")
Back to Journals » OncoTargets and Therapy » Volume 13
Review of Research on the Role of Irisin in Tumors
Authors Zhang D, Tan X, Tang N, Huang F, Chen Z, Shi G
Received 8 January 2020
Accepted for publication 7 May 2020
Published 19 May 2020 Volume 2020:13 Pages 4423—4430
DOI https://doi.org/10.2147/OTT.S245178
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Carlos E Vigil
Deguo Zhang,1,2 Xueying Tan,2 Nan Tang,1 Fei Huang,3 Zengyin Chen,1,* Guangjun Shi2,*
1Department of Hepatobiliary Surgery, The Affiliated Qingdao Chengyang District People’s Hospital of Qingdao University, Qingdao 266109, Shandong Province, People’s Republic of China; 2Department of Hepatobiliary Surgery, The Affiliated Qingdao Municipal Hospital of Qingdao University, Qingdao 266000, Shandong Province, People’s Republic of China; 3Department of Gastrointestinal Hernia Surgery, Qianxinan People’s Hospital, Xingyi 562400, Guizhou Province, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zengyin Chen
Department of Hepatobiliary Surgery, The Affiliated Qingdao Chengyang District People’s Hospital of Qingdao University, No. 600, Changcheng Road, Qingdao 266109, Shandong Province, People’s Republic of China
Tel +86 178 0623 1116
Email [email protected]
Guangjun Shi
Department of Hepatobiliary Surgery, The Affiliated Qingdao Municipal Hospital of Qingdao University, No. 1, Jiao Zhou Road, Shibei District, Qingdao 266000, Shandong Province, People’s Republic of China
Tel +86 186 6167 5603
Email [email protected]
Abstract: Irisin is a newly discovered exercise-induced cytokine, produced by the proteolytic hydrolysis of fibronectin type III domain-containing protein 5 (FNDC5). Irisin is widely distributed in the human body and is involved in the browning of white adipose tissue, improving insulin resistance, improving cognitive function, and regulating bone metabolism. Recent studies have shown that irisin concentration is elevated in a variety of tumor tissues as compared with that in normal tissues. However, irisin has different effects on the proliferation and apoptosis of tumor cells in breast cancer, lung cancer, and liver cancer through various mechanisms. Irisin plays an important role in the occurrence, development, and metastasis of different tumors, suggesting that irisin can be used as a potential target for tumor diagnosis and treatment. Therefore, studying the expression and function of irisin in tumors may be of great significance for the prevention and treatment of tumors. This article reviews the research progress on the role of irisin in tumors.
Keywords: irisin, cancer, diagnosis, treatment
Introduction
Irisin is an exercise-induced cytokine that promotes the conversion of white adipose tissue to brown adipose tissue. Brown adipose tissue contains a large concentration of uncoupling protein-1 (UCP-1), which can dissipate the energy generated by oxidization in the mitochondria as thermal energy and promote energy consumption, thus playing a role in regulating energy metabolism.1 Recent studies have documented that irisin is involved in the occurrence and development of a variety of tumors. For example, researchers recently found that patients with bladder or prostate cancer had significantly lower serum irisin levels than the control subjects.2,3 The results indicate that irisin can be a potential target for tumor diagnosis and treatment. Studies have shown that exercise can reduce the risk of cancer and improve patient prognosis through direct or indirect anticancer mechanisms such as metabolism, epigenetics, and mental health.4,5 It is known that continuous exercise is related to the production of irisin, suggesting that irisin may be a part of the molecular mechanisms underlying the benefits of exercise. The present study reviews the structure and function of irisin and the research on the role of irisin in tumors (Table 1).
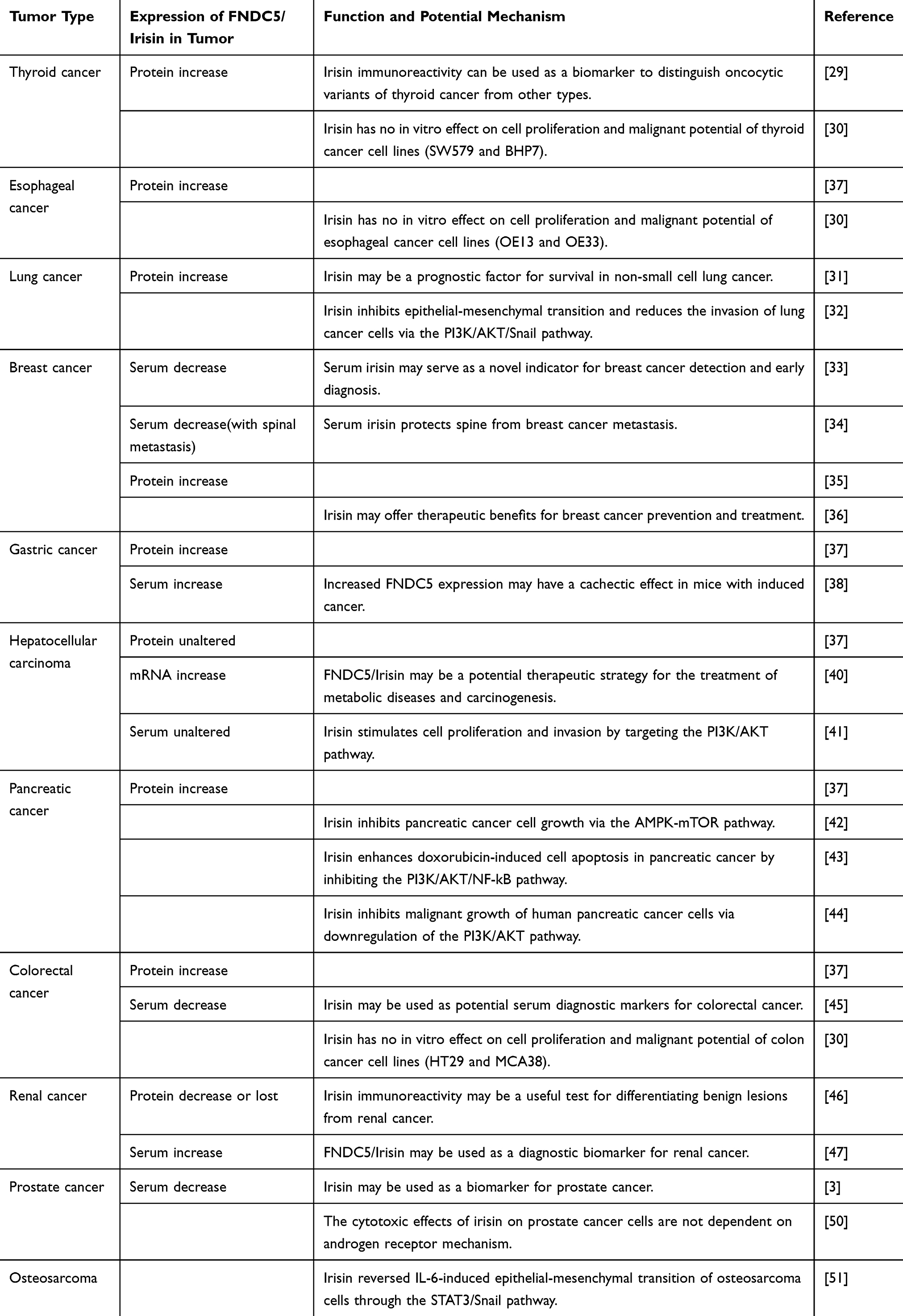 |
Table 1 Expression of FNDC5/Irisin and Related Molecular Mechanism in Tumors |
Structure and Biological Characteristics of Irisin
The precursor of irisin, FNDC5, is composed of 209 amino acids with a relative molecular weight of about 3.2 × 104. The sequence from the N-terminus to the C-terminus comprises signal peptide (SP), fibronectin Ⅲ domain (FND), hydrophobic domain (H), and c-terminal domain (C).1,6 FNDC5 is hydrolyzed by protease at amino acid site 30 and site 142 to produce irisin under the action of peroxisome proliferator-activated receptor gamma coactivator 1-α (PGC1-α). Irisin is an N-glycosylated protein factor with a relative molecular weight of 2.2 × 104, consisting of 112 amino acid residues. Irisin has two glycosylation sites, and the relative molecular weight after deglycosylation is about 1.5 × 10.4,7 Irisin is highly conservative and its amino acid sequence is identical between humans and mice.1
Irisin was first found in the cells of skeletal muscle. It has been proven that irisin is expressed in almost all tissues and organs of eukaryotic organisms.8,9 It was found that FNDC5 is highly expressed in skeletal muscles and muscle-rich tissues (such as pericardium and rectum), moderately expressed in the heart, and weakly expressed in the brain, kidney, liver, lung, and adipose tissue.10 In addition, FNDC5 is also expressed in the ovary, testis, and thyroid gland. Irisin can also be found in tissue and body fluids (such as plasma, cerebrospinal fluid, saliva, and milk).11–13
Physiological Function of Irisin
Inducing Browning of White Fat
Studies have shown that exercise can induce the expression of PGC1-α in skeletal muscle and promote the expression of FNDC5, thereby resulting in an increase in serum irisin levels and “browning” of white adipose tissue.14,15 White adipose tissue and brown adipose tissue are two different types of adipose tissues in the human body, which have different functions. White adipose tissue is mainly composed of triacylglycerol, which is the main constituent of fat stored the body. It has important functions such as maintaining normal body temperature and protecting important organs. Promoting uncoupling of oxidative respiratory chain electron transfer, generating adenosine triphosphate (ATP), and promoting dissipation of energy in the form of thermal energy by using a large amount of UCP-1 in the mitochondria, thereby maintaining the energy balance in the body, are the main functions of brown adipose tissue.16
After intraperitoneal administration of 50 μg of FNDC5 antibody to rats after 7 days of exercise, the up-regulation of UCP-1 expression in subcutaneous adipose tissue caused by exercise was suppressed, confirming that FNDC5 promotes the transformation of white adipocytes into brown adipocytes.1 However, the mechanism underlying the browning of white fat is not clear. Studies have shown that irisin promotes the expression of related genes by activating the p38 MAPK/ERK pathway involved in the browning of white fat.7
Improving Insulin Resistance
Appropriate exercise causes skeletal muscle to secrete irisin, which increases glucose uptake and expression of glucose transporter 4, and to increase glucose utilization and sensitivity of skeletal muscle to insulin.17 Studies have shown that exogenous irisin significantly reduced insulin resistance in insulin-resistant rats in whom resistance was induced by a high-fat diet.1 Type-2 diabetes mellitus (T2DM) is a heterogeneous disease characterized by insulin resistance, with decreased insulin secretion and increased glucose production. Choi et al18 found that as compared to subjects with normal glucose tolerance, T2DM patients showed significantly reduced serum irisin levels, suggesting that irisin plays an important role in the development of T2DM. Therefore, irisin plays an important regulatory role in reducing insulin resistance and may become a new target for diabetes treatment.
Improving Cognitive Function
The continuous formation of hippocampal neurons is necessary for the survival of humans and other vertebrates and for coping with the external environment. Brain-derived neurotrophic factor (BNDF) can promote the growth, differentiation, survival, and repair of neurons and maintain the structure and function of the nervous system, thereby improving cognitive ability.19 Studies have shown that FNDC5 protein injections given into non-motile mice promoted the expression of the BNDF gene in mouse brains, thereby promoting neuronal growth; however, when the FNDC5 gene was knocked out in mouse embryonic stem cells, neuronal differentiation was inhibited.20,21 In addition, irisin was found to promote the proliferation of mouse H19-7 cells in vitro at a pharmacological concentration and promote the STAT3 signal transduction pathway required for sensory neuron development.22 These studies suggest that irisin plays a potential role in the treatment of cognitive decline in the elderly, including neuropsychiatric diseases such as Alzheimer’s disease. However, further exploratory research on neurological diseases and cognitive functions is needed.
Regulation of Bone Metabolism
Exercise is a good way to maintain bone formation and absorption balance and prevent diseases such as osteoporosis and bone metabolism disorders.23 During human movement, some cytokines are secreted due to the interactions between the bone and skeletal muscles, affecting the physiological state.24 Colaianni et al25 conducted a study in animal models and found that irisin helped prevent and restore bone loss and hind-limb muscle atrophy in mice. Anastasilakis et al26 found that proportion of fractures caused by osteoporosis in postmenopausal women was correlated to the level of irisin in the blood. Palermo et al27 found a negative correlation between serum irisin levels and vertebral fractures and that irisin’s protective effect on the bones was not achieved by increasing bone density. Colaianni et al28 injected low doses of irisin into male mice and observed a significant increase in their cortical bone mass and strength. However, the specific mechanism of irisin-mediated bone metabolism is not clear, and further research is needed in this regard.
Irisin and Cancer
Thyroid Cancer
Ugur et al29 carried out immunohistochemical studies to detect the expression of irisin in thyroid tumors. The results showed a significant increase in irisin synthesis in oncocytic variants of thyroid cancer as compared to that in other thyroid tumors. Therefore, irisin immunoreactivity can be used as a biomarker to distinguish oncocytic variants of thyroid cancer from other types. Moon et al30 treated obesity-related cancer cell lines with irisin at physiological (5 to 10 nmol/L) and higher than physiological/pharmacological (50 to 100 nmol/L) concentrations. After 36 h of incubation, irisin had no in vitro effects on cell proliferation and malignant potential of obesity-related cancer cell lines, namely endometrial (KLE and RL95-2), colon (HT29 and MCA38), thyroid (SW579 and BHP7), and esophageal (OE13 and OE33).
Lung Cancer
Katarzyna et al31 found that irisin was elevated in the cancer cells and tumor stromal fibroblasts of patients with non-small cell lung cancer (NSCLC). The expression levels of irisin in cancer cells vary between NSCLC subtypes, eg, the expression levels are higher in adenocarcinoma (AC) than in squamous-cell carcinoma (SCC). The expression of irisin in stromal fibroblasts may affect the proliferation of NSCLC cells, and this may be a prognostic factor for NSCLC patients. Shao et al32 treated human lung cancer cells with different concentrations (0, 10, 20, and 50 nM) of irisin for different time periods (0, 24, and 48 h), and the results showed that irisin significantly inhibited the proliferation, migration, and invasion of lung cancer cells in a time-dependent manner. With the increase in irisin concentration, the expression of E-cadherin increased, while the expression of N-cadherin, vimentin, and Snail decreased. At the same time, LY294002, an inhibitor of PI3K, could reverse the effect of irisin. The results showed that irisin could inhibit epithelial-mesenchymal transition (EMT) and the invasion of lung cancer cells through the PI3K/Akt/Snail pathway, suggesting that irisin can be a potential drug candidate for lung cancer metastasis.
Breast Cancer
Provatopoulou et al33 studied 51 healthy women and 101 patients with invasive ductal breast cancer and found that serum irisin levels were significantly reduced in breast cancer patients. According to the experimental data, irisin level was closely related to the occurrence of breast cancer and was an independent predictor of breast cancer. It is estimated that for every one-unit increase in irisin levels, the risk of breast cancer decreases by nearly 90%. Irisin could effectively discriminate breast cancer patients at a cut-off point of 3.21 μg/mL, with 62.7% sensitivity and 91.1% specificity. Meanwhile, the serum level of irisin was positively correlated with tumor stage. These results suggest that irisin can be used in the early diagnosis, differential diagnosis, and prognosis of breast cancer.
Studies have shown that obesity increases the mortality risk of breast cancer patients by causing an increase in the concentration of estrogen and elevating the risk of inflammation and metabolic syndrome (MS). MS is a clinical syndrome characterized by disorders of metabolic factors such as hypertension, obesity, and hyperglycemia. Zhang et al34 studied 148 breast cancer patients, of whom, 53 (35.8%) had spinal metastases. The results showed that the serum level of irisin was higher in MS patients. Patients without spinal metastases had higher serum levels of irisin than the controls. Multivariate analysis showed that serum irisin had a protective effect against spinal metastases in breast cancer patients after adjusting for age and body mass index (BMI). In addition, there was a positive correlation between serum irisin levels and patient BMI. The results suggest that irisin can protect the spine from breast cancer metastasis. Kuloglu et al35 studied the breast tissue by immunohistochemical staining and found that the expression level of irisin was significantly higher in invasive lobular carcinoma, intraductal papilloma, invasive ductal carcinoma, invasive papillary carcinoma, and mucinous carcinoma tissue than in normal breast tissue. Gannon et al36 used different concentrations of irisin (0.625–20 nmol/L) to treat breast cancer cells and found that irisin could significantly reduce the cell number, migration, and viability and enhance the cytotoxic effect of doxorubicin (Dox) in malignant MDA-MB-231 cells, without affecting non-malignant MCF-10a cells. In addition, irisin enhanced the activity of caspase-3 and caspase-7, thus inducing apoptosis of MDA-MB-231 cells, and inhibited the activity of nuclear factor kappa B (NF-κb), thus indicating its anti-inflammatory role.
Tumors of the Digestive System
Gastric Cancer
The immunohistochemical studies of Aydin et al37 showed that irisin was expressed in normal gastric tissues, and there was no significant difference the expression of irisin and that of gastric neuroendocrine carcinoma tissues. However, the expression of irisin in gastric adenocarcinoma and signet ring cell carcinoma tissues was significantly higher than that in normal gastric tissues. Altay et al38 induced gastric cancer in mice and detected the expression level of the FNDC5 gene in stomach tissue, skeletal muscle, and fat. The results showed that as compared with the control group, the gastric cancer group showed no significant change in the expression of FNDC5 in skeletal muscle but showed a significant increase in its expression in fat tissue. Increased FNDC5 expression in white and brown adipose tissues may have a cachectic effect in mice with induced cancer.
Hepatocellular Carcinoma (HCC)
Previous studies have shown that in the liver, irisin is produced by hepatocytes, Kupffer cells, and sinusoidal endothelial cells, but its role in the liver is still unknown.39 Gaggini et al40 studied the liver tissue of 18 patients with HCC undergoing liver transplantation and of 18 deceased donors and found no significant difference in plasma irisin content between patients with HCC and deceased donors. At the same time, irisin, stearoyl COA dehydrogenase-1 (SCD-1), sterol regulatory element binding factor-1 (Srebf-1), and Notch1 gene expression were detected in the liver tissue. In addition to Srebf-1, irisin, SCD-1, and Notch1 levels were significantly higher in HCC livers than in deceased donors’ livers. Gaggini et al speculate that during the development of liver cancer, irisin acts as a paracrine hormone to inhibit the production of lipids and limit the adipogenesis induced by cancer progression, thus exerting a protective effect on the liver. Aydin et al37 found irisin in the hepatocytes of normal liver tissue, but there was no significant difference in irisin levels between HCC liver and normal liver tissue, which is not consistent with the results of Gaggini et al. There was a lack of consistency in the differences in irisin mRNA and protein expressions between HCC and normal liver tissues in both studies. These differences could be attributed to different tissue sample collection standards and detection methods and to the changes in mRNA transcription and post-translational modifications, such as metabolic degradation of proteins. Shi et al41 found that irisin significantly increased cell proliferation, invasion, and migration by activating the PI3K/AKT pathway and decreasing the cytotoxicity of Dox.
Pancreatic Cancer
Liu et al42 studied the effect of different concentrations (0, 5, 10, 50, and 100 nM) of irisin on pancreatic cancer cell lines (MIA PaCa-2 and Panc03.27) for different time periods (24, 48, and 72 h). The results showed that irisin inhibited the growth of pancreatic cancer cells via the AMPK-mTOR pathway and suppressed the migration and invasion of pancreatic cancer cells by inhibiting EMT. In addition, Liu et al43 found that irisin increased the chemosensitivity of pancreatic cancer cells to Dox or gemcitabine by inactivating the PI3K/AKT/NF-kB signaling pathway. Zhang et al44 treated pancreatic cancer cell lines (PANC-1 and BxPC-3) with different concentrations (0, 10, 20 and 50 nM) of irisin for different time periods (24, 48, and 72 h). The results showed that irisin inhibited cell proliferation and induced apoptosis of pancreatic cancer cells in a dose-dependent manner. In addition, irisin decreased the migration and invasion of pancreatic cancer cells. Finally, Western blot analysis revealed that irisin downregulated the PI3K/AKT signaling pathway. Therefore, irisin could be further developed as a novel potential therapeutic agent for the treatment of pancreatic cancer.
Colorectal Cancer (CRC)
Aydin et al37 investigated the expression of irisin in colon cancers using immunocytochemistry. As compared with normal colon cells, colon adenocarcinoma and mucinous colon adenocarcinoma cells showed a significant increase in irisin immunoreactivity. Zhu et al45 recruited 76 patients with CRC and 40 healthy controls and measured the serum levels of activating transcription factor 3 (ATF3) and irisin by enzyme-linked immunosorbent assay (ELISA). The results showed that the serum level of ATF3 in CRC patients increased by 37.2% and the irisin level decreased by 23.3% compared with the levels in the healthy control group. CRC was independently related to serum ATF3 and irisin levels. After adjusting for age, sex, BMI, and other biochemical parameters, individuals with high ATF3 levels were 22.3 times more likely to develop colorectal cancer than those with low ATF3 levels, while those with high irisin levels were 78.0% less likely to develop CRC than those with low irisin levels. The combination of ATF3 and irisin improves the accuracy of diagnosis, suggesting that ATF3 and irisin can be used as potential serum diagnostic markers for CRC.
Renal Cancer
Kuloglu et al46 used immunohistochemical staining to analyze human renal cancers. Irisin immunoreactivity was absent in Fuhrman nuclear grade 1, 2, and 3 clear-cell renal-cell carcinoma (RCC) and papillary RCC samples and was low in chromophobe RCC samples. Irisin immunoreactivity may be a useful test for differential diagnosis of RCC. Altay et al47 recruited 23 renal cancer patients and 25 healthy controls and collected their blood samples and measured the serum levels of FNDC5/irisin and carcinoembryonic antigen (CEA) by ELISA. The results showed that the serum levels of FNDC5/irisin and CEA in renal cancer patients were significantly higher than those in healthy controls. In addition, FNDC5/irisin levels are more sensitive and specific than CEA levels. The results suggested that serum FNDC5/irisin could be used as a biomarker for the diagnosis of renal cancer.
Prostate Cancer
Prostate cancer is a hormone-dependent tumor, and about 90% of prostate cancer cases are related to androgen stimulation, so blocking the action of androgen is an important step in treating prostate cancer.48 However, prostate cancer has heterogeneity, and the effect of endocrine therapy on a few patients is not obvious.49 Tekin et al50 treated the androgen receptor-positive and negative human prostate cancer cells LNCaP and DU-145, respectively, with different concentrations (0, 0.1, 1, 10 and 100 nM) of irisin for 24 h. The results showed that irisin could inhibit the proliferation of prostate cancer cells in a dose-dependent manner. The results indicated that the toxic effect of irisin on prostate cancer cells does not depend on the androgen receptor, indicating the potential of this new approach in treating prostate cancer.
Osteosarcoma
Kong et al51 treated osteosarcoma cells U2OS and MG-63 with different concentrations (0, 25, 50, 100, and 200 ng/mL) of irisin for different time points (12, 24, and 48 h). The results showed that irisin inhibited the proliferation, migration, and invasion of U2OS and MG-63 osteosarcoma cells in a dose- and time-dependent manner. Meanwhile, irisin reversed the IL-6-induced action of EMT in osteosarcoma cells by regulating the expression of E-cadherin, vimentin, fibronectin, matrix metalloproteinases (MMP)-2, MMP-7, and MMP-9. In addition, irisin inhibited the phosphorylation of signal transducers and activators of transcription (STAT) 3 and the expression of Snail activated by IL-6 in osteosarcoma cells. Finally, the inhibition of STAT3 by WP1066 further enhanced the effect of irisin on the expression of EMT and Snail in osteosarcoma cells. The results suggest that irisin may be a potential drug for the treatment of osteosarcoma metastasis.
Conclusion
In summary, irisin plays an important role in the diagnosis, treatment, and prognosis of tumors. Considering the results of studies analyzed in this review, the protective effect of irisin has been demonstrated in several in vitro studies. However, there are still many contradictions in the research results of different experimental institutions, possibly due to differences in the (1) detection or experimental methods used, (2) detection reagents or antibodies used, (3) tissue or cell specificity, and (4) receptors or pathways. Therefore, follow-up experiments are needed to improve experimental methods and reagents used in the study of irisin. At the same time, the receptors, pathways, and mechanisms involved in irisin action need to be further clarified.
The increased expression of irisin in different tumors suggests that it is closely related to the occurrence and development of tumors, making it an attracting new target for tumor prevention research. However, the mechanism of tumorigenesis and development is very complicated, and often, multiple influencing factors and signaling pathways are involved. At present, there are a few reports on the effects of irisin on tumors. Therefore, in-depth study of the role of irisin in tumorigenesis and development will help create a new approach for the prevention and treatment of tumors.
Acknowledgments
This work was supported by the Shandong Province Key Research and Development Project (#2018GSF118057), Qingdao Science and Technology Bureau Project (#19-6-2-58-cg), Qingdao Livelihood Science and Technology Project (#17-3-3-40-nsh), and Qingdao Outstanding Health Professional Development Fund.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bostrom P, Wu J, Jedrychowski MP, et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature. 2012;481(7382):463–468. doi:10.1038/nature10777
2. Esawy MM, Abdel-Samd KM. The diagnostic and prognostic roles of serum irisin in bladder cancer. Curr Probl Cancer. 2019;100529. doi:10.1016/j.currproblcancer.2019.100529
3. Aslan R, Alp HH, Eryilmaz R, et al. Can the irisin be a biomarker for prostate cancer? A case control study. Asian Pac J Cancer Prev. 2020;21(2):505–509. doi:10.31557/APJCP.2020.21.2.505
4. Thomas RJ, Kenfield SA, Jimenez A. Exercise-induced biochemical changes and their potential influence on cancer: a scientific review. Br J Sports Med. 2017;51(8):640–644. doi:10.1136/bjsports-2016-096343
5. Friedenreich CM, Neilson HK, Lynch BM. State of the epidemiological evidence on physical activity and cancer prevention. Eur J Cancer. 2010;46(14):2593–2604. doi:10.1016/j.ejca.2010.07.028
6. Erickson HP. Irisin and FNDC5 in retrospect: an exercise hormone or a transmembrane receptor? Adipocyte. 2013;2(4):289–293. doi:10.4161/adip.26082
7. Zhang Y, Li R, Meng Y, et al. Irisin stimulates browning of white adipocytes through mitogen-activated protein kinase p38 MAP kinase and ERK MAP kinase signaling. Diabetes. 2014;63(2):514–525. doi:10.2337/db13-1106
8. Aydin S. Three new players in energy regulation: preptin, adropin and irisin. Peptides. 2014;56:94–110. doi:10.1016/j.peptides.2014.03.021
9. Aydin S, Kuloglu T, Aydin S, et al. A comprehensive immunohistochemical examination of the distribution of the fat-burning protein irisin in biological tissues. Peptides. 2014;61:130–136. doi:10.1016/j.peptides.2014.09.014
10. Huh JY, Panagiotou G, Mougios V, et al. FNDC5 and irisin in humans: I. Predictors of circulating concentrations in serum and plasma and II. mRNA expression and circulating concentrations in response to weight loss and exercise. Metabolism. 2012;61(12):1725–1738. doi:10.1016/j.metabol.2012.09.002
11. Aydin S, Kuloglu T, Aydin S. Copeptin, adropin and irisin concentrations in breast milk and plasma of healthy women and those with gestational diabetes mellitus. Peptides. 2013;47:66–70. doi:10.1016/j.peptides.2013.07.001
12. Aydin S, Aydin S, Kuloglu T, et al. Alterations of irisin concentrations in saliva and serum of obese and normal-weight subjects, before and after 45 min of a Turkish bath or running. Peptides. 2013;50:13–18. doi:10.1016/j.peptides.2013.09.011
13. Piya MK, Harte AL, Sivakumar K, et al. The identification of irisin in human cerebrospinal fluid: influence of adiposity, metabolic markers, and gestational diabetes. Am J Physiol Endocrinol Metab. 2014;306(5):E512–518. doi:10.1152/ajpendo.00308.2013
14. Castillo-Quan JI. From white to brown fat through the PGC-1alpha-dependent myokine irisin: implications for diabetes and obesity. Dis Model Mech. 2012;5(3):293–295. doi:10.1242/dmm.009894
15. Elsen M, Raschke S, Eckel J. Browning of white fat: does irisin play a role in humans? J Endocrinol. 2014;222(1):R25–38. doi:10.1530/JOE-14-0189
16. Sammons MF, Price DA. Modulation of adipose tissue thermogenesis as a method for increasing energy expenditure. Bioorg Med Chem Lett. 2014;24(2):425–429. doi:10.1016/j.bmcl.2013.12.016
17. Liu J. Irisin as an exercise-stimulated hormone binding crosstalk between organs. Eur Rev Med Pharmacol Sci. 2015;19(2):316–321.
18. Choi YK, Kim MK, Bae KH, et al. Serum irisin levels in new-onset type 2 diabetes. Diabetes Res Clin Pract. 2013;100(1):96–101. doi:10.1016/j.diabres.2013.01.007
19. Bekinschtein P, Cammarota M, Medina JH. BDNF and memory processing. Neuropharmacology. 2014;76(PtC):677–683. doi:10.1016/j.neuropharm.2013.04.024
20. Wrann CD, White JP, Salogiannnis J, et al. Exercise Induces Hippocampal BDNF through a PGC-1α/FNDC5 Pathway. Cell Metab. 2013;18(5):649–659. doi:10.1016/j.cmet.2013.09.008
21. Hashemi MS, Esfahani AK, Peymani M, et al. Zinc finger protein 521 overexpression increased transcript levels of Fndc5 in mouse embryonic stem cells. J Biosci. 2016;41(1):69–76. doi:10.1007/s12038-015-9578-5
22. Moon HS, Dincer F, Mantzoros CS. Pharmacological concentrations of irisin increase cell proliferation without influencing markers of neurite outgrowth and synaptogenesis in mouse H19-7 hippocampal cell lines. Metabolism. 2013;62(8):1131–1136. doi:10.1016/j.metabol.2013.04.007
23. Banfi G, Lombardi G, Colombini A, Lippi G. Bone metabolism markers in sports medicine. Sports Med. 2010;40(8):697–714. doi:10.2165/11533090-000000000-00000
24. Guo B, Zhang ZK, Liang C, et al. Molecular communication from skeletal muscle to bone: a review for muscle-derived myokines regulating bone metabolism. Calcif Tissue Int. 2017;100(2):184–192. doi:10.1007/s00223-016-0209-4
25. Colaianni G, Mongelli T, Cuscito C, et al. Irisin prevents and restores bone loss and muscle atrophy in hind-limb suspended mice. Sci Rep. 2017;7(1):2811. doi:10.1038/s41598-017-02557-8
26. Anastasilakis AD, Polyzos SA, Makras P, et al. Circulating irisin is associated with osteoporotic fractures in postmenopausal women with low bone mass but is not affected by either teriparatide or denosumab treatment for 3 months. Osteoporos Int. 2014;25(5):1633–1642. doi:10.1007/s00198-014-2673-x
27. Palermo A, Strollo R, Maddaloni E, et al. Irisin is associated with osteoporotic fractures independently of bone mineral density, body composition or daily physical activity. Clin Endocrinol. 2015;82(4):615–619. doi:10.1111/cen.12672
28. Colaianni G, Cuscito C, Mongelli T, et al. The myokine irisin increases cortical bone mass. Proc Natl Acad Sci USA. 2015;112(39):12157–12162. doi:10.1073/pnas.1516622112
29. Ugur K, Aydin S, Kuloglu T, et al. Comparison of irisin hormone expression between thyroid cancer tissues and oncocytic variant cells. Cancer Manag Res. 2019;11:2595–2603. doi:10.2147/CMAR.S201979
30. Moon HS, Mantzoros CS. Regulation of cell proliferation and malignant potential by irisin in endometrial, colon, thyroid and esophageal cancer cell lines. Metabolism. 2014;63(2):188–193. doi:10.1016/j.metabol.2013.10.005
31. Nowinska K, Jablonska K, Pawelczyk K, et al. Expression of Irisin/FNDC5 in cancer cells and stromal fibroblasts of non-small cell lung cancer. Cancers. 2019;11(10):1538. doi:10.3390/cancers11101538
32. Shao L, Li H, Chen J, et al. Irisin suppresses the migration, proliferation, and invasion of lung cancer cells via inhibition of epithelial-to-mesenchymal transition. Biochem Biophys Res Commun. 2017;485(3):598–605. doi:10.1016/j.bbrc.2016.12.084
33. Provatopoulou X, Georgiou GP, Kalogera E, et al. Serum irisin levels are lower in patients with breast cancer: association with disease diagnosis and tumor characteristics. BMC Cancer. 2015;15:898. doi:10.1186/s12885-015-1898-1
34. Zhang ZP, Zhang XF, Li H, et al. Serum irisin associates with breast cancer to spinal metastasis. Medicine. 2018;97(17):e0524. doi:10.1097/MD.0000000000010524
35. Kuloglu T, Celik O, Aydin S, et al. Irisin immunostaining characteristics of breast and ovarian cancer cells. Cell Mol Biol. 2016;62(8):40–44.
36. Gannon NP, Vaughan RA, Garcia-Smith R, Bisoffi M, Trujillo KA. Effects of the exercise-inducible myokine irisin on malignant and non-malignant breast epithelial cell behavior in vitro. Int J Cancer. 2015;136(4):E197–202. doi:10.1002/ijc.29142
37. Aydin S, Kuloglu T, Ozercan MR, et al. Irisin immunohistochemistry in gastrointestinal system cancers. Biotech Histochem. 2016;91(4):242–250. doi:10.3109/10520295.2015.1136988
38. Us Altay D, Keha EE, Ozer Yaman S, et al. Investigation of the expression of irisin and some cachectic factors in mice with experimentally induced gastric cancer. QJM. 2016;109(12):785–790. doi:10.1093/qjmed/hcw074
39. Komolka K, Albrecht E, Schering L, Brenmoehl J, Hoeflich A, Maak S. Locus characterization and gene expression of bovine FNDC5: is the myokine irisin relevant in cattle? PLoS One. 2014;9(1):e88060. doi:10.1371/journal.pone.0088060
40. Gaggini M, Cabiati M, Del Turco S, et al. Increased FNDC5/Irisin expression in human hepatocellular carcinoma. Peptides. 2017;88:62–66. doi:10.1016/j.peptides.2016.12.014
41. Shi G, Tang N, Qiu J, et al. Irisin stimulates cell proliferation and invasion by targeting the PI3K/AKT pathway in human hepatocellular carcinoma. Biochem Biophys Res Commun. 2017;493(1):585–591. doi:10.1016/j.bbrc.2017.08.148
42. Liu J, Song N, Huang Y, Chen Y. Irisin inhibits pancreatic cancer cell growth via the AMPK-mTOR pathway. Sci Rep. 2018;8(1):15247. doi:10.1038/s41598-018-33229-w
43. Liu J, Huang Y, Liu Y, Chen Y. Irisin enhances doxorubicin-induced cell apoptosis in pancreatic cancer by inhibiting the PI3K/AKT/NF-κB pathway. Med Sci Monit. 2019;25:6085–6096. doi:10.12659/MSM.917625
44. Zhang D, Zhang P, Li L, et al. Irisin functions to inhibit malignant growth of human pancreatic cancer cells via downregulation of the PI3K/AKT signaling pathway. Onco Targets Ther. 2019;12:7243–7249. doi:10.2147/OTT.S214260
45. Zhu H, Liu M, Zhang N, et al. Serum and adipose tissue mRNA levels of ATF3 and FNDC5/Irisin in colorectal cancer patients with or without obesity. Front Physiol. 2018;9:1125. doi:10.3389/fphys.2018.01125
46. Kuloglu T, Artas G, Yardim M, et al. Immunostaining characteristics of irisin in benign and malignant renal cancers. Biotech Histochem. 2019;94(6):435–441. doi:10.1080/10520295.2019.1586998
47. Altay DU, Keha EE, Karaguzel E, Mentese A, Yaman SO, Alver A. The diagnostic value of FNDC5/Irisin in renal cell cancer. Int Braz J Urol. 2018;44(4):734–739. doi:10.1590/s1677-5538.ibju.2017.0404
48. Palmberg C, Koivisto P, Visakorpi T, Tammela TL. PSA decline is an independent prognostic marker in hormonally treated prostate cancer. Eur Urol. 1999;36(3):191–196. doi:10.1159/000067996
49. Miyamoto H, Messing EM, Chang C. Androgen deprivation therapy for prostate cancer: current status and future prospects. Prostate. 2004;61(4):332–353. doi:10.1002/pros.20115
50. Tekin S, Erden Y, Sandal S, Yilmaz B. Is irisin an anticarcinogenic peptide. Med-Sci. 2015;4(2):2172–2180. doi:10.5455/medscience.2014.03.8210
51. Kong G, Jiang Y, Sun X, et al. Irisin reverses the IL-6 induced epithelial-mesenchymal transition in osteosarcoma cell migration and invasion through the STAT3/Snail signaling pathway. Oncol Rep. 2017;38(5):2647–2656. doi:10.3892/or.2017.5973
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.