")
Back to Journals » OncoTargets and Therapy » Volume 11
Resveratrol, an activator of SIRT1, induces protective autophagy in non-small-cell lung cancer via inhibiting Akt/mTOR and activating p38-MAPK
Authors Wang J, Li J, Cao N, Li Z, Han J, Li L
Received 6 December 2017
Accepted for publication 30 May 2018
Published 2 November 2018 Volume 2018:11 Pages 7777—7786
DOI https://doi.org/10.2147/OTT.S159095
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jianmin Xu
Jing Wang,1 Jiamei Li,2 Naiqing Cao,1 Zhen Li,3 Jingying Han,3 Li Li3
1Department of Respiration, Shandong Provincial Hospital Affiliated to Shandong University, Jinan, Shandong, China; 2Department of Pathology, Shandong Provincial Hospital Affiliated to Shandong University, Jinan, Shandong, China; 3Department of Laboratory Medicine, Shandong Provincial Hospital Affiliated to Shandong University, Jinan, Shandong, China
Background: Resveratrol, a natural polyphenolic phytoalexin, has potent anti-tumor activity. Recently, it was found to induce autophagy in cancer cells. However, the effects of resveratrol on autophagy in non-small-cell lung cancer (NSCLC) cells have not yet been clearly elucidated.
Materials and methods: A549 and H1299 cells were treated with different concentrations of resveratrol. Cell growth and apoptosis were measured by CCK-8 assay and flow cytometry, respectively. A549 cells were then treated with 200 µM resveratrol or SRT1720. Cell autophagy was detected by western blot and immunofluorescence.
Results: In this study, we found that resveratrol exerted the anti-tumor effect through inhibiting cell proliferation and promoting cell apoptosis in NSCLC cells dose-dependently. Resveratrol has also increased the relative expression of Beclin1 and LC3 II/I while decreased p62 expression, suggesting that resveratrol induced autophagy in NSCLC cells. In addition, resveratrol increased SIRT1 expression and SIRT1 activator SRT1720-induced autophagy of NSCLC cells. SIRT1 knockdown reduced resveratrol-induced autophagy significantly. These results indicated that resveratrol might induce autophagy through upregulating SIRT1 expression. Moreover, inhibiting autophagy by autophagy inhibitor 3-methyladenine or SIRT1 inhibitor nicotinamide significantly suppressed proliferation while promoted apoptosis compared with the resveratrol 200 µM group, suggesting that resveratrol-induced autophagy might act as a protective mechanism to promote NSCLC cell survival and inhibiting autophagy can enhance the anti-tumor effect of resveratrol. Besides that, resveratrol treatment inhibited Akt/mTOR while p38-MAPK was activated in NSCLC cells in a dose-dependent manner. Activating Akt/mTOR pathway by IGF-1 or inhibiting p-38-MAPK pathway by doramapimod significantly inhibited cell proliferation while increased cell apoptosis of NSCLC cells compared with the resveratrol 200 µM group.
Conclusion: Taken together, our findings suggest that resveratrol inhibited proliferation but induced apoptosis and autophagy via inhibiting Akt/mTOR and activating p38-MAPK pathway. Resveratrol-induced autophagy might act as a protective mechanism to promote NSCLC cell survival. Therefore, inhibition of autophagy may enhance the anti-tumor activity of resveratrol in NSCLC.
Keywords: resveratrol, SIRT1, autophagy, non-small-cell lung cancer
Introduction
Non-small-cell lung cancer (NSCLC), which includes adenocarcinoma, squamous cell carcinoma, large cell carcinoma, and several other types, is a significant global health problem presently.1 As one of the most common malignancies, NSCLC remains the leading cause of cancer-related death worldwide.2 Although great improvements have been achieved in early detection and the treatments for NSCLC, the prognosis for NSCLC is still poor, with an estimated survival rate of only 15% at 5 years.3 Therefore, searching for new and effective treatment is an urgent need for NSCLC patients.
Resveratrol (trans-3,4′,5-trihydroxystilbene) is a natural polyphenolic phytoalexin, which is found in red grape skins, red wine, and peanuts.4 Accumulating evidence indicated that resveratrol exerted various biological effects including anti-oxidation, inhibition of tumorigenesis, and inhibition of angiogenesis.5,6 It was reported that the effects of resveratrol appeared to be related to its ability to induce silent information regulator (Sir2α, also known as SIRT1) activity.7 SIRT1 is a member of the class III histone deacetylase (HDAC) family and is a redox-sensitive enzyme that needs cellular NAD as a cofactor for its deacetylation reactivity.8 Previous studies elucidated that SIRT1 exerts its tumor suppressive activity through suppressing proliferation, inflammation, and angiogenesis by inducing apoptosis and autophagy.9–11 However, studies on whether resveratrol could activate SIRT1 and exert anti-tumor effects in NSCLC are still few and need further investigations.
Autophagy is a cellular process in which intracellular contents including large protein complexes and dysfunctional organelles are transported to lysosomes for degradation and reuse.12 Through degrading and recycling unnecessary or dysfunctional cellular components, autophagy maintains intracellular homeostasis and prevents cellular damage under multiple stresses.13 Autophagy is reported to act as a double-edged sword in cancer survival.14 On the one hand, autophagy supported cancer cell survival through recycling cellular components and promoting energy production to meet the high metabolic demands of cancer cells. On the other hand, autophagy reduces cell instability and damage to prevent tumorigenesis.15
In this study, we explored the autophagy induction effect of resveratrol on NSCLC cells and examined the underlying molecular mechanisms. Our findings indicated that resveratrol activated SIRT1 to induce protective autophagy in NSCLC cells via inhibiting Akt/mTOR and activating p38-MAPK pathway. Therefore, inhibition of protective autophagy may enhance anti-tumor activity of resveratrol in NSCLC.
Materials and methods
Cell culture
NSCLC cell lines A549 and H1299 cells were purchased from American Type Culture Collection (Manassas, VA, USA). Cells were cultured in RPMI-1640 complete culture medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS, HyClone; GE Healthcare Life Science, Logan, UT, USA) in a humidified atmosphere of 5% CO2 at 37°C.
Reagents and antibodies
Resveratrol, 3-methyladenine (3-MA), and nicotinamide were obtained from Sigma-Aldrich Co. (St Louis, MO, USA) and dissolved in dimethyl sulfoxide. SRT1720 was obtained from Calbiochem-Novabiochem Co. (La Jolla, CA, USA). Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis detection kit was purchased from Multisciences (Shanghai, China). Insulin-like growth factor-1 (IGF-1) was purchased from PeproTech, Inc. (Rocky Hill, NJ, USA). Doramapimod was obtained from Medichem Express (Princeton, NJ, USA). Antibodies against Beclin1 (No 3738), LC3 I/II (No 12741), p62 (No 88588), SIRT1 (No 8469), p-Akt (No 4060), Akt (No 4685), p-mTOR (No 5536), mTOR (No 2983), p-p70S6K (No 9204), p70S6K (No 2708), p-p38 (No 4511), p-38 (No 8690), and GAPDH (No 5174) as well as horseradish peroxidase (HRP)-conjugated secondary anti-rabbit and anti-mouse antibodies were obtained from Cell Signaling Technology (Boston, MA, USA).
Cell proliferation assay
Cell proliferation was determined by cell counting kit-8 (CCK-8) assay. Cells were seeded in 96-well plates at 5×103 per well and incubated with different concentrations of resveratrol (25, 100, and 200 μM) for indicated time spans. Then, 10 μL of CCK-8 solution (Beyotime Institute of Biotechnology, Shanghai, China) was added to the cultures and incubated at 37°C for 2 hours. The absorbance was measured at 450 nm using a microplate spectrophotometer (Molecular Devices LLC, Sunnyvale, CA, USA). Triplicate wells were used in each group.
Cell apoptosis assay
Cells were washed twice with cold PBS and resuspended at 1×106 cells/mL in 400 μL binding buffer. After adding 5 μL of Annexin V-FITC, the cells were incubated for 15 minutes at 4°C in the dark. Then, 10 μL of PI was added, and the cells were incubated for another 5 minutes at 4°C in the dark. Cell apoptosis was analyzed in a flow cytometer (BD Biosciences, San Jose, CA, USA).
Western blot assay
A549 cells were lysed with RIPA lysis buffer (Beyotime Institute of Biotechnology) according to the manufacturer’s protocol. Same amount of proteins of each sample was separated by 10% SDS-PAGE gel and then transferred into polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA, USA). After blocking with 5% bovine serum albumin, the membrane was incubated with specific primary antibodies overnight at 4°C followed by incubation with HRP-conjugated secondary antibodies for 1 hour at room temperature, and signals were detected by electrochemical luminescence.
Fluorescence microscopy
A549 cells were transfected with GFP-LC3 plasmid by Lipofectamine 3000™ (Thermo Fisher Scientific). After A549 cells were treated with or without resveratrol or SRT1720, the number of puncta formation of GFP-LC3 was determined under fluorescent microscopy. Cells with >5 puncta were considered to have accumulated autophagosomes.
Cell transfection
A549 cells were seeded into 96-well plates at 5×103 cells per well and cultured in complete medium for transfection. Small interfering RNA against SIRT1 (SIRT1 siRNA) was synthesized by GenePharma (Shanghai, China) and was transfected into A549 cells using Lipofectamine 2000 reagent (Thermo Fisher Scientific) according to manufacturer’s protocol.
Statistical analysis
All data are represented as mean ± standard deviation (SD). Statistical software SPSS 19.0 was used for the assessment. Student’s t-test was used to compare the mean values of two groups, and one-way analysis of variance was used for comparing the mean values values of multiple samples. Values of P<0.05 were considered statistically significant.
Results
Resveratrol inhibits proliferation while induces apoptosis of NSCLC cells
In order to evaluate the potential therapeutic effect of resveratrol for NSCLC, NSCLC cell lines A549 and H1299 cells were used in our experiments. The CCK-8 assay was performed to detect cell proliferation of NSCLC cells treated with different concentrations of resveratrol. As shown in Figure 1A and B, resveratrol treatment inhibited cell proliferation of A549 and H1299 cells in a dose-dependent manner compared with the control group (*P<0.05, **P<0.01). To define whether resveratrol-induced cell death is associated with apoptosis, Annexin V/PI double staining and flow cytometry were used to detect apoptosis. As shown in Figure 1C and D, the resveratrol treatment promoted cell apoptosis of both A549 and H1299 cells dose-dependently compared with the control group (*P<0.05, **P<0.01). These results indicated that resveratrol inhibited cell proliferation while induced apoptosis in NSCLC cells. The A549 cells were chosen for our following experiments.
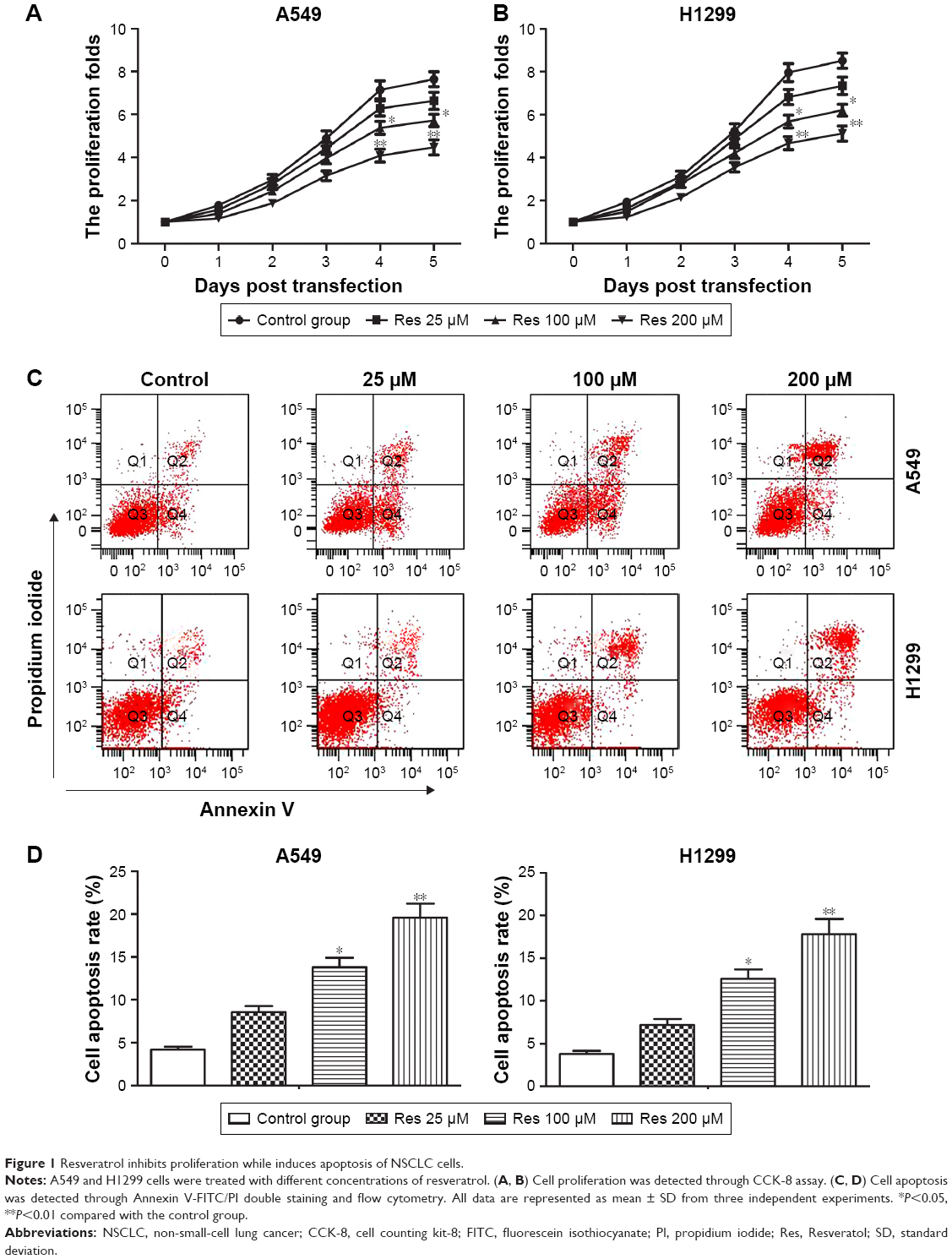 | Figure 1 Resveratrol inhibits proliferation while induces apoptosis of NSCLC cells. |
Resveratrol induces autophagy through increasing SIRT1 expression in NSCLC cells
We also observed that resveratrol treatment dose-dependently increased relative protein level of Beclin1 and LC3 II/I which were two well-validated biomarkers of autophagy while decreased relative protein level of p62 which can clear autophagosome, suggesting that resveratrol treatment induced autophagy in A549 cells (Figure 2A and B, *P<0.05, **P<0.01). We further examined the molecular mechanism underlying the resveratrol-induced autophagy. Our results from Western blot showed that resveratrol treatment upregulated the expression of SIRT1 significantly. We then examined the localization of LC3 to autophagosome formation as assessed through GFP-LC3 expression. LC3+ puncta formation observed as punctate dots of green fluorescence was significantly increased in the resveratrol 200 μM group compared with the control group. Therefore, we hypothesized that resveratrol might induce autophagy through upregulating SIRT1 expression. To verify our hypothesis, an SIRT1 activator SRT1720 (3 μM) was used to treat A549 cells for the induction of SIRT1 expression (Figure 2C and D, **P<0.01). SRT1720 treatment increased LC3+ puncta in cells remarkably compared with the control group (Figure 2E and F, *P<0.05, **P<0.01). Then, we transfected A549 cells with SIRT1 siRNA to decrease the expression of SIRT1 (Figure 2G and H, *P<0.05, #P<0.05). We observed that SIRT1 siRNA could counteract the increased Beclin1 expression induced by resveratrol significantly (Figure 2I and J, *P<0.05, #P<0.05). The abovementioned results verified our hypothesis that resveratrol induced autophagy by increasing SIRT1 expression in NSCLC cells.
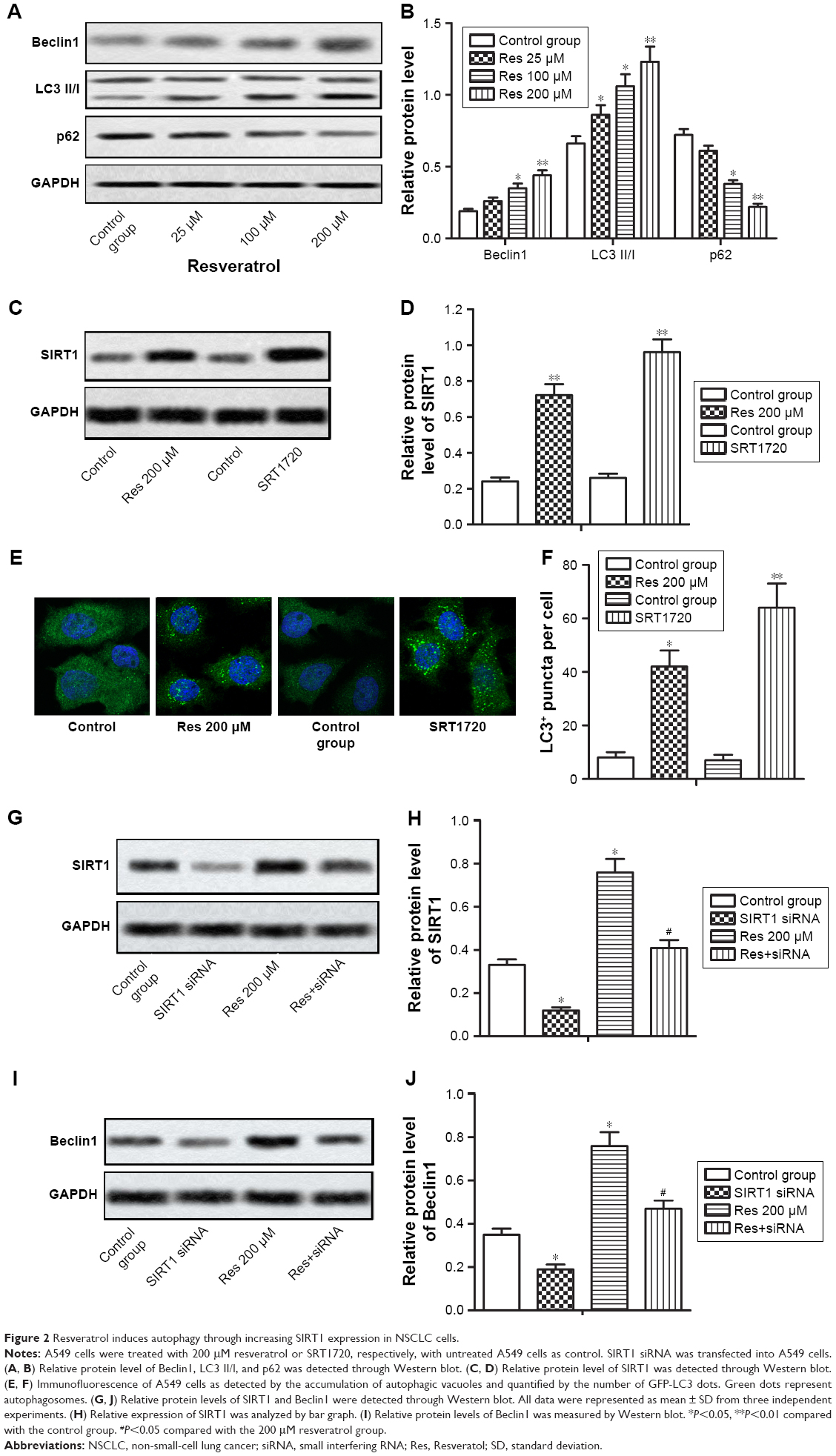 | Figure 2 Resveratrol induces autophagy through increasing SIRT1 expression in NSCLC cells. |
Treatment with autophagy inhibitor 3-MA or SIRT1 inhibitor nicotinamide enhances the anti-tumor effect of resveratrol
Since autophagy has dual roles in cancer cell survival and cell death, we then explored the effect of autophagy on the anti-tumor effect of resveratrol in A549 cells. We inhibited resveratrol-induced autophagy through treating A549 cell with autophagy inhibitor 3-MA (10 mmol/L) or SIRT1 inhibitor nicotinamide (5 mmol/L). Cell proliferation ability was determined by CCK-8 assay. The data showed that 3-MA or nicotinamide co-treatment both inhibited cell proliferation and promoted apoptosis compared with the resveratrol 200 μM group (Figure 3A and B, **P<0.01, #P<0.05, ##P<0.01). Our results demonstrated that inhibition of resveratrol-induced protective autophagy enhanced the anti-tumor effect of resveratrol in NSCLC cells.
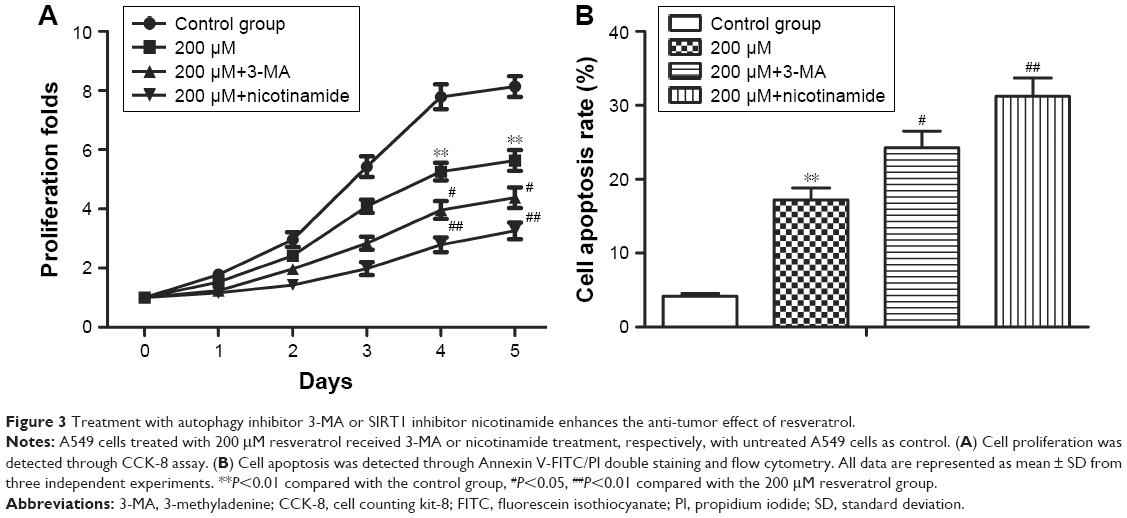 | Figure 3 Treatment with autophagy inhibitor 3-MA or SIRT1 inhibitor nicotinamide enhances the anti-tumor effect of resveratrol. |
Resveratrol induces autophagy via inhibiting Akt/mTOR and activating p38-MAPK pathway
Previous study indicated that resveratrol induced apoptosis and autophagy in T-cell acute lymphoblastic leukemia cells by inhibiting Akt/mTOR and activating p38-MAPK.16 To investigate whether these signaling pathways were also involved in resveratrol-induced apoptosis and autophagy in NSCLC cells, Western blot was performed to detect the activated state of associated proteins in A549 cells treated with different concentrations of resveratrol. Our results showed that resveratrol inhibited the phosphorylation of Akt, mTOR, and p-70S6K while increased the phosphorylation of p-38, thus resulted in the decreased ratio of p-Akt/Akt, p-mTOR/mTOR, p-p-70S6K/p-70S6K and increased ratio of p-p-38/p-38 in a dose-dependent manner (Figure 4A–E, *P<0.05, **P<0.01). As an Akt/mTOR activator, IGF-1 increased the phosphorylation of Akt and doramapimod decreased the phosphorylation of p-38 as a p-38-MAPK pathway inhibitor (Figure 4F–I, *P<0.05, **P<0.01, #P<0.05, ##P<0.01). Activating Akt/mTOR pathway by IGF-1 (50 ng/mL) or inhibiting p-38-MAPK pathway by doramapimod (10 μM) significantly inhibited cell proliferation while increased cell apoptosis of NSCLC cells compared with the resveratrol 200 μM group (Figure 4J and K, **P<0.01, #P<0.05). These results suggested that resveratrol induced protective autophagy via inhibiting Akt/mOR and activating p-38-MAPK signaling pathway. Therefore, activating Akt/mTOR or inhibiting p-38-MAPK signaling pathway can enhance the anti-tumor effect of resveratrol in NSCLC cells.
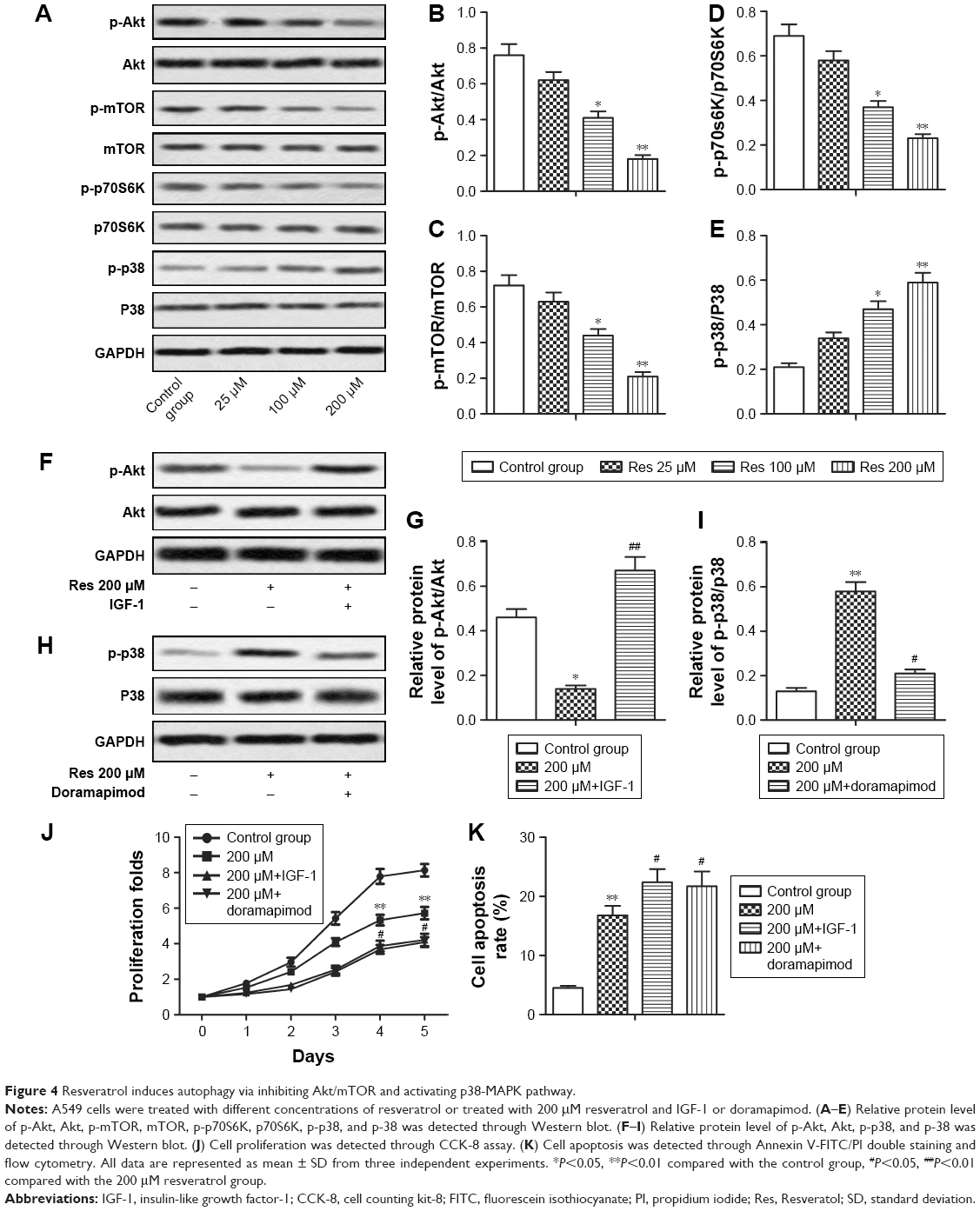 | Figure 4 Resveratrol induces autophagy via inhibiting Akt/mTOR and activating p38-MAPK pathway. |
Discussion
Resveratrol, a naturally occurring antibiotic plant, has been shown to have potent anti-tumor effect through suppressing proliferation and inducing apoptosis in multiple cancers.17,18 In this study, we also found that resveratrol, as an SIRT1 activator, can also induce protective autophagy which may inhibit the anti-tumor effect of resveratrol via inhibiting Akt/mTOR and activating p38-MAPK. Therefore, suppressing protective autophagy enhanced anti-tumor effect of resveratrol, making it an effective reagent against NSCLC.
Recently, natural compounds such as resveratrol have gained much attention for preventive and therapeutic strategies in cancer treatment including lung cancer.19,37 Previous studies have elucidated that resveratrol inhibited proliferation and induced apoptosis in various types of cancer cells. For instance, Lei et al reported that resveratrol inhibited proliferation and promoted differentiation and melanogenesis in HT-144 melanoma cells through inhibition of MEK/ERK kinase pathway.20 Research by Kim et al showed that resveratrol induced mitochondrial apoptosis and inhibited cell invasion and migration by inhibiting epithelial–mesenchymal transition in oral squamous cell carcinoma cells.21 Resveratrol was also reported to decrease cell viability and proliferation while induced apoptosis in H460 lung cancer cells significantly.22 Similarly, we also observed that resveratrol suppressed cell proliferation and induced apoptosis in NSCLC cells, indicating the anti-tumor effect of resveratrol in NSCLC progression.
Autophagy is a cellular process that long-lived proteins and damaged organelles are transported to lysosomes for degradation and reuse.12 Besides inducing apoptosis, resveratrol can also induce autophagy in cancer cells.16,23 Beclin1 is a specific marker of autophagy and is strongly involved in the autophagic process, especially in its early stages.24 The conversion of LC3I to LC3II contributes to the formation of autophagosome, and LC3II is often considered as an autophagosomal marker.25 P62 protein can clear the autophagosome in order to keep hemostasis.26 In this study, resveratrol treatment increased the expression of Beclin1 and LC3 II/I while decreased the expression of p62, indicating that resveratrol induced autophagy in NSCLC cells dose-dependently.
Resveratrol is reported to induce autophagy through activating SIRT1-mediated pathway in multiple types of cells.27–29 SIRT1 is an NAD+-dependent HDAC which is involved in multiple disease processes including cancer, vascular disease, and neurodegenerative disorders.30–32 SIRT1 can induce autophagy through deacetylation of Beclin1 and other autophagy mediators.33 Similarly, in this study, we observed that resveratrol treatment upregulated the expression of SIRT1 and induced autophagy significantly, indicating that resveratrol induced autophagy through activating SIRT1 in NSCLC cells. Adding SIRT1 activator SRT1720 further verified our hypothesis that increased expression of SIRT1 induced autophagy remarkably. In addition, knockdown of SIRT1 reduced resveratrol-induced autophagy significantly. These results elucidated that resveratrol acted as an SIRT1 activator to induce autophagy in NSCLC cells.
Autophagy can promote or suppress tumor cell survival. Autophagy enables cancer cells to survive under hostile stress while continuous stress and excessive autophagy could lead to autophagic cell death.34 A great number of research studies have demonstrated that autophagy played a cytoprotective role in cancer cells.15,35,36 Autophagy and apoptosis have a complex mutual relationship. In some cases, autophagy may act as an initiating factor for apoptosis-induction while in other cases autophagy can antagonize apoptosis. Here, we observed that inhibiting autophagy by autophagy inhibitor 3-MA or SIRT1 inhibitor nicotinamide significantly suppressed proliferation while promoted apoptosis compared with the resveratrol 200 μM group. These results elucidated that resveratrol-induced autophagy might act as a protective mechanism to promote NSCLC survival and inhibiting autophagy can enhance the anti-tumor effect of resveratrol.
Pathways of Akt/mTOR and p38-MAPK are also linked to resveratrol-induced apoptosis and autophagy. When nutrients are sufficient, the PI3K/Akt pathway activates mTOR to inhibit autophagy. During nutrient starvation and stress, the activity of mTOR is inhibited, leading to autophagy induction. Autophagy is one of the main mechanisms regulated by AMPK.15 Ge et al reported that resveratrol induced apoptosis and autophagy via inhibiting Akt/mTOR and activating p38-MAPK in T-cell acute lymphoblastic leukemia cells.16 Our present data also went in line with this study that resveratrol treatment inhibited Akt/mTOR while activated p38-MAPK in NSCLC cells in a dose-dependent manner. Moreover, activating Akt/mTOR pathway by IGF-1 or inhibiting p-38-MAPK pathway by doramapimod significantly inhibited cell proliferation but increased cell apoptosis of NSCLC cells compared with the resveratrol 200 μM group. Our results indicated that activating Akt/mTOR or inhibiting p-38-MAPK signaling pathway can enhance the anti-tumor effect of resveratrol in NSCLC cells.
Taken together, this study provides evidence that resveratrol exerted anti-tumor effect through inhibiting cell proliferation and enhancing cell apoptosis of NSCLC cells. However, resveratrol, acted as a SIRT1 activator, also induced protective autophagy to promote NSCLC survival via inhibiting Akt/mTOR and activating p38-MAPK. Therefore, activating Akt/mTOR or inhibiting p38-MAPK can suppress resveratrol-induced protective autophagy, which may enhance anti-tumor effects of resveratrol in NSCLC.
Acknowledgment
The present study was supported by Natural Science Foundation of Shandong Province (No ZR2015PH016) and Clinical Science and Technology Innovation Project (No 201503004).
Disclosure
The authors report no conflicts of interest in this work.
References
Siegel R, Naishadham D, Jemal A. Cancer statistics for Hispanics/Latinos, 2012. CA Cancer J Clin. 2012;62(5):283–298. | ||
Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108. | ||
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2017. CA Cancer J Clin. 2017;67(1):7–30. | ||
Jang M, Cai L, Udeani GO, et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science. 1997;275(5297):218–220. | ||
Tyihak E, Kiraly-Veghely Z, Moricz AM. Multiple beneficial effects of resveratrol and their chemical-biochemical basis. Nat Prod Commun. 2011;6(5):631–638. | ||
Tang FY, Su YC, Chen NC, Hsieh HS, Chen KS. Resveratrol inhibits migration and invasion of human breast-cancer cells. Mol Nutr Food Res. 2008;52(6):683–691. | ||
Sulaiman M, Matta MJ, Sunderesan NR, Gupta MP, Periasamy M, Gupta M. Resveratrol, an activator of SIRT1, upregulates sarcoplasmic calcium ATPase and improves cardiac function in diabetic cardiomyopathy. Am J Physiol Heart Circ Physiol. 2010;298(3):H833–H843. | ||
Gu XS, Wang ZB, Ye Z, et al. Resveratrol, an activator of SIRT1, upregulates AMPK and improves cardiac function in heart failure. Genet Mol Res. 2014;13(1):323–335. | ||
Kume S, Haneda M, Kanasaki K, et al. Silent information regulator 2 (SIRT1) attenuates oxidative stress-induced mesangial cell apoptosis via p53 deacetylation. Free Radic Biol Med. 2006;40(12):2175–2182. | ||
Kabra N, Li Z, Chen L, et al. SirT1 is an inhibitor of proliferation and tumor formation in colon cancer. J Biol Chem. 2009;284(27):18210–18217. | ||
Potente M, Ghaeni L, Baldessari D, et al. SIRT1 controls endothelial angiogenic functions during vascular growth. Genes Dev. 2007;21(20):2644–2658. | ||
Mizushima N. Autophagy: process and function. Genes Dev. 2007;21(22):2861–2873. | ||
Kroemer G, Marino G, Levine B. Autophagy and the integrated stress response. Mol Cell. 2010;40(2):280–293. | ||
Mathew R, Karantza-Wadsworth V, White E. Role of autophagy in cancer. Nat Rev Cancer. 2007;7(12):961–967. | ||
Zhao F, Huang W, Zhang Z, et al. Triptolide induces protective autophagy through activation of the CaMKKbeta-AMPK signaling pathway in prostate cancer cells. Oncotarget. 2016;7(5):5366–5382. | ||
Ge J, Liu Y, Li Q, et al. Resveratrol induces apoptosis and autophagy in T-cell acute lymphoblastic leukemia cells by inhibiting Akt/mTOR and activating p38-MAPK. Biomed Environ Sci. 2013;26(11):902–911. | ||
Empl MT, Cai H, Wang S, et al. Effects of a grapevine shoot extract containing resveratrol and resveratrol oligomers on intestinal adenoma development in mice: in vitro and in vivo studies. Mol Nutr Food Res. In press 2018. | ||
Chai R, Fu H, Zheng Z, Liu T, Ji S, Li G. Resveratrol inhibits proliferation and migration through SIRT1 mediated posttranslational modification of PI3K/AKT signaling in hepatocellular carcinoma cells. Mol Med Rep. 2017;16(6):8037–8044. | ||
Yousef M, Vlachogiannis IA, Tsiani E. Effects of resveratrol against lung cancer: in vitro and in vivo studies. Nutrients. 2017;9(11):E1231. | ||
Lei MJ, Dong Y, Sun CX, Zhang XH. Resveratrol inhibits proliferation, promotes differentiation and melanogenesis in HT-144 melanoma cells through inhibition of MEK/ERK kinase pathway. Microb Pathog. 2017;111:410–413. | ||
Kim SE, Shin SH, Lee JY, et al. Resveratrol induces mitochondrial apoptosis and inhibits epithelial-mesenchymal transition in oral squamous cell carcinoma cells. Nutr Cancer. 2018;70(1):125–135. | ||
Wright C, Iyer AKV, Yakisich JS, Azad N. Anti-tumorigenic effects of resveratrol in lung cancer cells through modulation of c-FLIP. Curr Cancer Drug Targets. 2017;17(7):669–680. | ||
Miki H, Uehara N, Kimura A, et al. Resveratrol induces apoptosis via ROS-triggered autophagy in human colon cancer cells. Int J Oncol. 2012;40(4):1020–1028. | ||
Sinha S, Levine B. The autophagy effector Beclin 1: a novel BH3-only protein. Oncogene. 2008;27(Suppl 1):S137–S148. | ||
Tanida I, Waguri S. Measurement of autophagy in cells and tissues. Methods Mol Biol. 2010;648:193–214. | ||
Wang S, Wang C, Yan F, et al. N-Acetylcysteine attenuates diabetic myocardial ischemia reperfusion injury through inhibiting excessive autophagy. Mediators Inflamm. 2017;2017:9257291. | ||
Zhao H, Chen S, Gao K, et al. Resveratrol protects against spinal cord injury by activating autophagy and inhibiting apoptosis mediated by the SIRT1/AMPK signaling pathway. Neuroscience. 2017;348:241–251. | ||
Zhang Y, Cao X, Zhu W, et al. Resveratrol enhances autophagic flux and promotes Ox-LDL degradation in HUVECs via upregulation of SIRT1. Oxid Med Cell Longev. 2016;2016:7589813. | ||
Deng H, Mi MT. Resveratrol attenuates Abeta25-35 caused neurotoxicity by inducing autophagy through the TyrRS-PARP1-SIRT1 signaling pathway. Neurochem Res. 2016;41(9):2367–2379. | ||
Maiese K. SIRT1 and stem cells: In the forefront with cardiovascular disease, neurodegeneration and cancer. World J Stem Cells. 2015;7(2):235–242. | ||
D’Onofrio N, Vitiello M, Casale R, Servillo L, Giovane A, Balestrieri ML. Sirtuins in vascular diseases: Emerging roles and therapeutic potential. Biochim Biophys Acta. 2015;1852(7):1311–1322. | ||
Duan W. Sirtuins: from metabolic regulation to brain aging. Front Aging Neurosci. 2013;5:36. | ||
Qiu G, Li X. The prognostic role of SIRT1-autophagy axis in gastric cancer. Dis Mark. 2016;2016:6869415. | ||
Song L, Wang Z, Wang Y, et al. Natural cyclopeptide RA-XII, a new autophagy inhibitor, suppresses protective autophagy for enhancing apoptosis through AMPK/mTOR/P70S6K pathways in HepG2 cells. Molecules. 2017;22(11):E1934. | ||
White E, DiPaola RS. The double-edged sword of autophagy modulation in cancer. Clin Cancer Res. 2009;15(17):5308–5316. | ||
Marino G, Niso-Santano M, Baehrecke EH, Kroemer G. Self-consumption: the interplay of autophagy and apoptosis. Nat Rev Mol Cell Biol. 2014;15(2):81–94. | ||
Rasheduzzaman M, Jeong JK, Park SY. Resveratrol sensitizes lung cancer cell to TRAIL by p53 independent and suppression of Akt/NF-kappaB signaling. Life sci. 2018;208:208–220. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.