")
Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 13
Research Progress on the Experimental Animal Model of Gestational Diabetes Mellitus
Authors He Y , Wu N , Yu W, Li L, OuYang H, Liu X, Qian M, Al-Mureish A
Received 17 July 2020
Accepted for publication 5 October 2020
Published 9 November 2020 Volume 2020:13 Pages 4235—4247
DOI https://doi.org/10.2147/DMSO.S270935
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Konstantinos Tziomalos
Yujing He,1 Na Wu,1,2 Wenshu Yu,1 Ling Li,1 Hong OuYang,1 Xinyan Liu,1 Meichen Qian,1 A Al-Mureish1
1Department of Endocrinology, Shengjing Hospital of China Medical University, Shenyang, People’s Republic of China; 2Clinical Skills Practice Teaching Center, Shengjing Hospital of China Medical University, Shenyang, 110004, People’s Republic of China
Correspondence: Na Wu
Department of Endocrinology, Shengjing Hospital of China Medical University, 36 Sanhao Road, Heping District, Shenyang, Liaoning Province 110004, People’s Republic of China
Tel +86 18940258445
Email [email protected]
Abstract: Gestational diabetes mellitus (GDM) refers to different degrees of abnormal glucose metabolism during pregnancy, where blood glucose levels do not reach the level of overt diabetes, accounting for 80– 90% of pregnancy with hyperglycemia. Hyperglycemia affects the pregnancy process, leading to a series of adverse maternal outcomes that have a profound impact on the future of the offspring. The establishing of an appropriate GDM model will provide theoretical basis for study GDM pathogenesis involves, the choice of resonable drugs and the observation the disease trends and outcomes. At present, there are many methods for establishing experimental GDM animal models and animal choices. This paper examines the different GDM models and their induction methods.
Keywords: gestational diabetes mellitus, animal model, chemical induction, high-sugar and high-fat diet, genetic manipulation
Introduction
With the gradual improvement of people’s living standards around the world, the incidence of diabetes has increased year on year, with the number of hyperglycemia in pregnancy increasing significantly. The prevalence of GDM varies greatly between countries and even between regions within a country. Epidemiological studies show an incidence of more than 9% GDM in the United States and between 3.0% and 21.2% in Asian countries.1 Hyperglycemia during pregnancy will cause various complications for pregnant women and fetuses, even endanger life and health, and will have a close impact on the outcome of pregnancy. It adversely affects the long-term blood glucose and blood lipid levels in the offspring. However, the pathogenesis of GDM has not yet been fully clarified. The establishment of ideal GDM animal models will therefore provide a theoretical basis for early detection and prevention of GDM and subsequent clinical medication and drug evaluation, in order to better control the occurrence and development of GDM and better improvement of the outcome of mothers and children.
In recent years, the main methods for establishing GDM animal models include experimental animal model, spontaneous animal model and transgenic animal model. Currently, many researchers select experimental animal models that is they use physical, chemical, and biological pathogenic factors to cause certain damage to animal tissues, organs and even the body as a whole to induce some functions, metabolism, or toxic side effects similar to those of human diseases. Researchers have adopted a variety of methods in the GDM study, and have used various animals for the experiment, including dogs, sheep, pigs, primates, and rodents.2 This study provides a brief review of the animals used in current researches, and the most commonly used methods of induction.
Introduction of GDM
GDM first appeared in the medical literature in the early 1970s. In 1979, the World Health Organization (WHO) identified GDM as an independent type of diabetes.3 The state of hyperglycemia during pregnancy is divided into gestational diabetes mellitus (GDM), overt diabetes mellitus (ODM) and pre-gestational diabetes mellitus (PGDM). Among these hyperglycemic variations, GDM refers to abnormal glucose metabolism in which blood glucose does not reach the level of overt diabetes during pregnancy, accounting for 80–90% of hyperglycemia during pregnancy.4 If the control of blood glucose in pregnant women is not good, it will not only increase the maternal risk of preeclampsia, abortion, stillbirth, premature birth, but will also lead to neonatal hypoglycemia, hyperbilirubinemia, overweight and other situations, and even cause long-term adverse effects on the blood glucose and the blood lipid of the offspring, and even endanger life and health in serious cases.5 At present, the diagnostic criteria for GDM in the different guidelines are different, as shown in Table 1.4
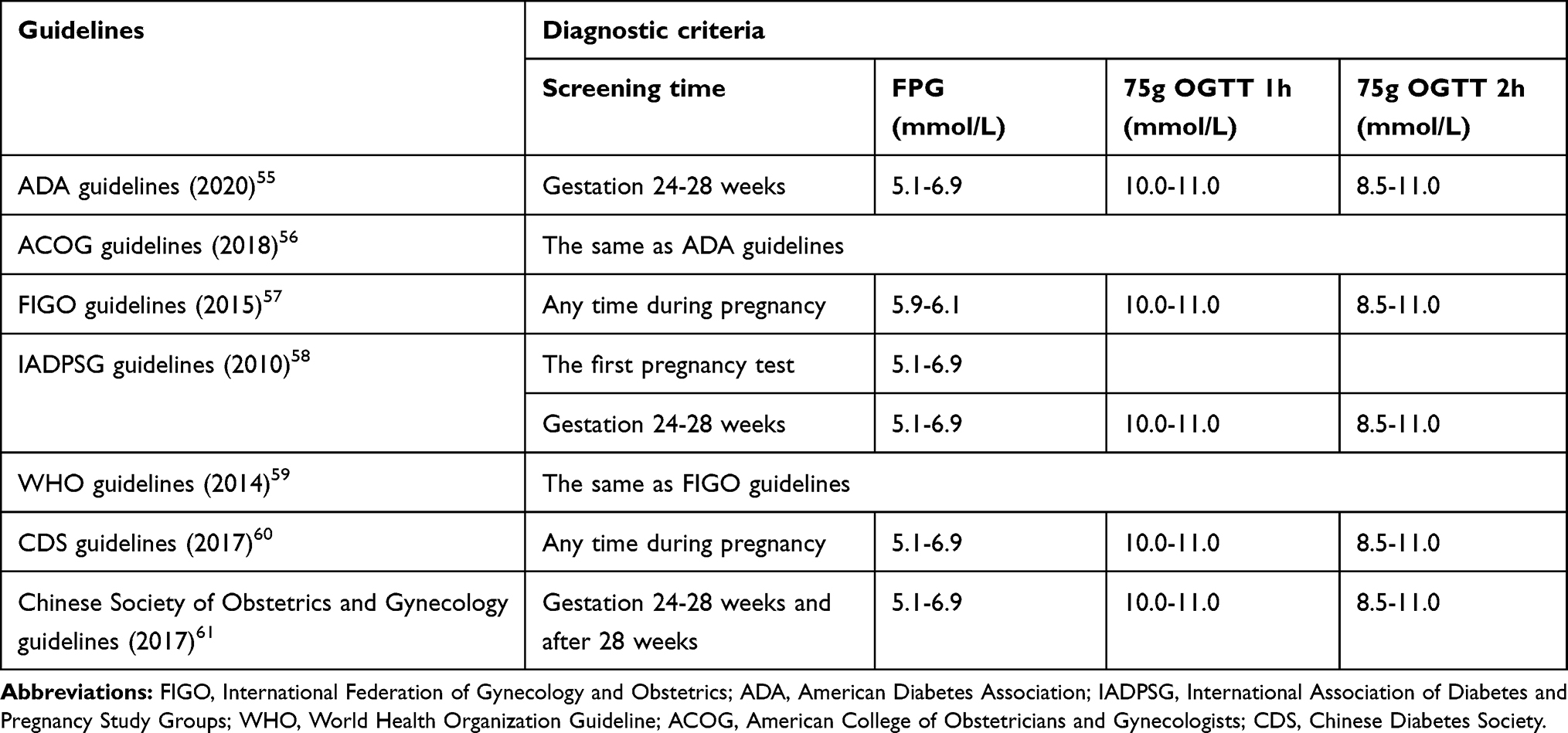 |
Table 1 Different Guidelines for GDM Diagnostic Criteria |
Relationship Between Animal Model and GDM
The animal model of human disease refers to the animal with human disease simulation performance established in various medical scientific research. There are three main animal models mentioned above: experimental animal model, spontaneous animal model and transgenic animal model. The spontaneous animal diseases are very similar to the corresponding human diseases in the occurrence, and development that occurs under natural conditions, so the application value of spontaneous animal models is very high, but the source of such models is limited and it is impossible to apply them in large quantities.6 Transgenic animal models are expensive with a long reproduction time, but the effective control of a single factor makes the experiment more scientific, rigorous, and reliable. The transgenic animal models can therefore help to find the cause inaccurately, and are more suitable for the mechanism of diabetes or drug targeted research. As a result, in the current research field of gestational diabetes, the method of constructing experimental animal models is often used, that is, researchers use physical, chemical and biological pathogenic factors to cause certain damage to the tissues, organs and even the entire body of animals, and some functions, metabolism or side effects similar to human diseases. Experimental animal models have the characteristics of high reproducibility, appropriate conditions, short experimental period, and easy control of disease characteristics, therefore they are commonly used in medical research.
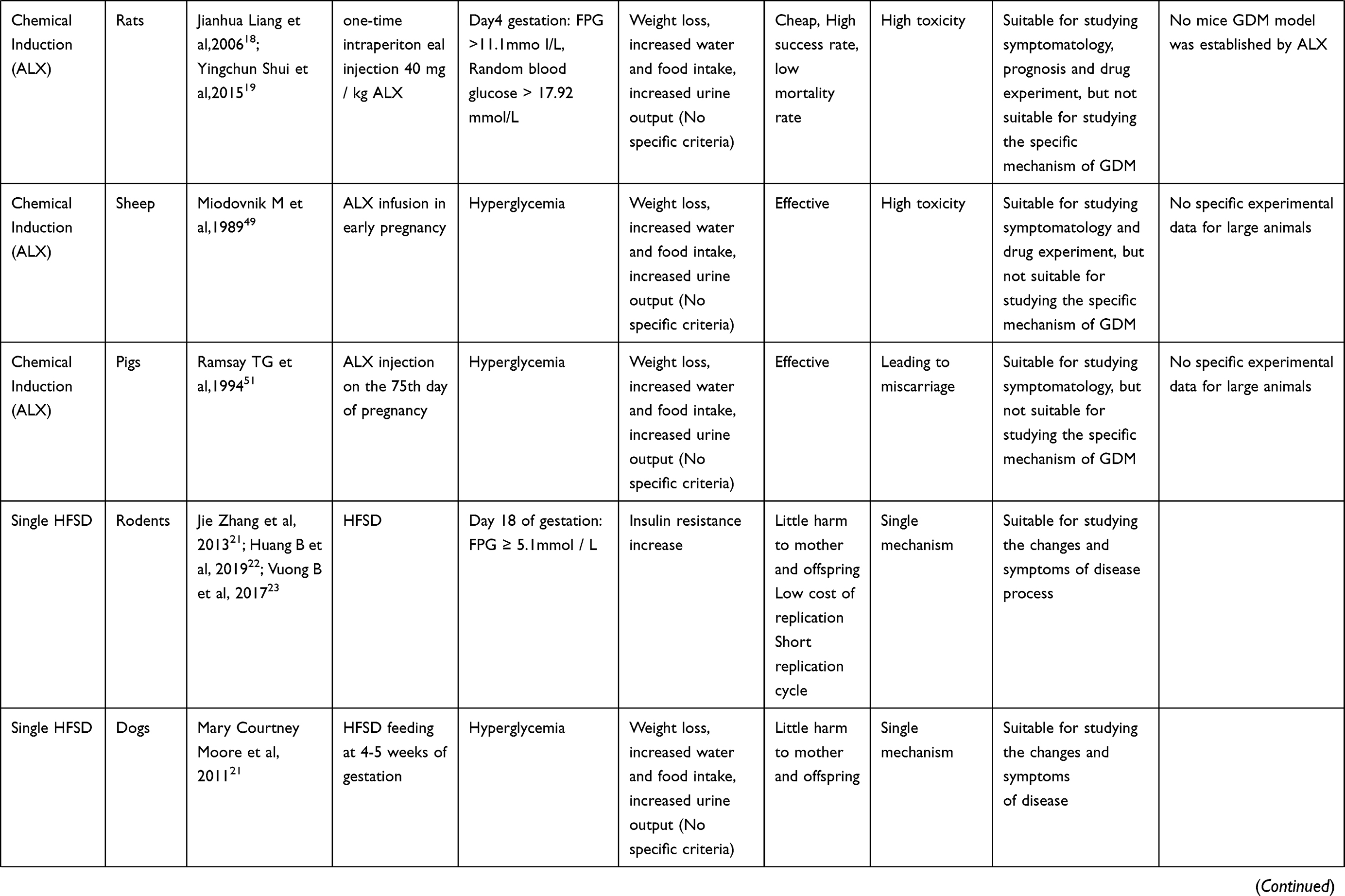
At present, the research focus on the pathogenesis of GDM is mainly reflected in the following aspects: insulin resistance; insulin antagonistic hormone release; abnormal insulin signal transduction pathway; cells, inflammatory factors; genetic factors. In the field which has been developed, animal experiments have made an indelible contribution. For example, when it comes to insulin resistance, through the study of rodents, we find that β-cell changes in pregnancy are similar to human beings. In other words, pancreatic β cells use a variety of adaptive mechanisms in early pregnancy, including increasing the quality, quantity and insulin secretion capacity, increasing the number of beta cells, and strengthening their functions to cope with the increase of insulin resistance in late pregnancy.7,8 Although the mechanism is not the same, it provides guidance for exploring the inducement and mechanism of human GDM, and also provides a signal for early prevention and late follow-up. Establishing an ideal animal model of GDM is helpful in better exploring the mechanism, causes and development process of GDM, helping to predict, prevent and treat GDM, and provide experimental basis for new methods of treatment for Chinese and Western medicine. The specific animal types and construction methods of the model are presented below and the criteria for successful modeling are shown in Table 2.
 |
 |
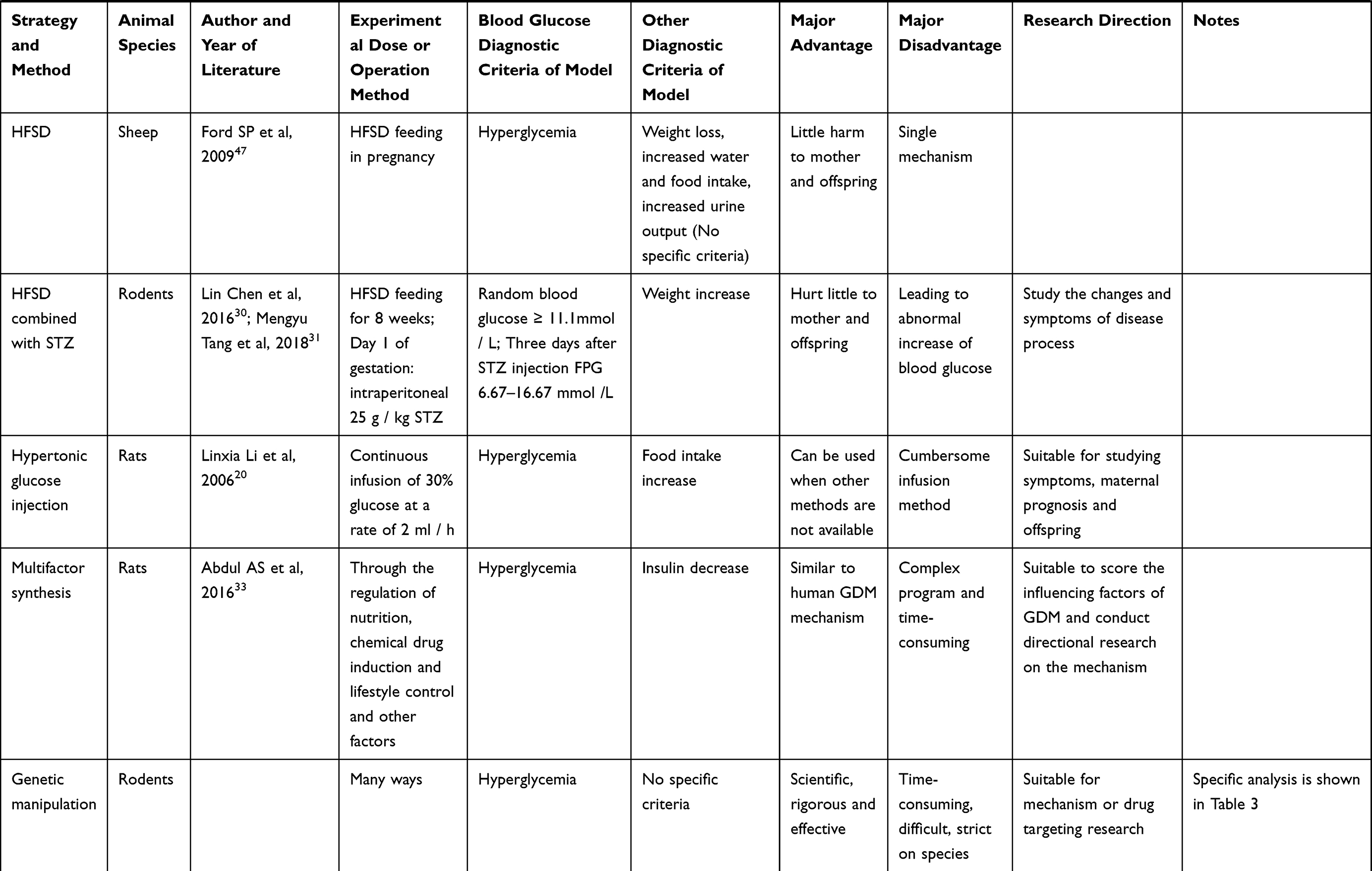 |
Table 2 Summary of Animal Model Construction |
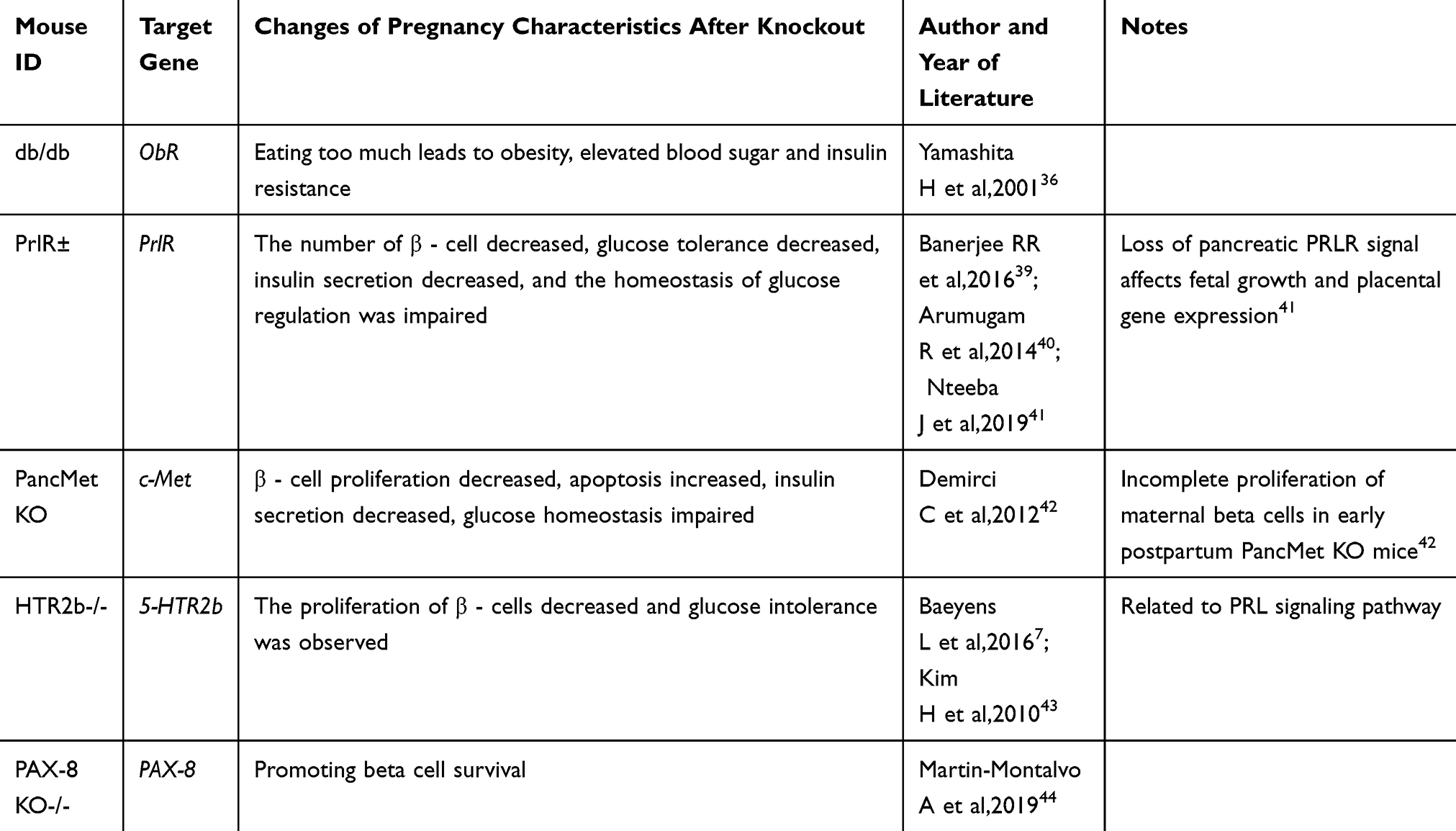 |
Table 3 GDM Genetic Manipulation Mouse Model |
Animal Model Analysis
Rodents
Chemical Induction
Streptozotocin is an antibiotic extracted from streptomycin, which has a highly selective toxic effect on the islet β cells of experimental animals, which can make insulin secretion insufficient and increase blood glucose, and is therefore one of the most commonly used drugs in animal models for diabetic induction.9 Since Rakieten et al first reported the effect of STZ on DM in 1963, many researchers used different doses of STZ to replicate DM animal models, and this method is still widely used in DM-related animal studies.10 STZ can induce a stable and rapid DM animal models, has low animal category selectivity even in small doses, and relatively has low tissue toxicity, and good stability. It may cause diabetes by damaging islet β cells with nitric oxide (NO) and free radicals, impairing the function of islet β cells, and reducing insulin synthesis. In the same species, the degree of cell damage caused by STZ depends on the amount of STZ, ie, low-dose STZ induces β cell apoptosis, and high-dose STZ causes β cell necrosis, which is very similar to human diabetes pathogenesis.11 Different doses are suitable for the construction of T1DM and T2DM models. The model induced by single large dose of STZ is more consistent with T1DM, while the model induced by small dose STZ is more consistent with T2DM.12 Based on this modeling method, it also has a certain guiding role in the construction of the GDM model. However, due to the embryonic factors caused by pregnancy, the application of STZ dose in the GDM model is limited, so the STZ dosage will also be discussed in the modeling process.
Previously, Hao et al had conducted experiments to compare the modeling rate and stability of the one-time high-dose STZ injection and multiple low-dose injection methods for the development of the gestational diabetic mice model. The membrane rate of 30mg/kg.bw (body weight) was 70%. Compared with the control group, the state of hyperglycemia lasted a long time and the weight gain decreased significantly. The conclusion was STZ 30mg/kg.bw, intraperitoneal injection 24 hours apart, three times injection is a relatively good method for establishing a model of gestational diabetes mice with has good stability.13 Li et al randomly divided 46 SD pregnant rats into low-dose, medium-dose and high-dose STZ groups in the preparation of the GDM models, injected 35, 45, 60 mg/kg.bw of STZ intraperitoneally on the day of pregnancy confirmation, measured the fasting blood glucose on the 3rd, 9th, 14th, and 19th days of pregnancy, observed the changes of bodyweight, water intake, food intake and urine volume. Pregnant rats in the 45 mg/kg STZ group were found to have 3 P’s of diabetes after medication, with the highest modeling success rate of 83.3% and the lowest rate of negative conversion.14 Meanwhile, in recent years, experimental data showed that the GDM rat model could be established by one-time intraperitoneal injection of 25 mg/kg.bw STZ to SD pregnant rats, the blood glucose of pregnant rats after modeling fluctuated between 6.7 and 8.3 mmol/L, and the modeling success rate was 100%. In an ideal clinical situation, overly high blood glucose levels during pregnancy can often be controlled, and the limited growth rate of the offspring in the uterus is significantly reduced, so at present, a single small dose 25 mg/kg.bw intraperitoneal injection of STZ in the establishment of a pregnant rat model is more in line with the clinical characteristics of human GDM and is one of the ideal methods for establishing experimental GDM rat models.15 Although STZ is expensive and has more applications in the construction of DM animal models, it has the advantages of small dosage, simple modeling method, low drug toxicity, and low mortality of modeling animals.
Alloxan is a β cell cytotoxic agent synthesized by the oxidation of uric acid. It is chemically unstable compound that generates oxygen free radicals and reacts with the SH group (mainly cysteine). It induces hyperpolarization of the cytoplasmic membrane potential, inhibits glucose-induced membrane impulses, which in turn inhibits voltage-dependent Ca2+ influx, destroys intracellular free Ca2 + concentration fluctuations, and affects the cell metabolism. Alloxan selectively damages the pancreatic islet β cells of various animals because the animals have high levels of SH groups in islet β cells, which impair the function of insulin-producing β-cells resulting in hyperglycemia and diabetes.16 Alloxan also damages β cells by inhibiting the expression of glucose transfer factor and glucokinase mRNA, which are necessary substances for the transport and metabolism of glucose.17
The effects of ALX on GDM induction are different with the route and dose in animals. Pregnant rats were randomly divided into groups administered intravenously and intraperitoneally. Pregnant rats were also divided randomly as intravenous high dose 60 mg/kg.bw, medium dose 40 mg/kg.bw and low dose 20 mg/kg.bw subgroups. The intraperitoneal administered pregnant rats were divided into high dose 120 mg/kg.bw and medium dose 100 mg/kg.bw subgroups. The effects of different doses of ALX and different routes of drug administration on the modeling success rate have been studied. It was observed that the 40 mg/kg.bw intravenous injection of ALX had a good modeling success rate of 91.7%, a low mortality rate of 8.3%, and a low negative conversion rate of 9.1%, and that the best dose for induction of GDM was chosen in pregnant rat models.18 ALX is highly toxic inducing great damage to animals for modeling, but is inexpensive in establishing a stable model with a high modeling success rate and low mortality. Therefore, alloxan-induced animal diabetes models are widely used in the development and selection of hypoglycemic drugs.19
Hypertonic Glucose Injection
Continuous intravenous injection of hypertonic glucose induces hyperglycemia during pregnancy, and is performed through intubation and fixation of the internal jugular veins after 16.5 days of mating. Since both STZ and ALX have a relatively high mortality rate in animals as well as pregnancy failure along with an unstable blood glucose levels in fetal rats,20 it was possible to establish a gestational hyperglycemia by continuous injection of 30% glucose into pregnant rats at a rate of 2 mL/h. However, the above-mentioned method of hypertonic glucose injection is slightly cumbersome and inconvenient to operate in actual experiments, and the method of hypertonic glucose injection is not very simple, whereas the GDM mechanism found to be more complicated so that it is not widely used in actual animal research.
Simple High-Sugar and High-Fat Diet Induction
The intake of sugars, lipids, and proteins increases during pregnancy. High lipid content in the diet pattern during pregnancy causes not only obesity but also GDM during pregnancy. Excessive intake of high-calorie foods in the diet increases the accumulation of fat in the body, promotes the secretion of obesity-causing adipokines, leading to insulin resistance through various related pathways. Combining the clinical features of gestational hyperglycemia and insulin resistance pathogenesis, dietary induction methods have been developed.
During the establishment of high sugar and high fat diet (HFSD) induced diabetes animal models, experimental animals are given a specific high-sugar and high-fat diet to significantly increase their fasting insulin and decrease their insulin sensitivity index, and thereby induce insulin resistance, which is one of the characteristics of GDM. A successful GDM was established in pregnant mice with a high sugar and high-fat diet. The modelling success rate of pregnant rats was 53.3%, and the fasting blood glucose level of the GDM model group was higher than that of the normal control group from the 10th day of pregnancy. On the 18th day of pregnancy, the fasting blood glucose level of the GDM model group was still higher than that of the normal control group. These were similar to the changes in the characteristics of human GDM blood glucose levels. At the same time, the FINS was measured and the insulin resistance index was calculated on the 18th day of pregnancy and the result indicated insulin resistance, which is consistent with the GDM pathophysiological mechanism.21 Huang et al also used this method to establish a GDM mouse model and conducted in-depth research on it.22 Vuong et al established a model and further studied the effect of GDM on the activation of microglia in the cerebellum and the chronic inflammatory response of the offspring.23 This kind of experiments eliminate the embryonic influence of drugs and to some extent, damages the mother’s organ function and restore the process of human GDM. In addition, high glucose and high-fat diet induction can lead to visceral fat increase, lipid metabolism disorder is one of the important pathophysiological characteristics of GDM.24 There is a certain degree of rationality.
High-Fat Diet Combined with Streptozotocin
The levels of CRP factors, leptin, and adiponectin have undergone special changes in the GDM process, which may become a new identification point for the discovery and prevention of GDM.25–29 Some researchers reported that after high-fat diet induction was combined with low-dose STZ intraperitoneal injection, the food intake, water intake and urine volume of mice increased significantly, the blood glucose level, insulin level, serum C-reactive protein content, leptin level increased significantly, while the adiponectin level was decreased significantly. However, the glucose level returned to normal after delivery, and the modeling success rate was approximately 60% with no mortality compared to control group suggesting that it was the ideal method for establishing gestational diabetes model.30 Tang et al explored the optimal HFSD scheme combined with STZ to establish gestational diabetes rat model, and determined the optimal dose of STZ. The model rate of 25 mg/kg group was 62.5%, the highest in the three groups, and the mortality rate was 12.5%, the lowest in the three groups. Blood glucose of pregnant rats increased significantly after STZ injection, and was stable in the 25 mg/kg group.31 The combination of low-dose STZ and HFSD can induce an animal model similar to diabetes. Compared with the simple HFSD model, the model setting rate is improved. Compared with the simple STZ induced model, it is more abundant and close to the actual human condition, which improves the animal model rate of this kind. It has more important medical research significance in the evaluation of drug efficacy in the treatment of diabetes.32
Combination of Multiple Factors
In recent years, Siti Hajar Abdul Aziz has provided a new method. A common diet was given to rats in the control group, and since the 10th week, rats in the GDM group have been fed HFSD. The mating behavior was then performed. At day 0 of pregnancy, 120 mg/kg.bw (body weight) of niacinamide was injected intraperitoneally, and 15 minutes later, 35 mg/kg.bw of STZ was dissolved in 0.1 mol/L citrate buffer (pH 4.5) and injected intraperitoneally. At the same time, the sleep mode of rats was controlled in order to induce stress. Various parameters of rats, including the blood glucose parameters, lipid profiles and genes that affect insulin signaling, have been measured. The results showed that GDM rats showed significantly higher body weight and blood glucose level, as well as lower plasma insulin levels and C-peptide compared with the controls.33 Although this method is cumbersome and time-consuming, it is closer to the real human GDM situation in the GDM mechanism. Based on these results, it can be concluded that an appropriate GDM animal model can be developed through a combination of factors such as nutrition, chemical drug induction, and lifestyle control. This kind of model can be used to further understand the pathophysiology of GDM and thus find new methods for GDM therapy.
Genetic Manipulation
Genetic factors are a very important part in the pathogenesis of GDM. Many studies have conducted in-depth researches on genes and genetics shown in Table 3.
Db/db mice are leptin receptor mutants (ObR), which are characterized by the inability to fully inhibit eating behavior, and their female mice are infertile, but the heterozygous females with mutant allele (db/+) are fertile and fasting blood glucose and insulin levels are normal in non-pregnant states. Excessive appetite of db/+ pregnant rats, however, leads to obesity, develops insulin resistance, and increase blood glucose, and phenotypic GDM is characterized by moderate glucose tolerance.34–36
The increase in prolactin is one of many physiological changes during the pregnancy of female mice and human beings. One of the key consequences of prolactin elevation is that the PRLR signal increases cell proliferation, which leads to an increase in pancreatic value in the presence of PRLR, and secretes more insulin to resist body changes and maintain blood glucose homeostasis in the body.37,38 In PRLR related experimental mice, heterozygous PRLR (PRLR ±) female mice are fertile. The destruction of PRLR gene leads to the failure of β cells to expand properly during pregnancy, which leads to the decrease in blood insulin levels and maternal glucose tolerance and damage to glucose homeostasis in vivo. Many people reported that after PRLR inactivation, β - cell volume decreased, glucose tolerance decreased and insulin secretion decreased.39,40 At present, some scholars have refined to study PRLR pdx1-d/d, and have successfully developed a mouse model for gestational diabetes mellitus.41
Studies have shown that the expression of hepatocyte growth factor (HGF) and c-Met will also be up-regulated during pregnancy. HGF is an insulin promoter of beta cells. In the study of PancMet KO mice, it was found that the pancreatic condition and blood glucose homeostasis of nonpregnant PancMet Ko female mice were normal, but glucose intolerance occurred after pregnancy, and the replication and apoptosis of islet β cells decreased, followed by increased blood glucose and impaired glucose tolerance. This indicates that the HGF/c-Met signal plays an important role in pregnant women, and its deletion or weakening may cause gestational diabetes mellitus.42
5-hydroxytryptamine (5-HT) acts on the downstream of prolactin signal to stimulate the proliferation of beta cells. During pregnancy, the expression of 5-hydroxytryptamine synthetase tryptophan hydroxylase-1 (Tph1) and the production of 5-HT in β - cells increase dramatically. Blocking 5-HTR2b signal in pregnant mice makes 5-HT unable to play a role, resulting in the reduction of β - cell proliferation and glucose intolerance. This study revealed a signaling pathway linking beta cells to hormonal changes during pregnancy, demonstrating the role of 5-HTR2b related genes in GDM.7,43
A new candidate gene PAX-8 has recently been identified. PAX-8 was induced in the islets of pregnant mice. It should be noted that under basic conditions, overexpression of PAX-8 in mouse islets could reduce the apoptosis by 40% and reduce cytokine-induced apoptosis. A similar decrease in caspase-3 cleaved by the apoptotic marker in the transfected β cells was also confirmed by immunostaining. Overall, these data suggest that PAX-8 promotes the survival of islet cell sand establishes a link between PAX-8 and GDM research.44
Dogs
Partial Pancreatectomy (PPX)
PPX is the most direct way to induce high blood glucose levels. After surgery, insulin-producing cells and various other pancreatic cells are significantly reduced, which further impairs the body’s ability to control blood glucose homeostasis. In a series of experiments at the beginning of the twentieth century, Markowitz et al performed total pancreatectomy for dogs in late pregnancy which caused hyperglycemia and diabetes in a few days, while Carlson et al pointed out that the similar results did not appear after similar operations. Moreover, the sample size of these early experiment is often relatively small, usually including 1–3 animals, and due to the occurrence of the pregnancy death, and spontaneous abortion, the experimental results are not ideal, and the conclusions are not convincing. Although PPX can successfully lead to abnormal blood glucose level during pregnancy, this is not specific. In-depth studies of the GDM mechanism in recent years have shown that human GDM is a spontaneous event caused by a combination of genetic, environmental, dietary, and other factors, rather than a sudden pancreatic dysfunction.45 Therefore, the PPX model cannot conduct in-depth research into the causes of GDM, but it has a certain role to play in studying the outcome of GDM.
HFSD
Mary Courtney Moore et al compared changes in the blood glucose between the normal meat diet group and the HFSD group during the 4–5th week of pregnancy. All dogs with a HFSD showed impaired glucose tolerance (IGT) or gestational diabetes (GDM).46 In the second trimester of pregnancy, the dog’s pancreas did not have an islet hypertrophy, hyperplasia or regeneration, combined with lack of pancreatic adaptability. In this study, systemic insulin sensitivity was reduced by 20–30% in the HFSD group. Although the role of genetic factors, environmental factors and other factors in GDM is not considered, this model provides a useful large animal model for the study of pathophysiology and treatment of GDM.
Sheep
HFSD
From 60 days before pregnancy to 75 days after pregnancy, many ewes were fed with high-fat diet. On day 75, the concentration of blood glucose and ewe insulin in the experimental group was significantly increased.47 This model is similar to other animal models based on high-fat diets or glucose injection, which ignores the genetic and environmental factors and achieves different pregnancy outcomes and effects on offspring for dietary factors.
Chemical Induction
STZ was used on the 85th day and 90th day of gestation for 14 pregnant ewes. The intravenous glucose tolerance test was performed before to the first administration of the drug, before the second administration of the drug, and 4 weeks after the last administration of the drug, and it was observed that the fasting blood glucose of the pregnant ewe was significantly increased. Insulin response from the experimental group showed that one STZ lost the second stage insulin response to glucose load, while the first stage insulin resistance to glucose load was observed after two doses.48 In this experiment, the STZ-induced destruction of pancreatic β-cells led to a reduction in insulin and weight gain, resulting in changes in maternal glucose and insulin responses. The study provided a large animal model suited to gestational diabetes research.
Miodovnik et al took intravenous injection of 40 mg/kg.bw ALX in the maternal inferior vena. Before ALX injection and 1, 3, and 5 days after ALX administration, the plasma glucose, insulin and glucagon of pregnant ewes were measured, and a series of symptoms of GDM in pregnant sheep were found. However, two animals had miscarriage between day 1 and day 3 following the administration of ALX.49 In the case of an increase in the number of experimental animals, J P lips and others also tried to construct such models. The results were similar in that all non-abortion ewes had symptoms of hyperglycemia and insulin resistance, but the rate of abortion was also relatively high.50 Therefore, this model is more suitable for the acute study of GDM. If we want to conduct a long-term study, we need to give appropriate amount of insulin to prevent serious acidosis and death of the mother.
Pigs
Chemical Induction
On the 80th day of gestation, STZ was injected into pregnant pigs. In the high-dose group, serum glucose concentration increased to hyperglycemia level and the insulin concentration decreased.51 But again, this method is more suitable for the study of the consequences and treatment of GDM.
On the 75th day of gestation, the pregnant pigs were divided into two groups and injected with buffer or ALX and the blood glucose level of pregnant pigs injected with ALX increased by 300%, confirming the successful induction of GDM model.52 This model is similar to other chemically induced animal models, but the systems of small pigs are highly similar to those of humans, and thus pigs have many applications in the establishment of diabetes models, which provides a certain basis for the establishment of GDM animal models.
Summary
In conclusion, animal models of different types and conditions can be used for different research purposes, and the modeling standards are also different. There are many methods used for different factors affecting GDM, such as high-fat diet or glucose injection, which can be used in dietary factors research; methods such as gene knockout can be used during the research on genetic factors. It is worth mentioning that in the way of chemical drug induction, such as STZ, ALX and other induced models, drugs will lead to permanent destruction of beta cells, resulting in GDM symptoms that still exist after pregnancy has ended. It won’t be like human GDM progress, when the end of pregnancy, the high glucose level and insulin resistance phenomenon are relieved. The current solution is to transplant normal islets into experimental animals, but this method requires high experimental technology and cost.53,54 Even if it is successful in rodents, it may not be able to find new specific directions for research due to different physiological mechanisms. However, large animals are more similar to humans, which may be applicable in large animal experiments and further exploration is needed. In the drug experiment of STZ, the time of injection of STZ is still worth discussing. At present, STZ is usually used to construct the animal model of GDM after animal pregnancy in domestic and foreign experiments, and it is used more in rodent experiments. The time of STZ injection is mostly 0–6 days of gestation. Some literatures indicate that if STZ is injected prematurely, the disorder of glucose metabolism may directly lead to abortion or termination of pregnancy in rats. Therefore, STZ injection on the 6th day of gestation can relatively guarantee embryos survival rate. 11 Due to the complexity of GDM pathogenesis, in the future, the research direction will gradually shift to a combination of multiple factors. There is still a lack of experimental data for large animals, and HFSD and chemical induction methods are still the mainstream. However, because of the incidence of large animals is more similar to the actual situation of human GDM, in the follow-up study, there are still many problems to be solved, such as the islet function which cannot be recovered after pregnancy, and the mechanism of the model constructed by some method is too single. If conditions are allowed, large animals, including pigs, rabbits, sheep, non-human primates, etc., should be used for the establishment of GDM animal models in future research with a view tender standing of human GDM.
Statement of Ethics
This article does not contain any studies with human or animals performed by any of the authors.
Consent for Publication
Does not apply for this manuscript.
Acknowledgments
We gratefully acknowledge Yueyang Zhao for providing intellectual support and technical assistance.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was supported by the National Natural Science Foundation of China (81700706), the 345 Talent Project of Shengjing hospital and the Clinical research project of Liaoning diabetes medical nutrition prevention society (grant No. LNSTNBYXYYFZXH-RS01B).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Chiefari E, Arcidiacono B, Foti D, Brunetti A. Gestational diabetes mellitus: an updated overview. J Endocrinol Invest. 2017;40(9):899–909. doi:10.1007/s40618-016-0607-5
2. Xin L. Research progress of experimental gestational diabetes rat model. Prog Mod Obstet Gynecol. 2012;21(8):648–650.
3. Koopmans SJ, VanderMeulen J, Wijdenes J, Corbijn H, Dekker R. The existence of an insulin-stimulated glucose and non-essential but not essential amino acid substrate interaction in diabetic pigs. BMC Biochem. 2011;12:25. doi:10.1186/1471-2091-12-25
4. Yu W, Wu N, Li L, OuYang H, Qian M, Shen HA. Review of research progress on glycemic variability and gestational diabetes. Diabetes Metab Syndr Obes. 2020;13:2729–2741. doi:10.2147/DMSO.S261486
5. Yuan N, Zhai H, Du D, Shan X, Wang M. Advances in pathogenesis of gestational diabetes mellitus. J Guangxi Med Univ. 2019;36(2):321–324.
6. Song D, Ran L, Jiang R, Yingjie W. Progress of animal models in diabetes research. Chin J Comp Med. 2016;26(9):83–87.
7. Baeyens L, Hindi S, Sorenson RL, German MS. Beta-Cell adaptation in pregnancy. Diabetes Obes Metab. 2016;18(Suppl 1):63–70. doi:10.1111/dom.12716
8. Moyce BL, Dolinsky VW. Maternal beta-cell adaptations in pregnancy and placental signalling: implications for gestational diabetes. Int J Mol Sci. 2018;19(11):3467. doi:10.3390/ijms19113467
9. Lenzen S. The mechanisms of alloxan- and streptozotocin-induced diabetes. Diabetologia. 2008;51(2):216–226. doi:10.1007/s00125-007-0886-7
10. RAKIETEN N, RAKIETEN ML, NADKARNI MR. Studies on the diabetogenic action of streptozotocin (NSC-37917). Cancer Chemother Rep.1963;29:91-98
11. Jiang J, Chunwei W, Xing L, et al. Establishment of gestational diabetes rat model with different doses of streptozotocin. J Guiyang Med Coll. 2010;35(3):221–223.
12. Cui Y, Yang D, Zhang Q, et al. Comparative study on STZ induced type 2 and type 1 diabetic rat models. J Med Res. 2018;47(5):36–38.
13. Hao Q, Song C. Establishment of gestational diabetes mellitus mouse model by two methods. World Latest Med Inf Abstr. 2019;19(6):185–186.
14. Jia L, Wei B, Ta N, Chen B. Comparison of the stability of different doses of streptozotocin in the establishment of gestational diabetes mellitus rat model. J Fourth Mi Med Univ. 2009;30(22):2500–2502.
15. Zhuang J, Ying H, Wang D. Study on the establishment of gestational diabetes mellitus rat model with low-dose streptozotocin. Prog Mod Obstet Gynecol. 2014;8:607–610.
16. Drews G, Kramer C, Dufer M, Krippeit-Drews P. Contrasting effects of alloxan on islets and single mouse pancreatic beta-cells. Biochem J. 2000;352(Pt 2):389–397. doi:10.1042/bj3520389
17. Gai W, Schott-Ohly P, Schulte IWS, Gleichmann H. Differential target molecules for toxicity induced by streptozotocin and alloxan in pancreatic islets of mice in vitro. Exp Clin Endocrinol Diabetes. 2004;112(1):29–37. doi:10.1055/s-2004-815724
18. Liang J, Yang S, Wen M, Wei S. Establishment of diabetic pregnancy rat model by alloxan. Chin Maternal Child Health Res. 2006;17(6):454–456.
19. Shui Y, Xia X, Zhang Q, Zhang C, Yuanzhe W. Changes of dipeptidyl peptidase 4 activity in alloxan induced diabetic pregnant rats. Southeast Natl Defense Med. 2015;6:568–571.
20. Linxia L, Wang Y, Yan Q, et al. Establishment of gestational hyperglycemia animal model by continuous intravenous infusion of hypertonic glucose. Shanghai Med. 2006;29(5):306–308.
21. Zhang J. Establishment of mouse model of gestational diabetes mellitus induced by high fat and high sugar diet and Study on lipid metabolism characteristics: internal medicine. Ningxia Medical University; 2013. Available from: http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=D421746.
22. Huang B, Huang C, Zhao H, et al. Impact of GPR1 signaling on maternal high-fat feeding and placenta metabolism in mice. Am J Physiol Endocrinol Metab. 2019;316(6):E987–E997. doi:10.1152/ajpendo.00437.2018
23. Vuong B, Odero G, Rozbacher S, et al. Exposure to gestational diabetes mellitus induces neuroinflammation, derangement of hippocampal neurons, and cognitive changes in rat offspring. J Neuroinflammation. 2017;14(1):80. doi:10.1186/s12974-017-0859-9
24. Ren J, Ding Y, Yin L, et al. Effects of high-sucrose/high-fat diet feeding time on establishment of a mouse model of gestational diabetes mellitus. J Hyg Res. 2018;47(1):128–133.
25. Retnakaran R, Hanley AJ, Raif N, Connelly PW, Sermer M, Zinman B. C-reactive protein and gestational diabetes: the central role of maternal obesity. J Clin Endocrinol Metab. 2003;88(8):3507–3512. doi:10.1210/jc.2003-030186
26. Wolf M, Sandler L, Hsu K, Vossen-Smirnakis K, Ecker JL, Thadhani R. First-trimester C-reactive protein and subsequent gestational diabetes. Diabetes Care. 2003;26(3):819–824. doi:10.2337/diacare.26.3.819
27. Qiu C, Williams MA, Vadachkoria S, Frederick IO, Luthy DA. Increased maternal plasma leptin in early pregnancy and risk of gestational diabetes mellitus. Obstet Gynecol. 2004;103(3):519–525. doi:10.1097/01.AOG.0000113621.53602.7a
28. Kautzky-Willer A, Pacini G, Tura A, et al. Increased plasma leptin in gestational diabetes. Diabetologia. 2001;44(2):164–172. doi:10.1007/s001250051595
29. Zhou G, Cong L. Research progress on the relationship between adiponectin and pregnancy. Foreign Med. 2007;34(3):159–161.
30. Chen L, Huang X, Yang Y, et al. Establishment of gestational diabetes mellitus mouse model induced by high fat diet combined with STZ. Chin J Comp Med. 2016;26(3):15–18.
31. Tang M, Wenting X, Wang C, Wang L. Optimization of high fat diet combined with streptozotocin in induced gestational diabetes rat model. J Jiangsu Univ. 2018;28(5):393–396.
32. Yang R, Qingwang L, Zhao R. Comparison of the effects of alloxan and streptozotocin on diabetic mice. J Northwest Agric Forest Univ. 2006;34(2):17–20.
33. Abdul AS, John CM, Mohamed YN, et al. Animal model of gestational diabetes mellitus with pathophysiological resemblance to the human condition induced by multiple factors (Nutritional, pharmacological, and stress) in rats. Biomed Res Int. 2016;2016:9704607.
34. Kaufmann RC, Amankwah KS, Dunaway G, Maroun L, Arbuthnot J, Roddick JJ. An animal model of gestational diabetes. Am J Obstet Gynecol. 1981;141(5):479–482. doi:10.1016/S0002-9378(15)33263-4
35. Yamashita H, Shao J, Qiao L, Pagliassotti M, Friedman JE. Effect of spontaneous gestational diabetes on fetal and postnatal hepatic insulin resistance in Lepr(db/+) mice. Pediatr Res. 2003;53(3):411–418. doi:10.1203/01.PDR.0000049667.58071.7D
36. Yamashita H, Shao J, Ishizuka T, et al. Leptin administration prevents spontaneous gestational diabetes in heterozygous Lepr(db/+) mice: effects on placental leptin and fetal growth. Endocrinology. 2001;142(7):2888–2897. doi:10.1210/endo.142.7.8227
37. Holemans K, Caluwaerts S, Poston L, Van Assche FA. Diet-induced obesity in the rat: a model for gestational diabetes mellitus. Am J Obstet Gynecol. 2004;190(3):858–865. doi:10.1016/j.ajog.2003.09.025
38. Fujinaka Y, Takane K, Yamashita H, Vasavada RC. Lactogens promote beta cell survival through JAK2/STAT5 activation and Bcl-XL upregulation. J Biol Chem. 2007;282(42):30707–30717. doi:10.1074/jbc.M702607200
39. Banerjee RR, Cyphert HA, Walker EM, et al. Gestational diabetes mellitus from inactivation of prolactin receptor and MafB in Islet beta-Cells. Diabetes. 2016;65(8):2331–2341. doi:10.2337/db15-1527
40. Arumugam R, Fleenor D, Freemark M. Knockdown of prolactin receptors in a pancreatic beta cell line: effects on DNA synthesis, apoptosis, and gene expression. Endocrine. 2014;46(3):568–576. doi:10.1007/s12020-013-0073-1
41. Nteeba J, Kubota K, Wang W, et al. Pancreatic prolactin receptor signaling regulates maternal glucose homeostasis. J Endocrinol. 2019;241(1):71–83. doi:10.1530/JOE-18-0518
42. Demirci C, Ernst S, Alvarez-Perez JC, et al. Loss of HGF/c-Met signaling in pancreatic beta-cells leads to incomplete maternal beta-cell adaptation and gestational diabetes mellitus. Diabetes. 2012;61(5):1143–1152.
43. Kim H, Toyofuku Y, Lynn FC, et al. Serotonin regulates pancreatic beta cell mass during pregnancy. Nat Med. 2010;16(7):804–808. doi:10.1038/nm.2173
44. Martin-Montalvo A, Lopez-Noriega L, Jimenez-Moreno C, et al. Transient PAX8 expression in Islets during pregnancy correlates with beta-cell survival, revealing a novel candidate gene in gestational diabetes mellitus. Diabetes. 2019;68(1):109–118. doi:10.2337/db18-0285
45. Pasek RC, Gannon M. Advancements and challenges in generating accurate animal models of gestational diabetes mellitus. Am J Physiol Endocrinol Metab. 2013;305(11):E1327–E1338. doi:10.1152/ajpendo.00425.2013
46. Moore MC, Menon R, Coate KC, et al. Diet-induced impaired glucose tolerance and gestational diabetes in the dog. J Appl Physiol. 2011;110(2):458–467. doi:10.1152/japplphysiol.00768.2010
47. Ford SP, Zhang L, Zhu M, et al. Maternal obesity accelerates fetal pancreatic β-cell but not α-cell development in sheep: prenatal consequences. Am J Physiol Regul Integr Comp Physiol. 2009;297(3):R835–R843. doi:10.1152/ajpregu.00072.2009
48. Dickinson JE, Meyer BA, Chmielowiec S, Palmer SM. Streptozocin-induced diabetes mellitus in the pregnant ewe. Am J Obstet Gynecol. 1991;165(6 Pt 1):1673–1677. doi:10.1016/0002-9378(91)90013-H
49. Miodovnik M, Mimouni F, Berk M, Clark KE. Alloxan-induced diabetes mellitus in the pregnant ewe: metabolic and cardiovascular effects on the mother and her fetus. Am J Obstet Gynecol. 1989;160(5 Pt 1):1239–1244. doi:10.1016/0002-9378(89)90203-2
50. Lips JP, Jongsma HW, Eskes TK. Alloxan-induced diabetes mellitus in pregnant sheep and chronic fetal catheterization. Lab Anim. 1988;22(1):16–22. doi:10.1258/002367788780746638
51. Ramsay TG, Wolverton CK, Steele NC. Alteration in IGF-I mRNA content of fetal swine tissues in response to maternal diabetes. Am J Physiol. 1994;267(5 Pt 2):R1391–R1396.
52. Ezekwe MO, Ezekwe EI, Sen DK, Ogolla F. Effects of maternal streptozotocin-diabetes on fetal growth, energy reserves and body composition of newborn pigs. J Anim Sci. 1984;59(4):974–980. doi:10.2527/jas1984.594974x
53. Ryan EA, Tobin BW, Tang J, Finegood DT. A new model for the study of mild diabetes during pregnancy. Syngeneic islet-transplanted STZ-induced diabetic rats. Diabetes. 1993;42(2):316–323. doi:10.2337/diab.42.2.316
54. Damasceno DC, Netto AO, Iessi IL, et al. Streptozotocin-induced diabetes models: pathophysiological mechanisms and fetal outcomes. Biomed Res Int. 2014;2014:819065. doi:10.1155/2014/819065
55. American Diabetes Association. 2. Classification and diagnosis of diabetes: standards of medical care in diabetes–2020. Diabetes Care. 2020;43(Suppl1):S14–s31. doi:10.2337/dc20-S002
56. ACOG Practice Bulletin No. 190: Gestational Diabetes Mellitus. Obstet Gynecol. 2018;131(2):e49–e64.
57. Hod M, Kapur A, Sacks DA, et al. The International Federation of Gynecology and Obstetrics (FIGO) Initiative on gestational diabetes mellitus: A pragmatic guide for diagnosis, management, and care. Int J Gynaecol Obstet. 2015;131 Suppl 3:S173–S211.
58. Metzger BE, Gabbe SG, Persson B, et al. International association of diabetes and pregnancy study groups recommendations on the diagnosis and classification of hyperglycemia in pregnancy. Diabetes Care. 2010;33(3):676–682.
59. Diagnostic criteria and classification of hyperglycaemia first detected in pregnancy: a World Health Organization Guideline. Diabetes Res Clin Pract. 2014;103(3):341–363.
60. Society. CD. Guidelines for the prevention and control of type 2 diabetes in China (2017 Edition). Chin J Pract Intern Med. 2018;38 (4):292–344.
61. endocrinology Cso. Experts consensus on management of glycemic variability of diabetes mellitus. Drug Evaluation. 2017;014(17):5–8,14.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.