")
Back to Journals » Cancer Management and Research » Volume 13
Role of lncRNA NR2F1-AS1 and lncRNA H19 Genes in Hepatocellular Carcinoma and Their Effects on Biological Function of Huh-7
Authors Ji WC, Bao GJ, Yang FW, Sun L, Han R
Received 20 October 2020
Accepted for publication 26 December 2020
Published 3 February 2021 Volume 2021:13 Pages 941—951
DOI https://doi.org/10.2147/CMAR.S284650
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Ahmet Emre Eşkazan
Wen-Chao Ji,1 Guang-Jian Bao,1 Fang-Wu Yang,2 Lei Sun,1 Rui Han1
1Department of Hepatobiliary Surgery, Zaozhuang Municipal Hospital, Zaozhuang, 277100 Shandong Province, People’s Republic of China; 2General Surgery Department, Zaozhuang Mining Group Central Hospital, Zaozhuang, 277100 Shandong Province, People’s Republic of China
Correspondence: Rui Han
Department of Hepatobiliary Surgery, Zaozhuang Municipal Hospital, NO. 41 Longtou Road, Shizhong District, Zaozhuang, 277100, Shandong Province, People’s Republic of China
Tel + 86-18369221166
Email [email protected]
Objective: This research was designed to probe into the expression and related mechanism of lncRNA NR2F1-AS1 and H19 in hepatocellular carcinoma (HCC).
Methods: Forty-two HCC patients who came to our hospital from February 2018 to August 2019 were included into a research group (RG). Meanwhile, 46 healthy controls were regarded as a control group (CG). BEL-7402, Huh-7 human hepatoma cells and HL-7702 human normal liver cells were purchased, and the NR2F1-AS1 and H19 levels in serum and tissues of HCC patients were detected. PcDNA3.1-NR2F1-AS1, si-NR2F1-AS1, NC, pcDNA3.1-H19 and si-H19 were transfected into BEL-7402 and Huh-7 cells. The NR2F1-AS1 and H19 levels in samples were detected via qRT-PCR, and the expression of apoptosis-related proteins in cells was tested through WB. Cell proliferation, invasion, or apoptosis was detected by CCK8, Transwell or flow cytometry, respectively.
Results: The NR2F1-AS1 and H19 levels were high in human hepatoma cells, and AUCs of lncRNA NR2F1-AS1 and lncRNA H19 were both > 0.8. The lncRNA NR2F1-AS1 and lncRNA H19 were associated with HCC staging. After transfection of pcDNA3.1-NR2F1-AS1, si-NR2F1-AS1, NC, pcDNA3.1-H19, si-H19 BEL-7402 and Huh-7 cells, silencing NR2F1-AS1 and H19 expression can promote apoptosis and inhibit cell growth, while silencing their over-expression can inhibit the EMT process of Huh-7 cells.
Conclusion: lncRNA NR2F1-AS1 and lncRNA H19 genes are abnormally expressed in HCC. Furthermore, the two can suppress the EMT process of Huh-7 cells and promote apoptosis effectively.
Keywords: lncRNA H19, lncRNA NR2F1-AS1, hepatocellular carcinoma, HCC, trophoblast cells, invasion, apoptosis
Introduction
Hepatocellular carcinoma (HCC) is one of the complications of advanced cirrhosis. Its morbidity and mortality make it the sixth most common cancer around the world.1–4 At present, HCC is usually treated by liver transplantation clinically. However, due to the shortage of suitable liver organs, most advanced HCC patients are getting worse without a suitable liver. So, improving their survival rate is still a challenge for clinicians.5,6 With the deepening of research on long non-coding RNA (lncRNA) in tumor drug resistance, clinical medicine began to analyze lncRNA’s role as a molecular marker for detection and diagnosis of various human cancers and its abnormally expressed lncRNA in HCC.7–9
With the development of next-generation sequencing technology, genome-wide transcriptome analysis has become an effective method to detect new lncRNA in various disease states.10,11 A large number of studies have analyzed the disorders of lncRNAs related to liver cancer (LC) by genome-wide transcriptome; it is found that the lncRNA NR2F1-AS1 and lncRNA H19 levels are abnormally up-regulated.12,13 And it has been proved that lncRNA NR2F1-AS1 regulates miR-17/SIK1 axis to inhibit invasion and migration of cervical squamous cell carcinoma cells.14 Long non-coding RNA H19 is involved in sorafenib resistance of HCC by up-regulating miR‑675.10 The molecular mechanism of lncRNA NR2F1-AS1 and lncRNA H19 in HCC has not been completely characterized. Thus, this paper aims to provide a new theoretical basis for HCC diagnosis and treatment in molecular biology, influencing the role of lncRNA NR2F1-AS1 and lncRNA H19 genes in HCC and the biological function of Huh-7.
Data and Methods
General Data
Forty-two HCC patients diagnosed and treated in our hospital were included in the research group (RG). Simultaneously, 46 healthy controls were enrolled in the healthy control group (CG). Inclusion and exclusion criteria: (1) All subjects were diagnosed as HCC by our hospital, and the clinical diagnosis was made according to the International Health Organization (WHO) standard;15 patients did not receive neoadjuvant radiotherapy or chemotherapy before operation. (2) Patients who complicated with infectious diseases, other primary malignancy diseases, or complications affecting the level of target genes were excluded. All of them voluntarily took part in the experiment, signed an informed consent form, and cooperated with medical staff to complete relevant diagnosis and treatment work. This research was conducted in accordance with the “Declaration of Helsinki”. This research was approved by the Ethics Committee of our hospital and National Cancer Center.
Main Reagents and Detection Methods
Cell Culture
Human LC cells BEL-7402, Huh-7 and normal liver cells HL-7702 [L-02] were purchased from Bena Culture Collection (provided by frozen storage tube/T25 culture bottle resources), No. BNCC338237, BNCC337690, BNCC100012, respectively. Growth conditions: All cells were cultured at 37°C and 5%CO2 in a CM2-1 culture medium (90%RPMI-1640+10%FBS).
Construction and Transfection of Expression Vectors
All expression vectors were designed by Thermo Fisher Scientific, including NR2F1-AS1 low expression vector (si-NR2F1-AS1) and H19 low expression vector (si-H19). As to NR2F1-AS1 and H19 over-expression, the coding sequence (CDS) was amplified and inserted into pcDNA3.1 vector (pcDNA3.1-NR2F1-AS1, pcDNA3.1-H19) (constructed by FitGene Co., Ltd.), and miRNA targeted inhibitory sequence (miRNA-inhibitor), miRNA mimic sequence (miRNA-mimics) and miR negative control group (miR-NC, Guangzhou, Guangdong, China). Cells were transfected by Lipofectamine™ 2000 kit, and the operation steps were strictly in line with the instructions. siRNA sequence: siH19-1: 5ʹ-ACGAGGCACUGCGGCCCAG-3ʹ.
qRT-PCR
Under fasting condition, 5 mL blood was drawn from elbow vein of each participant. Serum samples were prepared by conventional methods. Total RNA in serum, tissue samples and cells was separated by Trizol reagent (Invitrogen, carlsbad, California, USA), and its concentration and quality were measured by Nanodrop. Next, it was reverse transcribed into cDNA using iScriptTM cDNA synthesis kit (Bio-Rad, Hercules, CA). PCR was performed on qRT-PCR detection system (Applied Biosystems, Foster City, CA). All Ct values were measured by 2−ΔΔCt method. The primer sequences are shown in Table 1.
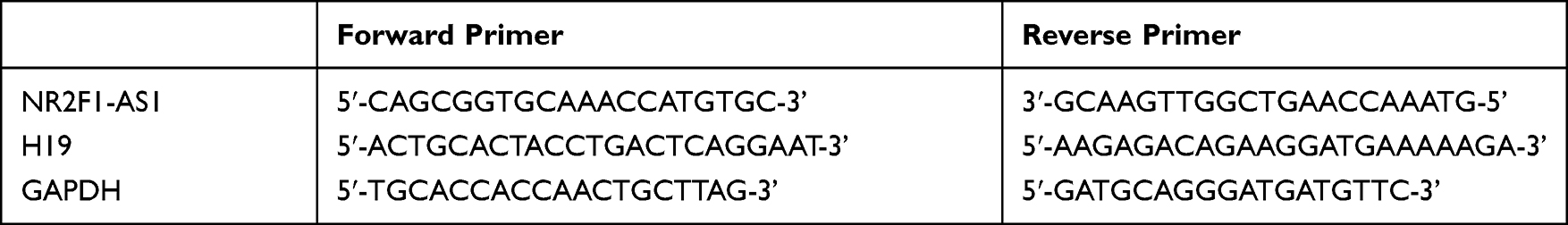 |
Table 1 Primer Sequence |
CCK8 Cell Proliferation Experiment
After logarithmic growth phase cells were digested with trypsin and re-suspended, the cell concentration was adjusted to 2×104/mL. They were inoculated into the plate and transfected with siRNA, miRNA or plasmid based on appropriate schemes. At each time point, CCK-8 reagent was added to each hole. Subsequently, the absorbance at 450 nm was measured by microplate reader to reflect the cytoactive.
Transwell Invasion Experiment
The cells were re-suspended in DMEM medium comprising 1% FBS at 4×105 cells/mL. Then, 200 μL cell suspension was added to the upper chamber, and 700 μL DMEM medium containing 10% FBS was added to the lower chamber. After the cells were incubated for 24 h to 48 h, those in the upper chamber were taken out and stained 20 min with 0.5% crystal violet. Five visual fields were observed randomly under an optical microscope, and the cells passing through the basement membrane of the chamber were counted.
Flow Cytometry Experiment
After the Huh-7 cells were transfected for 48 h, the apoptosis was measured. Collected by EDTA- trypsin, the cells were washed with PBS. Next, a 1×106 cells/mL suspension was prepared after cells were re-suspended by 100 μL AnnexinV Binding Buffer. Afterwards, 5 μL Annexin-V/FITC solution was added and incubated 15 min at 4°C, and 5 μL propidium iodide PI staining solution was added and incubated 5 min at 4°C. To take the average value, the test was repeated three times, and we analyzed it through flow cytometry (BD Biosciences, USA).
Western Blot (WB) Analysis
Protein samples were extracted by radioimmunoassay supplemented with protease inhibitors. Then, the protein was separated on 10% SDS-PAGE gel and transferred to PVDF membrane. The membrane was incubated with primary antibody and cultivated at 4 C all night. Then, the members were incubated with the secondary antibody. Finally, the protein was visualized by ECL detection system and quantified by ImageJ Software. β-actin was used as loading control. The primary antibody and secondary antibody goat anti-rabbit (HRP cross-linking) were bought from Abcam, Shanghai.
Statistical Methods
SPSS 20.0 (SPSS, Inc, Chicago, IL, USA) was used for statistical analysis. The normal distribution data were expressed by means±standard deviation (meas±SD), analyzed by Bonferroni post hoc test, and then assessed via two-tailed t-test and one-way or two-way analysis of variance. p<0.05 values were considered to be remarkably different.
Results
Expression of Target lncRNA Gene in HCC
Quantitative detection of gene expression by qRT-PCR:
(1) Compared with healthy controls, the lncRNA NR2F1-AS1 and lncRNA H19 levels in serum of HCC patients increased dramatically (all p<0.001). Receiver operating characteristic (ROC) curve found that AUCs of the two lncRNA were greater than 0.8.
(2) Compared with the paracancerous tissues, the lncRNA NR2F1-AS1 and lncRNA H19 levels in HCC tissues increased dramatically (all P<0.001).
(3) Compared with normal human normal liver cells HL-7702 [L-02], the lncRNA NR2F1-AS1 and lncRNA H19 levels in Huh-7 cells and BEL-7402 cells increased markedly (all P<0.001).
(4) In view of the median expression of the two target genes lncRNA, patients were divided into high expression group (HEG) and low expression group (LEG). It showed that the lncRNA NR2F1-AS1 and lncRNA H19 levels in the serum of stage III/IV HCC patients were higher and higher (all P<0.001) (Figures 1 and 2).
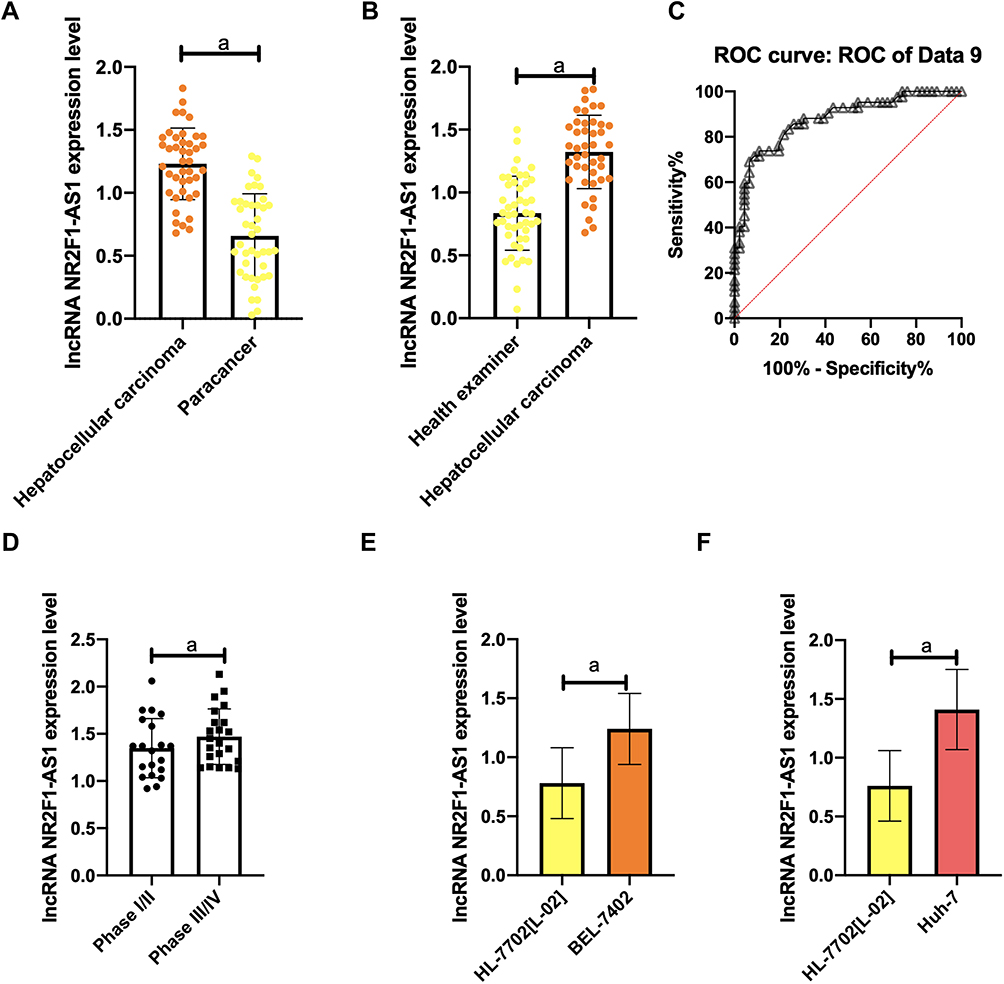 |
Figure 1 Gene expression of lncRNA NR2F1-AS1. (A) Compared with paracancerous tissues, lncRNA NR2F1-AS1 is markedly up-regulated in HCC tissues. (B) lncRNA NR2F1-AS1 expression in serum of HCC patients; (C) ROC curve analysis shows that AUC of lncRNA NR2F1-AS1 is > 0.8; (D) lncRNA NR2F1-AS1 expression in stage III/IV patients; (E) lncRNA NR2F1-AS1 expression in Huh-7; (F) lncRNA NR2F1-AS1 expression in BEL-7402. Note: aP<0.001. |
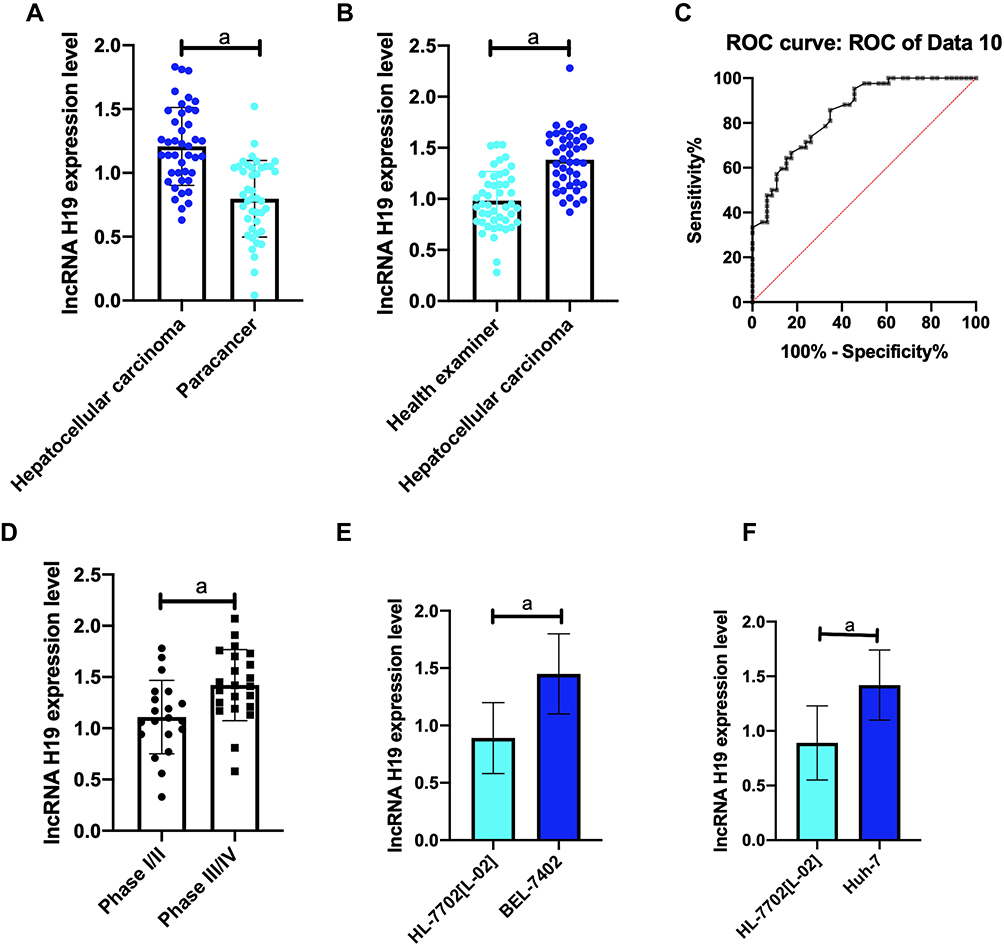 |
Figure 2 Gene expression of lncRNA H19. (A) The lncRNA H19 expression in HCC is markedly down-regulated compared with that in paracancerous tissues. (B) lncRNA H19 expression in serum of HCC patients; (C) ROC curve analysis shows that AUC of lncRNA H19 is > 0.8; (D) lncRNA H19 expression in stage III/IV patients; (E) lncRNA H19 expression in Huh-7; (F) lncRNA H19 expression in BEL-7402. Note: aP<0.001. |
Transfection of Cell Lines
(1) After Huh-7 cell transfection, the NR2F1-AS1 expression in the si-NR2F1-AS1 interference group was markedly lower than that in NC group, and the expression in the pcDNA3.1-NR2F1-AS1 group was markedly higher than that in NC group (all P<0.001); the H19 expression in the si-H19 interference group was markedly lower than that in NC group, and the expression in the pcDNA3.1-H19 group was markedly higher than that in NC group (all p<0.001) (Figures 3 and 4).
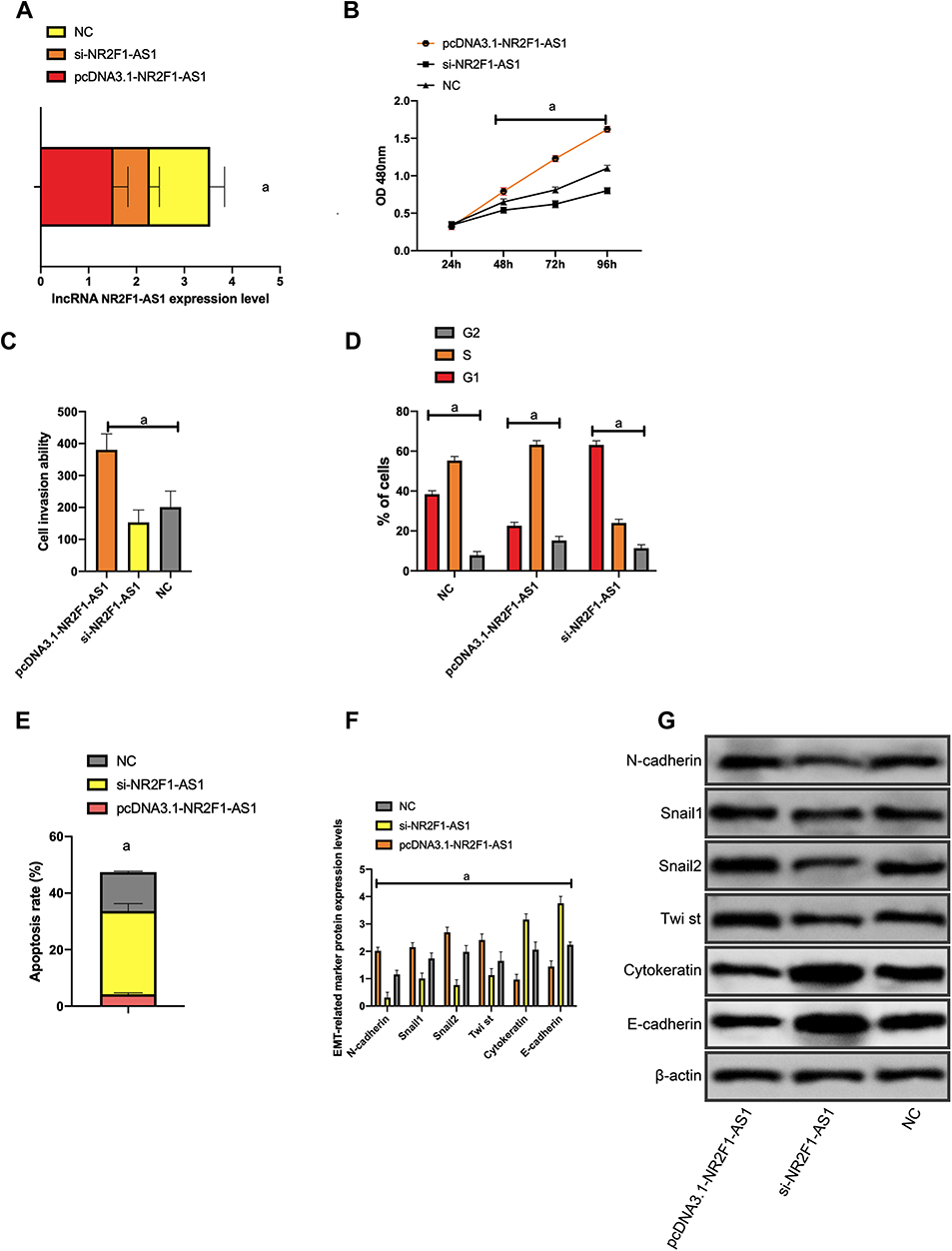 |
Figure 3 Effect of NR2F1-AS1 on biological function of Huh-7 cells NR2F1-AS1 expression after transfection of Huh-7 cells; (B) proliferation of Huh-7 cells after transfection; (C) invasion of Huh-7 cells after transfection; (D) cell cycle of Huh-7 after transfection; (E) apoptosis of Huh-7 after transfection; (F) expression of EMT-related markers after transfection; (G) WB figure. Note: aP<0.001. |
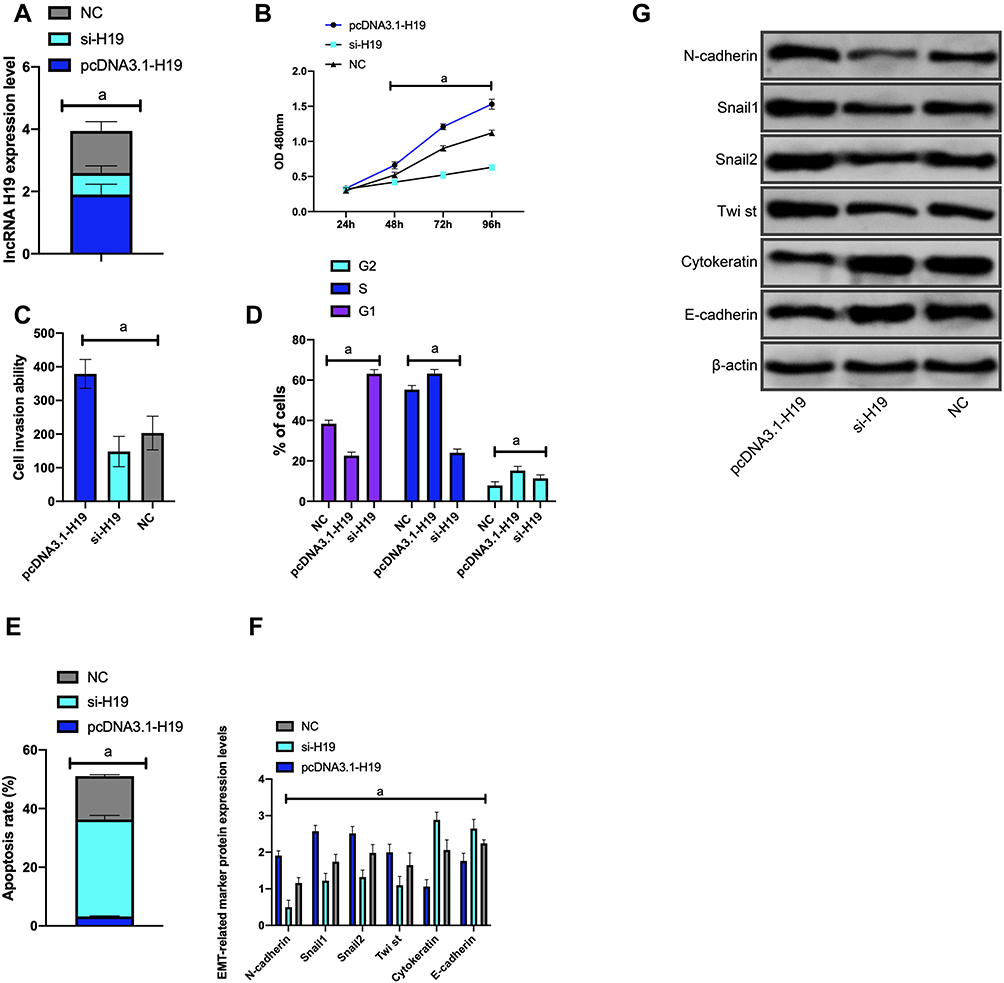 |
Figure 4 Effect of H19 on biological function of Huh-7 cells (A) H19 expression after transfection of Huh-7 cells; (B) proliferation of Huh-7 cells after transfection; (C) invasion of Huh-7 cells after transfection; (D) cell cycle of Huh-7 after transfection; (E) Huh-7 Apoptosis after transfection; (F) expression of EMT-related markers after transfection; (G) WB figure. Note: aP<0.001. |
(2) Compared with NC group, the proliferation and invasion of the si-NR2F1-AS1 interference group and the si-H19 interference group were obviously weakened, while those of the pcDNA3.1-NR2F1-AS1 group and the pcDNA3.1-H19 group were obviously enhanced (P<0.001); the apoptosis rate of the si-NR2F1-AS1 interference group and the si-H19 interference group decreased obviously, and the rate of the pcDNA3.1-NR2F1-AS1 group increased dramatically (P<0.001) (Figures 3 and 4).
(3) Compared with NC group, N-cadherin, SNAIL, Slug, CK18 and Vimentin in the si-NR2F1-AS1 interference group and the si-H19 interference group were dramatically down-regulated (P<0.001); E-cadherin in the si-NR2F1-AS1 interference group and the si-H19 interference group increased dramatically (P<0.001) (Figures 3 and 4).
Functional Target mRNA of lncRNA is Predicted by TargetScan, Starbase Database
(1) It was predicted that miR-363 had complementary binding sequence with NR2F1-AS1 in its 3ʹ-UTR, and the luciferase reporter plasmid of WT-H19 and mut-H19 binding site are shown in Figure 6A.
The luciferase reporter plasmid containing WT-H19 and miR-140-5p-mimics in Huh-7 cells reduced reporter gene activity (P<0.001). The miR-363 expression was detected by WB. miR-363 was down-regulated in Huh-7 cells after pcDNA3.1-NR2F1-AS1 transfection, and up-regulated after si-NR2F1-AS1 transfection (P<0.001). PcDNA3.1-NR2F1-AS1 and miR-363-mimics were co-transfected into Huh-7 cells. Functional analysis identified that after miR‐363-mimics transfection, the proliferation and invasion of Huh-7 cells decreased markedly, while the apoptosis increased (all P<0.01). pcDNA3.1-NR2F1-AS1/miR-363-mimics transfection could partially reverse the pro-apoptosis effect of pcDNA3.1-NR2F1-AS1 on Huh-7 cells. These results indicated that miR-363 was the functional target of NR2F1-AS1 in Huh-7 cells (Figure 5).
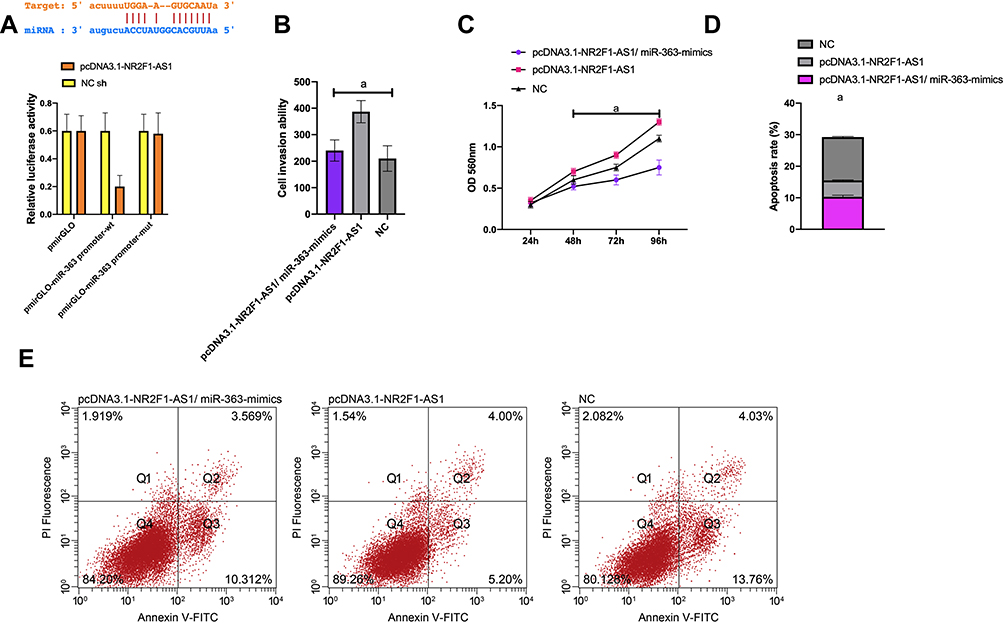 |
Figure 5 Dual-luciferase activity assay. (A) There is a binding site between NR2F1-AS1 and miR‐363 through relative luciferase activity-dual luciferase report assay; (B) invasion of Huh-7 cells after transfection; (C) cell proliferation; (D) apoptosis; (E) Flow cytometry. Note: aP<0.001. |
(2) miR-140-5p had complementary binding sequences with H19 in its 3ʹ-UTR, and the luciferase reporter plasmid and mut-H19 binding site of WT-NR2F1-AS1 are shown in Figure 6A.
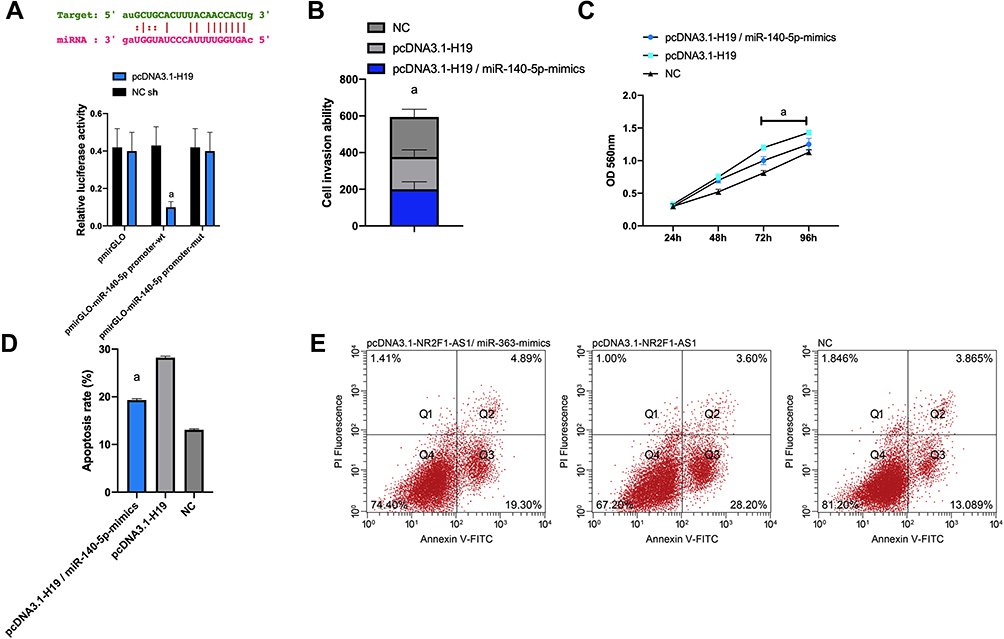 |
Figure 6 Dual-luciferase activity assay. (A) There is a binding site between H19 and miR-140-5p through the relative luciferase activity-dual luciferase report assay; (B) invasion of Huh-7 cells after transfection; (C) cell proliferation; (D) apoptosis; (E) Flow cytometry. Note: aP<0.001. |
The luciferase reporter plasmid comprising WT-NR2F1-AS1 and miR-363- mimics in Huh-7 cells reduced reporter gene activity (P<0.001). The miR-140-5p expression detected by WB showed that miR-140-5p was down-regulated in Huh-7 cells after pcDNA3.1-H19 transfection, and up-regulated after si-H19 transfection (P<0.001). pcDNA3.1-H19 and miR-140-5p-mimics were co-transfected into Huh-7 cells. Functional analysis manifested that after miR-140-5p-mimics transfection, the proliferation and invasion of Huh-7 cells decreased markedly, while the apoptosis increased (all P<0.01). Transfecting pcDNA3.1-H19/miR-140-5p-mimics could partially reverse the pro-apoptosis effect of pcDNA3.1-H19 on Huh-7 cells. These results indicated that miR-140-5p was the functional target of H19 in Huh-7 cells (Figure 6).
Effects of lncRNA NR2F1-AS1 and lncRNA H19 Gene on DNA Synthesis Phase (S Phase) of Huh-7 Cells
We monitored the cell cycle changes of Huh-7 cells transfected with pcDNA3.1-NR2F1-AS1/miR‐363-mimics and pcDNA3.1-H19/miR-140-5p-mimics.
After pcDNA3.1-NR2F1-AS1 and pcDNA3.1-H19 transfection, S-phase cells, EMT-related epithelial phenotypic protein level, and apoptosis-related protein expression decreased, while apoptosis-inhibiting protein expression increased.
After pcDNA3.1-NR2F1-AS1/miR-140-5p-mimics and pcDNA3.1-H19/miR-140-5p-mimics transfection, the number of S-phase cells and the level of EMT-related epithelial phenotypic proteins increased, the expression of pro-apoptotic protein was obviously up-regulated and that of anti-apoptotic protein was obviously down-regulated (P<0.05) (Figure 7).
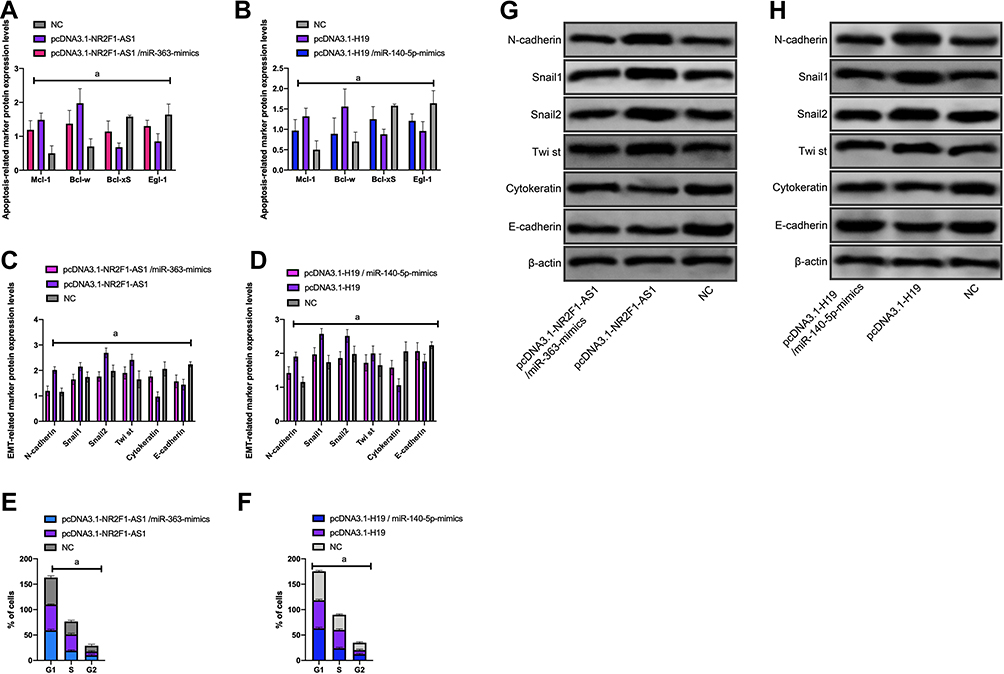 |
Figure 7 Effect of cell cycle and apoptosis-related proteins in HUH-7 cells. (A) Expression of apoptotic protein after pcDNA3.1-NR2F1-AS1/miR‐363-mimics transfection; (B) expression of apoptotic protein; (C) expression of EMT protein; (D) expression of EMT protein; (E) cell cycle; (F) cell cycle; (G) WB diagram; (H) WB diagram; aP<0.001. |
Discussion
HCC, as an aggressive malignancy with high recurrence rate, is always unsatisfactory in clinical treatment.16 It is widely verified that lncRNA takes part in tumorigenesis and regulation, so improving the research on “lncRNA regulates chemotherapy resistance of tumor cells and its influence on biological function of related tumor cells” is a vital purpose of the continuous development of bioinformatics prediction tools.17,18
lncRNA H19 is located on human chromosome 11p15, and H19 is up-regulated and has a strong carcinogenic function in cancer progression.19–21 Comparing the differences of lncRNA NR2F1-AS1 and lncRNA H19 between HCC and healthy people, we found that both of them were highly expressed in HCC patients’ serum and cancer tissues. The lncRNA NR2F1-AS1 up-regulated the carcinogenesis of FOXA1 in osteosarcoma by sponging miR-483-3p.22 In the cell transfection experiment, we silenced and over-expressed the NR2F1-AS1 and H19 levels in Huh-7 cells, and observed cell biological function. It was found that the invasion of HCC after inhibiting NR2F1-AS1 or H19 was obviously inhibited. This suggested that NR2F1-AS1 or H19 could influence the proliferation and apoptosis of Huh-7 cells, which could predict HCC progression. lncRNA mainly plays various roles in cancer by colliding with microRNA (miRNA). It was found that lncRNA AGAP2-AS1 might become a key inhibitor in HCC by competing with miR-574. This potential lncRNA-miRNA-regulatory target provided a novel therapeutic strategy for HCC.23 The lncRNA NR2F1-AS1 was highly expressed in esophageal squamous cell carcinoma tissues and cells. In addition, it also promoted the proliferation and metastasis of esophageal squamous cell carcinoma cells by regulating EMT.24
At last, we predicted that miR-363 and NR2F1-AS1 had complementary binding sequences, and miR-140-5p and H19 had complementary binding sequences by Targetscan6.2, and then analyzed the EMT process and cell cycle of H19 cells regulated by miRNA targeted by lncRNA NR2F1-AS1 and lncRNA H19. Many studies suggest that lncRNA can regulate G1/S-phase transition of cell cycle.25 While we found that transfecting pcDNA3.1-NR2F1-AS1/miR‐363-mimics and pcDNA3.1-H19/miR-140-5p-mimics could partially reverse EMT induction by pcDNA3.1-NR2F1-AS1 or H19, and restore the malignant proliferation of Huh-7 cells. The long non-coding RNA NR2F1-AS1 promoted the proliferation and migration of thyroid cancer cells by regulating miRNA-338-3p/CCND1 axis, but inhibited their apoptosis.26 lncRNA H19 mediated cisplatin resistance and migration of ovarian cancer cells during EMT.27 miR-675-3p derived from LncRNA H19 boosted epithelial-mesenchymal transition and stem cell growth of human pancreatic cancer cells.28 Combined with the results, we believe that knocking down lncRNA NR2F1-AS1 and lncRNA H19 can reduce the proliferation and metastasis of Huh-7 cells. What is more, over-expressing miR-363/miR-140-5p can suppress HCC progression promoted by lncRNA NR2F1-AS1/lncRNA H19.
In this research, we explored the ability of lncRNA NR2F1-AS1 and lncRNA H19 genes to interfere with the protein encoded by target gene mRNA. And we confirmed their function in HCC and influence on the biological function of Huh-7. Functional tests were conducted to explore the mechanism of regulating the behavior of Huh-7 cells by targeting miRNA with H19 and NR2F1-AS1. However, we still have some limitations. For one thing, we have not further explored the lncRNA-miRNA-mRNA network, for another, no related promotion studies have been made on other related signal pathways. Hence, we hope to screen and analyze more lncRNA and mRNA which are ceRNA in HCC in the future research, so as to provide more basis for the experiment.
Conclusion
To summarize, lncRNA NR2F1-AS1 and lncRNA H19 genes are abnormally high in HCC. Furthermore, the two can effectively inhibit the EMT process of Huh-7 cells and promote apoptosis.
Funding
The authors received no funding for this work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Pauline P, Boris G, Sophie G, et al. Intra-arterial idarubicin_lipiodol without embolization can provide prolonged complete response in hepatocellular carcinoma: a case report. J Oncol Pharm Pract. 2020;26:507–510. doi:10.1177/1078155219861422
2. Wanshui Y, Jing S, Ma Y, et al. A prospective study of dairy product intake and the risk of hepatocellular carcinoma in U.S. men and women. Int J Cancer. 2020;146:1241–1249. doi:10.1002/ijc.32423
3. Shao-Jiao M, Peng-Fei Y, Liu Y-P, et al. BCLAF1 promotes cell proliferation, invasion and drug-resistance though targeting lncRNA NEAT1 in hepatocellular carcinoma. Life Sci. 2020;242:
4. Jianchu W. miR-383 inhibits cell growth and promotes cell apoptosis in hepatocellular carcinoma by targeting IL-17 via STAT3 signaling pathway. Biomed Pharmacother. 2019;120:109551. doi:10.1016/j.biopha.2019.109551
5. Meiling L, Yu X, Lin L. ATGL promotes the proliferation of hepatocellular carcinoma cells via the p-AKT signaling pathway. J Biochem Mol Toxicol. 2019;33:e22391. doi:10.1002/jbt.22391
6. Yong J, Wenlei Z, You S, Li M, Lei L, Chen L. A nomogram for predicting depression in patients with hepatocellular carcinoma: an observational cross-sectional study. Int J Psychiatry Clin Pract. 2019;23:273–280. doi:10.1080/13651501.2019.1619777
7. Hande T, Ezgi B. lncRNA HOTAIR overexpression induced downregulation of c-Met signaling promotes hybrid epithelial/mesenchymal phenotype in hepatocellular carcinoma cells. Cell Commun Signal. 2020;18:110. doi:10.1186/s12964-020-00602-0
8. Jin LL, Jin Y, Yang H. Network of clinically-relevant lncRNAs-mRNAs associated with prognosis of hepatocellular carcinoma patients. Sci Rep. 2020;10:11124. doi:10.1038/s41598-020-67742-8
9. Wu J-H, Xu K, Liu J-H, et al. LncRNA MT1JP inhibits the malignant progression of hepatocellular carcinoma through regulating AKT. Eur Rev Med Pharmacol Sci. 2020;24:6647–6656. doi:10.26355/eurrev_202006_21651
10. Yongzi X, Yanhui L. Long non‑coding RNA H19 is involved in sorafenib resistance in hepatocellular carcinoma by upregulating miR‑675. Oncol Rep. 2020;44:165–173. doi:10.3892/or.2020.7608
11. Xuesong G. Overexpression of long noncoding RNAs (lncRNA) NF-κβ-interacting long noncoding RNA (NKILA) in ankylosing spondylitis is correlated with transforming growth factor β1 (TGF-β1), active disease and predicts length of treatment. Med Sci Monit. 2019;25:4244–4249. doi:10.12659/MSM.913813
12. Lei W, Shanshan Z, Y U M. LncRNA NR2F1-AS1 is involved in the progression of endometrial cancer by sponging miR-363 to target SOX4. Pharmazie. 2019;74:295–300. doi:10.1691/ph.2019.8905
13. Evodie P, Clément L, Collette J, et al. H19Enhancement of breast cancer cell aggressiveness by lncRNA and its mir-675 derivative: insight into shared and different actions. Cancers. 2020;12:. doi:10.3390/cancers12071730
14. Jie P, Fang H, Zhu W, et al. lncRNA nr2f1-as1 regulates mir-17/sik1 axis to suppress the invasion and migration of cervical squamous cell carcinoma cells. Reprod Sci. 2020;27:1534–1539. doi:10.1007/s43032-020-00149-y
15. Takeshi O, Nakamasa H, Masamichi T. Clinical outcomes of brain metastases from hepatocellular carcinoma: a multicenter retrospective study and a literature review. Int J Clin Oncol. 2018;23:1095–1100. doi:10.1007/s10147-018-1312-5
16. Jiangshan L, Xiaolin Z, Lu Y, Hao S, Zhang Z, Yang Y. Expression and significance of lncRNA-MINCR and cdk2 mRNA in primary hepatocellular carcinoma. Comb Chem High Throughput Screen. 2019;22:201–206. doi:10.2174/1386207322666190404151020
17. Ling L, Jingjing C, Cao M, Wang J, Wang S. Cao Mengyao NO donor inhibits proliferation and induces apoptosis by targeting PI3K/AKT/mTOR and MEK/ERK pathways in hepatocellular carcinoma cells. Cancer Chemother Pharmacol. 2019;84:1303–1314. doi:10.1007/s00280-019-03965-5
18. Yan S. Propofol exerts anticancer activity on hepatocellular carcinoma cells by raising lncRNA DGCR5. J Cell Physiol. 2020;235:2963–2972. doi:10.1002/jcp.29202
19. Aimin L, Saurav M, Haidan L, Jia P, Lee D-F, Zhao Z. H19, a long non-coding RNA, mediates transcription factors and target genes through interference of microRNAs in pan-cancer. Mol Ther Nucleic Acids. 2020;21:180–191. doi:10.1016/j.omtn.2020.05.028
20. Qiang C, Li P, Cao P. H19Genetic variant in long non-coding RNA modulates its expression and predicts renal cell carcinoma susceptibility and mortality. Front Oncol. 2020;10:785. doi:10.3389/fonc.2020.00785
21. Junyi L, Haozhuo X, Guangming D, He T, Huang W. Recombinant adenovirus (AdEasy system) mediated exogenous expression of long non-coding RNA H19 (lncRNA H19) biphasic regulating osteogenic differentiation of mesenchymal stem cells (MSCs). Am J Transl Res. 2020;12:1700–1713.
22. Shenglong L, Zheng K, Pei Y, Wang W, Zhang X. NR2F1-AS1Long noncoding RNA enhances the malignant properties of osteosarcoma by increasing forkhead box A1 expression via sponging of microRNA-483-3p. Aging. 2019;11:11609–11623. doi:10.18632/aging.102563
23. Xianjuan S, Yajing X, Hui C, Wang X, Ju S. Dysregulation of serum microRNA-574-3p and its clinical significance in hepatocellular carcinoma. Ann Clin Biochem. 2018;55:478–484. doi:10.1177/0004563217741908
24. Ren P, Zhang H. LncRNA NR2F1-AS1 promotes proliferation and metastasis of ESCC cells via regulating EMT. Eur Rev Med Pharmacol Sci. 2020;24:3686–3693. doi:10.26355/eurrev_202004_20831
25. Shibo Z, Jingqi Z, Yanhong C. Long non-coding RNA HOXA11-AS upregulates Cyclin D2 to inhibit apoptosis and promote cell cycle progression in nephroblastoma by recruiting forkhead box P2. Am J Cancer Res. 2020;10:284–298.
26. Feng G, Fu Q, Wang Y, Sui G. Long non-coding RNA NR2F1-AS1 promoted proliferation and migration yet suppressed apoptosis of thyroid cancer cells through regulating miRNA-338-3p/ CCND1 axis. J Cell Mol Med. 2019;23:5907–5919. doi:10.1111/jcmm.14386
27. Yuxian W, Yang Z, Jie H, Sun H, Jin Z. Long non-coding RNA H19 mediates ovarian cancer cell cisplatin-resistance and migration during EMT. Int J Clin Exp Pathol. 2019;12:2506–2515.
28. Feng W, Long R, Zhengkui Z. LncRNA H19-Derived miR-675-3p promotes epithelial-mesenchymal transition and stemness in human pancreatic cancer cells by targeting the STAT3 pathway. J Cancer. 2020;11:4771–4782. doi:10.7150/jca.44833
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.