")
Back to Journals » OncoTargets and Therapy » Volume 12
SNHG8 is upregulated in esophageal squamous cell carcinoma and directly sponges microRNA-411 to increase oncogenicity by upregulating KPNA2
Authors Song H, Song J, Lu L, Li S
Received 7 May 2019
Accepted for publication 10 August 2019
Published 28 August 2019 Volume 2019:12 Pages 6991—7004
DOI https://doi.org/10.2147/OTT.S214881
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tohru Yamada
This paper has been retracted.
Huali Song, 1 Jinxia Song, 2 Lianwei Lu, 3 Shoubo Li 4
1Department of Gastroenterology, Sunshine Union Hospital, Weifang, Shandong 261061, People’s Republic of China; 2Department of Oncology, Qingdao Eighth People’s Hospital, Qingdao, Shandong 266100, People’s Republic of China; 3Department of Imaging, Binhai Hospital, Weifang People’s Hospital, Weifang, Shandong 262737, People’s Republic of China; 4Department of Thoracic Surgery, People’s Hospital of Weifang Binhai Economic and Technological Development Zone, Weifang, Shandong 262737, People’s Republic of China
Correspondence: Shoubo Li
Department of Thoracic Surgery, People’s Hospital of Weifang Binhai Economic and Technological Development Zone, 05441 Xihai Road, Weifang, Shandong 262737, People’s Republic of China
Tel +86 1 356 269 9251
Email [email protected]
Background: The long noncoding RNA, small nucleolar RNA host gene 8 (SNHG8), is upregulated in multiple human cancer types. However, whether SNHG8 is aberrantly expressed in esophageal squamous cell carcinoma (ESCC) and its biological functions have yet to be elucidated. Thus, we aimed to determine the expression status of SNHG8 in ESCC, explore the effects of SNHG8 on the oncogenicity of ESCC, and investigate the potential underlying mechanisms.
Methods: SNHG8 expression in ESCC tissues and cell lines was determined via reverse-transcription quantitative polymerase chain reaction. The actions of SNHG8 on the malignant characteristics of ESCC were explored using CCK-8 assay, flow-cytometric analysis, Transwell migration and invasion assays, and tumor xenografts in nude mice.
Results: SNHG8 expression was significantly higher in ESCC tissues and cell lines. High SNHG8 expression was revealed to closely correlate with primary tumor invasion depth, lymph node metastases, TNM stage, and worse overall survival among patients with ESCC. Functional investigation showed that ablation of SNHG8 notably restricted ESCC cell proliferation, migration, and invasion while inducing apoptosis in vitro and hindered tumor growth in vivo. In the meantime, SNHG8 acted as a molecular sponge of microRNA-411 (miR-411) in ESCC. Furthermore, miR-411 exerted a tumor-suppressive effect on ESCC cells, and karyopherin alpha 2 (KPNA2) turned out to be a direct target gene of miR-411. Restoring KPNA2 expression neutralized the inhibitory effects of miR-411 overexpression on the malignant behaviors of ESCC cells. Moreover, silencing of miR-411 abrogated the influence of SNHG8 downregulation in ESCC cells.
Conclusion: SNHG8 may play oncogenic roles in the malignancy of ESCC by sponging miR-411 to increase KPNA2 expression. The SNHG8–miR-411–KPNA2 pathway may be a novel target for the treatment of patients with ESCC and offer potential biomarkers for the diagnosis and prognosis of ESCC.
Keywords: small nucleolar RNA host gene 8, microRNA-411, esophageal squamous cell carcinoma, karyopherin alpha 2
Introduction
Esophageal cancer ranks the seventh among most frequent cancer types and the sixth among the leading causes of cancer-related deaths globally.1 Esophageal cancer has two major histopathological subtypes: squamous cell carcinoma and adenocarcinoma.2 Esophageal squamous cell carcinoma (ESCC), which arises from the squamous epithelium of the esophagus, is the predominant histological type and accounts for ~95% of cases of esophageal cancer.3 Various risk factors, such as smoking, consumption of pickled vegetables and mycotoxin-contaminated foods, excessive drinking, and hot drinks, have been identified as significant contributors to oncogenesis, including progression of ESCC.4 Recently, the advances in the diagnostic and therapeutic approaches notably improved the prognosis of patients with ESCC; however, the clinical outcomes of patients with this disease remain unfavorable, with a 5-year survival rate of less than 21%.5 Therefore, exploring the mechanisms underlying ESCC formation and progression is of great importance for the identification of attractive therapeutic targets, which may improve the prognosis of patients with ESCC.
Long noncoding RNAs (lncRNAs) are a group of RNA molecules consisting of over 200 nucleotides and do not encode a protein.6 It is well-known that lncRNAs function as regulators of gene expression through diverse mechanisms, including genomic interactions, protein amounts, miRNA competition, and chromatin modifications.7,8 Recent reports proved that lncRNAs have crucial roles in diverse biological and pathological behaviors, particularly in carcinogenesis and cancer progression.9,10 Notably, many lncRNAs are abnormally expressed in ESCC, and their aberrant expression has been reported to be closely related to the malignancy of ESCC.11 For instance, LINC01980,12 SNHG6,13 and AK00179614 are upregulated in ESCC and play oncogenic roles in the malignant phenotypes. On the contrary, neighboring enhancer of FOXA2 (NEF),15 growth arrest–specific 5 (GAS5),16 and FER1L417 are downregulated in ESCC and can restrain cancer progression. Thus, lncRNAs may be promising targets for the treatment of patients with ESCC.
MicroRNAs (miRNAs) are a family of naturally occurring single-stranded noncoding short RNAs comprising of approximately 20–22 nucleotides.18 They are capable of recognizing and directly binding to the 3′ untranslated regions (3′-UTRs) of their target mRNAs via classic base pairing, resulting in mRNA degradation and/or translation inhibition.19 MiRNAs have emerged as important players with tumor-suppressive or oncogenic roles in tumorigenesis and tumor progression.20 Extensive studies have shown that a plethora of miRNAs are dysregulated in ESCC, and the deregulated miRNAs are implicated in the pathogenesis and progression of ESCC by modulating multiple pathological processes.21–23 Therefore, in-depth research into the regulatory roles of miRNAs in ESCC may provide novel opportunities for identifying effective techniques for the diagnosis and therapy of patients with ESCC.
Many recent studies indicate that SNHG8 is overexpressed in multiple types of human cancer,24–26 suggesting that this lncRNA may promote the aggressiveness of these cancers. However, whether SNHG8 is aberrantly expressed in ESCC and its biological functions have yet to be elucidated. Thus, in this study, we aimed to determine the expression status of SNHG8 in ESCC and elucidate its clinical value among patients with ESCC. In addition, the effects of SNHG8 on the oncogenicity of ESCC and the potential underlying mechanisms of these effects were explored in a series of experiments. Our study may provide a novel theoretical basis for understanding the molecular events responsible for ESCC pathogenesis.
Materials and methods
Collection of tissue specimens
In total, 51 patients diagnosed with ESCC who underwent surgical resection before receiving radiotherapy or chemotherapy at People’s Hospital of Weifang Binhai Economic and Technological Development Zone were chosen for our study. All tissue samples, including ESCC and matched adjacent normal tissue samples, were rapidly frozen in liquid nitrogen, and then stored in a −80 °C freezer for further use. We got the approval of the study protocol from the Ethics Committee of People’s Hospital of Weifang Binhai Economic and Technological Development Zone. Written informed consent was obtained from all the participators. All the protocols were in accordance with the Declaration of Helsinki.
Cell culture
Four human ESCC cell lines, Eca109, KYSE70, KYSE150, and TE-1, were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). A normal human esophageal epithelial cell line (HET-1A) was obtained from the American Type Culture Collection (ATCC). All the aforementioned cell lines were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% (v/v) heat-inactivated fetal bovine serum (FBS) and 1% (v/v) streptomycin/penicillin mixture (all from Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), and grown in a humidified 37 °C incubator supplied with 5% of CO2.
Oligonucleotides, plasmids, and cell transfection
Small interfering RNA (siRNA) targeting human lncRNA SNHG8 (si-SNHG8) or scrambled oligonucleotides (si-scramble) were chemically synthesized by RiboBio (Guangzhou, China). MiR-411 mimics, miRNA mimics negative control (miR-NC), miR-411 inhibitor and NC inhibitor were bought from GenePharma (Shanghai, China). KPNA2 overexpression plasmid pcDNA3.1-KPNA2 (pc-KPNA2) and the empty pcDNA3.1 plasmid were constructed by the Chinese Academy of Sciences (Changchun, China). Cells were seeded in 6-well plates and subsequently transfected with the above-mentioned molecular constructs using the Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA). After different periods of incubation, the transfected cells were harvested and used for functional experiments.
Reverse-transcription quantitative polymerase chain reaction (RT-qPCR)
Isolation of total RNA was carried out using the RNeasy Mini Kit (Qiagen, Valencia, CA, USA). For quantification of SNHG8 and KPNA2 expression, a PrimeScript® RT Reagent Kit (Takara Biotechnology Co., Ltd., Dalian, China) was employed to reversely transcribe total RNA into complementary DNA (cDNA). The generated cDNA was then subjected to qPCR using SYBR® Premix Ex TaqTM II (Takara Biotechnology Co., Ltd.). To measure miR-411 expression, the miScript Reverse Transcription Kit and miScript SYBR Green PCR Kit (both from Qiagen GmbH, Hilden, Germany) were utilized for reverse transcription and qPCR, respectively. GAPDH served as the internal control of SNHG8 and KPNA2, whereas U6 small nuclear RNA served as the internal reference for the expression of miR-411. Relative gene expression was calculated with the 2−ΔΔCt method.
Cell counting kit-8 (CCK-8) assay
After 24 h transfection, cells were collected and resuspended in the culture medium. In total, 100 µl of cell suspension containing 2 × 103 cells was inoculated into 96-well plates. Cells were incubated in the humidified incubator at 37 °C and 5% CO2 to determine the cellular proliferation at specific time points: 0, 1, 2, and 3 days after inoculation. At every time point, 10 µl of the CCK-8 solution (Dojindo Molecular Technologies, Inc., Kumamoto, Japan) was added into each well, followed by incubation at 37 °C for another 2 h. The absorbance of each well was detected at a 450 nm wavelength on a microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Cell apoptosis evaluation by flow-cytometric analysis
The apoptosis of transfected cells was determined by means of the annexin V fluorescein isothiocyanate (FITC) apoptosis detection kit (Biolegend, San Diego, CA, USA). Transfected cells were collected into a centrifuge tube and then washed twice with ice-cold phosphate-buffered saline (PBS; Gibco; Thermo Fisher Scientific, Inc.). After that, cells were resuspended in 100 µL of binding buffer, followed by staining with 5 µL of annexin V and 5 µL of propidium iodide (PI). After a reaction at room temperature for 15 min in darkness, the percentage of apoptotic cells was analyzed by flow cytometry (FACScan; BD Biosciences, Bedford, MA, USA).
Transwell migration and invasion assays
The capacity for cell migration was assessed using Transwell chambers (8 µm diameter; Corning Inc., Corning, NY, USA). After the transfected cells were cultured for 48 h, the cells were harvested, washed with PBS, and resuspended in DMEM without FBS. In total, 100 μL of suspension containing 5 × 104 cells was added into the upper chambers. The Transwell chambers were placed into a 24-well plate that had already been covered with 500 μL of DMEM containing 10% of FBS. At 24 h after inoculation, nonmigratory cells were gently removed, and the migratory cells were fixed with 100% methanol, stained with 0.5% crystal violet, washed with PBS, and imaged using an inverted microscope (Olympus Corporation, Tokyo, Japan). The Transwell invasion assays were similar to the migration assay except that the chambers were precoated with Matrigel (BD Biosciences). The migratory and invasive abilities were measured by counting respectively the migratory and invading cells.
Tumor xenografts in nude mice
Four- to 6-week-old female BALB/c nude mice were bought from the Experimental Animal Center of Jilin University (Changchun, China) and maintained under pathogen-free conditions. Equal numbers of si-SNHG8–transfected or si-scramble–transfected Eca109 cells were resuspended in 100 μl of FBS-free DMEM and next inoculated into nude mice by way of subcutaneous injections. The width and length of tumor xenografts was monitored every 2 days starting 2 weeks after the injection. All nude mice were euthanized by cervical dislocation 4 weeks after injection, and tumor xenografts were resected and stored for further use. The volume of tumor xenografts was calculated via the formula: volume =0.5× (length × width2). The animal experiment was approved by the Research Ethics Committee of People’s Hospital of Weifang Binhai Economic and Technological Development Zone, and was carried out following the Animal Protection Law of the People’s Republic of China-2009.
Prediction of target genes of SNHG8 and miR-411
StarBase 3.0 (http://starbase.sysu.edu.cn/) and LncBase Experimental v.2 (http://carolina.imis.athena-innovation.gr/diana_tools/web/index.php?r=lncbasev2%2findex-experimental) were utilized for the prediction of the binding site in SNHG8 for miR-411. The potential target of miR-411 was analyzed by bioinformatics analysis. The analysis was carried out in TargetScan (http://www.targetscan.org/vert_72/), StarBase 3.0, and miRDB (http://mirdb.org/).
Luciferase reporter assay
The wild-type (wt) SNHG8 that contained the predicted miR-411–binding site and mutant (mut) SNHG8 was chemically synthesized by GenePharma and integrated into pMIR-REPORT luciferase reporter plasmids (Promega Corporation, Madison, WI, USA) to produce the pMIR-SNHG8-wt (SNHG8-wt) and pMIR-SNHG8-mut (SNHG8-mut) reporter plasmids. The reporter plasmids, KPNA2-wt and KPNA2-mut, were also generated by GenePharma. For the reporter assay, cells were seeded in 24-well plates. When cells grew to 70% confluence, the reporter plasmids were cotransfected with miR-411 mimics or miR-NC by means of the Lipofectamine 2000 reagent. After 48 h transfection, transfected cells were processed for the detection of luciferase activity in a Dual-Luciferase Reporter Assay System (Promega Corporation). The relative luciferase activity was normalized to Renilla luciferase activity.
RNA immunoprecipitation (RIP) assay
The Magna RIP RNA-Binding Protein Immunoprecipitation Kit (EMD Millipore, Billerica, MA, USA) was employed to determine the interaction between miR-411 and SNHG8 in ESCC cells. Briefly, cell lysates were incubated with RIP buffer containing magnetic beads conjugated with a human anti-Argonaute 2 (Ago2) antibody or normal immunoglobulin G (IgG). After that, total RNA was isolated and then subjected to the analysis of miR-411 and SNHG8 expression by RT-qPCR.
Western blot analysis
Total protein was extracted with RIPA lysis and extraction buffer (Thermo Fisher Scientific, MA, USA), and its concentration was measured with the Bicinchoninic Acid Assay Kit (Beyotime Institute of Biotechnology, Haimen, China). Equal amounts of protein were loaded and separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis in a 10% gel, followed by transferring to a polyvinylidene difluoride membrane (Millipore, Bedford, MA, USA) and blocking with 5% fat-free milk diluted in Tris-buffered saline containing 0.1% of Tween 20. After that, the membranes were incubated with primary antibodies against KPNA2 (ab170495; Abcam, Cambridge, UK) or GAPDH (ab128915; Abcam) at 4 °C overnight. After three washes, a goat anti-rabbit horseradish peroxidase–conjugated secondary antibody (ab205718; Abcam) was incubated with the membranes. Finally, the protein signals were visualized using the Enhanced Chemiluminescence Western Blotting Kit (Beyotime Institute of Biotechnology). Densitometric analysis of the protein signals was performed in Quantity One software, version 4.62 (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
Two-tailed Student’s t-test was conducted to evaluate the difference between two groups. The comparison between multiple groups was conducted by one-way analysis of variance followed by Dunnett’s post hoc test. The correlation between SNHG8 and clinical parameters of the patients with ESCC was examined by the χ2 test. The overall survival rate was determined by the Kaplan–Meier method, and the results were assessed for statistical significance by the logrank test. P<0.05 was assumed to indicate a statistically significant difference.
Results
SNHG8 expression is high in ESCC and is correlated with clinical parameters
To evaluate the potential involvement of SNHG8 in ESCC, we first measured its expression in 51 pairs of ESCC and matched adjacent normal tissue samples by RT-qPCR. The data indicated that SNHG8 expression was much higher in ESCC tissues relative to the adjacent normal tissues (Figure 1A, P<0.05). In addition, the expression of SNHG8 was assessed in a panel of ESCC cell lines: Eca109, KYSE70, KYSE150, and TE-1. A normal human esophageal epithelial cell line (HET-1A) served as the control. Consistently with the findings in tissues, SNHG8 was found to be upregulated in the four ESCC cell lines (when compared with HET-1A cells), especially in Eca109 and TE-1 cells, which were therefore chosen for functional experiments (Figure 1B, P<0.05).
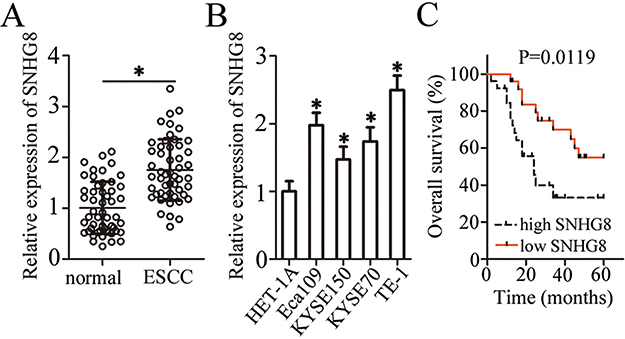 |
Figure 1 SNHG8 expression is high in ESCC tissues and cell lines. (A) RT-qPCR was carried out for the quantification of SNHG8 expression in 51 pairs of ESCC and matched adjacent normal tissue samples. *P<0.05 vs normal tissue samples. (B) SNHG8 expression was determined by RT-qPCR in four ESCC cell lines (Eca109, KYSE70, KYSE150, and TE-1) and a normal human esophageal epithelial cell line, HET-1A. *P<0.05 vs HET-1A. (C) Comparison of overall survival between patients with ESCC harboring high or low SNHG8 expression. P=0.0119. |
We also studied the clinical value of SNHG8 in ESCC by examining the association between SNHG8 expression and clinical parameters among the 51 patients. All the patients were assigned to either a low-SNHG8 expression group (n=26) or high-SNHG8 expression group (n=25) according to the median value of SNHG8 expression in ESCC tissues. Increased SNHG8 expression was obviously correlated with primary tumor invasion depth (P=0.012), lymph node metastases (P=0.001) and TNM stage (P=0.002) but was unrelated to the age (P=0.393), gender (P=0.565), and differentiation status (P=0.173) (Table 1). Notably, patients with high SNHG8 expression had shorter overall survival relative to the patients with low SNHG8 expression (Figure 1C, P=0.0119). These observations suggested that upregulation of SNHG8 in ESCC may be closely related to cancer progression.
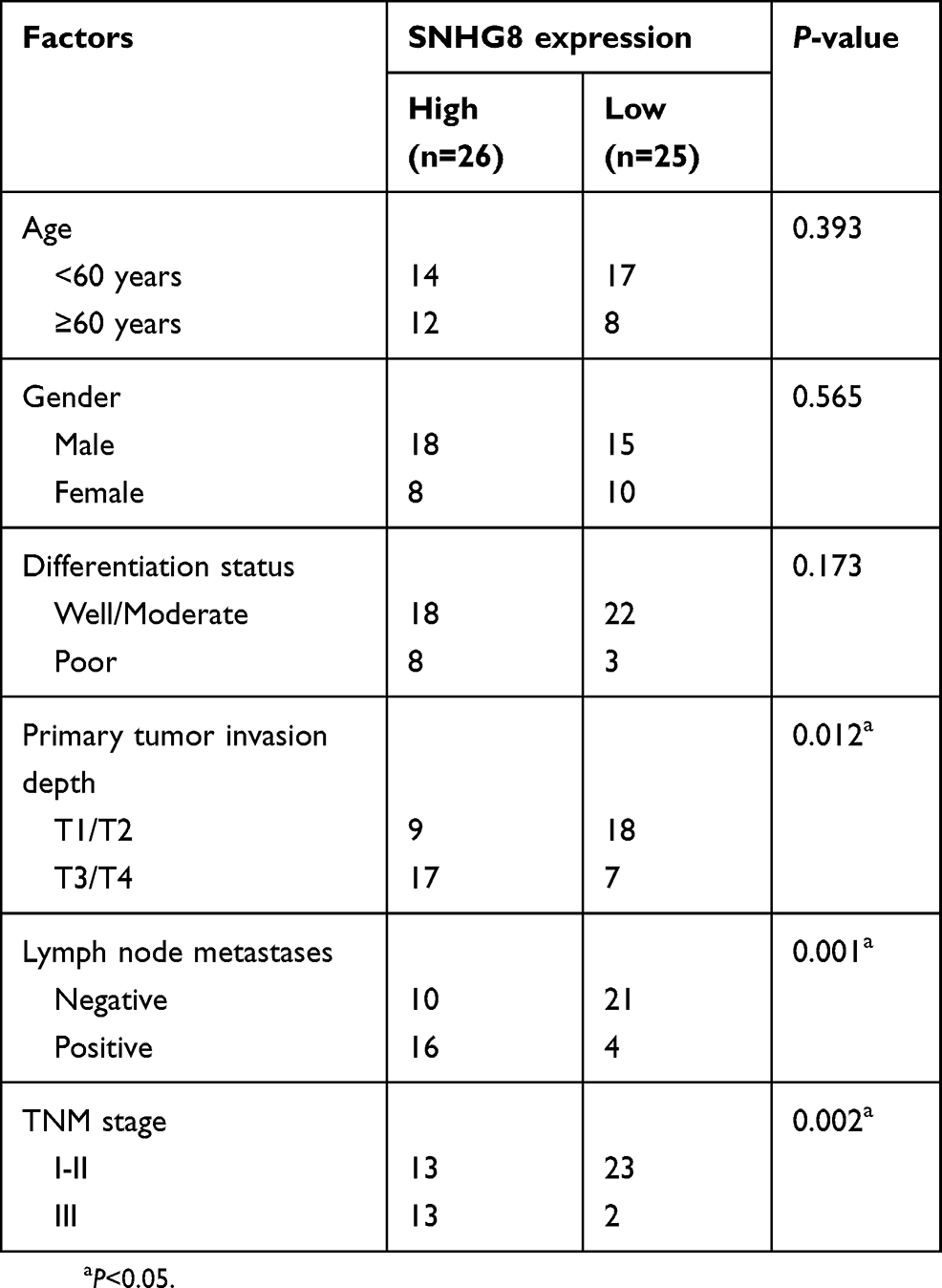 |
Table 1 The correlation between SNHG8 expression and ESCC clinicopathological factors |
SNHG8 silencing inhibits the proliferation, migration, and invasiveness and promotes the apoptosis of ESCC cells
To assess the influence of SNHG8 on the malignant characteristics of ESCC, Eca109 and TE-1 cells were transfected with si-SNHG8 or si-scramble. RT-qPCR analysis revealed that SNHG8 expression was efficiently decreased in Eca109 and TE-1 cells after si-SNHG8 transfection as compared with that in cells transfected with si-scramble, implying the successful SNHG8 silencing after transfection (Figure 2A, P<0.05). The effect of SNHG8 knockdown on the proliferation of Eca109 and TE-1 cells was determined in CCK-8 assay. Si-SNHG8 transfection markedly restrained the proliferative capacity of Eca109 and TE-1 cells compared with the si-scramble group (Figure 2B, P<0.05). In addition, the proportion of Eca109 and TE-1 apoptotic cells was substantially higher in the si-SNHG8 group than in the si-scramble group, as revealed by flow-cytometric analysis (Figure 2C, P<0.05). Furthermore, Transwell migration and invasion assays were conducted to determine whether SNHG8 is involved in the regulation of ESCC cell migration and invasion. The migratory (Figure 2D, P<0.05) and invasive (Figure 2E, P<0.05) abilities of both cell lines obviously decreased after the downregulation of SNHG8. These findings suggested that SNHG8 may exert an oncogenic action on the aggressiveness of ESCC cells in vitro.
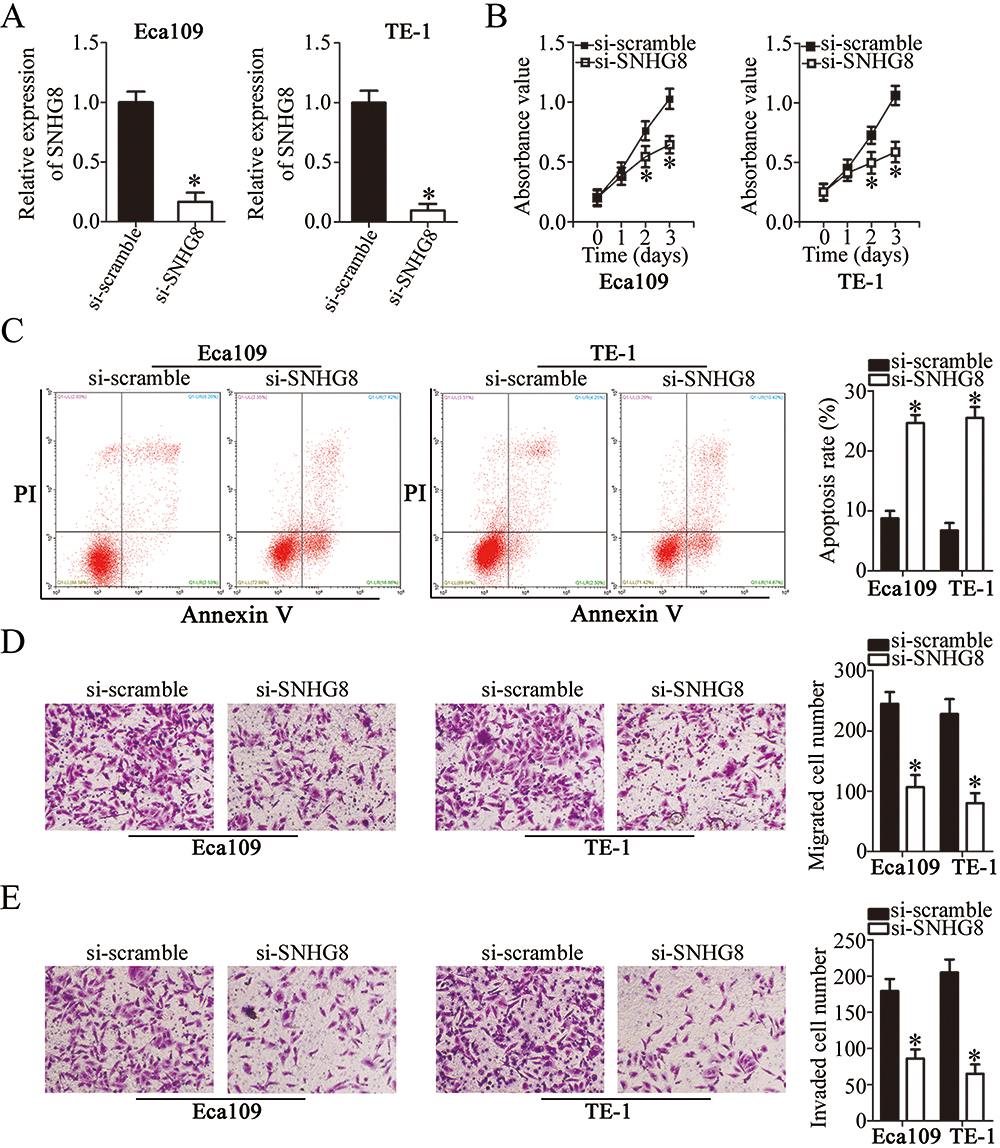 |
Figure 2 Suppression of SNHG8 inhibits Eca109 and TE-1 cell proliferation, migration, and invasion and induces their apoptosis. (A) Si-SNHG8 or si-scramble was transfected into Eca109 and TE-1 cells. SNHG8 expression following the transfection was examined by RT-qPCR. *P<0.05 vs si-scramble. (B) The CCK-8 assay was carried out to evaluate the proliferation of Eca109 and TE-1 cells after transfection of si-SNHG8 or si-scramble. *P<0.05 vs si-scramble. (C) Eca109 and TE-1 cells were transfected with si-SNHG8 or si-scramble and the proportion of apoptotic cells was measured via flow-cytometric analysis. *P<0.05 vs si-scramble. (D, E) Transwell migration and invasion assays uncovered the influence of SNHG8 silencing on the migratory capacity and invasiveness of Eca109 and TE-1 cells. *P<0.05 vs si-scramble. |
SNHG8 acts as a sponge for mir-411 in ESCC
Multiple studies have pointed out that lncRNAs may function as competitive endogenous RNAs by sponging miRNAs.27–29 Accordingly, we tested whether SNHG8 has such a role in ESCC by sponging certain miRNAs. To test our hypothesis, the potential miRNAs sponged by SNHG8 were predicted by bioinformatics analysis. Among these candidates, miR-411 (Figure 3A) was chosen for subsequent validation, based on its crucial roles in oncogenesis and tumor progression.30–40 The luciferase reporter assay was carried out to gain more insights into the potential interaction between SNHG8 and miR-411. The results indicated that miR-411 mimics–mediated upregulation of miR-411 (Figure 3B, P<0.05) significantly weakened the luciferase activity of the SNHG8-wt plasmid in Eca109 and TE-1 cells (P<0.05) but had no such effect on the SNHG8-mut plasmid (Figure 3C). In addition, the RIP assay revealed that miR-411 and SNHG8 were notably enriched in Ago2 pellets but not in the IgG (Figure 3D, P<0.05), indicating that miR-411 is an SNHG8-targeted miRNA.
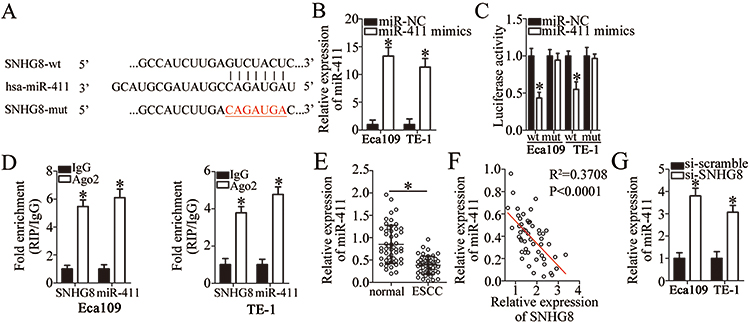 |
Figure 3 SNHG8 acts as a sponge of miR-411 in ESCC cells. (A) The binding site for miR-411 in SNHG8. Bioinformatic analysis was performed to search for the target miRNAs that can be sponged by SNHG8. (B) The expression of miR-411 in Eca109 and TE-1 cells—when they were transfected with miR-411 mimics or miR-NC—was determined via RT-qPCR. *P<0.05 vs miR-NC. (C) Eca109 and TE-1 cells were cotransfected with either miR-411 mimics or miR-NC and either SNHG8-wt or SNHG8-mut for 48 h, and next, the luciferase activities were quantified using a Dual-Luciferase Reporter Assay System. *P<0.05 vs miR-NC. (D) The RIP assay showed that both SNHG8 and miR-411 were obviously enriched in an AGO2 immunoprecipitate. IgG served as an internal control. *P<0.05 vs IgG. (E) RT-qPCR analysis revealed the expression levels of miR-411 in 51 pairs of ESCC and matched adjacent normal tissue samples. *P<0.05 vs normal tissue samples. (F) Spearman’s correlation analysis was applied to examine the association between SNHG8 and miR-411 expression in ESCC tissue samples. R2=0.3708, P<0.0001. (G) RT-qPCR evaluated miR-411 expression in Eca109 and TE-1 cells after si-SNHG8 or si-scramble transfection. *P<0.05 vs si-scramble. |
We next measured miR-411 expression in ESCC tissues and adjacent normal tissue samples by RT-qPCR. The expression of miR-411 was low in ESCC tissues as compared with adjacent normal tissues (Figure 3E, P<0.05). In addition, SNHG8 expression inversely correlated with miR-11 among ESCC tissue samples (Figure 3F; R2 = 0.3708, P<0.0001). Meanwhile, the data obtained in the RT-qPCR analysis revealed that silencing of SNHG8 increased miR-411 expression in Eca109 and TE-1 cells (Figure 3G, P<0.05). We concluded from the above results that SNHG8 acts as a sponge for miR-411 and decreases its amount in ESCC.
miR-411 exerts tumor-suppressive effects on the growth and metastasis of ESCC cells in vitro
To investigate the biological actions of miR-411 on ESCC cells, miR-411 mimics or miR-NC was introduced into Eca109 and TE-1 cells, and then a series of functional assays was conducted. CCK-8 assay and flow-cytometric analysis revealed that resumption of miR-411 expression notably decreased the proliferation (Figure 4A, P<0.05) and increased the apoptosis (Figure 4B, P<0.05) of Eca109 and TE-1 cells. We also observed in the Transwell migration and invasion assays that when Eca109 and TE-1 cells were transfected with miR-411 mimics, the migration (Figure 4C, P<0.05) and invasion (Figure 4D, P<0.05) remarkably decreased compared with that in miR-NC–transfected cells. Overall, miR-411 may have a tumor-suppressive function in the regulation of biological activities of ESCC.
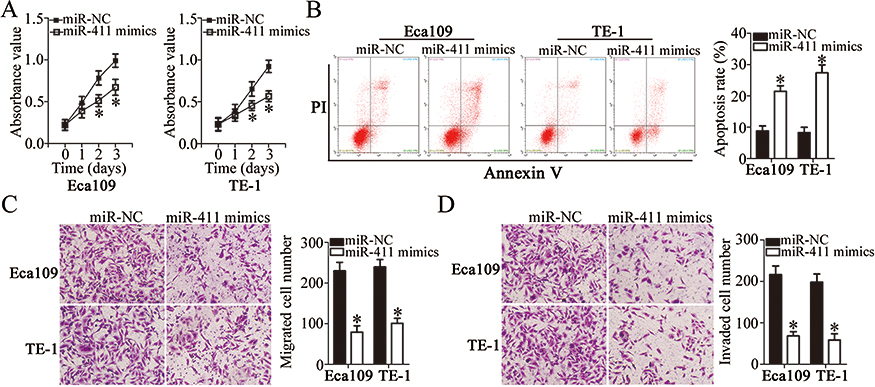 |
Figure 4 Ectopic miR-411 expression inhibits the proliferation, migration, and invasion and promotes the apoptosis of Eca109 and TE-1 cells. (A, B) The influence of miR-411 overexpression on the proliferation and apoptosis of Eca109 and TE-1 cells was tested by the CCK-8 assay and flow cytometry. *P<0.05 vs miR-NC. (C, D) The migratory and invasive abilities of miR-411–overexpressing Eca109 and TE-1 cells were investigated in Transwell migration and invasion assays. *P<0.05 vs miR-NC. |
KPNA2 mRNA is a direct target of miR-411 in ESCC
To elucidate the mechanisms underlying the activity of miR-411 in ESCC cells, online target exploratory software tools TargetScan, StarBase 3.0, and miRDB were employed to search for the putative target of miR-411. A highly conserved binging site for miR-411 in the 3′-UTR of KPNA2 was noted (Figure 5A), and this prediction was verified by the luciferase reporter assay. The KPNA2-wt and KPNA2-mut reporter plasmids were constructed, and cotransfected with miR-411 mimics or miR-NC into Eca109 and TE-1 cells. The luciferase activity of the KPNA2-wt plasmid was significantly decreased by miR-411 mimics transfection (P<0.05). In contrast, no obvious alterations in the luciferase activity of cells cotransfected with miR-411 and the KPNA2-mut plasmid were observed (Figure 5B). In addition, we determined the expression of KPNA2 mRNA and protein in miR-411–overexpressing Eca109 and TE-1 cells. Compared with the miR-NC group, the expression of KPNA2 mRNA (Figure 5C, P<0.05) and protein (Figure 5D, P<0.05) was notably suppressed in Eca109 and TE-1 cells following overexpression of miR-411. Furthermore, RT-qPCR analysis was applied to evaluate KPNA2 mRNA expression in ESCC tissue samples and revealed that KPNA2 mRNA expression was significantly higher in ESCC tissue samples than in adjacent normal tissues (Figure 5E, P<0.05). Besides, the expression of miR-411 turned out to be inversely correlated with KPNA2 mRNA expression among ESCC tissue samples (Figure 5F; R2 = 0.3186, P<0.0001). These results validated KPNA2 as a direct target of miR-411 in ESCC cells.
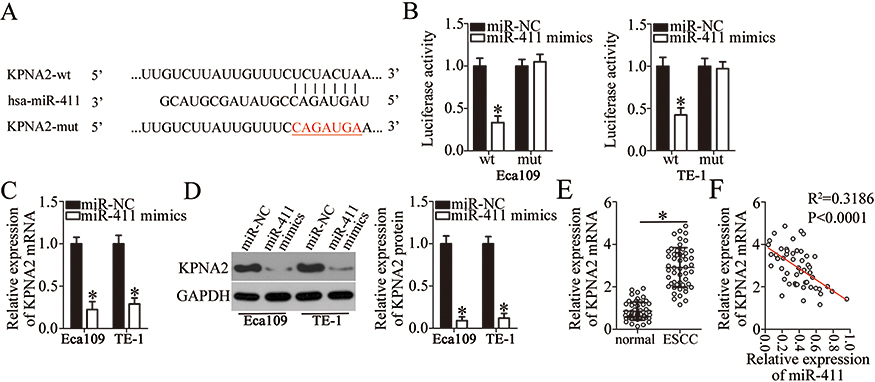 |
Figure 5 KPNA2 is validated as a direct target gene of miR-411 in ESCC cells. (A) The binding sequences for miR-411 in the wild-type KPNA2 3′-UTR. The mutant KPNA2 3′-UTR is also presented. (B) Eca109 and TE-1 cells were cotransfected with either miR-411 mimics or miR-NC and either the KPNA2-wt or KPNA2-mut reporter plasmid. The transfected cells were harvested after 48 h of incubation and then subjected to the detection of luciferase activity. *P<0.05 vs miR-NC. (C) RT-qPCR was carried out to assess KPNA2 mRNA expression in Eca109 and TE-1 cells after their transfection with miR-411 mimics or miR-NC. *P<0.05 vs miR-NC. (D) The protein expression of KPNA2 was determined by Western blotting in miR-411 mimics–transfected or miR-NC–transfected Eca109 and TE-1 cells. *P<0.05 vs miR-NC. (E) RT-qPCR analysis uncovered the expression profile of KPNA2 mRNA in the 51 pairs of ESCC and matched adjacent normal tissue samples. *P<0.05 vs normal tissue samples. (F) A negative correlation between miR-411 and KPNA2 mRNA levels in ESCC cells was demonstrated via Spearman’s correlation analysis. R2=0.3186, P<0.0001. |
Restoring KPNA2 expression neutralizes the tumor-suppressive influence of miR-411 on ESCC cells
After identifying KPNA2 as a direct target of miR-411, we determined whether KPNA2 silencing was essential for the tumor-suppressive effects of miR-411 in ESCC cells. First, miR-411–overexpressing Eca109 and TE-1 cells were transfected with KPNA2 overexpression plasmid pc-KPNA2 or pcDNA3.1 (empty vector). KPNA2 protein expression was found to be significantly downregulated in miR-411–overexpressing Eca109 and TE-1 cells but could be restored by cotransfection with pc-KPNA2 (Figure 6A, P<0.05). Subsequently, a series of functional assays revealed that the impact of miR-411 overexpression on ESCC cell proliferation (Figure 6B, P<0.05), apoptosis (Figure 6C, P<0.05), migration (Figure 6D, P<0.05), and invasion (Figure 6E, P<0.05) was partly reversed by KPNA2 restoration. Collectively, miR-411 exerted its tumor-suppressive actions on the behaviors of ESCC cells by decreasing KPNA2 expression.
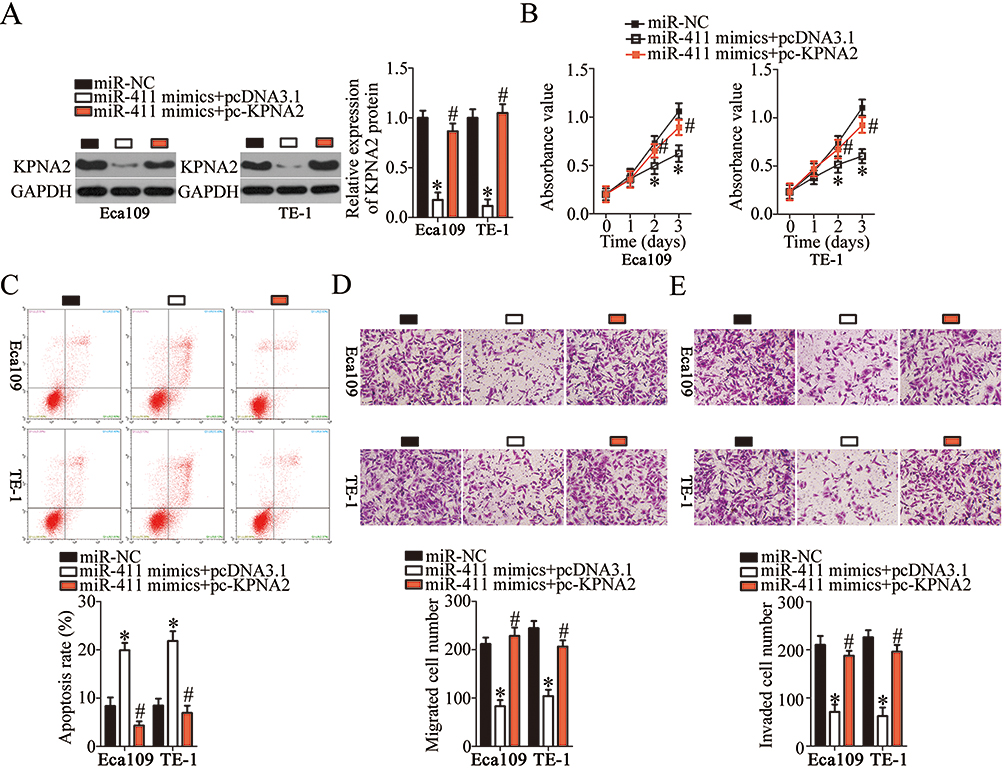 |
Figure 6 Reintroduction of KPNA2 prevents the tumor-suppressive effects of miR-411 upregulation on Eca109 and TE-1 cells. (A) MiR-411–overexpressing Eca109 and TE-1 cells were next transfected with pc-KPNA2 or pcDNA3.1. At 48 h after transfection, the cells were collected and subjected to Western blotting for the determination of KPNA2 protein expression. *P<0.05 vs miR-NC. #P<0.05 vs miR-411 mimics+pcDNA3.1. (B–E) The proliferation, apoptosis, migration, and invasiveness of Eca109 and TE-1 cells treated as described above were evaluated by the CCK-8 assay, flow cytometry, and Transwell migration and invasion assays, respectively. *P<0.05 vs miR-NC. #P<0.05 vs miR-411 mimics+pcDNA3.1. |
SNHG8 affects the malignant behaviors of ESCC cells through regulation of the miR-411–KPNA2 axis
To confirm that silencing of SNHG8 has an inhibitory influence on the malignant behaviors of ESCC cells via the miR-411–KPNA2 axis, Eca109 and TE-1 cells were cotransfected with si-SNHG8 and miR-411 inhibitor or NC inhibitor; then cell proliferation, apoptosis, migration, and invasion were analyzed by CCK-8, flow-cytometric, and Transwell migration and invasion assays, respectively. First, RT-qPCR analysis was carried out to measure miR-411 expression in Eca109 and TE-1 cells after miR-411 inhibitor or NC inhibitor transfection. The data revealed that transfection of miR-411 inhibitor significantly downregulated miR-411 in Eca109 and TE-1 cells (Figure 7A, P<0.05). Meanwhile, si-SNHG8–mediated upregulation of miR-411 (Figure 7B, P<0.05) and downregulation of KPNA2 protein (Figure 7C, P<0.05) were partially reversed in Eca109 and TE-1 cells after cotransfection with miR-411 inhibitor. In addition, cotransfection with miR-411 inhibitor attenuated the effects of SNHG8 silencing on the proliferation (Figure 7D, P<0.05), apoptosis (Figure 7E, P<0.05), migration (Figure 7F, P<0.05), and invasiveness (Figure 7G, P<0.05) of Eca109 and TE-1 cells. In a word, these results suggested that SNHG8 promotes the malignant behaviors of ESCC cells through the miR-411–KPNA2 axis.
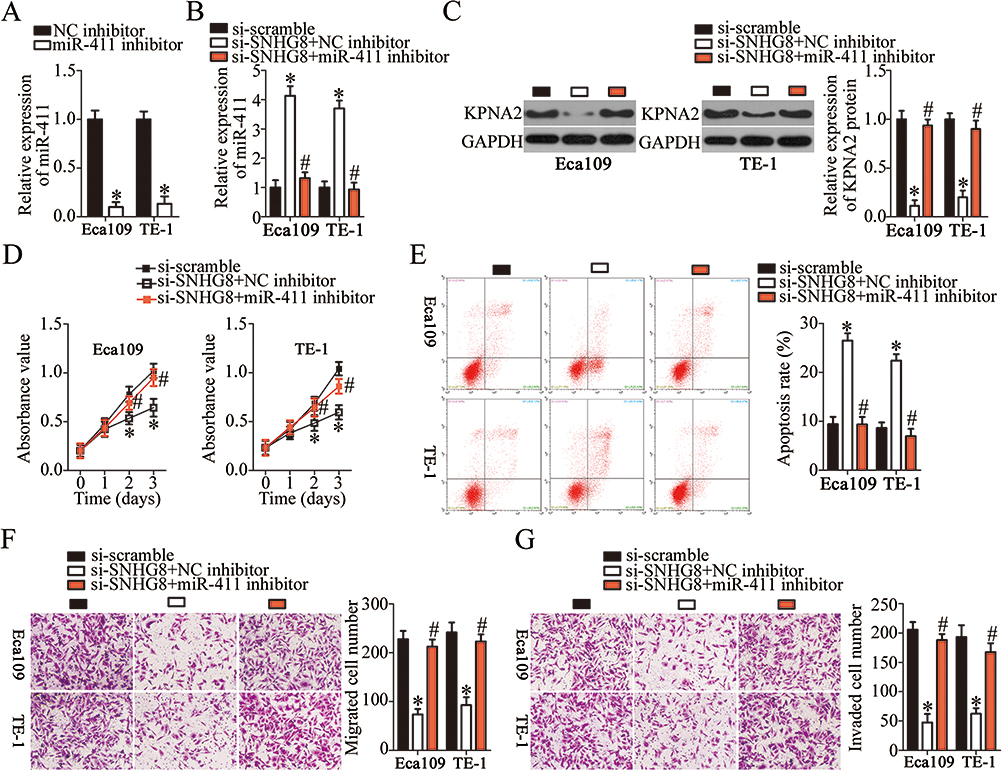 |
Figure 7 SNHG8 regulates the malignant characteristics of Eca109 and TE-1 cells through the miR-411–KPNA2 axis. (A) The transfection efficiency of the miR-411 inhibitor in Eca109 and TE-1 cells was evaluated by RT-qPCR analysis. The NC inhibitor served as the control. *P<0.05 vs NC inhibitor. (B, C) RT-qPCR and Western blotting were respectively utilized to assess miR-411 and KPNA2 protein expression in Eca109 and TE-1 cells following cotransfection with si-SNHG8 and either miR-411 inhibitor or NC inhibitor. *P<0.05 vs si-scramble. #P<0.05 vs si-SNHG8+NC inhibitor. (D–G) The proliferation, apoptosis, migration, and invasiveness of the aforementioned cells were evaluated by the CCK-8 assay, flow cytometry, and Transwell migration and invasion assays, respectively. *P<0.05 vs si-scramble. #P<0.05 vs si-SNHG8+NC inhibitor. |
SNHG8 knockdown inhibits the xenograft growth of ESCC cells in vivo
Tumor xenografts in nude mice were finally implemented to determine the influence of SNHG8 on the tumor growth of ESCC cells in vivo. Obvious inhibition of the tumor volume was observed in the si-SNHG8 group compared with the si-scramble group (Figure 8A and B, P<0.05). At 4 weeks after tumor cell inoculation, all nude mice were euthanized, and the tumor xenografts were excised and weighed. The average weight of tumor xenografts was significantly lower in the si-SNHG8 group than in the si-scramble group (Figure 8C, P<0.05). In addition, RT-qPCR analysis suggested that the tumor xenografts deriving from si-SNHG8–transfected Eca109 cells harbored SNHG8 underexpression (Figure 8D, P<0.05) and miR-411 overexpression (Figure 8E, P<0.05). Furthermore, the protein level of KPNA2 turned out to be lower in the nude mice injected with the SNHG8 knockdown Eca109 cells (Figure 8F, P<0.05). These results revealed that SNHG8 knockdown hinders tumor growth of ESCC cells in vivo via upregulation of miR-411 and a consequent decrease of KPNA2 expression.
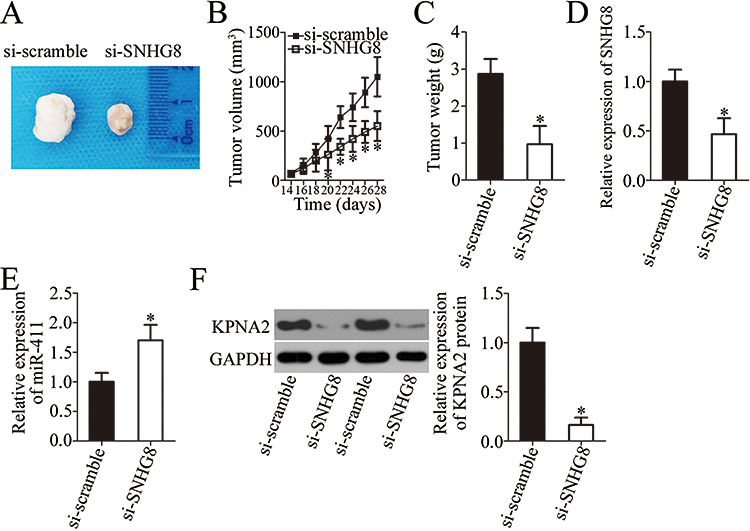 |
Figure 8 The SNHG8 knockdown impairs the tumor growth in vivo via the miR-411–KPNA2 axis. (A) The images of tumor xenografts of nude mice injected with si-SNHG8–transfected or si-scramble–transfected cells. (B) Tumor volume was examined starting at 2 weeks after inoculation of si-SNHG8–transfected or si-scramble–transfected cells. *P<0.05 vs si-scramble. (C) Tumor weight in the mice injected with cells transfected with si-SNHG8 or si-scramble after 4 weeks. *P<0.05 vs si-scramble. (D, E) The expression levels of SNHG8 and miR-411 in tumor xenografts were investigated via RT-qPCR analysis. *P<0.05 vs si-scramble. (F) Extraction of total protein was carried out, and the total-protein samples were subjected to KPNA2 protein quantification. *P<0.05 vs si-scramble. |
Discussion
An increasing number of studies has revealed the abnormal expression of lncRNAs in ESCC.41–43 The dysregulation of lncRNAs has been demonstrated to closely correlate with the malignancy of ESCC by affecting tumor processes.44–46 Therefore, investigating the roles of lncRNAs in ESCC progression may be essential for the identification of novel potential targets for anticancer therapy. In this study, we first measured the expression of SNHG8 in ESCC cells and then evaluated its clinical significance. In addition, the influence of SNHG8 suppression on the aggressive phenotypes of ESCC cells was investigated, including cellular proliferation, apoptosis, migration, and invasion in vitro as well as tumor growth in vivo. Notably, the underlying mechanisms by which SNHG8 may regulate the tumorigenic processes in ESCC were identified in detail.
SNHG8 is upregulated in pancreatic adenocarcinoma, and its upregulation significantly correlates with tumor stage and differentiation grade.24 Patients with pancreatic adenocarcinoma having higher SNHG8 expression show worse overall survival than do patients with lower expression.24 SNHG8 is also overexpressed in hepatocellular carcinoma.25 SNHG8 is validated as an independent prognostic factor of tumor recurrence in patients with hepatocellular carcinoma.25 Furthermore, SNHG8 is overexpressed in non–small cell lung cancer tissues and cell lines.26 Patients with non–small cell lung cancer overexpressing SNHG8 show shorter overall survival and shorter progression-free survival relative to the patients with low SNHG8 expression.26 However, the expression profile and clinical value of SNHG8 in ESCC have remained elusive and merit research. In this study, we found that SNHG8 expression is high in ESCC tissues and cell lines. Upregulation of SNHG8 manifested a close relation with primary tumor invasion depth, lymph node metastases and TNM stage. Patients with ESCC overexpressing SNHG8 showed worse overall survival than did patients with low SNHG8 expression. These findings suggest that SNHG8 might be a promising biomarker for the diagnosis and prognosis of ESCC.
SNHG8 plays oncogenic roles in tumorigenesis and tumor progression. For instance, downregulation of SNHG8 inhibits cell proliferation, induces cell cycle arrest, promotes apoptosis, and reduces the chemoresistance of pancreatic adenocarcinoma cells.24 In hepatocellular carcinoma, silencing of SNHG8 suppresses cell growth, invasion, and lung metastasis in vitro and in vivo.25 In non–small cell lung cancer, SNHG8 knockdown results in inhibition of cell proliferation and metastasis in vitro, increase in cell apoptosis and cell cycle arrest, and suppression in tumor growth in vivo.26 Nevertheless, the relation between SNHG8 and ESCC pathogenesis has been unknown. Herein, we revealed that SNHG8 silencing restricted cell proliferative, migratory, and invasive abilities of ESCC cells; increased the percentage of apoptotic cells; and hindered the growth of transplanted tumors in vivo. These observations proved the crucial participation of SNHG8 in the aggressive behaviors of ESCC, and therefore SNHG8 might be a novel target for the treatment of patients with this disease.
Multiple mechanisms by which SNHG8 regulates tumorigenic processes have been well elucidated, including sponging miR-149-5p and regulation of PPM1F expression in hepatocellular carcinoma,25 targeting of the miR-542-3p–CCND1–CDK6 axis in non–small cell lung cancer,26 and regulation of c-MET expression by sponging miR-152 in endometrial carcinoma.47 Herein, the oncogenic actions of SNHG8 in ESCC were mediated by sponging of miR-411 and consequent upregulation of KPNA2. KPNA2, a member of the importin α family, performs key functions in nucleocytoplasmic transport.48 KPNA2 is overexpressed in both ESCC tissues and cell lines. High KPNA2 expression shows an obvious correlation with poor differentiation, tumor depth, lymphatic invasion, venous invasion, and tumor stage.49 The prognosis of patients with ESCC overexpressing KPNA2 is shorter than that of patients with low KPNA2 expression, as revealed by univariate analysis.49 Functionally, KPNA2 plays tumor-promoting roles in regulating the malignant phenotype of ESCC through promoting cell proliferation and colony formation and by inhibiting G2–M arrest.50 Hence, the KPNA2 knockdown due to SNHG8 silencing and miR-411 upregulation could be a promising therapeutic strategy against ESCC.
MiR-411 is upregulated in hepatocellular carcinoma,30 lung cancer,31,32 and osteosarcoma;33 on the contrary, it is downregulated in breast cancer,34,35 renal cell cancer,36 colorectal cancer,37 cervical cancer,38 and bladder cancer.39,40 Nevertheless, whether miR-411 is dysregulated in ESCC and whether its dysregulation contributes to ESCC oncogenesis has been unclear. In this study, we demonstrated that miR-411 is downregulated in ESCC. MiR-411 is sponged by SNHG8 and directly downregulates KPNA2 to inhibit the oncogenicity of ESCC. Therefore, the SNHG8-miR-411-KPNA2 axis may provide a new theoretical basis for the exploration of the molecular pathogenesis of ESCC.
Conclusion
In summary, this study revealed that SNHG8 may perform oncogenic functions in the progression of ESCC by sponging miR-411 to upregulate KPNA2. SNHG8, miR-411, and KPNA2 seem to be interrelated and offer novel targets for ESCC therapy as well as potential diagnostic and prognostic biomarkers.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Nagai H, Kim YH. Cancer prevention from the perspective of global cancer burden patterns. J Thorac Dis. 2017;9(3):448–451. doi:10.21037/jtd.2017.02.75
2. Napier KJ, Scheerer M, Misra S. Esophageal cancer: A Review of epidemiology, pathogenesis, staging workup and treatment modalities. World J Gastrointest Oncol. 2014;6(5):112–120. doi:10.4251/wjgo.v6.i5.112
3. Chen W, He Y, Zheng R, et al. Esophageal cancer incidence and mortality in China, 2009. J Thorac Dis. 2013;5(1):19–26. doi:10.3978/j.issn.2072-1439.2013.01.04
4. Arnold M, Soerjomataram I, Ferlay J, Forman D. Global incidence of oesophageal cancer by histological subtype in 2012. Gut. 2015;64(3):381–387. doi:10.1136/gutjnl-2014-308124
5. Kollarova H, Machova L, Horakova D, Janoutova G, Janout V. Epidemiology of esophageal cancer–an overview article. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2007;151(1):17–20.
6. Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136(4):629–641. doi:10.1016/j.cell.2009.02.006
7. Khalil AM, Guttman M, Huarte M, et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc Natl Acad Sci U S A. 2009;106(28):11667–11672. doi:10.1073/pnas.0904715106
8. Guttman M, Rinn JL. Modular regulatory principles of large non-coding RNAs. Nature. 2012;482(7385):339–346. doi:10.1038/nature10887
9. Sarfi M, Abbastabar M, Khalili E. Long noncoding RNAs biomarker-based cancer assessment. J Cell Physiol. 2019. doi:10.1002/jcp.28417
10. Sun Y, Ma L. New Insights into Long Non-Coding RNA MALAT1 in Cancer and Metastasis. Cancers. 2019;11:2. doi:10.3390/cancers11020216
11. Fanelli GN, Gasparini P, Coati I, et al. LONG-NONCODING RNAs in gastroesophageal cancers. Noncoding RNA Res. 2018;3(4):195–212. doi:10.1016/j.ncrna.2018.10.001
12. Zhang S, Liang Y, Wu Y, et al. Upregulation of a novel lncRNA LINC01980 promotes tumor growth of esophageal squamous cell carcinoma. Biochem Biophys Res Commun. 2019;513(1):73–80.
13. Zhang Y, Li R, Ding X, Zhang K, Qin W. Upregulation of long non-coding RNA SNHG6 promote esophageal squamous cell carcinoma cell malignancy and its diagnostic value. Am J Transl Res. 2019;11(2):1084–1091.
14. Zong MZ, Shao Q, An XS. Expression and prognostic significance of long noncoding RNA AK001796 in esophageal squamous cell carcinoma. Eur Rev Med Pharmacol Sci. 2019;23(1):181–186. doi:10.26355/eurrev_201901_16763
15. Zhang J, Hu SL, Qiao CH, et al. LncRNA-NEF inhibits proliferation, migration and invasion of esophageal squamous-cell carcinoma cells by inactivating wnt/beta-catenin pathway. Eur Rev Med Pharmacol Sci. 2018;22(20):6824–6831. doi:10.26355/eurrev_201810_16150
16. Wang G, Sun J, Zhao H, Li H. Long Non-Coding RNA (lncRNA) Growth Arrest Specific 5 (GAS5) suppresses esophageal squamous cell carcinoma cell proliferation and migration by inactivating phosphatidylinositol 3-kinase (PI3K)/AKT/Mammalian Target of Rapamycin (mTOR) signaling pathway. Med Sci Monit. 2018;24:7689–7696. doi:10.12659/MSM.910867
17. Ma W, Zhang CQ, Li HL, et al. LncRNA FER1L4 suppressed cancer cell growth and invasion in esophageal squamous cell carcinoma. Eur Rev Med Pharmacol Sci. 2018;22(9):2638–2645. doi:10.26355/eurrev_201805_14958
18. Teoh SL, Das S. The role of microRNAs in diagnosis, prognosis, metastasis and resistant cases in breast cancer. Curr Pharm Des. 2017;23(12):1845–1859. doi:10.2174/1381612822666161027120043
19. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. doi:10.1016/j.cell.2009.01.002
20. Shukla GC, Singh J, Barik S. MicroRNAs: Processing, Maturation, Target Recognition and Regulatory Functions. Mol Cell Pharmacol. 2011;3(3):83–92.
21. Sun C, Zhang X, Chen Y, Jia Q, Yang J, Shu Y. MicroRNA-365 suppresses cell growth and invasion in esophageal squamous cell carcinoma by modulating phosphoserine aminotransferase 1. Cancer Manag Res. 2018;10:4581–4590. doi:10.2147/CMAR.S157858
22. Zhao Y, Wang Y, Xing G. miR-516b functions as a tumor suppressor by directly modulating CCNG1 expression in esophageal squamous cell carcinoma. Biomed Pharmacother. 2018;106:1650–1660.
23. Xiao Q, Chen T, Wu Y, et al. MicroRNA6753p promotes esophageal squamous cell cancer cell migration and invasion. Mol Med Rep. 2018;18(4):3631–3640. doi:10.3892/mmr.2018.9372
24. Song Y, Zou L, Li J, Shen ZP, Cai YL, Wu XD. LncRNA SNHG8 promotes the development and chemo-resistance of pancreatic adenocarcinoma. Eur Rev Med Pharmacol Sci. 2018;22(23):8161–8168. doi:10.26355/eurrev_201812_16508
25. Dong J, Teng F, Guo W, Yang J, Ding G, Fu Z. lncRNA snhg8 promotes the tumorigenesis and metastasis by sponging miR-149-5p and predicts tumor recurrence in hepatocellular carcinoma. Cell Physiol Biochem. 2018;51(5):2262–2274. doi:10.1159/000495871
26. Chen C, Zhang Z, Li J, Sun Y. SNHG8 is identified as a key regulator in non-small-cell lung cancer progression sponging to miR-542-3p by targeting CCND1/CDK6. Onco Targets Ther. 2018;11:6081–6090. doi:10.2147/OTT.S170482
27. Hao NB, He YF, Li XQ, Wang K, Wang RL. The role of miRNA and lncRNA in gastric cancer. Oncotarget. 2017;8(46):81572–81582. doi:10.18632/oncotarget.19197
28. Chen Y, Lin Y, Bai Y, Cheng D, Bi Z. A Long Noncoding RNA (lncRNA)-Associated Competing Endogenous RNA (ceRNA) network identifies eight lncrna biomarkers in patients with osteoarthritis of the knee. Med Sci Monit. 2019;25:2058–2065. doi:10.12659/MSM.915555
29. He M, Lin Y, Xu Y. Identification of prognostic biomarkers in colorectal cancer using a long non-coding RNA-mediated competitive endogenous RNA network. Oncol Lett. 2019;17(3):2687–2694. doi:10.3892/ol.2019.9936
30. Xia K, Zhang Y, Cao S, et al. miR-411 regulated ITCH expression and promoted cell proliferation in human hepatocellular carcinoma cells. Biomed Pharmacother. 2015;70:158–163. doi:10.1016/j.biopha.2015.01.001
31. Zhao Z, Qin L, Li S. miR-411 contributes the cell proliferation of lung cancer by targeting FOXO1. Tumour Biol. 2016;37(4):5551–5560. doi:10.1007/s13277-015-4425-8
32. Wang SY, Li Y, Jiang YS, Li RZ. Investigation of serum miR-411 as a diagnosis and prognosis biomarker for non-small cell lung cancer. Eur Rev Med Pharmacol Sci. 2017;21(18):4092–4097.
33. Xu N, Yang W, Liu Y, Yan F, Yu Z. MicroRNA-411 promoted the osteosarcoma progression by suppressing MTSS1 expression. Environ Sci Pollut Res Int. 2018;25(12):12064–12071. doi:10.1007/s11356-018-1331-9
34. Guo L, Yuan J, Xie N, et al. miRNA-411 acts as a potential tumor suppressor miRNA via the downregulation of specificity protein 1 in breast cancer. Mol Med Rep. 2016;14(4):2975–2982. doi:10.3892/mmr.2016.5645
35. Zhang Y, Xu G, Liu G, et al. miR-411-5p inhibits proliferation and metastasis of breast cancer cell via targeting GRB2. Biochem Biophys Res Commun. 2016;476(4):607–613. doi:10.1016/j.bbrc.2016.06.006
36. Zhang X, Zhang M, Cheng J, Lv Z, Wang F, Cai Z. MiR-411 functions as a tumor suppressor in renal cell cancer. Int J Biol Markers. 2017;32(4):e454–e460. doi:10.5301/ijbm.5000261
37. Zhao J, Xu J, Zhang R. MicroRNA-411 inhibits malignant biological behaviours of colorectal cancer cells by directly targeting PIK3R3. Oncol Rep. 2018;39(2):633–642. doi:10.3892/or.2017.6135
38. Shan D, Shang Y, Hu T. MicroRNA-411 inhibits cervical cancer progression by directly targeting STAT3. Oncol Res. 2019;27(3):349–358. doi:10.3727/096504018X15247361080118
39. Jin H, Sun W, Zhang Y, et al. MicroRNA-411 downregulation enhances tumor growth by upregulating MLLT11 expression in human bladder cancer. Mol Ther Nucleic Acids. 2018;11:312–322. doi:10.1016/j.omtn.2018.03.003
40. Liu Y, Liu T, Jin H, Yin L, Yu H, Bi J. MiR-411 suppresses the development of bladder cancer by regulating ZnT1. Onco Targets Ther. 2018;11:8695–8704. doi:10.2147/OTT.S173750
41. Zhang Y, Chen W, Pan T, Wang H, Zhang Y, Li C. LBX2-AS1 is activated by ZEB1 and promotes the development of esophageal squamous cell carcinoma by interacting with HNRNPC to enhance the stability of ZEB1 and ZEB2 mRNAs. Biochem Biophys Res Commun. 2019;511(3):566–572. doi:10.1016/j.bbrc.2019.02.079
42. Zhang C, Jiang F, Su C, Xie P, Xu L. Upregulation of long noncoding RNA SNHG20 promotes cell growth and metastasis in esophageal squamous cell carcinoma via modulating ATM-JAK-PD-L1 pathway. J Cell Biochem. Epub 2019 Feb 14. doi:10.1002/jcb.28444
43. Cao T, Shen J, Pan W, Li C, Qiao Z. Upregulation of long noncoding RNA ANRIL correlates with tumor progression and poor prognosis in esophageal squamous cell carcinoma. J Buon. 2018;23(6):1862–1866.
44. Abraham JM, Meltzer SJ. Long noncoding RNAs in the pathogenesis of barrett’s esophagus and esophageal carcinoma. Gastroenterology. 2017;153(1):27–34. doi:10.1053/j.gastro.2017.04.046
45. Hou X, Wen J, Ren Z, Zhang G. Non-coding RNAs: new biomarkers and therapeutic targets for esophageal cancer. Oncotarget. 2017;8(26):43571–43578. doi:10.18632/oncotarget.16721
46. Shen WJ, Zhang F, Zhao X, Xu J. LncRNAs and esophageal squamous cell carcinoma - implications for pathogenesis and drug development. J Cancer. 2016;7(10):1258–1264. doi:10.7150/jca.14869
47. Yang CH, Zhang XY, Zhou LN, et al. LncRNA SNHG8 participates in the development of endometrial carcinoma through regulating c-MET expression by miR-152. Eur Rev Med Pharmacol Sci. 2018;22(6):1629–1637. doi:10.26355/eurrev_201803_14698
48. Tseng SF, Chang CY, Wu KJ, Teng SC. Importin KPNA2 is required for proper nuclear localization and multiple functions of NBS1. J Biol Chem. 2005;280(47):39594–39600. doi:10.1074/jbc.M508425200
49. Sakai M, Sohda M, Miyazaki T, et al. Significance of karyopherin-{alpha} 2 (KPNA2) expression in esophageal squamous cell carcinoma. Anticancer Res. 2010;30(3):851–856.
50. Ma S, Zhao X. KPNA2 is a promising biomarker candidate for esophageal squamous cell carcinoma and correlates with cell proliferation. Oncol Rep. 2014;32(4):1631–1637. doi:10.3892/or.2014.3381
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.