")
Back to Journals » OncoTargets and Therapy » Volume 12
Silencing Of MAGI1 Promotes The Proliferation And Inhibits Apoptosis Of Glioma Cells Via The Wnt/β-Catenin And PTEN/AKT Signaling Pathways
Authors Lu Y, Sun W, Zhang L, Li J
Received 11 May 2019
Accepted for publication 15 October 2019
Published 13 November 2019 Volume 2019:12 Pages 9639—9650
DOI https://doi.org/10.2147/OTT.S215400
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Federico Perche
Yongzhi Lu,1 Wei Sun,2 Liang Zhang,3 Junyao Li4
1Department of Oncology, Qingdao Municipal Hospital, Qingdao, Shandong Province 266000, People’s Republic of China; 2Department of Neurology, Qingdao Third People’s Hospital, Qingdao, Shandong Province 266000, People’s Republic of China; 3Department of Critical Care Medicine, Qingdao Eighth People’s Hospital, Qingdao, Shandong Province 266000, People’s Republic of China; 4Department of Emergency, Qingdao Municipal Hospital, Qingdao, Shandong Province 266000, People’s Republic of China
Correspondence: Junyao Li
Department of Emergency, Qingdao Municipal Hospital, Qingdao, Shandong Province 266000, People’s Republic of China
Email [email protected]
Background: Membrane-associated guanylate kinase (MAGUK) with inverted orientation protein 1 (MAGI1) is a novel member of the MAGUK family with a vital role in tumor progression related to invasion and metastasis. However, the function of MAGI1 in glioma is currently unknown. We therefore analyzed the expression of MAGI1 protein in human glioma samples, glioma cell lines and glioma stem cells (GSCs), and explored its effects on glioma cell proliferation and apoptosis.
Methods: MAGI1 expression in glioma tissues was examined by Western blotting and real-time polymerase chain reaction and its relationships with clinical pathological features were analyzed. The effects of MAGI1 knockdown on the proliferation of glioma cell lines and GSCs were detected by CCK8 and colony-formation assays, and apoptosis was assessed by flow cytometry. We also investigated the effects of MAGI1 silencing on protein expression levels of epithelial-mesenchymal transition biomarkers, as well as β-catenin, cyclin D1, PTEN and phospho-Akt by Western blotting.
Results: MAGI1 was significantly downregulated in glioma tissues and its expression was related to cancer progression. Silencing of MAGI1 in both glioma cell lines and GSCs enhanced proliferation and inhibited apoptosis. MAGI1 knockdown also significantly increased the expression levels of N-cadherin, vimentin, β-catenin, cyclin D1 and phospho-Akt and reduced the expression of E-cadherin and PTEN.
Conclusions: Our results indicated that MAGI1 might play a vital role in glioma progression and may represent a potential therapeutic target for the treatment of glioma.
Keywords: glioma, MAGI1, β-catenin, EMT, proliferation, knockdown
Introduction
Brain tumors are the leading cause of death in children with cancer and the third leading cause of cancer-related deaths in people aged 15–34 years.1 Gliomas are one of the most common primary brain tumors, accounting for about 80% of all cases, with overall incidence rates for all gliomas of 4.67–5.73 per 100,000 persons.2 According to the International Classification of Diseases-Oncology, version 3 (ICD-O-3) and World Health Organization (WHO) grade, gliomas are typically categorized as astrocytic, oligodendrocytic, or a mix of both cell types.3 Glioblastoma multiforme is a grade IV glioma that is notorious for its high degree of invasiveness and destructiveness, as well as its resistance to therapy. Accumulating evidences demonstrated that glioma stem cells (GSCs) contribute to not only for the initiation and progression of glioma but also for the recurrence and metastasis.4–6 Thus, the study of the potential mechanisms of GSCs is necessary for the pathogenesis of GSCs-targeting therapeutic technique.
Previous research has resulted in rapid advances in the discovery of some of the key molecular mechanisms, as well as the genetic and epigenetic factors, underpinning gliomas. However, despite progress in medical treatments such as surgery, radiotherapy, and chemotherapy, the prognosis of glioma remains poor, especially for glioblastoma which has a median survival of only 14.6 months.7 There is thus an urgent need to identify novel mechanisms that might indicate new possibilities for the treatment of glioma.
Membrane-associated guanylate kinase (MAGUK) with inverted orientation protein 1 (MAGI1) is a member of the MAGUK protein family, which comprises peripheral membrane proteins involved in the formation of specialized cell-cell junctions.8 MAGI proteins are localized at apical junctions and mainly function through binding to β-catenin and other apical proteins.9 MAGI1 can regulate cell-cell adhesion as a molecular scaffold protein through multiple protein interaction domains, including PSD-95/disks large/zonula occludens (PDZ), Src homology 3, WW, and guanylate kinase motif domains.10 MAGI1 interacts with a variety of PDZ-binding molecules including N-methyl-D aspartate receptors,11 brain-specific angiogenesis inhibitor 1,12 mNET1,10 and β-catenin.9 In addition, MAGI1 can bind the guanine nucleotide exchange factor for junction remodeling of the small GTPase RAP1 and is required for VE-cadherin-dependent Rap1 activation.13 MAGI1 also binds to the tumor suppressor phosphatase and tensin homolog (PTEN) and further stabilizes it at the membrane, resulting in increased PTEN activity.14 Overall, these results suggest that MAGI-1 may be act as a modulator of multiple signaling pathways.
MAGI1 also plays an important role in stabilizing cadherin mediated cell–cell conjunctions and inhibiting non-transformed epithelial cells. MAGI-1 reduced the proportions of human papillomavirus 16- and 18-positive proliferating cells and served as a negative regulator of cell proliferation.15 Previous reports have demonstrated that MAGI1 is associated with diverse cancers. MAGI1 expression was decreased in several types of human T-cell leukemia cell lines,16 and was down-regulated in solid tumors of the digestive system, including gastric cancer and hepatocellular carcinoma, and was correlated with disease prognosis.17,18 Furthermore, MAGI1 was capable of inducing an epithelial-like morphology and suppressing Wnt signaling, cell migration, and invasion in colorectal cancer cells.19 However, the relevance of MAGI1 in glioma remains unknown. In the present study, we examined the expression levels of MAGI1 in glioma tissues. In glioma cell lines and GSCs, we also determined the effect of MAGI1 knockdown on apoptosis, proliferation, and migration, and the role of the Wnt/β-catenin and PTEN/Akt signaling pathway in the actions of MAGI1.
Materials And Methods
Tissue Specimens
This study was approved by the Ethics Committee of Qingdao Municipal Hospital. All procedures involving human participants were performed in accordance with the ethical standards of the Committee and with the 1964 Helsinki declaration and its later amendments. Signed informed consent was obtained from all patients before surgery. Glioma specimens and corresponding normal brain tissues were obtained from 68 patients who underwent surgery. All tumor samples were classified as grade I, II, III, or IV based on the WHO classification guidelines. The clinical pathological characteristics are shown in Table 1. No patients had any history of chemotherapy or radiotherapy before surgery. The specimens were immediately frozen in liquid nitrogen after dissection and stored at −80°C until use.
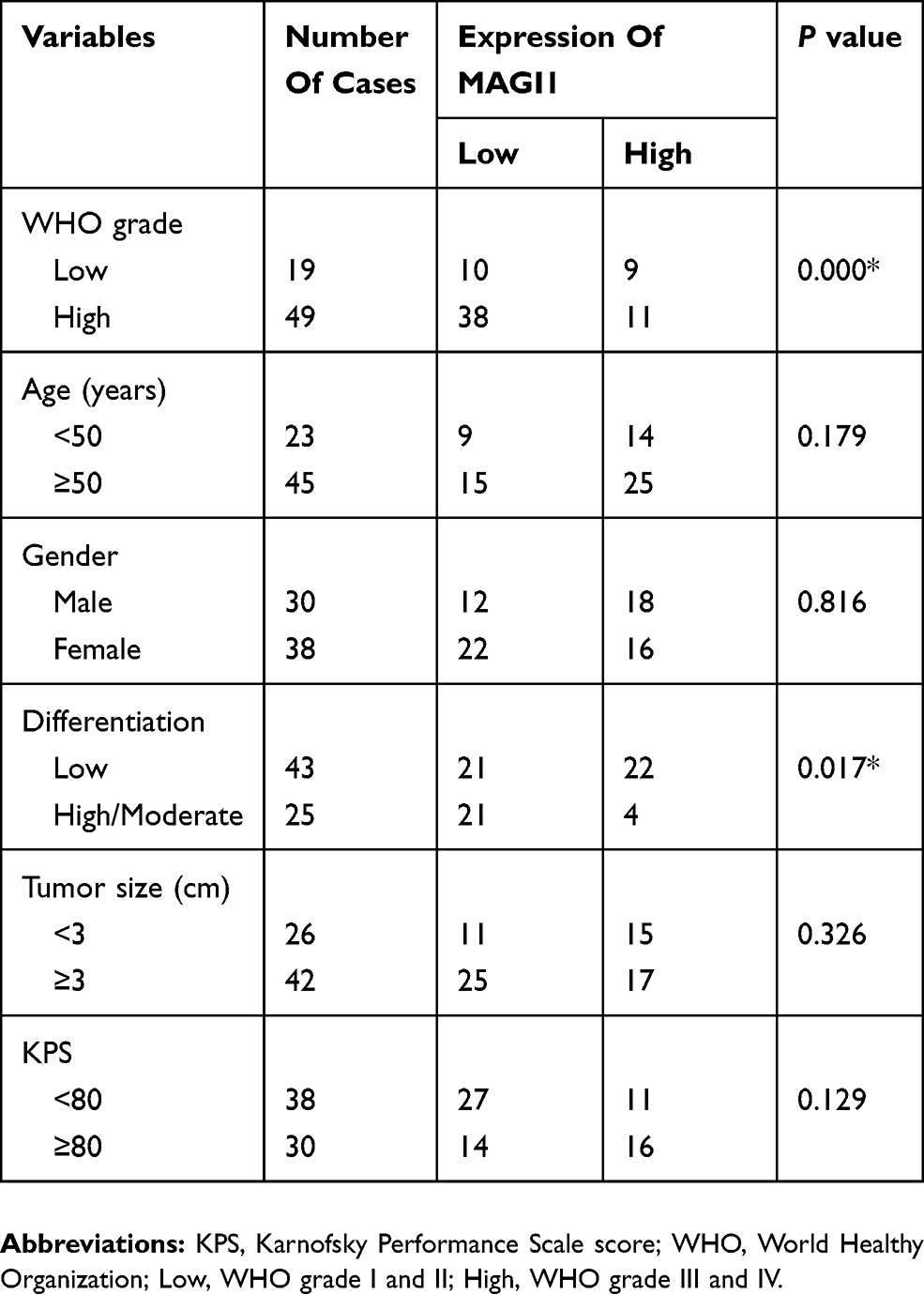 |
Table 1 Clinicopathologic Features Of 68 Glioma Patients |
Cell Culture And Transfection
Five human glioma cell lines (U87MG, U251, U118-MG, SHG44, LN229) were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). All procedures were performed on a sterile console. Cells were cultured in Dulbecco’s Modified Eagle’s Medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (Sigma-Aldrich, St. Louis, MO, USA) and 1% penicillin/streptomycin (Sigma-Aldrich) in a cell incubator with 5% CO2 at 37°C. The procedures used to establish GSCs have been previously described.20 Briefly, CD133-positive cells were sorted via magnetic cell sorting technology and were cultured in serum-free DMEM/F12 (SFM) containing 20 ng/mL human recombinant epidermal growth factor (EGF), 20 ng/mL human recombinant basic fibroblast growth factor (bFGF), and B27 (1:50) (all from Invitrogen, Carlsbad, CA, USA); half of the volume of medium was replaced every 3 days. This study was approved by the Ethics Committee of Qingdao Municipal Hospital and written informed consent was obtained from all patients providing tissue specimens. The following small hairpin RNAs (shRNAs) designed to silence endogenous MAGI1 in glioma cell lines were purchased from Shanghai Genechem Company (China): shMAGI#1: 5′-GAGGTTATCCATTGCCTTT-3′; shMAGI#2: 5′-ACCTATGAAGGAAACTATT-3′; and shRNA control: 5′-TTCTCCGAACGTGTCACGTTT-3′. The glioma cells were treated at a final concentration of 2 μg/mL.
Quantitative Real-Time Polymerase Chain Reaction (PCR) Analysis
Total RNA was extracted from glioma tissues and control samples using TRIzol (Invitrogen) and reverse transcribed using a PrimeScript RT Reagent Kit (RR047A; Takara). cDNA amplification was then performed using SYBR Premix Ex Taq (RR820; Takara Biotechnology, Dalian, China) in a LightCycler 480. PCR was carried out for 40 cycles according to the following procedure: 95°C for 30 s, annealing for 30 s, and 60°C for 34 s. Each sample was analyzed in duplicate. Relative mRNA expression levels of the target genes were calculated using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as an internal control. The primer sequences used for PCR were: MAGI1, forward: CCTTCAGTCTGGCTCTAAGC, reverse: GTGTGCTCCTCTTGTTCACC; GAPDH, forward: CAAAGACCTGTACGCCAACA, reverse: GAAGCATTTGCGGTGGAC.
Cell Proliferation Assay
Cell proliferation was examined using a Cell Counting Kit 8 (CCK8) assay (Dojindo, Kumamoto, Japan). Briefly, glioma cells were seeded at 2×103 cells/well in 96-well plates. After the cells were cultured for 4 days, 10 µL of CCK-8 reagent was added for 1 h. The optical density (OD) was measured at 490 nm using a spectrophotometer.
Western Blot Assay
Glioma tissues or cells were homogenized and lysed with radioimmunoprecipitation assay lysis buffer supplemented with 1 mmol/l phenylmethylsulfonyl fluoride and phosphatase inhibitor (Pierce; Thermo Fisher Scientific, Inc, IL), and then centrifuged at 12,000 rpm for 10 min at 4°C. Protein concentration was measured using a bicinchoninic acid kit (Takara Biotechnology, Dalian, China). Immunoblotting was performed according to the standard protocol and the proteins were transferred to polyvinylidene membranes (Millipore, Billerica, MA, USA), blocked with TBST with 5% skimmed milk for 2 h at room temperature, and then incubated at 4°C overnight with either anti-MAGI1, anti-E-cadherin, anti-N-cadherin, anti-Bax, anti-Bcl2, anti-caspase3 (all 1:1,000, Abcam, Cambridge, MA, UK), anti-vimentin, anti-PTEN, anti phospho-Akt, anti β-catenin, anti-cyclin D1 (all 1:1,000, Cell Signaling Technology, Beverly MA, USA), or anti-GAPDH antibodies (1:1,000, Santa Cruz Biotechnology, Santa Cruz, CA, USA). After three washes in TBST buffer, the membranes were incubated with horseradish peroxidase-conjugated IgG antibody for 2 h at room temperature. Immunoblot bands were quantified relative to GAPDH using Image Pro Plus software (Media Cybernetics, Inc.).
Colony-Formation Assay
Cell proliferation was evaluated by colony-formation assay. Glioma cells were seeded at 500–1,000 cells/well in 6-well plates and placed in a cell incubator with 5% CO2 at 37°C. The cells were stained with crystal violet and colony numbers were counted after 14 days of treatment.
Flow Cytometric Analysis
Glioma cells were seeded at a density of 2×105 cells/well in six-well plates, harvested, and resuspended in 100 μL binding buffer to achieve a concentration of 1×106 cells/mL. The cells were then stained with 5 μL annexin V-fluorescein isothiocyanate and 10 μL propidium iodide (BD Bioscience) according to the manufacturer’s guidelines and analyzed by flow cytometry (NovoCyte; ACEA). The data were analyzed using CellQuest software.
Animal And Subcutaneous Xenotransplantation Model
Animal care and experimental procedures were approved by Animal Ethics Committee of Qingdao Municipal Hospital. All experiments were carried out and received human care. Luciferase stably expressing U87 cells were transfected with Lenti-MAGI1 or Lenti-vector. Twenty 4-week-old male BALB/c nude mice (Linchang, Shanghai, China) were injected subcutaneously with 1×106 of the respective cells. Each mice was subcutaneously injected into the neck with 1×106 cells. Two weeks later, the tumor was visible. After five injections, the mice were sacrificed and the tumor tissues were integrally stripped out and counted for further study.
Statistical Analysis
All statistical analyses were performed using SPSS Software version 20.0 (SPSS, Inc., Chicago, IL, USA). Results were calculated as mean ± standard deviation. Differences between groups were compared by one-way ANOVA and Spearman correlation test. Data were analyzed and graphs were created using GraphPad Prism 5 software. A p value <0.05 was considered significant.
Results
MAGI1 Is Downregulated In Human Glioma Tissues
We evaluated the mRNA expression levels of MAGI1 in 68 human glioma samples and adjacent non-tumorous tissues by real time-PCR and Western blot. MAGI1 mRNA levels were significantly decreased in human glioma tissues compared with adjacent non-tumorous tissues (p<0.01) (Figure 1A). This result was confirmed by Western blot assay, which showed lower MAGI1 protein levels in glioma tissues compared with adjacent brain tissues (p<0.01) (Figure 1B). Besides, the results also showed that the MAGI1 protein expression were the lowest in the high grade of glioma tissues (grade IV) (Figure 1C)
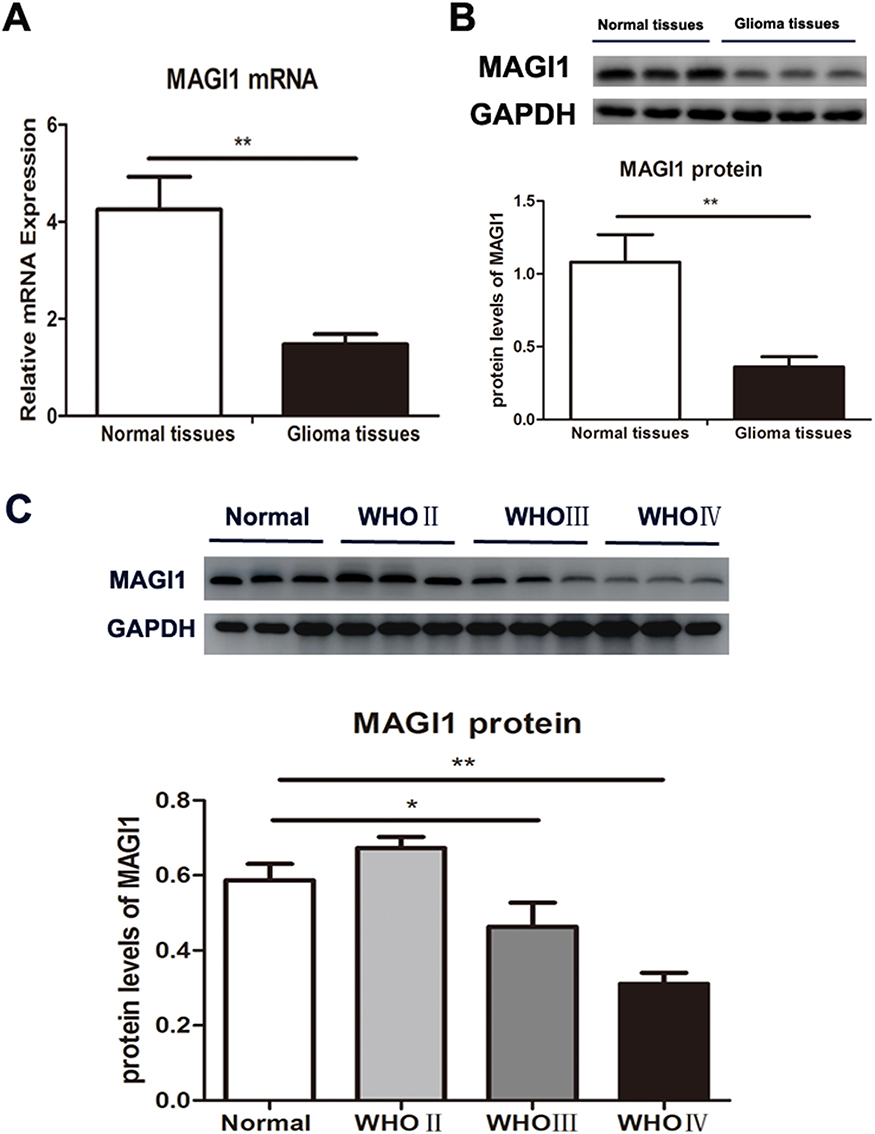 |
Figure 1 MAGI1 expression in glioma tissues and normal brain tissues. (A) MAGI1 mRNA expression was detected by real-time PCR and the ratio of MAGI1/GAPDH was determined by giving a mean net density in human glioma tissues compared with adjacent non-tumorous tissues. (B) MAGI1 protein expression was determined by Western blot and the ratio of MAGI1/GAPDH was determined by giving a mean net density in human glioma tissues compared with adjacent non-tumorous tissues. (C) MAGI1 protein levels were determined by Western blot analysis across the four subtypes and the ratio of MAGI1/GAPDH was determined by giving a mean net density. WHO: World Healthy Organization; Low: WHO grade I and II); High: WHO grade III and IV; Data were presented as the means ± standard errors. *p<0.05 and **p<0.01 was considered as statistically significant. Each lane represented a different sample. |
Correlation Between MAGI1 Expression And Clinical Pathology In Patients With Glioma
The clinical data for 68 patients with glioma were assessed to evaluate the relationships between MAGI1 expression levels and clinical pathological features (Table 1). We divided the patients into high- and low-MAGI1-expression groups. MAGI1 expression was significantly negatively correlated with WHO grade and differentiation degree (p<0.01), but there was no significant correlation between MAGI1 expression and any other clinical features, including sex, age, tumor size, and Karnofsky performance score.
MAGI1 Expression In Glioma Cell Lines
We examined the MAGI1 expression levels in glioma cell lines (U251, U87-MG, U118-MG, SHG44 and LN229) and showed that MAGI1 protein levels were highest in U87, LN229 and U251 cell lines (Figure 2A and B). These cell lines were therefore selected for subsequent assays.
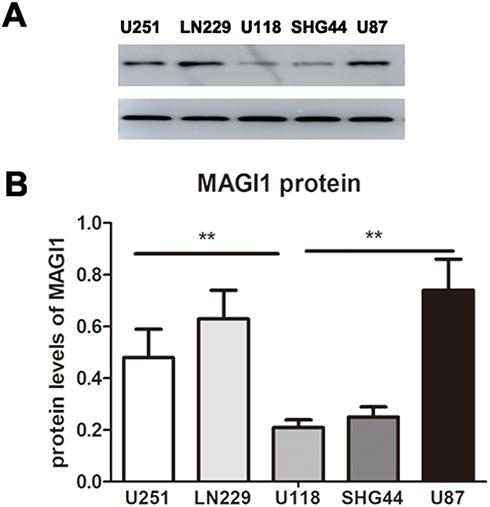 |
Figure 2 Expression of MAGI1 in glioma cell lines. (A) MAGI1 protein levels were determined by Western blot in five glioma cell lines (U251, U87-MG, U118-MG, SHG44 and LN229). (B) The ratio of MAGI1/GAPDH in glioma cell lines was determined by giving a mean net density. Data were presented as the means ± standard errors. **p<0.01 was considered significant. |
MAGI1 Knockdown Promotes Proliferation Of Glioma Cells
We assessed the role of MAGI1 in glioma cells by silencing its expression using shRNA in U87, U251 cells lines and GSCs. The transfection efficiency was evaluated by Western blot assay (Figure 3A). MAGI1 protein expression was dramatically reduced in both glioma cell lines and GSCs transfected with shMAGI1 compared with controls. We therefore used shMAGI1#1 and shMAGI1#2 for further studies.
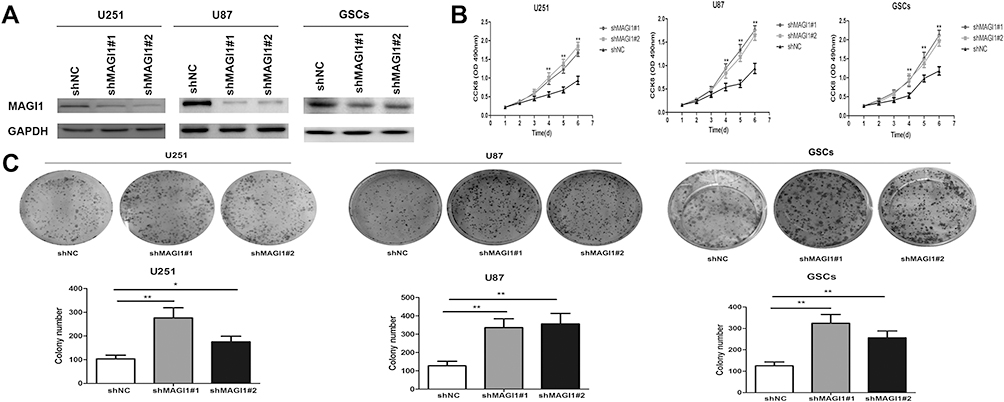 |
Figure 3 Knockdown of MAGI1 promotes cell proliferation in glioma cell lines and glioma stem cells (GSCs). (A) The expression of MAGI1 was examined after shMAGI1 transfection. (B) After stable transfection with shMAGI1 in U87, U251 cells and GSCs, the proliferation of glioma cells was detected by the CCK8 assay. Knockdown of MAGI1 notably enhanced the proliferation of U87, U251 cells and GSCs. (C) The colony forming assay indicated that knockdown of MAGI1 increased cell growth in U87, U251 cells and GSCs, Data are presented as the means ± standard errors. *p<0.05, **p<0.01 was considered significant. |
We detected the effects of MAGI1 knockdown on glioma cell growth by CCK8 and colony-forming assays. The proliferation of U87, U251 cells lines and GSCs in the control vector-transfected (shNC) and shMAGI1-transfected groups was examined by CCK8 assay over 6 days. The proliferation of shMAGI1 cells was dramatically enhanced after 96 h in both glioma cell lines and GSCs compared with the shNC group (p<0.01) (Figure 3B). Furthermore, MAGI1 knockdown also significantly increased the colony-formation capacities of U87, U251 cells lines and GSCs. The colony-forming counts were significantly increased compared with cells transfected with shNC (p<0.01) (Figure 3C). These findings indicated that silencing of MAGI1 could enhance the growth of glioma cells.
MAGI1 Knockdown Reduces Apoptosis Of Glioma Cells
We further analyzed the effect of MAGI1 on cell apoptosis by flow cytometry and Western blot assay. Apoptosis rates of U87, U251 cells lines and GSCs were significantly reduced in shMAGI1 cells compared with the shNC group (all p<0.01) (Figure 4A). In addition, Bax and cleaved-caspase 3 protein levels were significantly reduced while Bcl2 expression was enhanced in MAGI1-knockdown cells (all p<0.01) (Figure 4B), indicating that silencing MAGI1 significantly inhibited apoptosis in glioma cells.
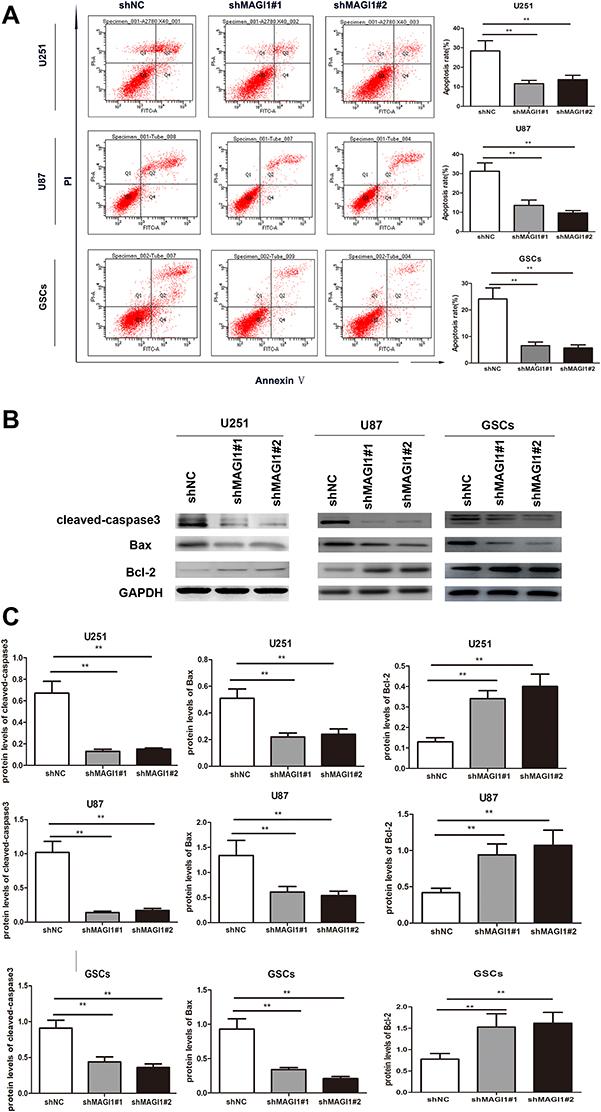 |
Figure 4 Knockdown of MAGI1 inhibits cell apoptosis in glioma cell lines and glioma stem cells (GSCs). (A) The apoptotic rates was determined by flow cytometry in U87, U251 cells and GSCs. Knockdown of MAGI1 inhibited cell apoptosis in U87, U251 cells and GSCs. (B) Western blot assay evaluated the expression of the cleaved-caspase 3, Bax and Bcl-2 proteins in U87, U251 cells and GSCs. (C) The ratios of the relative protein/GAPDH in glioma cells were determined by giving a mean net density. Data were presented as the means ± standard errors. **p<0.01 was considered significant. |
MAGI1 Knockdown Promotes Epithelial-Mesenchymal Transition (EMT) Phenotype In Glioma Cells
EMT is a vital process in tumor progression and metastasis, and is characterized by down-regulation of E-cadherin and upregulation of N-cadherin and vimentin. We therefore examined the effect of MAGI1 on the EMT process in U87, U251 cells lines and GSCs. E-cadherin expression was significantly suppressed (p<0.01) (Figure 5A and B), while vimentin and N-cadherin expression were significantly increased in shMAGI1-transfected U87 cells (all p<0.01). Moreover, silencing of MAGI1 also significantly downregulated E-cadherin and up-regulated vimentin and N-cadherin expression in U251 cells and GSCs compared with the shNC group (all p<0.01). These results suggested that silencing MAGI1 promoted the EMT phenotype in glioma cells.
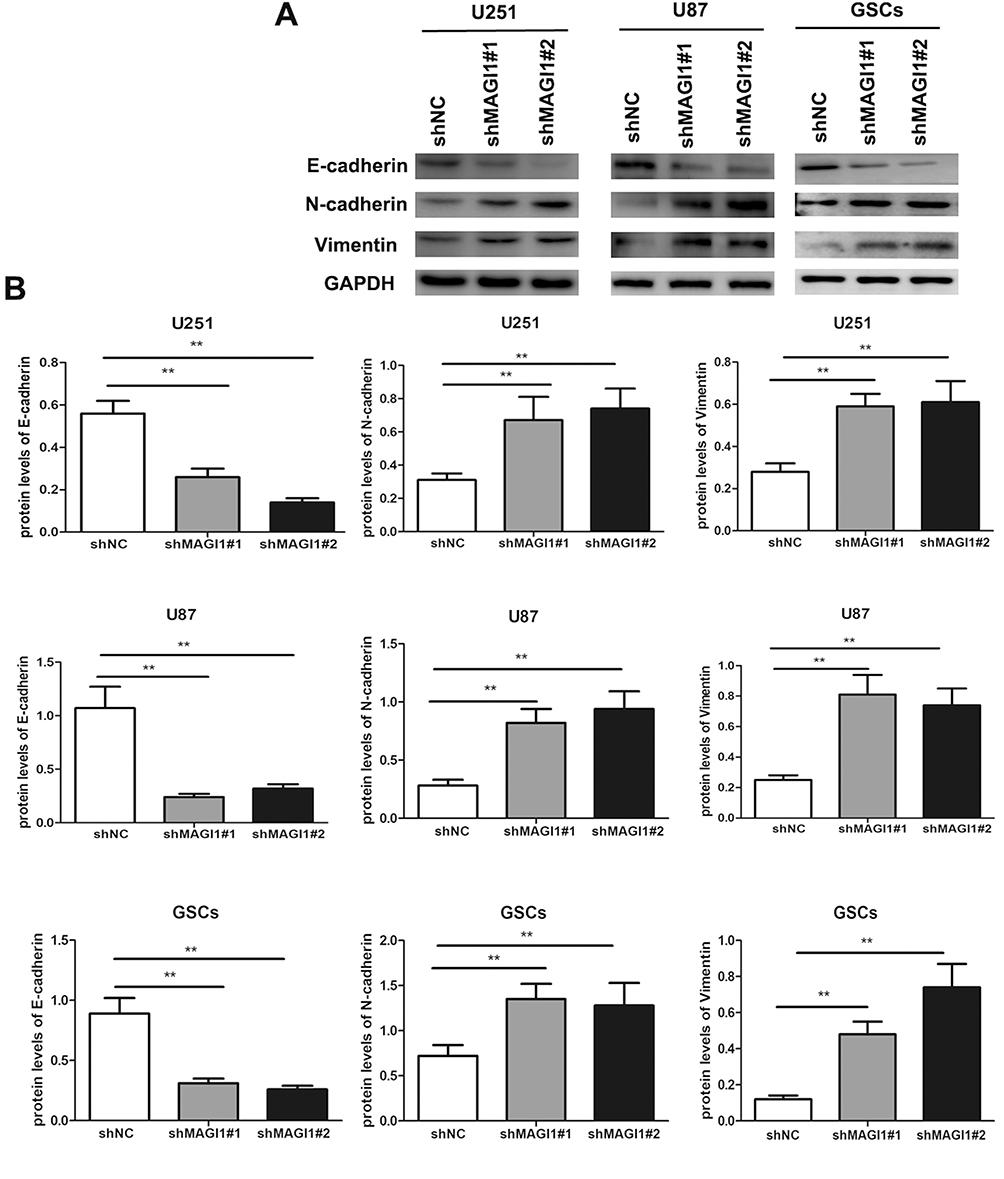 |
Figure 5 Knockdown of MAGI1 increases the EMT process in glioma cell lines and glioma stem cells (GSCs). (A) The protein expression levels of E-cadherin, N-cadherin and Vimentin were measured by Western blotting in U87, U251 and GSCs. (B) The ratios of EMT biomarkers were determined by giving a mean net density. Data were presented as the means ± standard errors. **p<0.01 was considered significant. |
MAGI1 Knockdown Functions Via Wnt/Β-Catenin And PTEN/Akt Signaling Pathways In Glioma Cells
The classic Wnt/β-catenin signaling pathway plays an important role in cell proliferation and can promote EMT in many cancers. We examined the effects of MAGI silencing on activation of the Wnt/β-catenin and PTEN/Akt signaling pathways in. We analyzed the expression levels of β-catenin and cyclin D1, which are correlated with the Wnt/β-catenin signaling pathway, in U87 and U251 cells. Silencing MAGI1 significantly upregulated β-catenin and cyclin D1 levels in both glioma cell lines (both p<0.01) (Figure 6A and B). Furthermore, expression levels of the tumor suppressor PTEN were significantly reduced and levels of phospho-Akt were significantly increased in both cell lines after shMAGI1 transfection compared with the shNC group (both p<0.01).
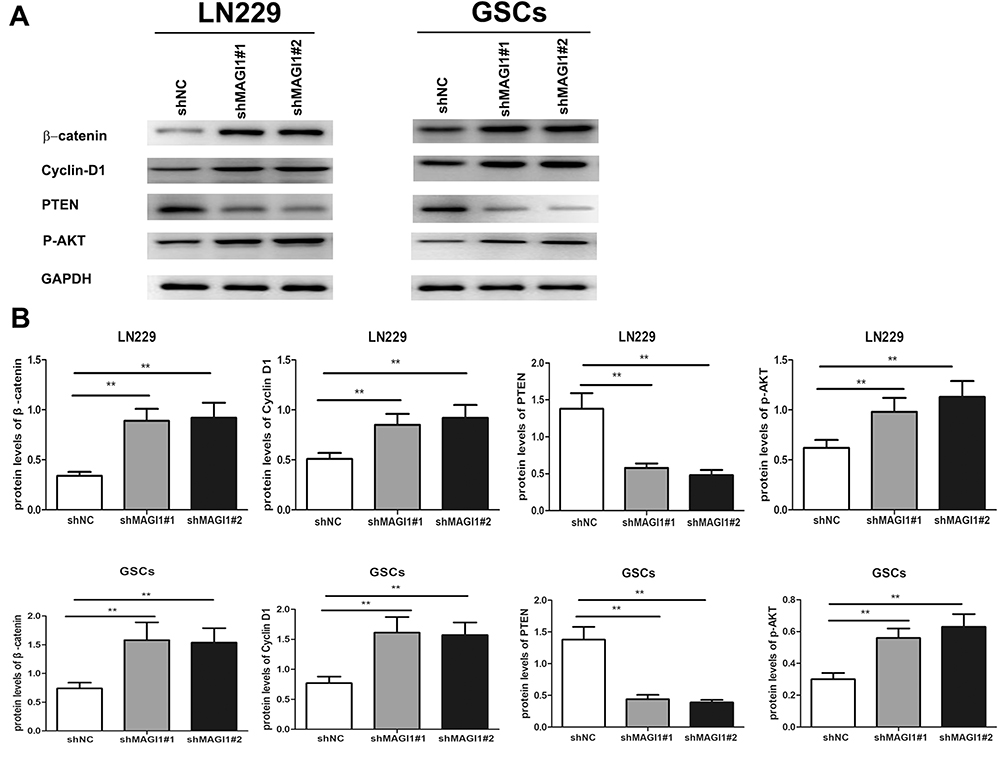 |
Figure 6 MAGI1 Knockdown functions through Wnt/β-catenin and PTEN/AKT signaling pathway in LN229 (PTEN wild-type) and glioma stem cells (GSCs). (A) The protein expression levels of β-catenin, cyclin D1, PTEN and p-AKT were measured by Western blotting in LN229 and GSCs. (B) The ratios of β-catenin, cyclin D1, PTEN and p-AKT were determined by giving a mean net density in LN229 and GSCs. Data were presented as the means ± standard errors. **p<0.01 was considered significant. |
Overexpression Of MAGI1 Inhibits The Tumorigenesis Of Glioma In Vivo
To further investigate whether overexpression of MAGI1 suppress tumor growth in vivo, a xenograft model using GSCs was performed, and subcutaneous injection with Lenti-MAGI1 or Lenti-vector in the nude mice was established. The results showed that the tumor volume was significantly decreased in nude mice with Lenti-MAGI1 when compared with vector-transfected cells (Figure 7A and B). Western blot assay confirmed that the protein levels of MAGI1 were significantly increased in the Lenti-MAGI1 group tumor cells than Lenti-vector group (Figure 7C).
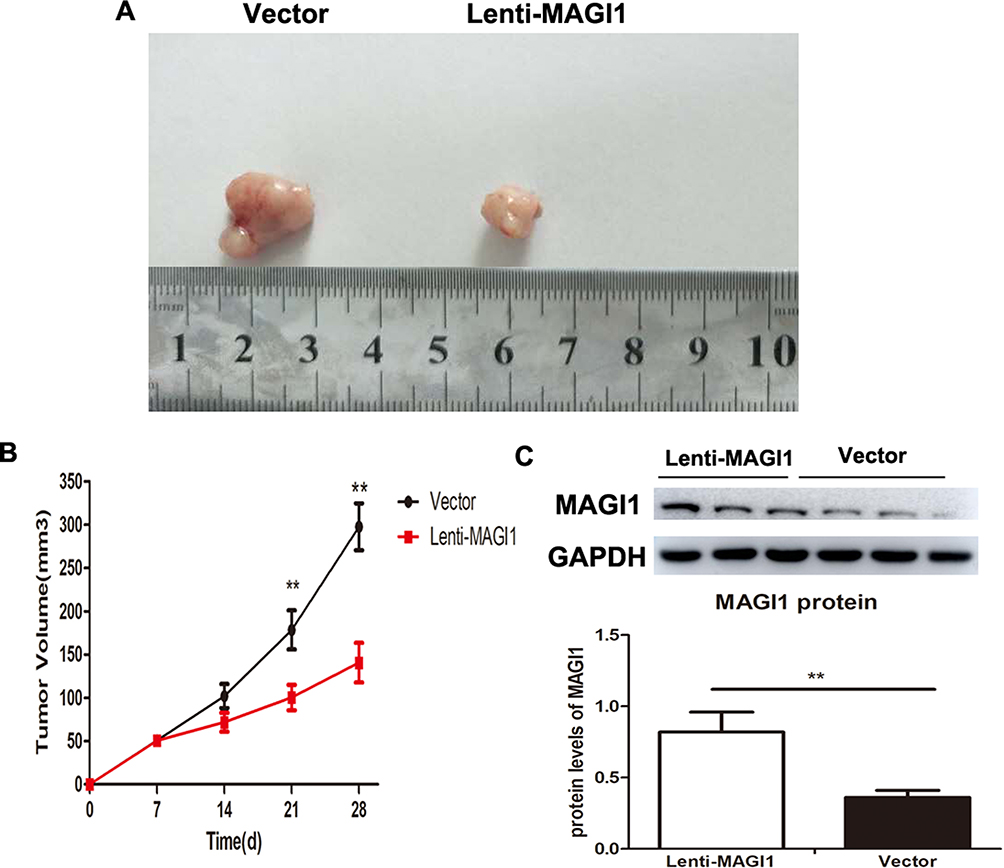 |
Figure 7 The effect of MAGI1 knockdown on tumorigenesis in vivo. (A) GSCs/Vector and GSCs/MAGI1 stabled cells were counted and injected subcutaneously in nude mice respectively. After 28 days, the mice were sacrificed and the tumor tissues were integrally stripped out. (B) The difference in tumor volume was counted in nude mice with Lenti-MAGI1 when compared with vector-transfected cells. (C) MAGI1 protein levels were determined by Western blot and the ratio of MAGI1/GAPDH was determined by giving a mean net density. Data were presented as the means ± standard errors. **p<0.01 was considered significant. |
Discussion
Glioma is the most common central nervous system tumor and is characterized by a high degree of malignancy and high recurrence rate. Despite the development of novel treatment, the prognosis of glioma remains poor,21,22 and novel therapeutic targets are needed to improve its diagnosis and treatment.
In this study, we showed that MAGI1 mRNA and protein expression levels were significantly decreased in human glioma samples. In addition, downregulation of MAGI1 was closely related to the degree of differentiation and WHO stage of the glioma. Silencing of MAGI1 expression with shRNA promoted cell proliferation and the EMT process and inhibited apoptosis in glioma cell lines and GSCs. Moreover, silencing of MAGI1 increased activation of the Wnt/β-catenin and PETN/AKT signaling pathways in glioma cells.
MAGI1 is a novel member of the MAGUK family that plays a vital role in tumor progression related to invasion and metastasis.23,24 MAGI1 has been shown to act as a tumor suppressor in multiple cancers, including hepatocarcinoma, cervical, gastric, and colorectal cancers,15,18,19,25–27 and MAGI1 expression was negatively related to cell progression in cancer cell lines.18,19,27 To the best of our knowledge, the current study provides the first evidence for the expression and contribution of MAGI1 in human glioma. Our results were in line with earlier studies showing that MAGI1 was significantly decreased in human glioma samples. We also evaluated the relationship between MAGI1 expression and clinical pathological features in patients with glioma and demonstrated that MAGI1 was negatively correlated with the degree of differentiation and WHO stage. Moreover, MAGI1 expression was reduced in well-differentiated gliomas and enhanced in invasive gliomas, suggesting that MAGI1 might participate in the development of glioma and contribute to its prognosis. We also verified the effects of MAGI1 in glioma by silencing its expression with shRNA, which indicated that MAGI1 significantly promoted the proliferation of U87 and U251 glioma cells. Overall, these findings suggested that MAGI1 may function as tumor suppressor in the pathogenesis of glioma.
In the current study, MAGI1 knockdown significantly reduced apoptosis rates in glioma cell lines and GSCs. Moreover, MAGI1 silencing also reduced cleaved caspase 3 and changed the Bax/Bcl2 ratio by downregulating Bax and upregulating Bcl2. Previous reports demonstrated that the Bax/Bcl2 ratio contributed to the secretion of cytochrome into the cytosol, leading to caspase activation.28 In addition, MAGI1 was also shown to regulate the apoptotic reaction in papillomavirus-positive tumor cells.15 These studies suggested that MAGI1 silencing may be involved in the apoptosis of glioma cells.
EMT is associated with the invasion and metastasis of cancer cells, and with enhanced tumor metastasis. Reduced expression of the epithelial biomarker E-cadherin and elevated expression of the mesenchymal biomarker N-cadherin are the main characteristics of EMT.29 Cancer cells with an EMT phenotype were associated with tumor metastasis and a poor prognosis in patients.30 Gliomas with EMT demonstrated increased invasion and metastasis, suggesting that EMT contributes to the progression of glioma.15–17,31–33 The current results showed that MAGI1 knockdown significantly reduced E-cadherin and increased vimentin and N-cadherin expression in glioma cell lines and GSCs, indicating that downregulation of MAGI1 promoted tumor progression by enhancing the EMT process in glioma cells.
We also examined the role of MAGI1 in the pathogenesis of tumor suppression in glioma. The Wnt/β-catenin signaling pathway has been reported to contribute to the regulation of various of processes, such as cell proliferation, invasion, metastasis, and differentiation,34,35 and to serve as a promoter in gastric cancer.36 Dysregulation of Wnt/β-catenin signaling has been identified as a vital pathogenesis of glioma development.37 β-catenin is the crucial downstream regulator of the classic Wnt signaling pathway and overexpression of β-catenin is highly related to the progression of glioma. Inhibition of the Wnt/β-catenin signaling pathway thus represent a potential therapeutic approach for glioma. Zaric et al showed that MAGI1 knockdown promoted Wnt signaling and reduced E-cadherin and β-catenin localization in colorectal cancer cell lines.19 In the current study, silencing of MAGI1 increased Wnt signaling pathway activation and upregulated the target gene cyclin D1 in glioma cells. These results indicated that MAGI1 knockdown promoted glioma progression by activating the Wnt/β-catenin signaling pathway. PTEN acts as a vital modulator of cell proliferation, migration, and invasion.38 PTEN is viewed as a classic tumor suppressor gene, and its downregulation is associated with the development of various tumors.39,40 PTEN decreased Akt phosphorylation and further suppressed tumor invasion and proliferation.41 Malignant gliomas were shown to have high levels of PTEN gene mutations, resulting in increased Akt phosphorylation and enhanced tumor progression.42 Interestingly, an earlier study demonstrated that PTEN functioned as a tumor suppressor by interacting with MAGI1, and that change of MAGI1 enhanced the mutual effect with PTEN.14 In the current study, silencing of MAGI1 in glioma cells enhanced Akt phosphorylation and reduced PTEN expression, indicating that MAGI1 might be involved in glioma progression via the PTEN/Akt signaling pathway. In addition, overexpression of MAGI1 significantly inhibited tumor growth in vivo as compared with vehicle treatment. It is indicated that overexpressing of MAGI1 could suppress tumor progression of glioma in vivo.
In conclusion, this study provided novel evidence demonstrating that MAGI1 contributed to glioma progression. Silencing MAGI1 promoted cell proliferation and the EMT process and inhibited apoptosis of glioma cell lines and glioma stem cells by inhibiting the Wnt/β-catenin and PTEN/AKT signaling pathways, suggesting that MAGI1 may be an effective therapeutic target for glioma.
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Buckner JC, Brown PD, O’Neill BP, Meyer FB, Wetmore CJ, Uhm JH. Central nervous system tumors. Mayo Clinic Proc. 2007;82(10):1271–1286. doi:10.4065/82.10.1271
2. Gousias K, Markou M, Voulgaris S, et al. Descriptive epidemiology of cerebral gliomas in northwest Greece and study of potential predisposing factors, 2005–2007. Neuroepidemiology. 2009;33(2):89–95. doi:10.1159/000222090
3. Louis DN, Ohgaki H, Wiestler OD, et al. The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 2007;114(2):97–109. doi:10.1007/s00401-007-0243-4
4. Bexell D, Svensson A, Bengzon J. Stem cell-based therapy for malignant glioma. Cancer Treat Rev. 2013;39(4):358–365. doi:10.1016/j.ctrv.2012.06.006
5. Reya T, Morrison SJ, Clarke MF, Weissman IL. Stem cells, cancer, and cancer stem cells. Nature. 2001;414(6859):105–111. doi:10.1038/35102167
6. Singh SK, Clarke ID, Terasaki M, et al. Identification of a cancer stem cell in human brain tumors. Cancer Res. 2003;63(18):5821–5828.
7. Stupp R, Hegi ME, Mason WP, et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009;10(5):459–466. doi:10.1016/S1470-2045(09)70025-7
8. Dobrosotskaya IY, James GL. MAGI-1 interacts with beta-catenin and is associated with cell-cell adhesion structures. Biochem Biophys Res Commun. 2000;270(3):903–909. doi:10.1006/bbrc.2000.2471
9. Kawajiri A, Itoh N, Fukata M, et al. Identification of a novel beta-catenin-interacting protein. Biochem Biophys Res Commun. 2000;273(2):712–717. doi:10.1006/bbrc.2000.3002
10. Dobrosotskaya I, Guy RK, James GL. MAGI-1, a membrane-associated guanylate kinase with a unique arrangement of protein-protein interaction domains. J Biol Chem. 1997;272(50):31589–31597. doi:10.1074/jbc.272.50.31589
11. Hirao K, Hata Y, Ide N, et al. A novel multiple PDZ domain-containing molecule interacting with N-methyl-D-aspartate receptors and neuronal cell adhesion proteins. J Biol Chem. 1998;273(33):21105–21110. doi:10.1074/jbc.273.33.21105
12. Ide N, Hata Y, Nishioka H, et al. Localization of membrane-associated guanylate kinase (MAGI)-1/BAI-associated protein (BAP) 1 at tight junctions of epithelial cells. Oncogene. 1999;18(54):7810–7815. doi:10.1038/sj.onc.1203153
13. Sakurai A, Fukuhara S, Yamagishi A, et al. MAGI-1 is required for Rap1 activation upon cell-cell contact and for enhancement of vascular endothelial cadherin-mediated cell adhesion. Mol Biol Cell. 2006;17(2):966–976. doi:10.1091/mbc.e05-07-0647
14. Kotelevets L, van Hengel J, Bruyneel E, Mareel M, van Roy F, Chastre E. Implication of the MAGI-1b/PTEN signalosome in stabilization of adherens junctions and suppression of invasiveness. FASEB J. 2005;19(1):115–117. doi:10.1096/fj.04-1942fje
15. Kranjec C, Massimi P, Banks L. Restoration of MAGI-1 expression in human papillomavirus-positive tumor cells induces cell growth arrest and apoptosis. J Virol. 2014;88(13):7155–7169. doi:10.1128/JVI.03247-13
16. Kozakai T, Takahashi M, Higuchi M, et al. MAGI-1 expression is decreased in several types of human T-cell leukemia cell lines, including adult T-cell leukemia. Int J Hematol. 2018;107(3):337–344. doi:10.1007/s12185-017-2359-1
17. Zhang G, Liu T, Wang Z. Downregulation of MAGI1 associates with poor prognosis of hepatocellular carcinoma. J Invest Surg. 2011;25(2):93–99. doi:10.3109/08941939.2011.606875
18. Jia S, Lu J, Qu T, et al. MAGI1 inhibits migration and invasion via blocking MAPK/ERK signaling pathway in gastric cancer. Chin J Cancer Res. 2017;29(1):25–35. doi:10.21147/j.issn.1000-9604.2017.01.04
19. Zaric J, Joseph JM, Tercier S, et al. Identification of MAGI1 as a tumor-suppressor protein induced by cyclooxygenase-2 inhibitors in colorectal cancer cells. Oncogene. 2012;31(1):48–59. doi:10.1038/onc.2011.218
20. Wu N, Xiao L, Zhao X, et al. miR-125b regulates the proliferation of glioblastoma stem cells by targeting E2F2. FEBS Lett. 2012;586(21):3831–3839. doi:10.1016/j.febslet.2012.08.023
21. Ostrom QT, Gittleman H, Fulop J, et al. CBTRUS statistical report: primary brain and central nervous system tumors diagnosed in the United States in 2008–2012. Neuro-Oncology. 2015;17 Suppl 4:iv1–iv62.
22. Wang X, Chen JX, Zhou Q, et al. Statistical report of central nervous system tumors histologically diagnosed in the Sichuan Province of China from 2008 to 2013: a West China glioma center report. Ann Surg Oncol. 2016;23(Suppl 5):946–953. doi:10.1245/s10434-016-5410-1
23. Nguyen MM, Nguyen ML, Caruana G, Bernstein A, Lambert PF, Griep AE. Requirement of PDZ-containing proteins for cell cycle regulation and differentiation in the mouse lens epithelium. Mol Cell Biol. 2003;23(24):8970–8981. doi:10.1128/MCB.23.24.8970-8981.2003
24. Wegmann F, Ebnet K, Du Pasquier L, Vestweber D, Butz S. Endothelial adhesion molecule ESAM binds directly to the multidomain adaptor MAGI-1 and recruits it to cell contacts. Exp Cell Res. 2004;300(1):121–133. doi:10.1016/j.yexcr.2004.07.010
25. Zhang G, Liu T, Wang Z. Downregulation of MAGI1 associates with poor prognosis of hepatocellular carcinoma. J Invest Surg. 2012;25(2):93–99. doi:10.3109/08941939.2011.606875
26. Feng X, Jia S, Martin TA, Jiang WG. Regulation and involvement in cancer and pathological conditions of MAGI1, a tight junction protein. Anticancer Res. 2014;34(7):3251–3256.
27. Zhang G, Wang Z. MAGI1 inhibits cancer cell migration and invasion of hepatocellular carcinoma via regulating PTEN. J Cent South Univ Med Sci. 2011;36(5):381–385. doi:10.3969/j.issn.1672-7347.2011.05.002
28. Kordezangeneh M, Irani S, Mirfakhraie R, Esfandyari-Manesh M, Atyabi F, Dinarvand R. Regulation of BAX/BCL2 gene expression in breast cancer cells by docetaxel-loaded human serum albumin nanoparticles. Med Oncol. 2015;32(7):208. doi:10.1007/s12032-015-0652-5
29. Sun L, Fang J. Epigenetic regulation of epithelial-mesenchymal transition. Cell Mol Life Sci. 2016;73(23):4493–4515. doi:10.1007/s00018-016-2303-1
30. Katsuno Y, Lamouille S, Derynck R. TGF-beta signaling and epithelial-mesenchymal transition in cancer progression. Curr Opin Oncol. 2013;25(1):76–84. doi:10.1097/CCO.0b013e32835b6371
31. Bhat KP, Salazar KL, Balasubramaniyan V, et al. The transcriptional coactivator TAZ regulates mesenchymal differentiation in malignant glioma. Genes Dev. 2011;25(24):2594–2609. doi:10.1101/gad.176800.111
32. Mahabir R, Tanino M, Elmansuri A, et al. Sustained elevation of Snail promotes glial-mesenchymal transition after irradiation in malignant glioma. Neuro-Oncology. 2014;16(5):671–685. doi:10.1093/neuonc/not239
33. Yan YR, Xie Q, Li F, et al. Epithelial-to-mesenchymal transition is involved in BCNU resistance in human glioma cells. Neuropathology. 2014;34(2):128–134. doi:10.1111/neup.12062
34. Anastas JN, Moon RT. WNT signalling pathways as therapeutic targets in cancer. Nat Rev Cancer. 2013;13(1):11–26. doi:10.1038/nrc3419
35. Jang GB, Kim JY, Cho SD, et al. Blockade of Wnt/beta-catenin signaling suppresses breast cancer metastasis by inhibiting CSC-like phenotype. Sci Rep. 2015;5:12465. doi:10.1038/srep12465
36. Radulescu S, Ridgway RA, Cordero J, et al. Acute WNT signalling activation perturbs differentiation within the adult stomach and rapidly leads to tumour formation. Oncogene. 2013;32(16):2048–2057. doi:10.1038/onc.2012.224
37. Gao L, Chen B, Li J, et al. Wnt/beta-catenin signaling pathway inhibits the proliferation and apoptosis of U87 glioma cells via different mechanisms. PLoS One. 2017;12(8):e0181346. doi:10.1371/journal.pone.0181346
38. Coustan-Smith E, Song G, Clark C, et al. New markers for minimal residual disease detection in acute lymphoblastic leukemia. Blood. 2011;117(23):6267–6276. doi:10.1182/blood-2010-12-324004
39. Shang Y, Guo XX, Li WW, et al. Cucurbitacin-B inhibits neuroblastoma cell proliferation through up-regulation of PTEN. Eur Rev Med Pharmacol Sci. 2014;18(21):3297–3303.
40. Li XT, Wang HZ, Wu ZW, et al. miR-494-3p regulates cellular proliferation, invasion, migration, and apoptosis by PTEN/AKT signaling in human glioblastoma cells. Cell Mol Neurobiol. 2015;35(5):679–687. doi:10.1007/s10571-015-0163-0
41. Yamada KM, Araki M. Tumor suppressor PTEN: modulator of cell signaling, growth, migration and apoptosis. J Cell Sci. 2001;114(Pt 13):2375–2382.
42. Vivanco I, Sawyers CL. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev Cancer. 2002;2(7):489–501. doi:10.1038/nrc839
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.