")
Back to Journals » Infection and Drug Resistance » Volume 16
Resistance and Molecular Characteristics of Methicillin-Resistant Staphylococcus aureus and Heterogeneous Vancomycin-Intermediate Staphylococcus aureus
Authors Liang J, Hu Y, Fu M, Li N, Wang F, Yu X, Ji B
Received 10 October 2022
Accepted for publication 10 January 2023
Published 23 January 2023 Volume 2023:16 Pages 379—388
DOI https://doi.org/10.2147/IDR.S392908
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Jin Liang,1,* Yuanfang Hu,1,* Mingxia Fu,2 Na Li,1 Fengxia Wang,1 Xiaojun Yu,1 Bing Ji1
1Department of Laboratory Medicine, Binzhou Medical University Hospital, Binzhou, People’s Republic of China; 2Department of Laboratory Medicine, Binzhou People’s Hospital, Binzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Bing Ji, Email [email protected]
Objective: To investigate the molecular epidemiology of Staphylococcus aureus (S. aureus) isolated from patients in the Yellow River Delta region and to construct antibiotic resistance profiles in different genetic backgrounds.
Methods: Antibiotics susceptibility testing, staphylococcal protein A (spa) typing and accessory gene regulator (agr) typing were performed for all the 204 strains. Isolates with the positive mecA gene and heterogeneous vancomycin-intermediate S. aureus (hVISA) were subjected to multilocus sequence typing (MLST) and Staphylococcal chromosomal cassettes mec (SCCmec) typing.
Results: Thirty-nine MRSA strains were identified by mecA gene. Twenty-two hVISA isolates including 9 MRSA and 13 methicillin-sensitive S. aureus (MSSA) strains were confirmed, the rest isolates (n = 182) were vancomycin-susceptible S. aureus (VSSA). 94.9% of MRSA and 63.6% of hVISA belonged to agrI. Fifty-seven distinct spa types including 5 novel types were mainly t309 (30.9%), t078 (11.8%) and t437 (11.8%). Fourteen sequence types (STs) containing 3 new STs were classified into 3 clone complexes (CCs) and 7 singletons among MRSA and hVISA isolates. Most MRSA isolates (87.2%) belonged to type Ⅳ SCCmec.
Conclusion: The predominant genotype among MRSA population was ST59-t437-agrI-IVa (53.8%), followed by ST72-t2431-agrI-IVF (15.4%). ST72 and CC5 (ST5/965/7197) were the most common hVISA clones. Both CC59 (ST59/7437) and ST72 clones were resistant to erythromycin and clindamycin among MRSA population. Strains of MSSA with phenotypic hVISA (MS-hVISA) exhibited a striking genetic diversity accompanied by the diversification of drug resistance patterns.
Keywords: Staphylococcus aureus, heterogeneous vancomycin-intermediateS. aureus, accessory gene regulator, multilocus sequence typing, erythromycin
Introduction
For more than 140 years since its discovery, Staphylococcus aureus has remained a pathogen of clinical concern. Among the various models of S. aureus, the real challenge is the emergence of MRSA.1 Over a long period of evolution, MRSA acquired the mecA gene encoding a penicillin-binding protein (PBP2a) that displays low affinity for most semisynthetic penicillins by horizontal transfer, resulting in resistance to β-lactams.2 Due to the misuse of antibiotics in humans, the “cunning” MRSA has integrated antibiotic resistance genes into its genome in various ways, leading to the emergence of multidrug-resistant strains, making it more difficult to treat, and as a result, the spread of MRSA has become more unchecked worldwide. To understand the molecular epidemiology from different geographical distributions, researchers adopt typing methods including SCCmec, spa, agr and MLST as a means for the characterization and discrimination of isolates based on their genetic characteristics. However, the antibiotics susceptibility results of strains from the same background differed. ST22 lineage from a Kuwaiti hospital showed a high resistance rate of 69.2% and 84.6% to gentamicin and trimethoprim-sulfamethoxazole, respectively,3 in contrast, the clone from China displayed no resistance activity to both antibiotics.4 Strains ST8-IVa-t008 from China and Germany were both susceptible to clindamycin but differed in resistance to ciprofloxacin (100%, 37.5%).5,6 ST59-IV-t437 from England was resistant to erythromycin, clindamycin and tetracycline, however, this lineage from the Chinese pediatric population was phenotypically resistant to trimethoprim-sulfamethoxazole, ciprofloxacin and chloramphenicol in addition to the above antibiotics.7,8
Gram-positive bacterial infections including MRSA are majorly treated using glycopeptide antibiotics, in which vancomycin is the first-line agent. Vancomycin binds the D-alanine-D-alanine terminus of the murein precursor, forming a complex that in turn blocks transpeptidation as well as transglycosylation, leading to incomplete or inefficient peptidoglycan cross-linking during cell wall synthesis. The resultant cell wall is weak and unable to adapt to changing osmotic pressures.9 However, due to irrational use of vancomycin, hVISA/VISA strains which causes vancomycin treatment failure and prolonged hospitalization have emerged. The data suggested that the prevalence of hVISA/VISA in China (7.98%) is higher than the global average (3.39–3.68%), and causes the risk of persistent infection and higher treatment costs.10–12 The aim of this study was to determine the molecular epidemiology of MRSA and hVISA/VISA strains in patients from the Yellow River Delta Regional Center (a tertiary care hospital) and to construct antibiogram under different molecular characteristics, thus providing a theoretical basis for clinicians to execute empirical drug administration well.
Materials and Methods
Collection of Bacterial Isolates
A total of 204 S. aureus strains were obtained from inpatients and outpatients from September 2019 to February 2021 which involved 25 sections as part of the routine laboratory procedures at the Binzhou Medical University Hospital. These strains were isolated from pus (n = 78, 38.2%), sputum (n = 40, 19.6%), secretions (n = 29, 14.2%), transudation (n = 24, 11.8%), blood (n = 17, 8.3%) and other seven sources (n < 10, <4.9% each). Strains were initially identified using matrix-assisted laser desorption ionization-time of flight mass spectrometry MALDI-TOF (Biomérieux, France) and stored at −70°C.
Antimicrobial Susceptibility Testing
All S. aureus isolates were subjected to antibiotic susceptibility testing using the automated VITEK 2 Compact system (Biomérieux, France). The antibiotics agents used were penicillin (PEN), oxacillin (OXA), cefoxitin (FOX), ceftarolin (CPT), gentamicin (GEN), levofloxacin (LVX), moxifloxacin (MFX), erythromycin (ERY), clindamycin (CLI), rifampin (RIF), trimethoprim-sulfamethoxazole (SXT), vancomycin (VAN), teicoplanin (TEC), daptomycin (DAP), and linezolid (LZD). In addition, we used the rapid E-test method to determine the MICs for 3 agents. Briefly, 50 µL of bacterial suspensions with turbidity adjusted to 0.5 McFarland’s standard were evenly spread on BHI agar plates. E-test strips for amikacin (AMK), chloramphenicol (CHL) and minocycline (MNO) were placed on the plate. Susceptible, intermediate and resistant breakpoints for these antibiotics agents were determined using the Clinical and Laboratory Standards Institute (CLSI) interpretation criteria.13 Strains that are resistant to 3 kinds or more of antibiotics are defined as MDR.
Identification of MRSA and hVISA Isolates
The criteria of resistance towards oxacillin (MIC ≥ 2 μg/mL) and/or cefoxitin (MIC ≥ 4 μg/mL) of phenotypic MRSA in S. aureus isolates was performed according to CLSI guidelines.13 mecA was screened by PCR as a confirmatory experiment for MRSA. The primers were mecA147-F 5’ -GTGAAGATATACCAAGTGATT-3’ and mecA147-R 5’ -ATGCGCTATAGATTGAAAGGAT-3’. Since there are no clear guidelines for identification of hVISA in CLSI, we performed “gold standard” test population analyses profile– area under curve (PAP/AUC) after positive primary screening to reduce the workload.14,15 Briefly, randomly isolated colonies were diluted using 0.85% sodium chloride solution, and their density adjusted to be equivalent to that of 0.5 McFarland’s standard (1.5×108 CFU/mL). Then, 10 uL of suspensions were spot inoculated onto Brain-Heart infusion agar (BHIA) plates containing casein digested by trypsin (16 g/L) and vancomycin (2, 3, 4 mg/L). The BHIA plates were incubated at 35°C for 48 h. If the number of colonies ≥1 within 48 h, the parental strain was designated as suspected hVISA. Positive primary parent bacterial cultures from overnight growth in 8 mL tryptone soy broth (TSB) liquid medium were suspended in 0.85% sterile saline to a density corresponding to 0.5 McFarland’s standard. Then, the original suspensions were diluted to 10−3 (105 CFU/mL) and 10−6 (102 CFU/mL). Fifty uL aliquot of start cell suspension of each isolate and its serial dilution was spread on BHI agar plate containing increasing concentrations of vancomycin (0, 0.5, 1, 2, 3, 4, 6, 8, 12 mg/L). Plates were incubated at 35°C for 48 h. The hVISA reference strain (Mu3) was used as the positive control while the ATCC29213 strain was the negative control. The plate with the best growth of a single colony in three different concentrations was selected for manual counting, and the actual number of CFU determined as the dilution multiplied by the number of colonies in the plate under that dilution and expressed in log10 CFU/mL. The Graphpad Prism software was used to draw the curve of cell number versus vancomycin concentration and to calculate the area under the curve (AUC). The PAP/AUC ratio was calculated as AUCstrain/AUCMu3. The accepted criteria for confirmation was AUCstrain/AUCMu3 <0.9 for VSSA, >1.3 for VRSA, and a ratio of 0.9 to 1.3 was considered as hVISA.
Accessory Gene Regulator (Agr) and Staphylococcal Protein a (Spa) Typing
All isolates were characterized by the combination of spa typing and agr locus analysis as previously described.16,17
Multi Locus Sequence Typing (MLST) Determination
The MLST scheme was followed using seven housekeeping genes (arcC, aroE, glpF, gmk, pta, tpi, and yqiL) on MRSA and hVISA isolates.18 Allelic profiles containing 7 allele numbers for each strain were assigned a sequence type (ST) by the MLST database (https://pubmlst.org/saureus/). Phylogenetic tree construction was performed by the Neighbor-Joining method using mega7 software (https://www.megasoftware.net/) to cluster STs with close genetic backgrounds into identical clonal complexes (CCs).
Identification of the SCCmec
Multiplex PCR was performed to determine the SCCmec type and subtype as previously described.19
Statistical Analysis
The IBM SPSS version 22.0 software was used for analyses. The Pearson chi-square test or Fisher’s exact test was used for categorical variables. P < 0.05 was the threshold for significance.
Results
Clinical Characteristics of S. aureus
Thirty-nine MRSA strains harboured the mecA gene based on PCR analysis. Fifty phenotypic methicillin-resistance strains were obtained, among which 49 resistance to cefoxitin, 43 resistance to oxacillin and 42 resistance to both agents. Among two mecA-positive strains, one was susceptible to cefoxitin and oxacillin and another was susceptible to oxacillin. Of the 12 strains absent methicillin-resistance determinant, mecA, six were resistant to cefoxitin, one to oxacillin, and the remaining five were tolerant to both.
Twenty-two isolates (9 MRSA and 13 MSSA exhibiting hVISA phenotypes, ie MR-hVISA and MS-hVISA) were categorized as hVISA with no vancomycin-intermediate S. aureus (VISA) strains. These hVISA isolates were significantly prevalent in MRSA than in MSSA (23.1% vs 7.9%, p = 0.014).
The distribution of isolates in each section was shown in Table S1. Strains from the thyroid unit accounted for most (11.8%) of S. aureus, but only three MRSA isolates were found, and none of the strains was hVISA. The pediatric department had the highest proportion of MRSA (50%) and the largest number of hVISA isolates (n = 8). None of the S. aureus from dermatology harboured mecA gene, but two cases were hVISA isolates (MSSA-6219 for 0.911 and MSSA-6550 for 0.95).
S.aureus from pus sources were the most abundant, accounting for 38.24% (data was not shown). MRSA strains were mainly isolated from pus and sputum (12/39, 30.8%, respectively), whereas, hVISA was more common in blood samples (8/22, 36.4%). Notably, MR-hVISA was blood or sputum-borne (n = 5 and 4, respectively), while MS-hVISA was of wide origin (secretions, n = 4; blood, n = 3; sputum, n = 3; pus, n = 2; transudation, n = 1).
Molecular Types of S. aureus
The 204 S. aureus isolates included 57 different spa types that belonged to four agr groups which were not duplicated. The most common spa types were t309 (63/204, 30.9%). agrI had 37 spa types with the largest number of isolates (n = 165), among which 63 were divided into t309, 24 had t078, 22 exhibited t437, while the remaining 56 isolates exhibited 34 spa types. Seventeen isolates belonged to agrII were grouped into 8 spa types with the most abundant being t002. Six isolates covering 3 spa types belonged to agrIII group. Sixteen isolates classified as agrⅣ yielded 9 spa types. Five novel spa types, namely t20226, t20227, t20228, t20229 and t20230 were observed.
t034, t309, t437 and t062 were distributed in both MRSA and MSSA groups, with t437 being more prevalent in the MRSA group (56.1% vs 1.2%, p = 0.001), whereas, t309 was more rich in the MSSA group (37.0% vs 5.1%, p = 0.001). All strains belonging to t078 and t002 were MSSA, in contrast, isolates classified as t2431 were all MRSA. There were no MRSA bacteria in group agr III and agr Ⅳ (Table S2).
A large number of spa types were distributed in both hVISA and VSSA, with t002, t062 and t2431 being more abundant in hVISA. None of the strains in the agr group Ⅳ was termed as hVISA.
MLST revealed 8 distinct STs for the MRSA isolates, of which ST59 (61.5%) was the most prevalent sequence type, followed by ST72 (20.5%), ST22 (5.1%), and other STs (ST9, ST965, ST1232, newly assigned ST7196 and ST7437). There were 10 different STs among hVISA isolates. Specifically, 9 MR-hVISA belonged to ST72 (n = 4), ST59 (n = 3), ST965 (n = 1) and novel ST7196 (n = 1). Thirteen MS-hVISA were assigned to ST25 (n = 4), ST5 (n = 2), while the remaining four STs had one isolate each. The phylogenetic tree based on alignment of 3186 single nucleotide polymorphisms of 7 housekeeping genes revealed that ST59/7437, ST15/5459 and ST5/965/7197 were from 3 clonal complexes (CC59, CC15 and CC5) in which the strains evolved in close evolutionary distance to each other (Figure 1).
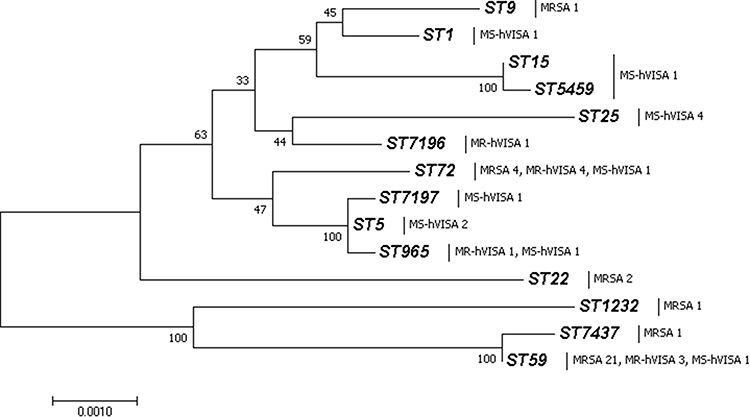 |
Figure 1 Molecular Phylogenetic analysis of all MRSA and hVISA isolate detected for MLST data by Neighbor-Joining method. ST59 and ST7437 belonged to CC59, CC15 contained ST15 and ST5459, ST965, ST7197 and ST5 grouped into CC5. |
SCCmec typing revealed that 87.2% of the MRSA isolates were type IV, which contained only two subtypes IVa (n = 24) and IVF (n = 10). Among the remaining isolates, 3 carried SCCmec III, one was SCCmec Ⅴ, and the last one failed to type. All the 9 MR-hVISA were classified as type IV.
A high correlation between the molecular typing of the strains was observed. ST-SCCmec combination showed that all ST72-positive isolates were grouped into SCCmec IVF. CC59 was determined as type IVa, except for one ST59 strain that was identified as SCCmec III. ST(CC)-spa combination indicated CC5 shared t002, t2431 strains belonged to ST72. ST-agr combination revealed that ST1 was grouped into agr III. CC5, CC15 and monomeric ST9 were identified as agr II. The remaining STs or CCs were classified as agrI.
ST59-t437-agrI-Ⅳa was the most prevalent genotype, occurring in 53.8% of the MRSA population (n = 21), followed by ST72-t2431-agrI-ⅣF (n = 6, 15.4%). Among the nine MR-hVISA isolates, the genotypes were ST59-t437-agrI-Ⅳa (33.3%), ST72-t2431-agrI-ⅣF (33.3%) with ST72-t5155-agrI-ⅣF, ST72-t20226-agrI-ⅣF and ST965-t062-agrII-ⅣF (11.1% for each phenotype). Molecular typing analysis revealed a wide genetic diversity among the 13 MS-hVISA isolates, which included 9 different STs, 10 distinct spa types and 3 agr groups.
Antimicrobial Activities
All S. aureus isolates were sensitive to FOX, LZD, DAP, TEC, VAN and MNO. Resistance rates of MRSA and hVISA groups to PEN, ERY and CLI were above 75% (Table 1).
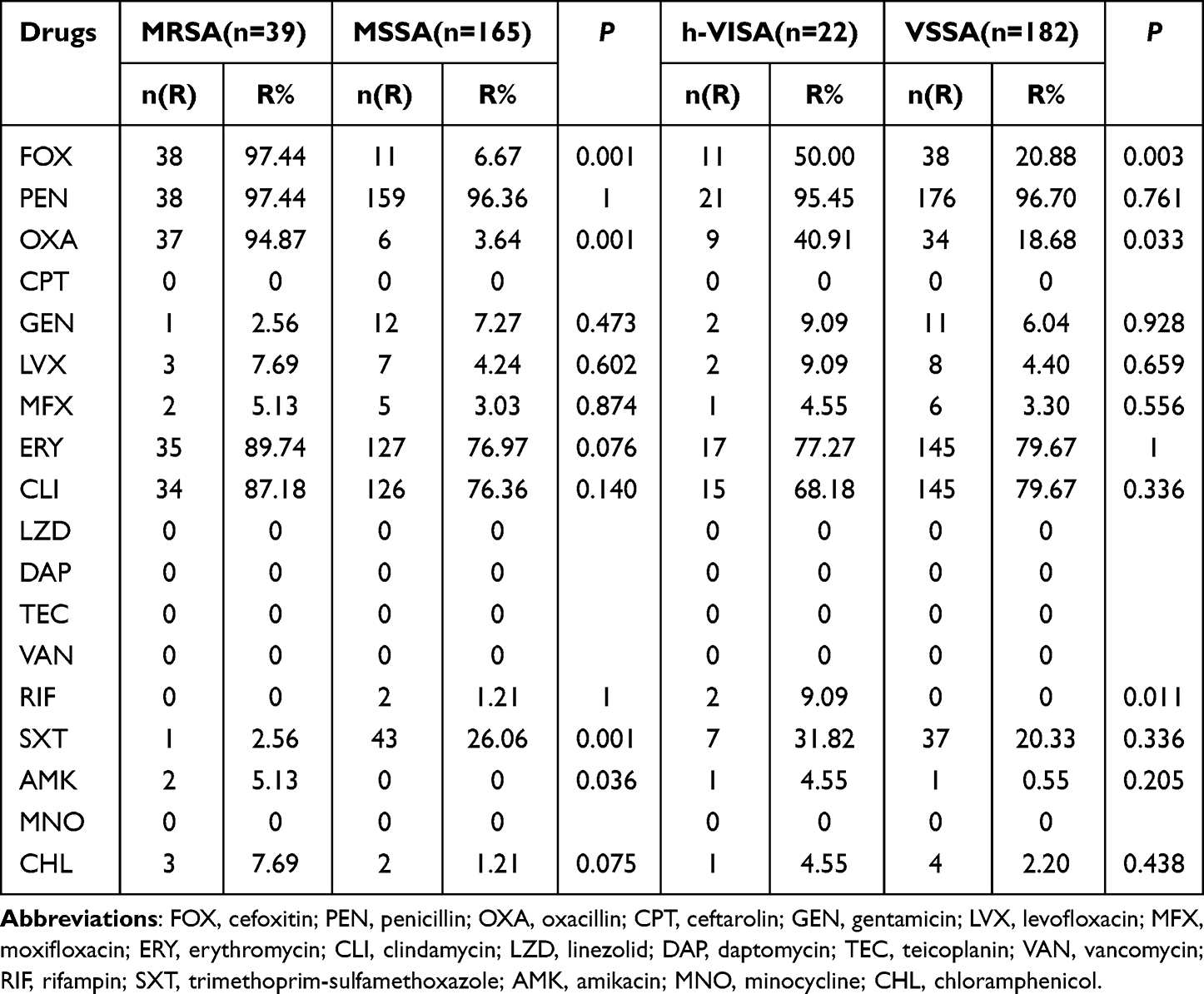 |
Table 1 Resistance Rate of S.aureus |
Compared to the MSSA group, the MRSA group was more resistant to FOX (97.44%), OXA (94.87%), AMK (5.13%) and CPT (92.31%), but exhibited low resistance to SXT (2.56% vs 26.06%). The MDR rate of MRSA isolates was significantly higher than that of MSSA isolates (92.3% vs 30.3%, p < 0.001). The most prevalent genotype among MRSA population ST59-t437-agrI-IVa (21/39, 53.8%) displayed resistance profile as FOX-OXA-PEN-ERY-CLI (17/21, 81.0%), consistant with other STs (Table 2).
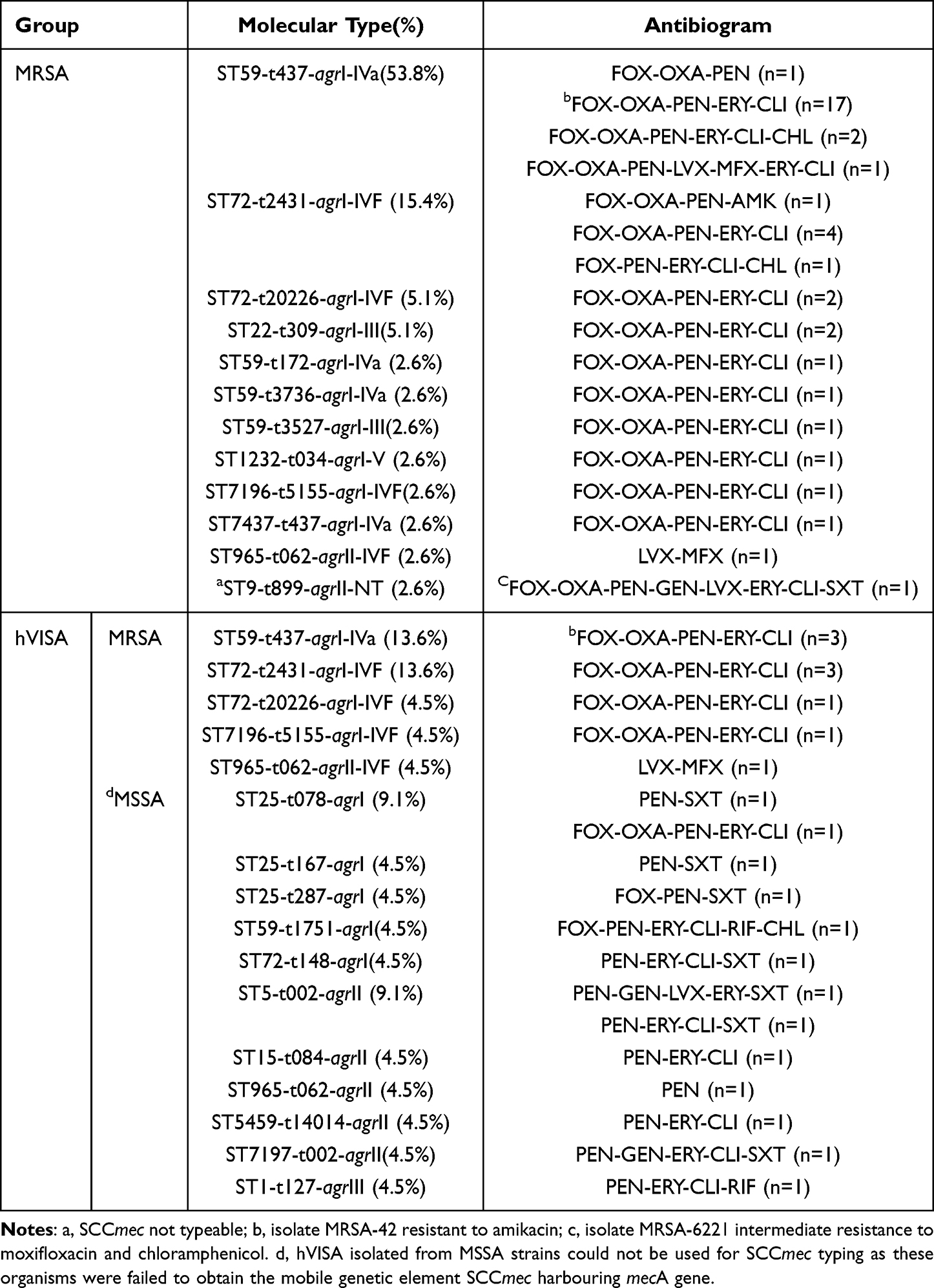 |
Table 2 Antibiogram of MRSA and hVISA Isolates in the Context of Different Genotypes |
Isolates from the hVISA group against FOX (50%), OXA (40.9%) and RIF (9.1%) were significantly higher than those from the VSSA group. Only 2 MS-hVISA strains were resistant to rifampicin while 2 MRSA (one MR-hVISA) strains were resistant to AMK. There were significant differences in MDR rate between the two groups (68.2% vs 39%, p = 0.009). 77.8% (7/9) of MR-hVISA strains expressed phenotypic resistance to FOX-OXA-PEN-ERY-CLI, while MS-hVISA isolates showed different resistance profiles. We found that in the MSSA population, MS-hVISA strains were more resistant to SXT than those without the hVISA phenotype (53.8% vs 23.7%, p = 0.040).
Discussion
As sample sizes have increased, laboratory personnel have found an increasing discordance between cefoxitin and oxacillin assays, or in both phenotypic and genetic analyses.20–22 Isolates MR-hVISA-33 and MRSA-6539 were mecA positive yet phenotypically susceptible to oxacillin (OS-MRSA), indicating the likely occurrence of frameshift mutations or single base substitutions in regions of nucleotide repeats within mecA, which have the ability to reverse into resistance when exposed to antibiotics.23,24 The other remaining 12 strains, despite exhibiting phenotypic resistance to oxacillin or/and cefoxitin, were not identified by PCR for the mecA gene and had negative results in the SCCmec typing assay, so we did not treat them as MRSA.
The regional clone prevalent in the region belongs to ST59-IVa-t437 in agreement with the findings of Li and Wang.25,26 Our findings are different from those of other national studies. ST8 strains were the most prevalent (59.8%) of PVL-positive MRSA in Japan, while ST59 clone consisted of 6 strains (6.5%).27 The main MRSA type in the UK was ST22,28 which accounted for 5.1% in our findings. The most prevalent molecular types of MRSA in the United States in 2015 were ST5 and ST8.29,30 Neither of which were not identified among our MRSA isolates; however, ST5 was present in 9.1% of hVISA isolates in our study.
The isolation rate of hVISA in S. aureus isolates accounted for 10.8%, which came from 23.1% of MRSA and 7.9% of MSSA. Most previous studies have focused on exploring hVISA from blood-borne MRSA isolates.31–33 however, our current study demonstrated that hVISA can be identified from a wide range of clinical samples and that a certain percentage of MSSA strains also belonged to hVISA. Previous studies have demonstrated that hVISA can persist in blood for a long time, which may be the reason why hVISA is more common in blood-borne MRSAs in our study.33 Also we suggest that the investigation of hVISA should be conducted from S. aureus from different sample sources. In our study, 22.7% of hVISA isolates belonged to ST72 and CC5 (ST5/965/7197), followed by CC59 (ST59) and ST25 (18.2%, respectively), which was different from some studies where ST5 clones were highly prevalent between hVISA strains (92.3%) in Latin American, but ST59 and ST72 were not found.34 A systematic review and meta-analysis showed that ST239 was the most prevalent hVISA clone (58.62%), accompanied by ST5 (14.45%), while both ST72 and ST59 were less than 3.5%.35 These studies imply the diversity of molecular genetic background of hVISA isolates.
97.4% and 92.3% of MRSA were susceptible to gentamicin and levofloxacin, respectively, which was significant when compared to findings by Riccobono et al (75%, 23.3%, with p = 0.002 and <0.001, respectively), but significantly low susceptibility to erythromycin (7.7% vs 29%, p = 0.005).36 The reason for this difference may have been due to selective pressures of this bacteria with different geographical distribution to the drug.
Our study showed a multidrug resistance pattern of MRSA represented by ST59-t437 exhibiting β-lactams, macrolides (erythromycin) and lincosamides (clindamycin), while being sensitive to rifampicin, levofloxacin and moxifloxacin (100%, 92.3% and 92.3%, respectively). The lower cost of resistance adaptation gives this clone an advantage in terms of growth and competition. The previously dominant genotype ST239 was not identified in our study, as it was shown that this clone is resistant to rifampicin, levofloxacin, moxifloxacin and tetracycline, and the additional acquired resistance gene necessarily leads to its evolutionary burden resulting in the loss of its advantage to become dominant.25,37 ST59 is only regionally prevalent, with major outbreaks in East Asia and sporadic occurrences in Australia, the Netherlands, and the United States.38–41 In the United States, 72.1% of ST59-t437 strains were erythromycin resistant, but the resistance rate to clindamycin was low (11.8%), which was inconsistent with our study (Table 1), presuming that the genome of ST59 clones in the United States contained the erythromycin resistance gene ermA, but lacked the lincosamides resistance gene ermB present in the East Asian ST59 clones.41 In addition to t437 and SCCmec IVa, the ST59 genealogy also contains t172, t3527, t3736 and type III SCCmec, but in low proportions, suggesting a diversity of gene structures in this clone; however, these did not alter the resistance phenotype of this clone, despite the presence of type III SCCmec carrying a higher number of resistance genes (plasmid pT181, transposon Tn554 and ψTn554).42
MRSA was common in pediatrics (50%), with the most frequently identified genotypes being ST72-IVF-t2431-agrI (6/11,54.5%), and secondly, ST59-IVa-t437-agrI(4/11,36.4%). Except for strain MRSA-110, which was resistance for only β-lactams and amikacin, the antibiotics susceptibility testing results of the two strain groups with very different molecular characteristics were almost identical, and the antibiogram was erythromycin-clindamycin-β-lactam, excluding MR-6539, which was additionally tolerant to chloramphenicol, indicating that the structural differences of resistance genes between the two lineages were not significant.
In addition, pediatrics was also the major source of hVISA in this study (31.8%). Notably, although MS-hVISA did not contain SCCmec, two of the three strains were more resistance to rifampicin or SXT than the five MR-hVISA strains with consistent resistance profiles, suggesting that mutations in the rpoB gene occurred during the evolution of the strain from methicillin-sensitive to heterogeneous intermediated resistance to vancomycin, or through the integration of other mobile genetic elements that confer resistance to erythromycin, clindamycin and SXT. This may also account for the higher resistance rate of MS-hVISA to trimethoprim-sulfamethoxazole than MSSA found in our study. Also, without considering the pediatric origin, a longitudinal view of the entire hVISA and VSSA populations, the more widespread distribution of the mecA responsible for resistance to cefoxitin and oxacillin in hVISA and the identification of only two rifampicin-resistant isolates among 204 strains as hVISA do not suggest that mecA and mutations in the rpoB gene associated with reduced susceptibility to vancomycin (Table 1).43,44
ST72-IVF-t2431-agrI just became the children clone in this region, contrary to the findings of the recognized “pediatric clone” ST5-IV.45,46 The results of antibiotics susceptibility testing revealed that the most common genotype ST59-MRSA-IV/V, representing the Chinese pediatric strain, had a resistance rate of more than 78.6% to erythromycin, but the resistance rate to clindamycin varies greatly (83.3% vs 14.3%, respectively).47,48 ST72-IV has been isolated in a small amount among the MRSA population of Chinese children, showing sensitivity to erythromycin and clindamycin.47 However, sporadically ST72-IVc from Japan and Portugal were resistant to both agents.49,50 There is no specific resistance spectrum for the dominant clone ST72-IV in the Korean pediatric population, but extensive data, including analysis of data from adult ST72-IV, demonstrated that this genotype is mostly sensitive to clindamycin, with varying rates of resistance to erythromycin.51–53 All ST72-IVF identified in our study were resistant to erythromycin and clindamycin, except strain MRSA-110 (Table 2).
SCCmec type IV contains many subtypes (Ⅳa to n), but we did not find any effect of subtypes on the resistance patterns, suggesting a weakened relationship between subtypes and resistance, Because (i) there is no SCCmec structure but still drug resistance (ii) no other resistance genes were found to be present in the type IV SCCmec structure except for mecA.42 Although it has been shown that in adults, ST72-IV causes lower mortality than ST5-II,52 nevertheless such a high number of MRSA and hVISA in pediatrics should be of concern to clinicians. Vancomycin and others still possess excellent antibacterial activity against MRSA and hVISA and can be used for anti-infective treatment caused by both (Table 1).
In summary, the frequency of MRSA and hVISA in the Yellow River Delta was 19.1% and 10.8%, respectively, with the predominant genotypes of MRSA were ST59-t437-agrI-IVa and ST72-t2431-agrI-IVF, and the majority were resistant to erythromycin and clindamycin. The primary clones of hVISA were CC5-agrII and ST72-agrI, but there was heterogeneity in spa typing. For MR-hVISA, phenotypic resistance was the same as MRSA. However, the resistance profiles of MS-hVISA to the antibiotics varies according to the diversity of molecular genetic background. Pediatrics as a primary source of MRSA, especially hVISA, is worrisome and highlights the necessity of rational use of drugs and improved management.
Ethical Statement
This study was approved by the Institutional Ethics Board of Binzhou Medical University Hospital (KYLL-2022-47).
Acknowledgments
We are grateful to Binzhou Medical University Hospital for their assistance in our study. We also thank the curators of the University of Würzburg, Germany for assigning new alleles, profiles and spa- types.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was supported by a grant from Natural Science Foundation of Shandong Province (project no. ZR2020MH309).
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Barber M. Methicillin-resistant staphylococci. J Clin Pathol. 1961;14:385–393. doi:10.1136/jcp.14.4.385
2. Hartman BJ, Tomasz A. Low-affinity penicillin-binding protein associated with beta-lactam resistance in Staphylococcus aureus. J Bacteriol. 1984;158(2):513–516. doi:10.1128/jb.158.2.513-516.1984
3. Boswihi SS, Udo EE, Monecke S, et al. Emerging variants of methicillin-resistant Staphylococcus aureus genotypes in Kuwait hospitals. PLoS One. 2018;13(4):e195933. doi:10.1371/journal.pone.0195933
4. Zhou W, Jin Y, Liu X, Chen Y, Shen P, Xiao Y. Comparison of genetic features and evolution of global and Chinese strains of Community-Associated Methicillin-Resistant staphylococcus aureus ST22. Microbiol Spectr. 2022;10(1):e203721. doi:10.1128/spectrum.02037-21
5. Wang X, Zhao H, Wang B, et al. Identification of methicillin-resistant Staphylococcus aureus ST8 isolates in China with potential high virulence. Emerg Microbes Infect. 2022;11(1):507–518. doi:10.1080/22221751.2022.2031310
6. Klein S, Hannesen J, Zanger P, Heeg K, Boutin S, Nurjadi D. Entry of Panton-Valentine leukocidin-positive methicillin-resistant Staphylococcus aureus into the hospital: prevalence and population structure in Heidelberg, Germany 2015-2018. Sci Rep. 2020;10(1):13243. doi:10.1038/s41598-020-70112-z
7. Ellington MJ, Perry C, Ganner M, et al. Clinical and molecular epidemiology of ciprofloxacin-susceptible MRSA encoding PVL in England and Wales. Eur J Clin Microbiol Infect Dis. 2009;28(9):1113–1121. doi:10.1007/s10096-009-0757-x
8. Geng W, Yang Y, Wang C, Deng L, Zheng Y, Shen X. Skin and soft tissue infections caused by community-associated methicillin-resistant staphylococcus aureus among children in China. Acta Paediatr. 2010;99(4):575–580. doi:10.1111/j.1651-2227.2009.01645.x
9. Walsh C. Deconstructing vancomycin. Science. 1999;284(5413):442–443. doi:10.1126/science.284.5413.442
10. Shen P, Zhou K, Wang Y, et al. High prevalence of a globally disseminated hypervirulent clone, Staphylococcus aureus CC121, with reduced vancomycin susceptibility in community settings in China. J Antimicrob Chemother. 2019;74(9):2537–2543. doi:10.1093/jac/dkz232
11. Zhang S, Sun X, Chang W, Dai Y, Ma X. Systematic review and Meta-Analysis of the epidemiology of Vancomycin-Intermediate and heterogeneous Vancomycin-Intermediate staphylococcus aureus isolates. PLoS One. 2015;10(8):e136082. doi:10.1371/journal.pone.0136082
12. Shariati A, Dadashi M, Moghadam MT, van Belkum A, Yaslianifard S, Darban-Sarokhalil D. Global prevalence and distribution of vancomycin resistant, vancomycin intermediate and heterogeneously vancomycin intermediate Staphylococcus aureus clinical isolates: a systematic review and meta-analysis. Sci Rep. 2020;10(1):12689. doi:10.1038/s41598-020-69058-z
13. Humphries R, Bobenchik AM, Hindler JA, Schuetz AN. Overview of changes to the clinical and laboratory standards institute performance standards for antibiotics susceptibility testing, m100. J Clin Microbiol. 2021;59(12):e21321. doi:10.1128/JCM.00213-21
14. Wootton M, Howe RA, Hillman R, Walsh TR, Bennett PM, Macgowan AP. A modified population analysis profile (PAP) method to detect hetero-resistance to vancomycin in Staphylococcus aureus in a UK hospital. J Antimicrob Chemother. 2001;47(4):399–403. doi:10.1093/jac/47.4.399
15. van Hal SJ, Wehrhahn MC, Barbagiannakos T, et al. Performance of various testing methodologies for detection of heteroresistant vancomycin-intermediate Staphylococcus aureus in bloodstream isolates. J Clin Microbiol. 2011;49(4):1489–1494. doi:10.1128/JCM.02302-10
16. Harmsen D, Claus H, Witte W, et al. Typing of Methicillin-Resistant Staphylococcus aureus in a University Hospital Setting by Using Novel Software for spa Repeat Determination and Database Management. J Clin Microbiol. 2003;41(12):5442–5448. doi:10.1128/JCM.41.12.5442-5448.2003
17. Wang W, Lee S, Chiueh T, Lu J. Molecular and Phenotypic Characteristics of Methicillin-Resistant and Vancomycin-Intermediate Staphylococcus aureus Isolates from Patients with Septic Arthritis. J Clin Microbiol. 2009;47(11):3617–3623. doi:10.1128/JCM.00539-09
18. Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol. 2000;38(3):1008–1015. doi:10.1128/JCM.38.3.1008-1015.2000
19. Mcclure-Warnier J, Conly JM, Zhang K. Multiplex PCR assay for typing of staphylococcal cassette chromosome mec types I to V in methicillin-resistant Staphylococcus aureus. J Vis Exp. 2013. doi:10.3791/50779
20. Felten A, Grandry B, Lagrange PH, Casin I. Evaluation of Three Techniques for Detection of Low-Level Methicillin-Resistant Staphylococcus aureus (MRSA): a Disk Diffusion Method with Cefoxitin and Moxalactam, the Vitek 2 System, and the MRSA-Screen Latex Agglutination Test. J Clin Microbiol. 2002;40(8):2766–2771. doi:10.1128/JCM.40.8.2766-2771.2002
21. Ho PL, Liu MC, Tong MK, et al. Evaluation of disc diffusion tests and agar screening for predicting mecA-mediated oxacillin resistance in Staphylococcus lugdunensis revealed a cefoxitin-susceptible, mecA-positive S. Lugdunensis clonal complex 27 clone. J Glob Antimicrob Resist. 2020;20:260–265. doi:10.1016/j.jgar.2019.08.021
22. Miller SA, Karichu J, Kohner P, et al. Multicenter Evaluation of a Modified Cefoxitin Disk Diffusion Method and PBP2a Testing To Predict mecA -Mediated Oxacillin Resistance in Atypical Staphylococcus aureus. J Clin Microbiol. 2017;55(2):485–494. doi:10.1128/JCM.02211-16
23. Gargis AS, Yoo BB, Lonsway DR, et al.. Difficult-To-Detect staphylococcus aureus : MecA-Positive isolates associated with oxacillin and cefoxitin False-Susceptible results. J Clin Microbial. 2020) doi: 128/JCM.02038–19
24. Goering RV, Swartzendruber EA, Obradovich AE, Tickler IA, Tenover FC. Emergence of oxacillin resistance in stealth Methicillin-Resistant staphylococcus aureus due to mecA sequence instability. Antimicrob Agents Chemother. 2019;63(8):e00558. doi:10.1128/AAC.00558-19
25. Li S, Sun S, Yang C, et al. The Changing Pattern of Population Structure of Staphylococcus aureus from Bacteremia in China from 2013 to 2016: ST239-030-MRSA Replaced by ST59-t437. Front Microbiol. 2018;9:332. doi:10.3389/fmicb.2018.00332
26. Wang B, Xu Y, Zhao H, et al. Methicillin-resistant Staphylococcus aureus in China: a multicentre longitudinal study and whole-genome sequencing. Emerg Microbes Infect. 2022;11(1):532–542. doi:10.1080/22221751.2022.2032373
27. Yamaguchi T, Nakamura I, Sato T, et al. Changes in the Genotypic Characteristics of Community-Acquired Methicillin-Resistant Staphylococcus aureus Collected in 244 Medical Facilities in Japan between 2010 and 2018: a Nationwide Surveillance. Microbiol Spectr. 2022:e227221. doi:10.1128/spectrum.02272-21
28. Holden MT, Hsu LY, Kurt K, et al. A genomic portrait of the emergence, evolution, and global spread of a methicillin-resistant Staphylococcus aureus pandemic. Genome Res. 2013;23(4):653–664. doi:10.1101/gr.147710.112
29. Kyung-Hwa P, Kerryl EG, James RU, et al. Molecular epidemiology of Staphylococcus aureus bacteremia in a single large Minnesota medical center in 2015 as assessed using MLST, core genome MLST and spa typing. PLoS One. 2017;12(6). doi:10.1371/journal.pone.0174953
30. Wang X, Zhao H, Wang B, et al. Identification of methicillin-resistant Staphylococcus aureus ST8 isolates in China with potential high virulence. Emerg Microbes Infect. 2022;11(1):507–518. doi:10.1080/22221751.2022.2031310
31. Musta AC, Riederer K, Shemes S, et al. Vancomycin MIC plus heteroresistance and outcome of methicillin-resistant Staphylococcus aureus bacteremia: trends over 11 years. J Clin Microbiol. 2009;47(6):1640–1644. doi:10.1128/JCM.02135-08
32. Ozmen CB, Tekeli A, Karahan ZC. Evaluation of the presence and characterization of Vancomycin-Intermediate and heterogeneous Vancomycin-Intermediate level resistance among bloodstream isolates of Methicillin-Resistant staphylococcus aureus. Microb Drug Resist. 2020;26(3):238–244. doi:10.1089/mdr.2019.0178
33. Hanaki H, Cui L, Ikeda-Dantsuji Y, et al. Antibiotic susceptibility survey of blood-borne MRSA isolates in Japan from 2008 through 2011. J Infect Chemother. 2014;20(9):527–534. doi:10.1016/j.jiac.2014.06.012
34. Castro BE, Berrio M, Vargas ML, et al. Detection of heterogeneous vancomycin intermediate resistance in MRSA isolates from Latin America. J Antimicrob Chemother. 2020;75(9):2424–2431. doi:10.1093/jac/dkaa221
35. Zhang S, Sun X, Chang W, Dai Y, Ma X. Systematic review and Meta-Analysis of the epidemiology of Vancomycin-Intermediate and heterogeneous Vancomycin-Intermediate staphylococcus aureus isolates. PLoS One. 2015;10(8):e136082. doi:10.1371/journal.pone.0136082
36. Riccobono E, Giani T, Baldi G, et al. Update on activity of dalbavancin and comparators against clinical isolates of Gram-positive pathogens from Europe and Russia (2017-2018), and on clonal distribution of MRSA. Int J Antimicrob Ag. 2021;59(2):106503.
37. Baines SL, Holt KE, Schultz MB, et al. Convergent adaptation in the dominant global hospital clone ST239 of methicillin-resistant Staphylococcus aureus. Mbio. 2015;6(2):e80. doi:10.1128/mBio.00080-15
38. Diep BA, Carleton HA, Chang RF, Sensabaugh GF, Perdreau-Remington F. Roles of 34 virulence genes in the evolution of hospital- and community-associated strains of methicillin-resistant Staphylococcus aureus. J Infect Dis. 2006;193(11):1495–1503. doi:10.1086/503777
39. Chuang YY, Huang YC. Molecular epidemiology of community-associated meticillin-resistant Staphylococcus aureus in Asia. Lancet Infect Dis. 2013;13(8):698–708. doi:10.1016/S1473-3099(13)
40. Coombs GW, Monecke S, Ehricht R, et al. Differentiation of clonal complex 59 community-associated methicillin-resistant Staphylococcus aureus in Western Australia. Antimicrob Agents Chemother. 2010;54(5):1914–1921. doi:10.1128/AAC.01287-09
41. Ward MJ, Goncheva M, Richardson E, et al. Identification of source and sink populations for the emergence and global spread of the East-Asia clone of community-associated MRSA. Genome Biol. 2016;17(1):160. doi:10.1186/s13059-016-1022-0
42. Lakhundi S, Zhang K. Methicillin-Resistant Staphylococcus aureus: molecular Characterization, Evolution, and Epidemiology. Clin Microbiol Rev. 2018;31(4). doi:10.1128/CMR.00020-18
43. Guo Y, Wang B, Rao L, et al. Molecular characteristics of Rifampin-Sensitive and -Resistant isolates and characteristics of rpoB gene mutations in Methicillin-Resistant staphylococcus aureus. Infect Drug Resist. 2021;14:4591–4600. doi:10.2147/IDR.S336200
44. Mlynarczyk-Bonikowska B, Kowalewski C, Krolak-Ulinska A, Marusza W. Molecular mechanisms of Drug Resistance in Staphylococcus aureus. Int J Mol Sci. 2022;23(15):8088. doi:10.3390/ijms23158088
45. Hirose M, Aung MS, Fukuda A, Murata Y, Saitoh M, Kobayashi N. Prevalence and Genetic Characteristics of Methicillin-Resistant Staphylococcus aureus and Coagulase-Negative Staphylococci Isolated from Oral Cavity of Healthy Children in Japan. Microb Drug Resist. 2019;25(3):400–407. doi:10.1089/mdr.2018.0333
46. Moreno MP, Vargas JM, Vivaldo S, et al. Molecular epidemiology of methicillin-resistant Staphylococcus aureus from different population groups in Argentina. J Glob Antimicrob Resist. 2020;23:82–86. doi:10.1016/j.jgar.2020.07.016
47. Wang X, Liu Q, Zhang H, et al. Molecular Characteristics of Community-Associated Staphylococcus aureus Isolates From Pediatric Patients with Bloodstream Infections Between 2012 and 2017 in Shanghai, China. Front Microbiol. 2018;9:1211. doi:10.3389/fmicb.2018.01211
48. Zhao R, Wang X, Wang X, et al. Molecular characterization and virulence gene profiling of methicillin-resistant Staphylococcus aureus associated with bloodstream infections in southern China. Front Microbiol. 2022;13:1008052. doi:10.3389/fmicb.2022.1008052
49. Aung MS, Urushibara N, Kawaguchiya M, et al. Clonal Diversity and Genetic Characteristics of Methicillin-Resistant Staphylococcus aureus Isolates from a Tertiary Care Hospital in Japan. Microb Drug Resist. 2019;25(8):1164–1175. doi:10.1089/mdr.2018.0468
50. Tavares DA, Sa-Leao R, Miragaia M, de Lencastre H. Large screening of CA-MRSA among Staphylococcus aureus colonizing healthy young children living in two areas (urban and rural) of Portugal. Bmc Infect Dis. 2010;10:110. doi:10.1186/1471-2334-10-110
51. Sung JY, Lee J, Choi EH, Lee HJ. Changes in molecular epidemiology of community-associated and health care-associated methicillin-resistant Staphylococcus aureus in Korean children. Diagn Microbiol Infect Dis. 2012;74(1):28–33. doi:10.1016/j.diagmicrobio.2012.05.018
52. Park KH, Chong YP, Kim SH, et al. Community-associated MRSA strain ST72-SCCmecIV causing bloodstream infections: clinical outcomes and bacterial virulence factors. J Antimicrob Chemother. 2015;70(4):1185–1192. doi:10.1093/jac/dku475
53. Park SH, Park C, Yoo JH, et al. Emergence of community-associated methicillin-resistant Staphylococcus aureus strains as a cause of healthcare-associated bloodstream infections in Korea. Infect Control Hosp Epidemiol. 2009;30(2):146–155. doi:10.1086/593953
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.