Antimicrob Chemother. 2005;55(4):562–565. doi:10.1093/jac/dkh553
44. Allmeier H, Cresnar B, Greck M, Schmitt R. Complete nucleotide sequence of Tn1721: gene organization and a novel gene product with features of a chemotaxis protein. Gene. 1992;111(1):11–20. doi:10.1016/0378-1119(92)90597-I
45. Doi Y, Hazen TH, Boitano M, et al. Whole-genome assembly of Klebsiella pneumoniae coproducing NDM-1 and OXA-232 carbapenemases using single-molecule, real-time sequencing. Antimicrob Agents Chemother. 2014;58(10):5947–5953. doi:10.1128/AAC.03180-14
46. DuBois JL, Ojha S. Production of dioxygen in the dark: dismutases of oxyanions. In: Sustaining Life on Planet Earth: Metalloenzymes Mastering Dioxygen and Other Chewy Gases. Springer; 2015:45–87.doi:10.1007/978-3-319-12415-5_3
47. Trine M, Sørum H. Functional Tn5393-like transposon in the R plasmid pRAS2 from the fish pathogen Aeromonas salmonicida subspecies salmonicida isolated in Norway. Appl Environ Microbiol. 2000;66(12):5533–5535. doi:10.1128/AEM.66.12.5533-5535.2000
48. Chen Y-T, Shu H-Y, Li L-H, et al. Complete nucleotide sequence of pK245, a 98-kilobase plasmid conferring quinolone resistance and extended-spectrum-β-lactamase activity in a clinical Klebsiella pneumoniae isolate. Antimicrob Agents Chemother. 2006;50(11):3861–3866. doi:10.1128/AAC.00456-06
49. Zautner AE, Bunk B, Pfeifer Y, et al. Monitoring microevolution of OXA-48-producing Klebsiella pneumoniae ST147 in a hospital setting by SMRT sequencing. J Antimicrob Chemother. 2017;72(10):2737–2744. doi:10.1093/jac/dkx216
50. Chen CM, Yu WL, Huang M, et al. Characterization of IS26‐composite transposons and multidrug resistance in conjugative plasmids from Enterobacter cloacae. Microbiol Immunol. 2015;59(9):516–525. doi:10.1111/1348-0421.12289
51. Noguchi N, Takada K, Katayama J, Emura A, Sasatsu M. Regulation of transcription of the mph (A) gene for Macrolide 2′-Phosphotransferase I in Escherichia coli: characterization of the regulatory gene mphR (A). J Bacteriol. 2000;182(18):5052–5058. doi:10.1128/JB.182.18.5052-5058.2000
52. Partridge SR, Ginn AN, Paulsen IT, Iredell JR. pEl1573 carrying blaIMP-4, from Sydney, Australia, is closely related to other IncL/M plasmids. Antimicrob Agents Chemother. 2012;56(11):6029–6032. doi:10.1128/AAC.01189-12
53. San Millan A, Peña-Miller R, Toll-Riera M, et al. Positive selection and compensatory adaptation interact to stabilize non-transmissible plasmids. Nat Commun. 2014;5:5208. doi:10.1038/ncomms6208
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.