")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 18
Role of Increased Syncytin-1 Expression in Pathogenesis of Anti-N-Methyl-d-Aspartate Receptor Encephalitis
Authors Qiao S, Sun QY, Zhang SC , Zhang RR, Wu YJ, Wang ZH, Liu XW
Received 15 May 2022
Accepted for publication 21 July 2022
Published 29 July 2022 Volume 2022:18 Pages 1535—1543
DOI https://doi.org/10.2147/NDT.S374798
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yuping Ning
Shan Qiao,1,2,* Quan-Ye Sun,3,* Shan-Chao Zhang,1 Ran-Ran Zhang,4 Yu-Jiao Wu,4 Zhi-Hao Wang,4 Xue-Wu Liu4,5
1Department of Neurology, The First Affiliated Hospital of Shandong First Medical University & Shandong Provincial Qianfoshan Hospital, Jinan, People’s Republic of China; 2Department of Medical Genetics, School of Basic Medical Sciences, Cheeloo College of Medicine, Shandong University, Jinan, People’s Republic of China; 3Research Center of Translational Medicine, Central Hospital Affiliated to Shandong First Medical University, Jinan, People’s Republic of China; 4Department of Neurology, Qilu Hospital of Shandong University, Jinan, People’s Republic of China; 5Institute of Epilepsy, Shandong University, Jinan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xue-Wu Liu, Email [email protected]
Purpose: Syncytin-1 may play a role in several neuropsychiatric disorders, but its function in anti-N-methyl-D-aspartate receptor (anti-NMDAR) encephalitis is unknown. The purpose of this study was to examine the possible mechanism of action of syncytin-1 in patients with anti-NMDAR encephalitis.
Patients and Methods: Twenty patients with anti-NMDAR encephalitis and eight controls were recruited. The protein levels of syncytin-1 in serum were determined using an enzyme-linked immunosorbent assay, and the transcript levels of syncytin-1 were determined using real-time quantitative PCR. Flow cytometry was used for peripheral blood lymphocyte subset detection. Further, the relationship between syncytin-1 levels and clinical features of anti-NMDAR encephalitis and peripheral blood lymphocyte subsets was analyzed.
Results: Compared with those in controls, higher syncytin-1 levels and percentage of B cells (CD3-CD19+) were observed in patients with anti-NMDAR encephalitis. Among anti-NMDAR encephalitis patients, the level of syncytin-1 positively correlated with the proportion of B cells and modified Rankin scale score at onset and after immunotherapy and negatively correlated with the proportion of CD3+ T cells.
Conclusion: An increased expression of Syncytin-1 is associated with the pathogenesis of anti-NMDAR encephalitis, providing evidence for elucidating the pathogenesis of the disease and suggesting novel therapeutic targets. Further, this study clarifies the role of syncytin-1 in neuroimmune disorders.
Keywords: autoimmune encephalitis, neuroinflammation, syncytin-1, B cells, pathogenesis
Introduction
Anti-N-methyl-d-aspartate receptor (anti-NMDAR) encephalitis, the most common autoimmune encephalitis, often manifests as cognitive dysfunction, seizures, psychobehavioral abnormalities, and movement disorders.1,2 Most patients respond well to immunotherapy, while some patients have intractable seizures and various levels of cognitive impairment, despite being treated with immunotherapy.3,4 Clarifying its pathogenesis is of great significance for its early diagnosis and treatment. The pathogenesis of anti-NMDAR encephalitis is considered to be predominantly mediated by humoral immunity and B lymphocytes.5,6 In recent years, “B-lymphocyte depletion” therapy, such as rituximab (RTX), has achieved good results in anti-NMDAR encephalitis patients, confirming the important role of B lymphocytes in the development of this disease.7 Additionally, the peripheral blood T cell subsets of patients may be able to serve as an indicator of the disease process in anti-NMDAR encephalitis and may contribute to its pathogenesis.8 However, anti-NMDAR encephalitis has not been adequately studied with regard to its etiology and pathogenesis. Understanding its etiology and pathogenesis and identifying new biomarkers and diagnostic and therapeutic targets are important for improving current treatment outcomes.
Human endogenous retrovirus-W (HERV-w), a member of the human retrovirus family, plays a vital role in various diseases, such as cancers, inflammatory disease, and autoimmune conditions.9,10 A glycoprotein encoded by HERV-W, syncytin-1, is highly expressed in trophoblast cells of the placenta and plays an important role in promoting trophoblast cell fusion.11 Syncytin-1 has gradually attracted attention in research on neurological diseases, such as multiple sclerosis (MS), neuropsychiatric disorders, and neurodegenerative diseases.12–14 Specific infections (eg, herpesvirus infection) can transactivate HERV-W sequences to upregulate syncytin-1, inducing the secretion of proinflammatory factors from peripheral blood mononuclear cells (PBMCs) to cause neuroinflammation, mediating the pathogenesis of MS.10,15 Several studies have shown that syncytin-1 is highly expressed in the serum of patients with MS and that its levels are correlated with the MS stage, active/remission phase, and treatment outcome. Based on these findings, Syncytin-1 levels may be useful as a marker for monitoring MS progression and predicting prognosis.9,10,16 Another study found syncytin-1 mRNA transcription in plasma samples from patients with schizophrenia, suggesting that syncytin-1 can also play a role in schizophrenia.17–19 Collectively, these findings indicate the important role of syncytin-1 in neuroimmune regulation. To the best of our knowledge, there has been no report on the role of syncytin-1 in anti-NMDAR encephalitis pathogenesis.
Thus, this study sought to uncover the role of syncytin-1 in anti-NMDAR encephalitis and the possible mechanisms of action with the goal of identifying potential biomarkers and therapeutic targets. Serum levels of syncytin-1 were measured in patients with anti-NMDAR encephalitis to achieve this goal.
Materials and Methods
Study Design and Population
Between January 2019 and December 2020, 20 patients with anti-NMDAR encephalitis and 8 controls were recruited from Qilu Hospital of Shandong University, China. We included patients with the following criteria: (1) acute or subacute onset of at least one clinical symptom, such as seizures, cognitive dysfunction, or speech disturbance; (2) positive anti-NMDAR antibody in serum or cerebrospinal fluid (CSF); and (3) reasonable exclusion of other diseases. We excluded patients who failed to be present for follow-ups and those with incomplete data. Anti-NMDAR encephalitis was diagnosed according to the published diagnostic criteria by Graus et al.20 Healthy blood donors were recruited from the health check-up center as controls. Samples were obtained before immunotherapy from patients with anti-NMDAR encephalitis in a state of active disease.
Sample Collection and Processing
PBMCs and serum were subsequently separated on centrifugation of peripheral blood samples obtained via venipuncture. Before use, PBMCs were frozen in fetal bovine serum supplemented with 10% DMSO (Sigma, USA). Samples were immediately stored at −80 °C until experiments were conducted.
Syncytin-1 protein levels in serum were measured using the human Syncytin-1 (ERVWE1) ELISA kit (MyBioSource, Cat.# MBS9318236), according to the manufacturer’s protocol. The results from the triplicate experiments were expressed as mean ± SEM.
RNA Isolation and qPCR Analysis
Qiagen’s RNeasy Plus mini kit was used to extract RNA from PBMCs. We used PrimeScript RT reagent Kit with gDNA Eraser (TaKaRa) to convert 1000 ng of RNA into cDNA for real-time PCR testing. The ABI 7500HT fast block PCR system (Applied Biosystems) was used to perform real-time quantitative analyses. PCR was conducted at 95°C for 10 minutes and 40 cycles of 15 seconds at 95°C and 1 minute at 60°C. The comparative cycle threshold (**Ct) method was used to calculate the relative expression levels using 18S ribosomal RNA as the endogenous control. The primers were as follows: syncytin-1: Forward 5’-TCATATCTTAAGCCCCGCAAC-3’, reverse 5’-CGCCAATGCCAGTACCTAGT-3’; GAPDH: forward 5′-GCACCGTCAAGGCTGAGACC-3′, reverse 5’-ATGGTGGTGAAGACGCCAGT-3’.
Flow Cytometry
Flow cytometry for peripheral blood lymphocyte subset detection was performed according to the manufacturer’s instructions (BD Bioscience, USA). After red cell lysis, whole blood was stained for six lymphocyte surface markers, following a standard multicolor flow cytometry procedure. In total, five subpopulations were examined: T cell (CD3+), B cells (CD3-CD19+), CD4+ T cell (CD3+ CD4+), CD8+ T cell (CD3+ CD8+), and NK cells (CD3-CD16+ CD56+).
Statistical Analysis
Variables with normal continuous distribution are expressed as means and standard deviations, whereas variables with non-normal distributions are expressed as medians and IQRs. We used the χ2 test or Fisher’s exact test to analyze categorical data, and for continuous variables, Student’s t-test was applied. To identify correlations between syncytin-1 levels and clinical features, Spearman correlation coefficient was used. Statistical analyses were conducted using SPSS 26.0 and GraphPad Prism 8.0. P values < 0.05 indicated statistical significance.
Results
Characteristics of the Study Population
Table 1 summarizes the clinicodemographic characteristics of the 20 anti-NMDAR encephalitis patients and 8 controls. A significant between-group difference was not observed in sex or age distribution (p > 0.05). In the anti-NMDAR encephalitis group, the median age at onset was 10 (IQR, 6.25–13.75) years, and the median modified Rankin scale (mRS) score at onset was 4 (range, 3–5). The common clinical symptoms were mental behavioral disorders (80%), seizures (75%), memory impairment (60%), and dyskinesia (55%). No patient had tumors as complications. Overall, 14 (70%) patients showed infection symptoms, such as fever and respiratory tract infection, before onset. Further, eight (40%) patients had abnormal brain magnetic resonance imaging signals, and 14 (70%) patients had abnormal electroencephalography findings.
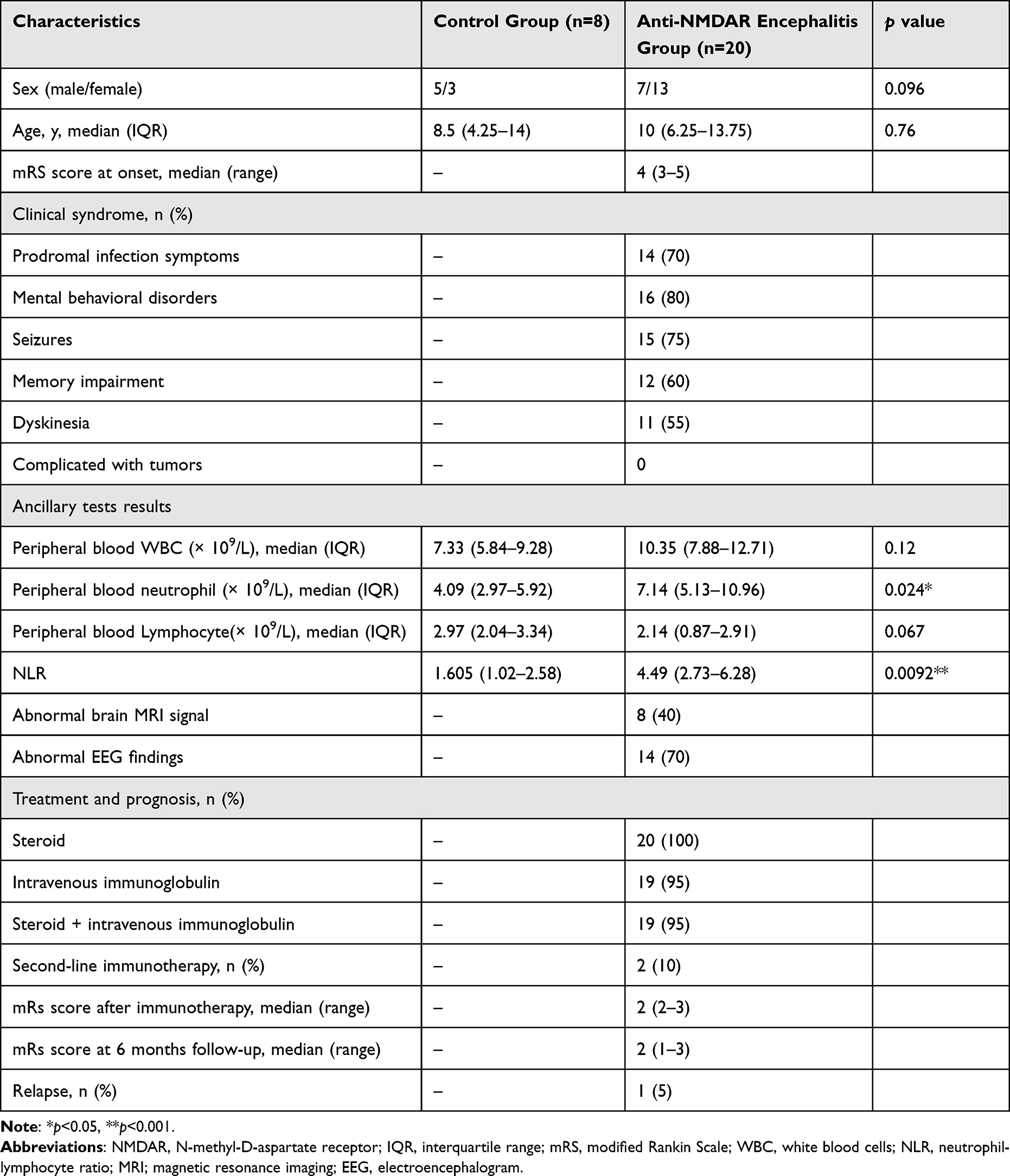 |
Table 1 Participant Characteristics by Group |
All 20 (100%) patients were treated with steroids, of whom 19 (95%) were treated with steroid and intravenous immunoglobulin. Two (10%) patients received second-line immunotherapy (both RTX). The median mRS score after immunotherapy was 2 (range, 2–3) and 2 (range, 1–3) at the 6-month follow-up. One patient (5%) experienced recurrence that mainly manifested as a fluctuation of original symptoms. The results of the auxiliary examination showed no significant difference in peripheral blood leukocyte (white blood cell) and lymphocyte counts between the two groups (p=0.12 and p=0.067, respectively). However, the median peripheral blood neutrophil count was significantly higher in the anti-NMDAR encephalitis group than in the control group (7.14 (IQR: 5.13–10.96) × 109/L vs 4.09 (IQR:2.97–5.92) × 109/L; p=0.024). The median neutrophil-to-lymphocyte ratio was also significantly higher in anti-NMDAR encephalitis (4.49 [2.73–6.28] vs 1.605 [1.02–2.58], p=0.0092).
Syncytin-1 Expression in the Control and Anti-NMDAR Encephalitis Groups
Syncytin-1 was measured using real-time PCR testing of peripheral blood monocytes (PBMC) from participants. The results showed a significant decrease in the syncytin-1 mRNA levels of patients with anti-NMDAR encephalitis when compared with those of controls (p=0.0009). In addition, the serum levels of syncytin-1 protein were also significantly higher in the anti-NMDAR encephalitis group (p=0.0003) (Figure 1).
 |
Figure 1 Expression level of syncytin-1 in anti-NMDAR encephalitis patients and controls. (A) Levels of syncytin-1 protein in serum and (B) mRNA expression in PBMC; ***p<0.0001. |
Characteristics of Peripheral Blood Lymphocyte Subsets in Controls and Anti-NMDAR Encephalitis Patients
With respect to characteristics of the peripheral blood lymphocyte subsets, a significant difference did not exist between groups in the peripheral blood CD3+ lymphocytes (p=0.1087) and the percentages of CD3+ CD4+ T cells (p=0.326), CD3+ CD8+ T cells (p=0.6818), and CD3+ CD4+/CD3+ CD8+ (p=0.9923) in the peripheral blood. However, the percentage of B cells (CD3-CD19+) was higher in the anti-NMDAR encephalitis group than in the control group (p=0.0002) but the percentage of CD3-CD16+ CD56+ lymphocytes was lower (p=0.0018) (Figure 2).
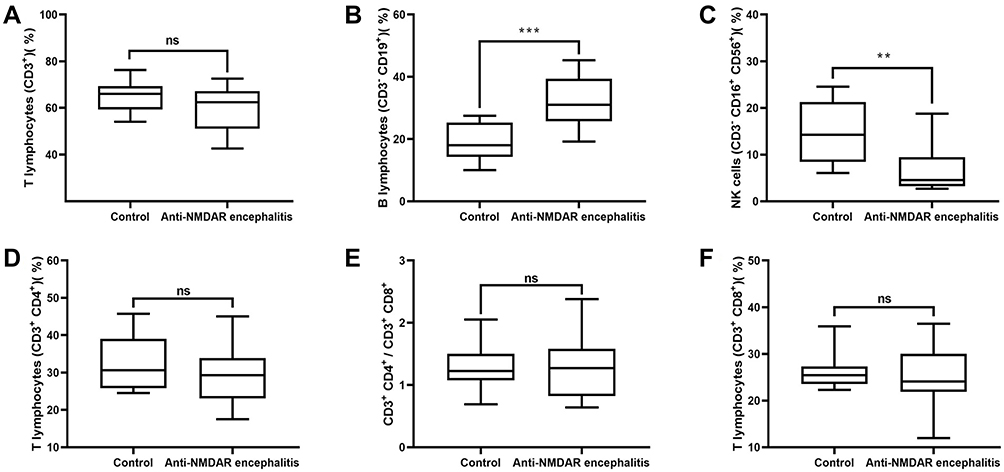 |
Figure 2 Characteristics of peripheral blood lymphocyte subsets in the control and anti-NMDAR encephalitis groups. (A) Percentages of CD3+ lymphocytes in peripheral blood in the two groups. (B) Percentages of B cells (CD3−CD19+) in peripheral blood in the two groups. (C) Percentages of CD3−CD16+ CD56+ lymphocytes in peripheral blood in the two groups. (D) Percentages of CD3+ CD4+ T cells in peripheral blood in the two groups. (E) CD3+ CD4+/CD3+ CD8+ in peripheral blood in the two groups. (F) Percentages of CD3+ CD8+ T cells in peripheral blood in the two groups. ns, no significant between-group difference; **p<0.001;***p<0.0001. |
Correlation of Syncytin-1 Expression Levels with Clinical Features and Peripheral Lymphocyte Subsets in Anti-NMDAR encephalitis
Positive correlation was found between the protein level of syncytin-1 in serum from patients with anti-NMDAR encephalitis and the proportion of CD3-CD19+ lymphocytes (r=−0.5279, 95% CI: −0.7866–0.1113, p=0.0168), while it was negatively correlated with the proportion of peripheral blood CD3+ lymphocytes (r=0.6707, 95% CI: 0.3245–0.8584, p=0.0012). In addition, the protein concentration of syncytin-1 in serum positively correlated with the mRS score at onset (r=0.6592, 95% CI: 0.3059–0.8529, p=0.0016) and after immunotherapy (r=0.6312, 95% CI: 0.2618–0.8393, p=0.0028) (Figure 3).
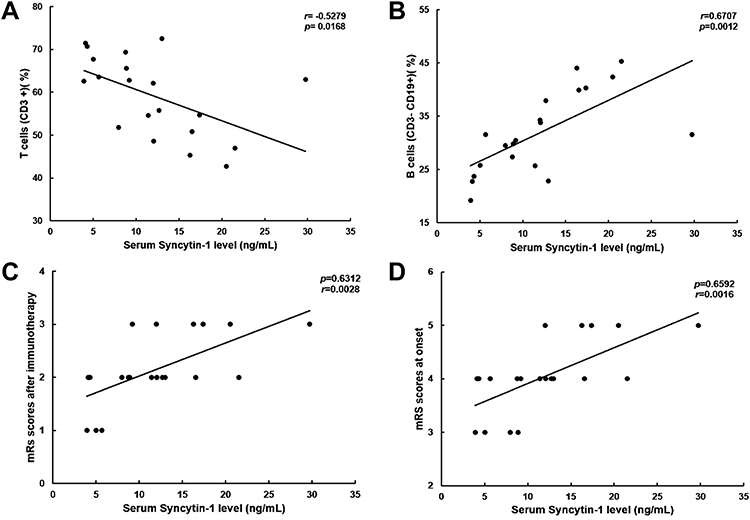 |
Figure 3 Correlation of syncytin-1 expression levels with clinical features and peripheral lymphocyte subsets in patients with anti-NMDAR encephalitis. (A) Correlation between syncytin-1 levels and the proportion of peripheral blood CD3+ lymphocytes in patients with anti-NMDAR encephalitis. (B) Correlation between syncytin-1 levels and the proportion of CD3−CD19+ lymphocytes in patients with anti-NMDAR encephalitis. (C) Correlation between syncytin-1 levels and the mRS score after immunotherapy in patients with anti-NMDAR encephalitis. (D) Correlation between syncytin-1 levels and the mRS score at onset in patients with anti-NMDAR encephalitis. |
Discussion
Till date, the role of syncytin-1 in anti-NMDAR encephalitis has not been clarified. Syncytin-1 levels in serum were significantly elevated in the acute phase of anti-NMDAR encephalitis, according to this study. To the best of our knowledge, this study is the first to report such findings and focus on the relationship between syncytin-1 levels and the clinical features of anti-NMDAR encephalitis and peripheral blood lymphocyte subsets.
Syncytin-1 expression is closely associated with the development of several autoimmune diseases, including type 1 diabetes, chronic inflammatory demyelinating polyneuropathy, and MS. Previous studies have suggested that syncytin-1 protein has proinflammatory and superantigenic activities. In addition, it exerts neurotoxic effects in vitro and in humanized or transgenic animal models, potentially causing neuroinflammation, neurodegeneration, immune system alterations, and stress responses.9,12,22,23 Several studies have reported upregulated syncytin-1 in serum and/or CSF of MS and schizophrenia, and it is thus believed to be a key factor in their pathogenesis.10,24 Rolland et al reported that syncytin-1 was involved in the pathogenesis of MS by acting on the Toll-like receptor-4 signaling pathway and potentially causing oligodendrocyte inflammatory injury.25 Collectively, these findings indicate the important role of syncytin-1 in neuroinflammatory diseases and immune regulatory processes.
Autopsy pathology of patients with anti-NMDAR encephalitis showed that the brain tissue was infiltrated by numerous inflammatory cells, which supported that its pathogenesis was mediated by immune inflammation.26 Previous studies on the regulatory mechanism of syncytin-1 in its pathogenesis have not been reported. Our study showed that syncytin-1 is highly expressed in serum in the acute phase of anti-NMDAR encephalitis and is positively correlated with the mRS score at onset and after immunotherapy. These findings indicate that syncytin-1 may play a critical role in the pathogenesis of anti-NMDAR encephalitis and is a potential biomarker for monitoring disease progression.
Increasing evidence implicates HERVs in the pathogenesis of neurological and neuropsychiatric disorders, such as MS, motor neuron disease, and schizophrenia.12,17,21 HERV-W is normally highly expressed in placental tissues, and its expression is inhibited owing to the high methylation of the CpG site in non-placental tissues.17,27,28 HERV-W is transactivated by triggers to express syncytin-1, and common triggers include infections (eg, herpes simplex virus infection, Epstein–Barr virus [EBV],) and tumors.11,29 It has been reported that transactivation of HERV sequences by herpesviruses, particularly EBV, may play a role in MS pathogenesis. Mameli et al discovered that ebvgp350 could induce the transcription of HERV-W env in monocytes, which was critical for proinflammatory cytokine release.30 Further, in multiple cancer tissues and cell lines, such as those of breast cancer and teratoma, abnormal RNA sequences of multiple HERV families have been detected.31,32
As observed in our study, syncytin-1 is activated and highly expressed in the acute phase of anti-NMDAR encephalitis; triggers and mechanisms of its activation are unknown. Previous studies have suggested that tumors—especially teratomas—and viral infections may be important triggers for the development of anti-NMDAR encephalitis.4,8,33 Some patients with anti-NMDAR encephalitis have concomitant or secondary tumors, with ovarian teratomas being the most common. Some patients with herpes simplex virus 1 encephalitis also develop anti-NMDAR encephalitis within weeks of the first encephalitis onset.34–36 No patient in our cohort developed tumors, but 70% of the patients exhibited prodromal infection symptoms prior to onset. This suggests that syncytin-1 is triggered and activated by infectious factors and participate in the pathogenesis of anti-NMDAR encephalitis. As such, long-term follow-up is crucial, and the possibility of tumors in patients cannot yet be ruled out.
As a major component of humoral immunity, B lymphocytes play a crucial role in the pathogenesis of autoimmune encephalitis.5,37,38 Autopsy pathology of patients with anti-NMDAR encephalitis showed that a large number of B cells and CD138+ cells were distributed in the perivascular, interstitial, and perivascular spaces, which provided evidence for the involvement of B cells in immune regulation.26 B cell-targeting therapies, including RTX, are effective in anti-NMDAR encephalitis, especially in recurrent and severe patients.6 These results illustrate an important immunoregulatory role for B cells in the pathogenesis of anti-NMDAR encephalitis. In the current study, the percentage of B cells (CD3-CD19+) was higher in the anti-NMDAR encephalitis group than in the control group, and the protein level of syncytin-1 in serum of patients with anti-NMDAR encephalitis positively correlated with the proportion of B cells. These findings suggest that B lymphocytes play a dominant role in syncytin-1-mediated pathogenesis of anti-NMDAR encephalitis. Although the protein level of syncytin-1 in serum negatively correlated with the proportion of peripheral blood CD3+ T lymphocytes, no significant differences were found in peripheral blood lymphocyte counts and CD4+ and CD8+ T cells between anti-NMDAR encephalitis patients and controls. Thus, we cannot conclude a major regulatory role for T lymphocytes in the pathogenesis of syncytin-1-mediated anti-NMDAR encephalitis.
A limitation of this study is the small sample size, which made it difficult to perform multivariate prognostic analyses; hence, there may have been a selection bias. Dynamic observation of patients according to their condition changes was also lacking. Selection bias and geographical factors may interfere with the results. Given the risk of invasive examination, there is a lack of observation of syncytin-1 expression in CSF. In future, studies with large sample sizes, multicenter involvement, and dynamic monitoring are needed to elucidate the in-depth mechanism of the role of syncytin-1 in the pathogenesis of anti-NMDAR encephalitis.
Conclusion
Anti-NMDAR encephalitis is associated with increased expression of syncytin-1. B lymphocytes may play a dominant role in syncytin-1-mediated pathogenesis of anti-NMDAR encephalitis, providing evidence for elucidating the pathogenesis of this disease and establishing therapeutic targets from a new perspective. Further, this study sheds new light on the function of syncytin-1 in neuropsychiatric and neuroimmune disorders.
Abbreviations
CSF, cerebrospinal fluid; ELISA, enzyme-linked immunosorbent assay; HERV-w, human endogenous retrovirus-W; IQR, interquartile range; mRS, modified Rankin Scale; MS, multiple sclerosis; NMDAR, N-methyl-D-aspartate receptor; PBMCs, peripheral blood mononuclear cells; RTX, rituximab.
Data Sharing Statement
Data support the findings of present study can be available from the corresponding author upon reasonable request.
Ethics Approval and Informed Consent
The Ethics Committee of Qilu Hospital of Shandong University approved this study (NO. KYLL-202008-044) and it was conducted according to the Declaration of Helsinki.
Consent for Publication
All participants were informed about the purpose of the study, and all participants or their legal guardians provided written consent.
Acknowledgments
The authors thank all the participants and their families for their cooperation.
Funding
This work was supported by the National Natural Science Foundation (No. 81873786).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Jafarpour S, Santoro JD. Autoimmune encephalitis. Pediatr Rev. 2022;43(4):198–211. doi:10.1542/pir.2021-005096
2. Qiao S, Wu HK, Liu LL, et al. Characteristics and prognosis of autoimmune encephalitis in the East of China: a multi-center study. Front Neurol. 2021;12:642078. doi:10.3389/fneur.2021.642078
3. Ball C, Fisicaro R, Morris L 3rd, et al. Brain on fire: an imaging-based review of autoimmune encephalitis. Clin Imaging. 2022;84:1–30. doi:10.1016/j.clinimag.2021.12.011
4. Shan W, Yang H, Wang Q. Neuronal surface antibody-medicated autoimmune encephalitis (limbic encephalitis) in China: a multiple-center, retrospective study. Front Immunol. 2021;12:621599. doi:10.3389/fimmu.2021.621599
5. Sun B, Ramberger M, O’Connor KC, RJM B-R, Irani SR. The B cell immunobiology that underlies CNS autoantibody-mediated diseases. Nat Rev Neurol. 2020;16(9):481–492. doi:10.1038/s41582-020-0381-z
6. Zhang XT, Wang CJ, Wang BJ, Guo SG. The short-term efficacy of combined treatments targeting B cell and plasma cell in severe and refractory Anti-N-methyl-D-aspartate receptor encephalitis: two case reports. CNS Neurosci Ther. 2019;25(1):151–153. doi:10.1111/cns.13078
7. Sveinsson O, Granqvist M, Forslin Y, Blennow K, Zetterberg H, Piehl F. Successful combined targeting of B- and plasma cells in treatment refractory anti-NMDAR encephalitis. J Neuroimmunol. 2017;312:15–18. doi:10.1016/j.jneuroim.2017.08.011
8. Liba Z, Kayserova J, Elisak M, et al. Anti-N-methyl-D-aspartate receptor encephalitis: the clinical course in light of the chemokine and cytokine levels in cerebrospinal fluid. J Neuroinflammation. 2016;13(1):55. doi:10.1186/s12974-016-0507-9
9. Bahrami S, Gryz EA, Graversen JH, Troldborg A, Stengaard Pedersen K, Laska MJ. Immunomodulating peptides derived from different human endogenous retroviruses (HERVs) show dissimilar impact on pathogenesis of a multiple sclerosis animal disease model. Clin Immunol. 2018;191:37–43. doi:10.1016/j.clim.2018.03.007
10. Garcia-Montojo M, Rodriguez-Martin E, Ramos-Mozo P, et al. Syncytin-1/HERV-W envelope is an early activation marker of leukocytes and is upregulated in multiple sclerosis patients. Eur J Immunol. 2020;50(5):685–694. doi:10.1002/eji.201948423
11. Guo L, Gu F, Xu Y, Zhou C. Increased copy number of syncytin-1 in the trophectoderm is associated with implantation of the blastocyst. PeerJ. 2020;8:e10368. doi:10.7717/peerj.10368
12. Wang X, Huang J, Zhu F. Human endogenous retroviral envelope protein Syncytin-1 and inflammatory abnormalities in neuropsychological diseases. Front Psychiatry. 2018;9:422. doi:10.3389/fpsyt.2018.00422
13. Bjerregard B, Ziomkiewicz I, Schulz A, Larsson LI. Syncytin-1 in differentiating human myoblasts: relationship to caveolin-3 and myogenin. Cell Tissue Res. 2014;357(1):355–362. doi:10.1007/s00441-014-1930-9
14. Antony JM, Izad M, Bar-Or A, et al. Quantitative analysis of human endogenous retrovirus-W env in neuroinflammatory diseases. AIDS Res Hum Retroviruses. 2006;22(12):1253–1259. doi:10.1089/aid.2006.22.1253
15. Brutting C, Stangl GI, Staege MS. Vitamin D, Epstein-Barr virus, and endogenous retroviruses in multiple sclerosis - facts and hypotheses. J Integr Neurosci. 2021;20(1):233–238. doi:10.31083/j.jin.2021.01.392
16. Lycke J. Trials of antivirals in the treatment of multiple sclerosis. Acta Neurol Scand. 2017;136(Suppl 201):45–48. doi:10.1111/ane.12839
17. Yan Q, Wu X, Zhou P, et al. HERV-W envelope triggers abnormal dopaminergic neuron process through DRD2/PP2A/AKT1/GSK3 for schizophrenia risk. Viruses. 2022;14(1):1. doi:10.3390/v14010145
18. Tamouza R, Meyer U, Foiselle M, et al. Identification of inflammatory subgroups of schizophrenia and bipolar disorder patients with HERV-W ENV antigenemia by unsupervised cluster analysis. Transl Psychiatry. 2021;11(1):377. doi:10.1038/s41398-021-01499-0
19. Aftab A, Shah AA, Hashmi AM. Pathophysiological role of HERV-W in schizophrenia. J Neuropsychiatry Clin Neurosci. 2016;28(1):17–25. doi:10.1176/appi.neuropsych.15030059
20. Graus F, Titulaer MJ, Balu R, et al. A clinical approach to diagnosis of autoimmune encephalitis. Lancet Neurol. 2016;15(4):391–404. doi:10.1016/S1474-4422(15)00401-9
21. Kristensen MK, Christensen T. Regulation of the expression of human endogenous retroviruses: elements in fetal development and a possible role in the development of cancer and neurological diseases. APMIS. 2021;129(5):241–253. doi:10.1111/apm.13130
22. Fominykh V, Brylev L, Gaskin V, et al. Neuronal damage and neuroinflammation markers in patients with autoimmune encephalitis and multiple sclerosis. Metab Brain Dis. 2019;34(5):1473–1485. doi:10.1007/s11011-019-00452-x
23. Latifi T, Zebardast A, Marashi SM. The role of human endogenous retroviruses (HERVs) in Multiple Sclerosis and the plausible interplay between HERVs, Epstein-Barr virus infection, and vitamin D. Mult Scler Relat Disord. 2022;57:103318. doi:10.1016/j.msard.2021.103318
24. Marrodan M, Alessandro L, Farez MF, Correale J. The role of infections in multiple sclerosis. Mult Scler. 2019;25(7):891–901. doi:10.1177/1352458518823940
25. Rolland A, Jouvin-Marche E, Viret C, Faure M, Perron H, Marche PN. The envelope protein of a human endogenous retrovirus-W family activates innate immunity through CD14/TLR4 and promotes Th1-like responses. J Immunol. 2006;176(12):7636–7644. doi:10.4049/jimmunol.176.12.7636
26. Martinez-Hernandez E, Horvath J, Shiloh-Malawsky Y, Sangha N, Martinez-Lage M, Dalmau J. Analysis of complement and plasma cells in the brain of patients with anti-NMDAR encephalitis. Neurology. 2011;77(6):589–593. doi:10.1212/WNL.0b013e318228c136
27. Soygur B, Sati L, Demir R. Altered expression of human endogenous retroviruses syncytin-1, syncytin-2 and their receptors in human normal and gestational diabetic placenta. Histol Histopathol. 2016;31(9):1037–1047.
28. Wang X, Wu X, Huang J, Li H, Yan Q, Zhu F. Human endogenous retrovirus W family envelope protein (HERV-W env) facilitates the production of TNF-alpha and IL-10 by inhibiting MyD88s in glial cells. Arch Virol. 2021;166(4):1035–1045. doi:10.1007/s00705-020-04933-8
29. Wang X, Liu Z, Wang P, et al. Syncytin-1, an endogenous retroviral protein, triggers the activation of CRP via TLR3 signal cascade in glial cells. Brain Behav Immun. 2018;67:324–334. doi:10.1016/j.bbi.2017.09.009
30. Mameli G, Poddighe L, Mei A, et al. Expression and activation by Epstein Barr virus of human endogenous retroviruses-W in blood cells and astrocytes: inference for multiple sclerosis. PLoS One. 2012;7(9):e44991. doi:10.1371/journal.pone.0044991
31. Fu Y, Zhuang X, Xia X, Li X, Xiao K, Liu X. Correlation between promoter hypomethylation and increased expression of Syncytin-1 in non-small cell lung cancer. Int J Gen Med. 2021;14:957–965. doi:10.2147/IJGM.S294392
32. Zhang M, Liang JQ, Zheng S. Expressional activation and functional roles of human endogenous retroviruses in cancers. Rev Med Virol. 2019;29:2.
33. Bien CG. Management of autoimmune encephalitis. Curr Opin Neurol. 2021;34(2):166–171. doi:10.1097/WCO.0000000000000909
34. Dubey D, Pittock SJ, Kelly CR, et al. Autoimmune encephalitis epidemiology and a comparison to infectious encephalitis. Ann Neurol. 2018;83(1):166–177. doi:10.1002/ana.25131
35. Venkatesan A, Benavides DR. Autoimmune encephalitis and its relation to infection. Curr Neurol Neurosci Rep. 2015;15(3):3. doi:10.1007/s11910-015-0529-1
36. Blackburn KM, Wang C. Post-infectious neurological disorders. Ther Adv Neurol Disord. 2020;13:1756286420952901. doi:10.1177/1756286420952901
37. Handono K, Pratama MZ, Sari DK, et al. Effect of active immunization with IL-17A on B cell function and infection risk in pristane-induced lupus model. Int J Rheum Dis. 2018;21(6):1277–1286. doi:10.1111/1756-185X.13325
38. Sabatino JJ
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.